
Extent and Severity of Caliciopsis Canker in New England, USA: An Emerging Disease of Eastern White Pine (Pinus strobus L.)

 ,
,
Abstract
:1. Introduction

2. Experimental Section
2.1. Study Area
2.2. Site Selection
2.3. Sampling
2.4. Isolation and Identification of Caliciopsis pinea
2.5. GIS Plot Data
2.6. Statistical Analyses
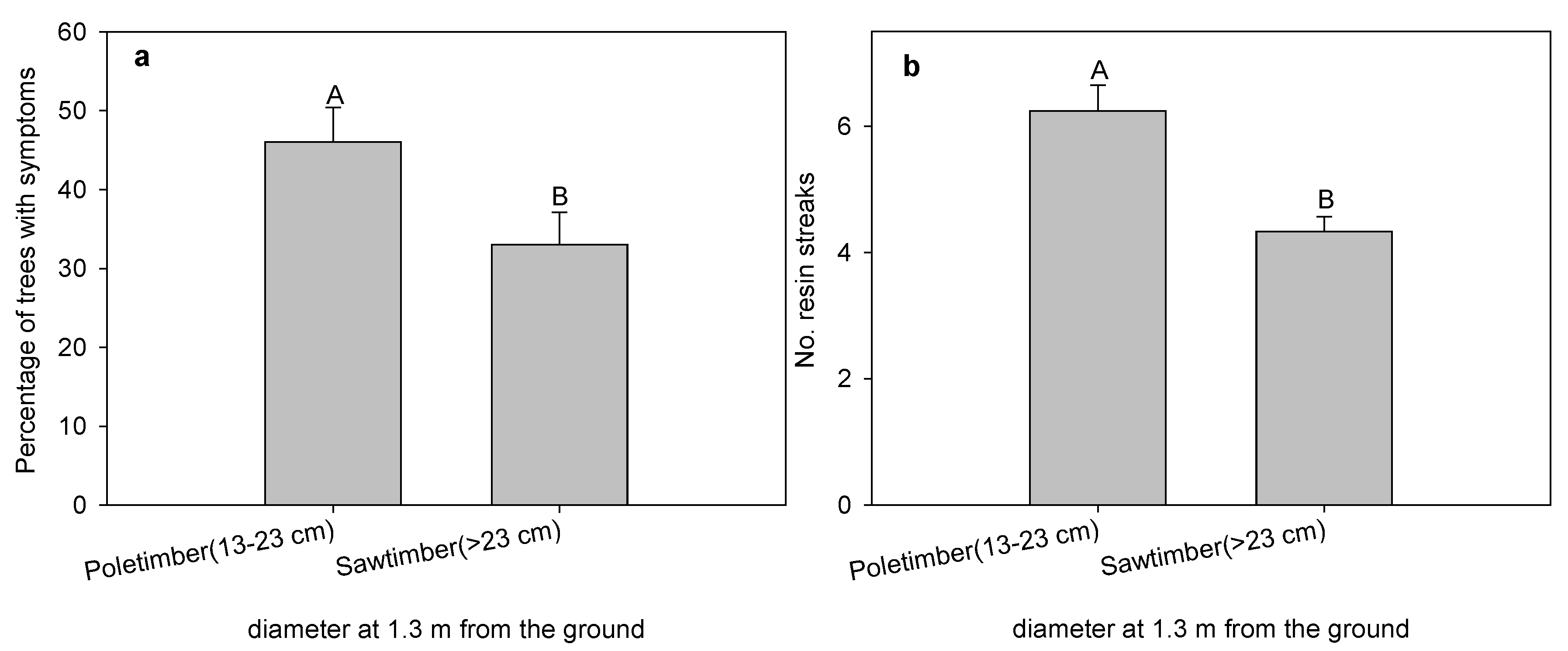
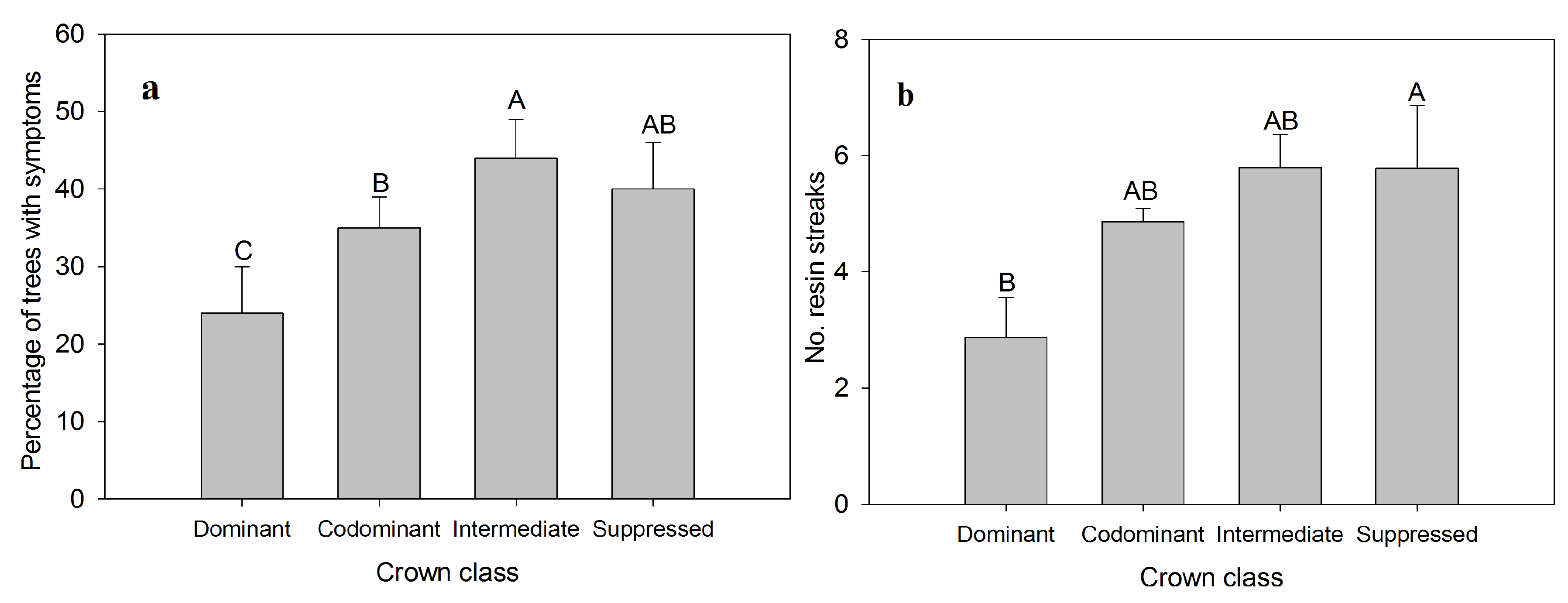
3. Results and Discussion
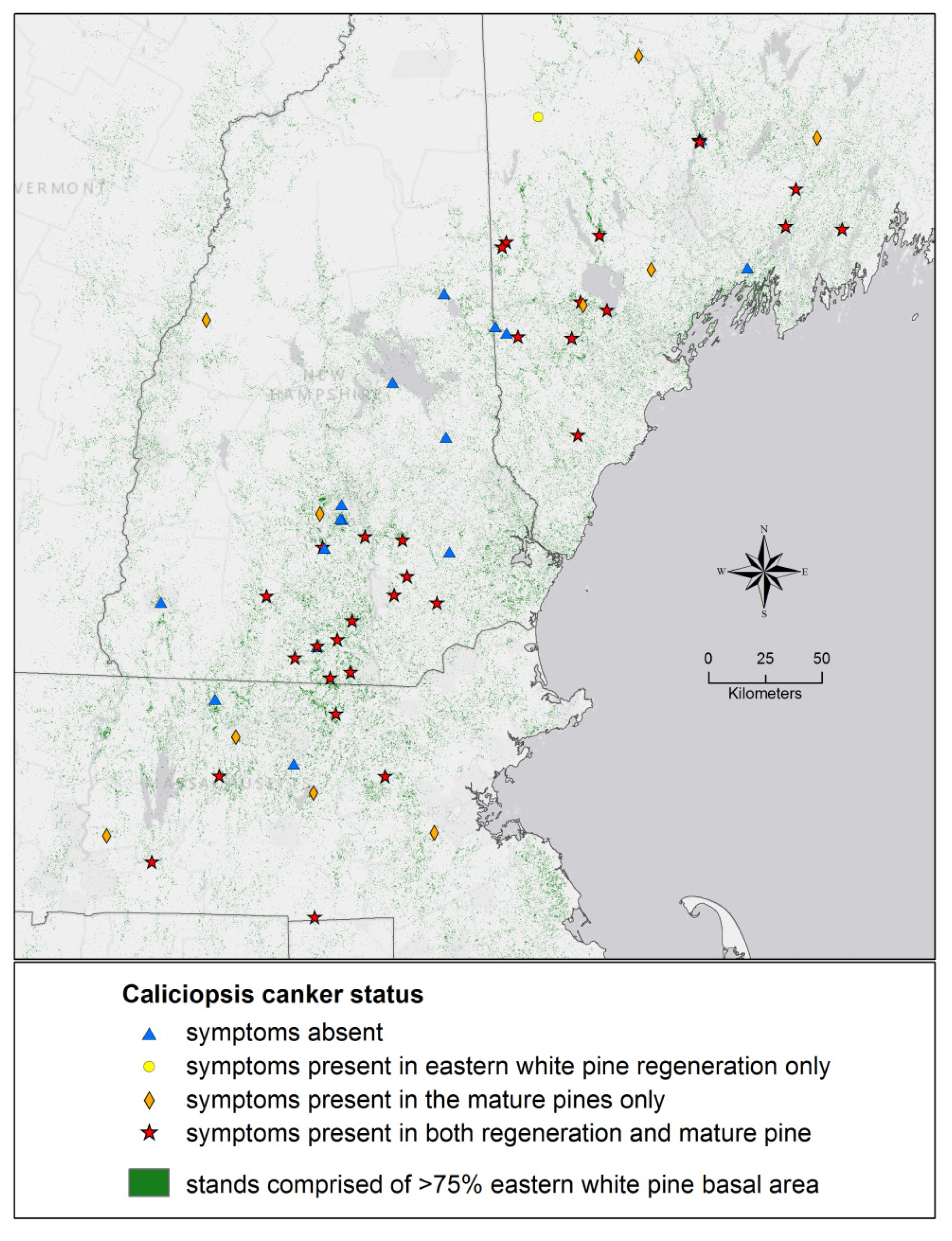


{kind=link}
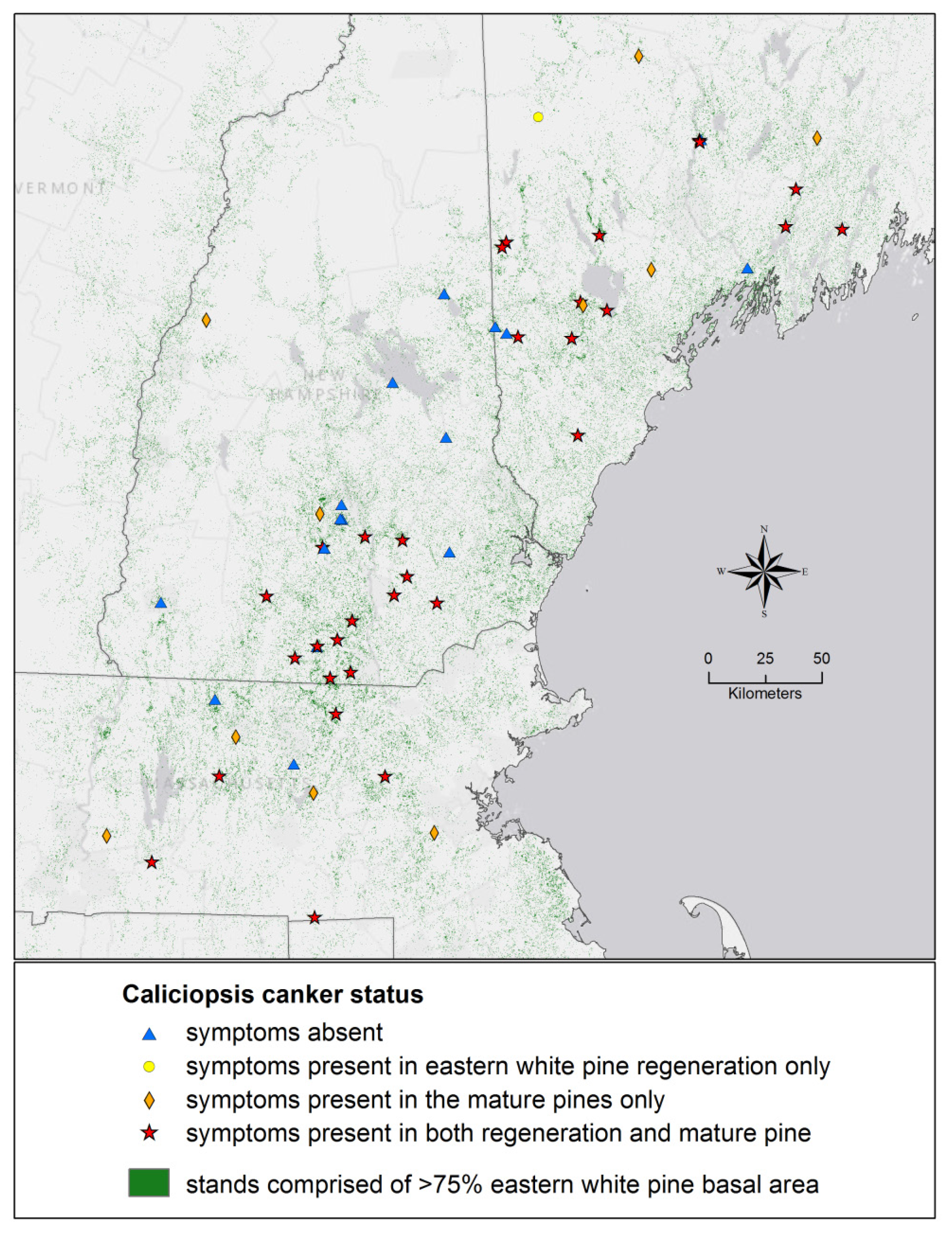{kind=link}
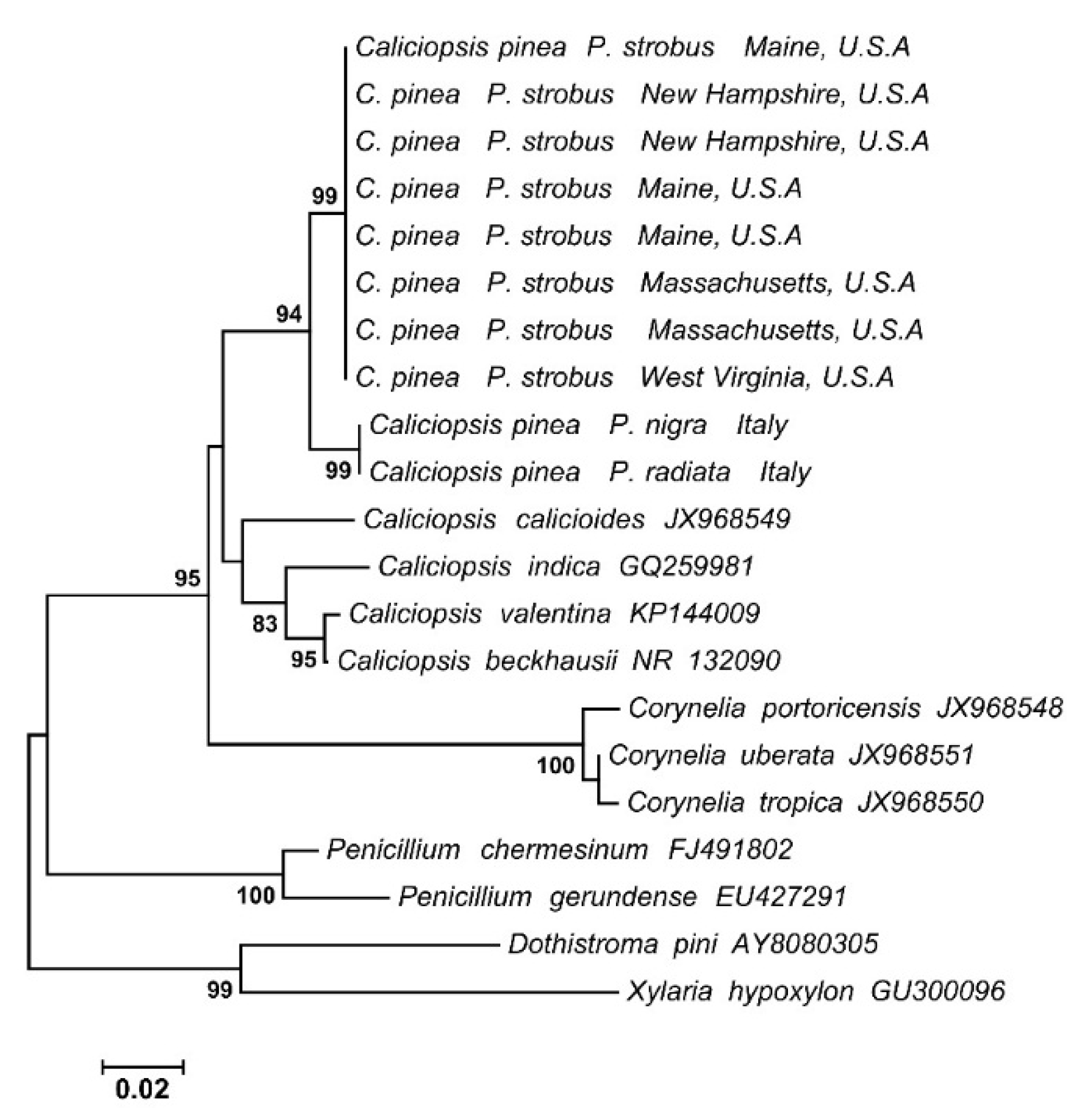{kind=link}
