High NDVI and Potential Canopy Photosynthesis of South American Subtropical Forests despite Seasonal Changes in Leaf Area Index and Air Temperature

Abstract
:1. Introduction
2. Methods
2.1. Study Area
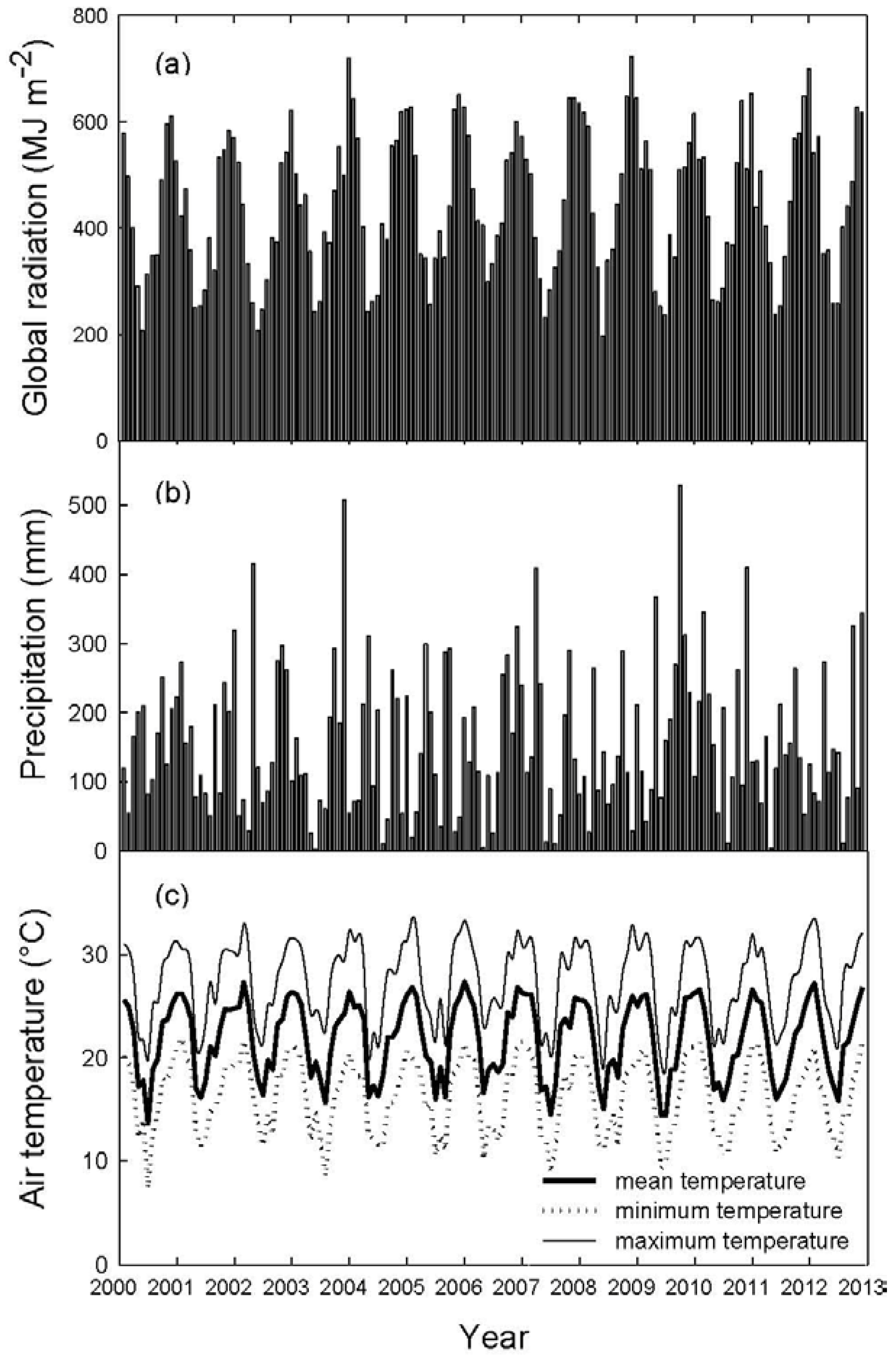
2.2. Field Measurements

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Family | Species | Origin | Phenology |
|---|---|---|---|
| Rutaceae | Balfourodendron riedelianum (Engl.) Engl. | Native | Brevideciduous |
| Meliaceae | Cabralea canjerana (Vell.) Mart. | Native | Evergreen |
| Meliaceae | Cedrela fissilis Vell. | Native | Deciduous |
| Bombacaceae | Ceiba speciosa (A.St.-Hil., A.Juss. & Cambess.) Ravenna | Native | Deciduous |
| Boraginaceae | Cordia trichotoma (Vell.) Arráb. ex Steud. | Native | Deciduous |
| Fabaceae | Holocalyx balansae Micheli | Native | Evergreen |
| Lauraceae | Ocotea diospyrifolia (Meisn.) Mez. | Native | Evergreen |
| Fabaceae | Lonchocarpus muehlbergianus Hassl. | Native | Brevideciduous |
| Fabaceae | Parapiptadenia rígida (Benth.)Brenan | Native | Brevideciduous |