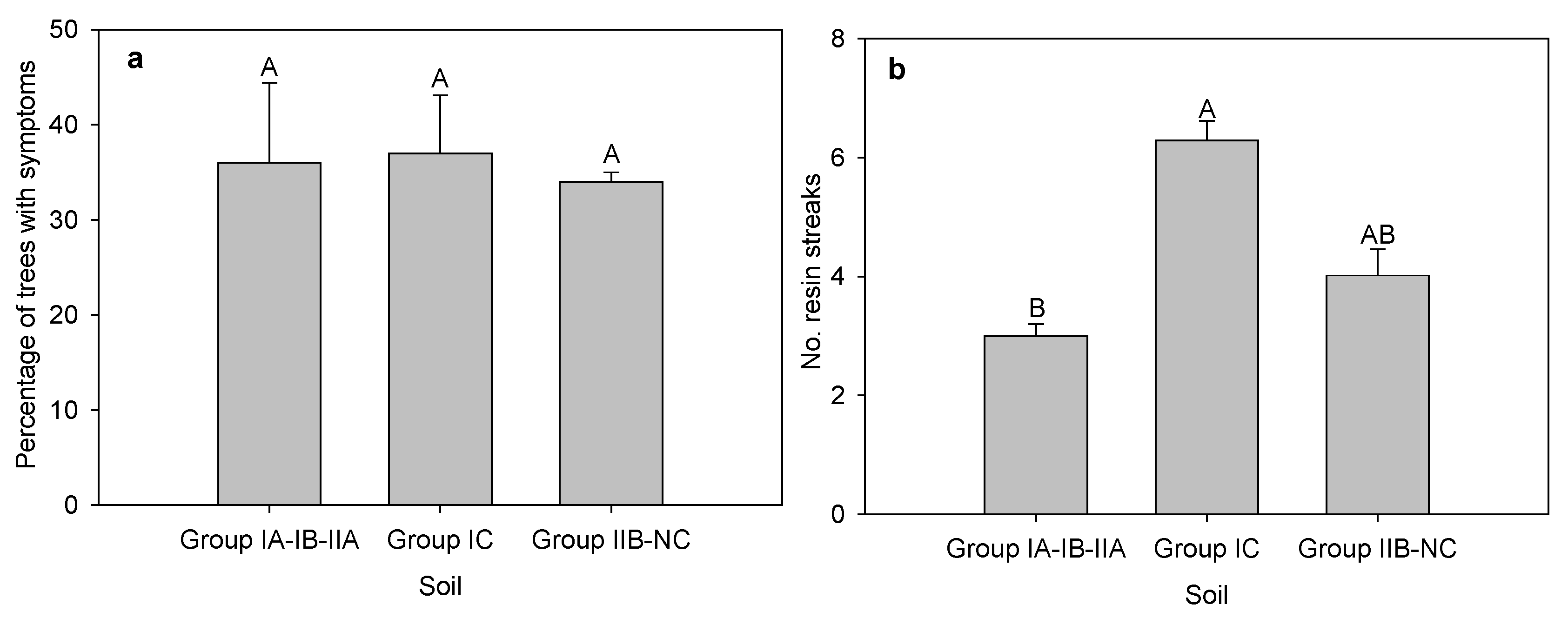{kind=link}
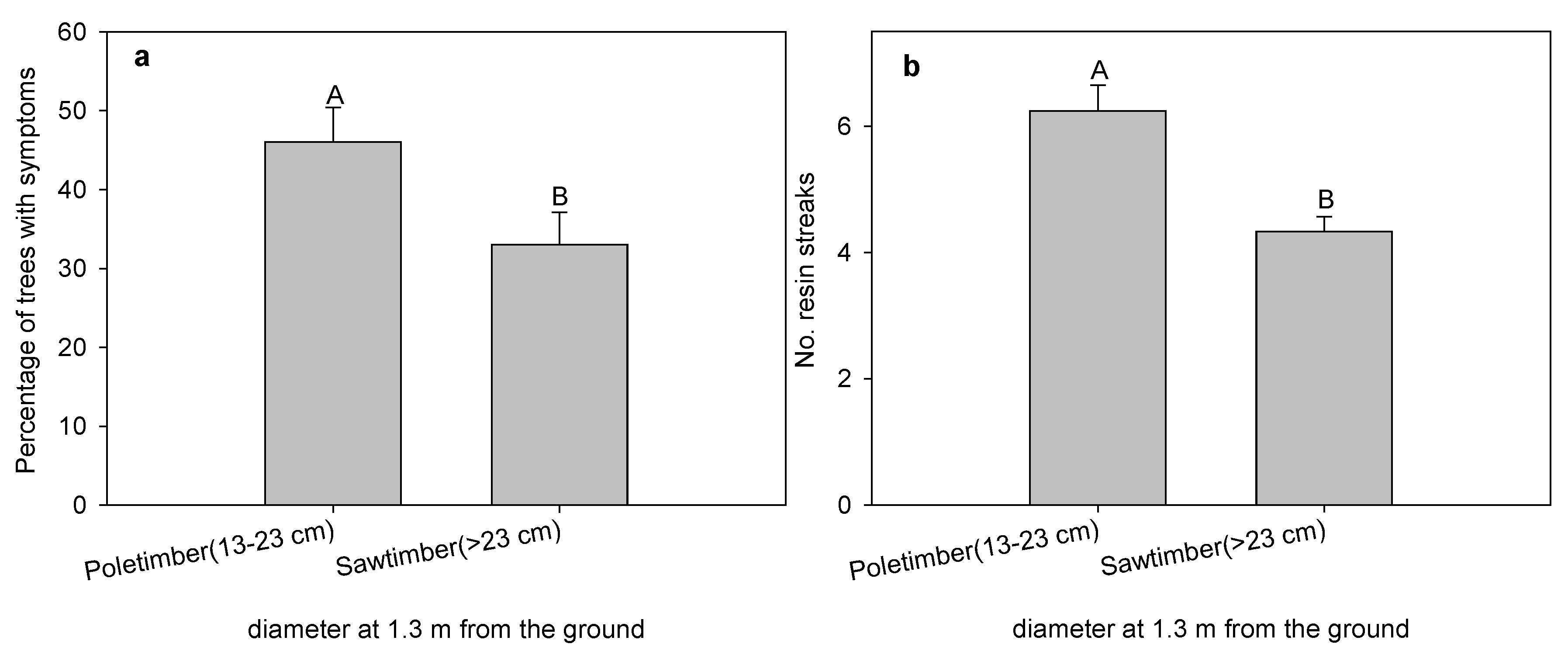{kind=link}
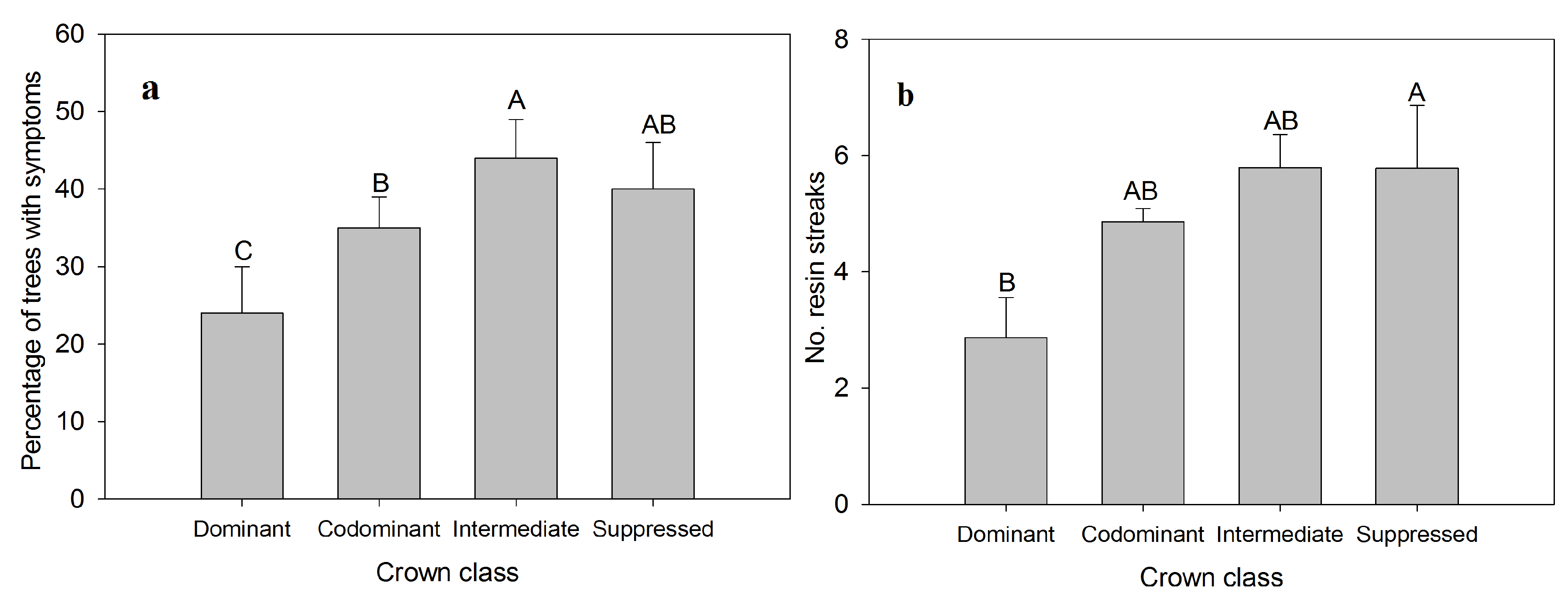{kind=link}
| Soil Group | Description | Percentage Sites with Caliciopsis Canker Symptoms | No. Sites |
|---|---|---|---|
| IA-IB-IIA | well drained, loamy, favourable moisture conditions, fertility suitable for hardwoods | 59 | 27 |
| IC | excessively drained, very dry, coarse textured soils derived from glacial outwash, fertility adequate for softwoods but not hardwoods | 86 | 21 |
| IIB-NC | sites with poorly drained soils and low fertility | 78 | 9 |

4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References and Notes
- Farr, D.F.; Rossman, A.Y. Fungal Databases, Systematic Mycology and Microbiology Laboratory, ARS, USDA. Available online: http://nt.ars-grin.gov/fungaldatabases/ (accessed on 17 September 2015).
- Luchi, N.; Capretti, P.; Migliorini, D.; Santini, A. Caliciopsis Canker: A Damaging Disease on Plantations in Italy. Abstracts of the Joint IUFRO 7.02.02 “Foliage, shoot and stem diseases of forest trees” and 7.03.04 “Diseases and insects in forest nurseries”. In Proceedings of Working Parties Meeting, Uppsala, Sweden, 7–12 June 2015; Swedish University of Agricultural Sciences: Uppsala, Sweden, 2015; p. 21. [Google Scholar]
- Funk, A. Studies in the genus Caliciopsis. Can. J. Bot. 1963, 41, 530–543. [Google Scholar] [CrossRef]
- Ray, W.W. Pathogenicity and cultural experiments with Caliciopsis pinea. Mycologia 1936, 28, 201–208. [Google Scholar] [CrossRef]
- Delatour, C. Caliciopsis pinea: Further results of artificial inoculations. Ann. Sci. for., Paris. 1969, 26, 285–295. [Google Scholar] [CrossRef]
- Lombard, K. Caliciopsis canker (pine canker) Caliciopsis pinea. In UNH Cooperative Extension Publication; UNH: Durham, NH, USA, 2003. [Google Scholar]
- Asaro, C. What is killing white pine in the highlands of West Virginia? In Forest Health Review May 2011; Virginia Department of Forestry: Charlottesville, VA, USA, 2011. [Google Scholar]
- Murphy, B. New Hampshire forest officials to study white pine canker. Wall Street Journal 2012. [Google Scholar]
- USDA Forest Service. 2000 Forest Health Highlights New Hampshire. Available online: http://www.fs.fed.us/foresthealth/fhm/fhh/fhh-00/nh/nh%20_00.pdf (accessed on 16 November 2015).
- Widmann, R.; McWilliams, W. An Overview of the White Pine Resource in New England Using Forest Inventory and Analyses Data. In Managing White Pine in a New Millennium; Bennett, K., Desmarais, K., Eds.; University of New Hampshire Cooperative Extention: Hillsborough, NH, USA, 2003; pp. 1–8. [Google Scholar]
- Yamasaki, M. White Pine as Wildlife Habitat. In Managing White Pine in a New Millennium; Benett, K., Desmarais, K., Eds.; University of New Hampshire Cooperative Extention: Hillsborough, NH, USA, 2003; pp. 33–36. [Google Scholar]
- McCormack, H.W. The morphology and development of Caliciopsis pinea. Mycologia 1936, 28, 188–196. [Google Scholar] [CrossRef]
- Sperduto, D.D.; Nichols, W.F. Natural Communities of New Hampshire; New Hampshire Natural Heritage Bureau: Concord, NH, USA, 2004. [Google Scholar]
- Environmetal Protection Agency. Level III and IV Ecoregions of the Continental United States. Available online: http://www.epa.gov/wed/pages/ecoregions/level_iii_iv.htm (accessed on 16 September 2015).
- Forest Inventory and Analysis National Program-USDA Forest Service. Available online: http://www.fia.fs.fed.us/ (accessed on 3 September 2015).
- Krist, F.J.; Ellenwood, J.R.; Woods, M.E.; McMahan, A.J.; Cowardin, J.P.; Ryerson, D.E.; Sapio, F.; Zweifler, M.O.; Romero, S.A. 2013–2014 National Insect and Disease Forest Risk Assessment. Available online: http://www.fs.fed.us/foresthealth/technology/nidrm.shtml#NIDRMReport (accessed on 16 June 2015).
- Kohler, A.; Murat, C.; Costa, M. High Quality Genomic DNA Extraction Using CTAB and Qiagen genomic-tip. Version 2; INRA Nancy Equipe Ecogénomique UMR Iam. Available online: http://1000.fungalgenomes.org/home/wp-content/uploads/2013/02/genomicDNAProtocol-K0511.pdf (accessed on 16 November 2015).
- Peck, C. Report of the Botanis. Ann. Rep. N.Y. State Mus. Nat. Hist. 1880, 33, 32. [Google Scholar]
- Broders, K.D.; Munck, I.; Wyka, S.; Iriarte, G.; Beaudoin, E. Characterization of fungal pathogens associated with White Pine Needle Damage (WPND) in Northeastern North America. Forests 2015, 6, 4088–4104. [Google Scholar] [CrossRef]
- Griffith, G.E.; Omernik, J.M.; Bryce, S.A.; Royte, J.; Hoar, W.D.; Homer, J.W.; Keirstead, D.; Metzler, K.J.; Hellyer, G. Ecoregions of New England. Available online: http://www.epa.gov/wed/pages/ecoregions/new_eng_eco.htm (accessed on 16 January 2015).
- USDA Agriultural Research Service. USDA Plant Hardiness Zone Map. Available online: http://planthardiness.ars.usda.gov (accessed on 16 January 2015).
- USDA Natural Resources Conservation Service. Soil Data Access. Available online: http://sdmdataaccess.nrcs.usda.gov/ (accessed on 16 January 2015).
- US Geological Survey. Elevation. Available online: http://nationalmap.gov/elevation.html (accessed on 16 January 2015).
- University of New Hampshire Cooperative Extension. Important Forest Soil Groups. Available online: https://extension.unh.edu/goodforestry/html/app-soils.htm (accessed on 16 January 2014).
- O’Hara, R.B.; Kotze, D.J. Do not log-transform count data. Methods Ecol. Evol. 2010, 1, 118–122. [Google Scholar] [CrossRef]
- Fries, M.L. Relationships Between Rooting Restrictions, Radial Growth, and Drought Stress with White Pine (Pinus strobus) Decline in Southern Maine; The University of Maine: Orono, ME, USA, 2002. [Google Scholar]
- Leak, W.B.; Cullen, J.B.; Frieswyk, T.S. Dynamics of White Pine in New England; Research Paper NE-699; USDA Forest Service, Northeastern Area State and Private Forestry: Radnor, PA, USA, 1995. [Google Scholar]
- Ostry, M.E.; Laflamme, G.; Katovich, S.A. Silvicultural approaches for management of eastern white pine to minimize impacts of damaging agents. For. Pathol. 2010, 40, 332–346. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Munck, I.A.; Livingston, W.; Lombard, K.; Luther, T.; Ostrofsky, W.D.; Weimer, J.; Wyka, S.; Broders, K. Extent and Severity of Caliciopsis Canker in New England, USA: An Emerging Disease of Eastern White Pine (Pinus strobus L.). Forests 2015, 6, 4360-4373. https://doi.org/10.3390/f6114360
Munck IA, Livingston W, Lombard K, Luther T, Ostrofsky WD, Weimer J, Wyka S, Broders K. Extent and Severity of Caliciopsis Canker in New England, USA: An Emerging Disease of Eastern White Pine (Pinus strobus L.). Forests. 2015; 6(11):4360-4373. https://doi.org/10.3390/f6114360
Chicago/Turabian StyleMunck, Isabel A., William Livingston, Kyle Lombard, Thomas Luther, William D. Ostrofsky, Jennifer Weimer, Stephen Wyka, and Kirk Broders. 2015. "Extent and Severity of Caliciopsis Canker in New England, USA: An Emerging Disease of Eastern White Pine (Pinus strobus L.)" Forests 6, no. 11: 4360-4373. https://doi.org/10.3390/f6114360
APA StyleMunck, I. A., Livingston, W., Lombard, K., Luther, T., Ostrofsky, W. D., Weimer, J., Wyka, S., & Broders, K. (2015). Extent and Severity of Caliciopsis Canker in New England, USA: An Emerging Disease of Eastern White Pine (Pinus strobus L.). Forests, 6(11), 4360-4373. https://doi.org/10.3390/f6114360