| Sapotaceae | Chrysophyllum gonocarpum (Mart. & Eichler) Engl. | Native | Evergreen |
| Araucariaceae | Araucaria angustifolia (Bertol.) Kuntze | Native | Evergreen |
| Myrtaceae | Eucalyptus grandis W.Hill ex Maiden | Exotic | Evergreen |
| Pinaceae | Pinus elliotti elliotti × P. caribaea hondurensis (hybrid Pinus) | Exotic | Evergreen |
| Pinaceae | Pinus taeda L. | Exotic | Evergreen |
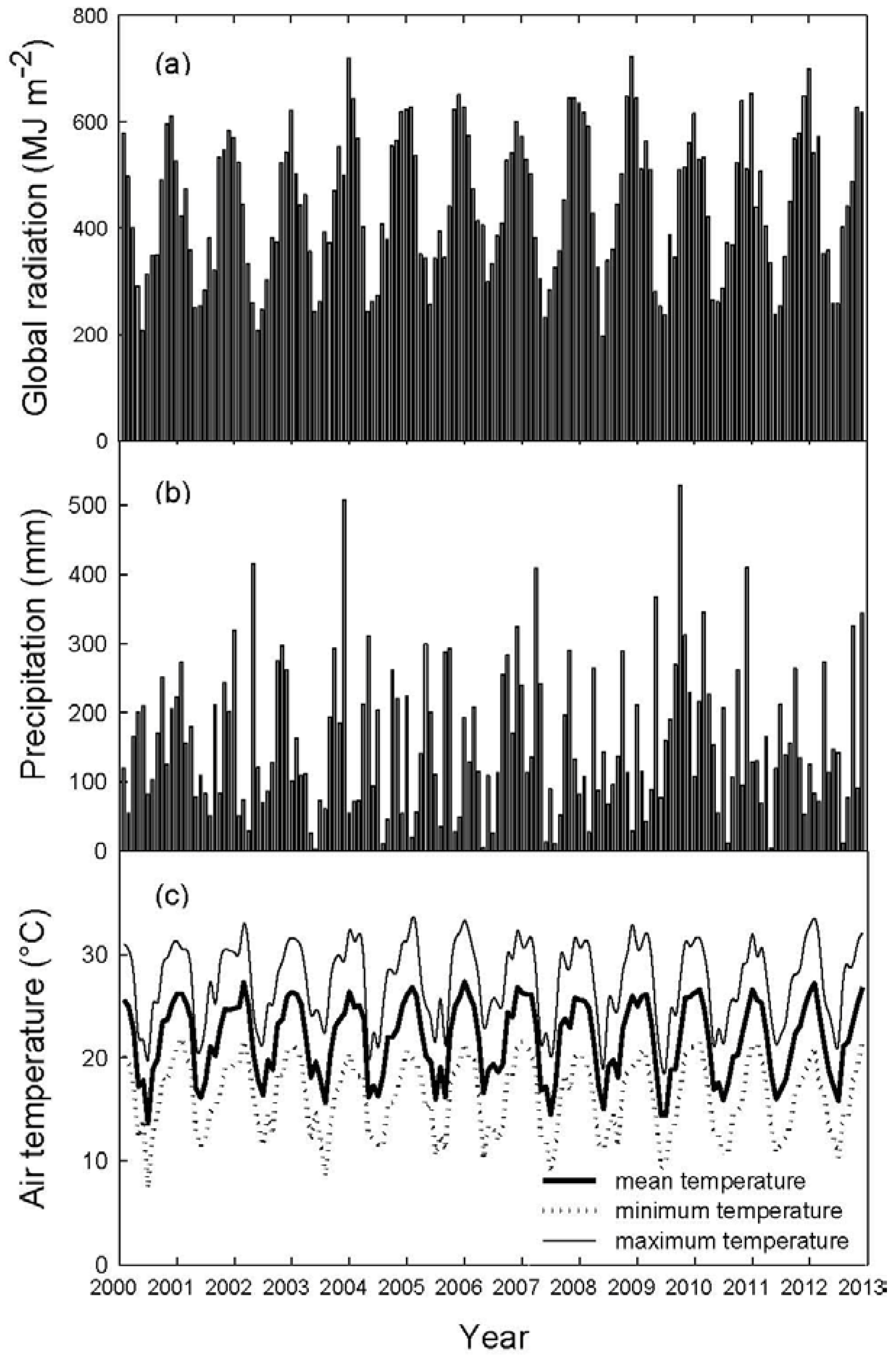
2.3. Remote Sensing and Meteorological Variables
| Study sites | Latitude (S) | Longitude (W) | NDVIN° pixels | LAIN° pixels |
|---|---|---|---|---|
| Native forest | 25°41′28.51′′ | 54°29′9.59′′ | 26 | 1 |
| 25°44′30.78′′ | 54°21′39.17′′ | 56 | 3 | |
| 25°48′16.13′′ | 54°15′6.77′′ | 14 | 2 | |
| P. taeda | 26°9′50.20′′ | 54°26′50.08′′ | 42 | 2 |
| 25°47′54.14′′ | 54°25′11.57′′ | 63 | 2 | |
| E. grandis | 26°6′34.80′′ | 54°26′10.68′′ | 10 | 2 |
| 26°7′43.13′′ | 54°34′33.16′′ | 9 | 1 | |
| A. angustifolia | 26°14′48.92′′ | 54°30′38.73′′ | 12 | 1 |
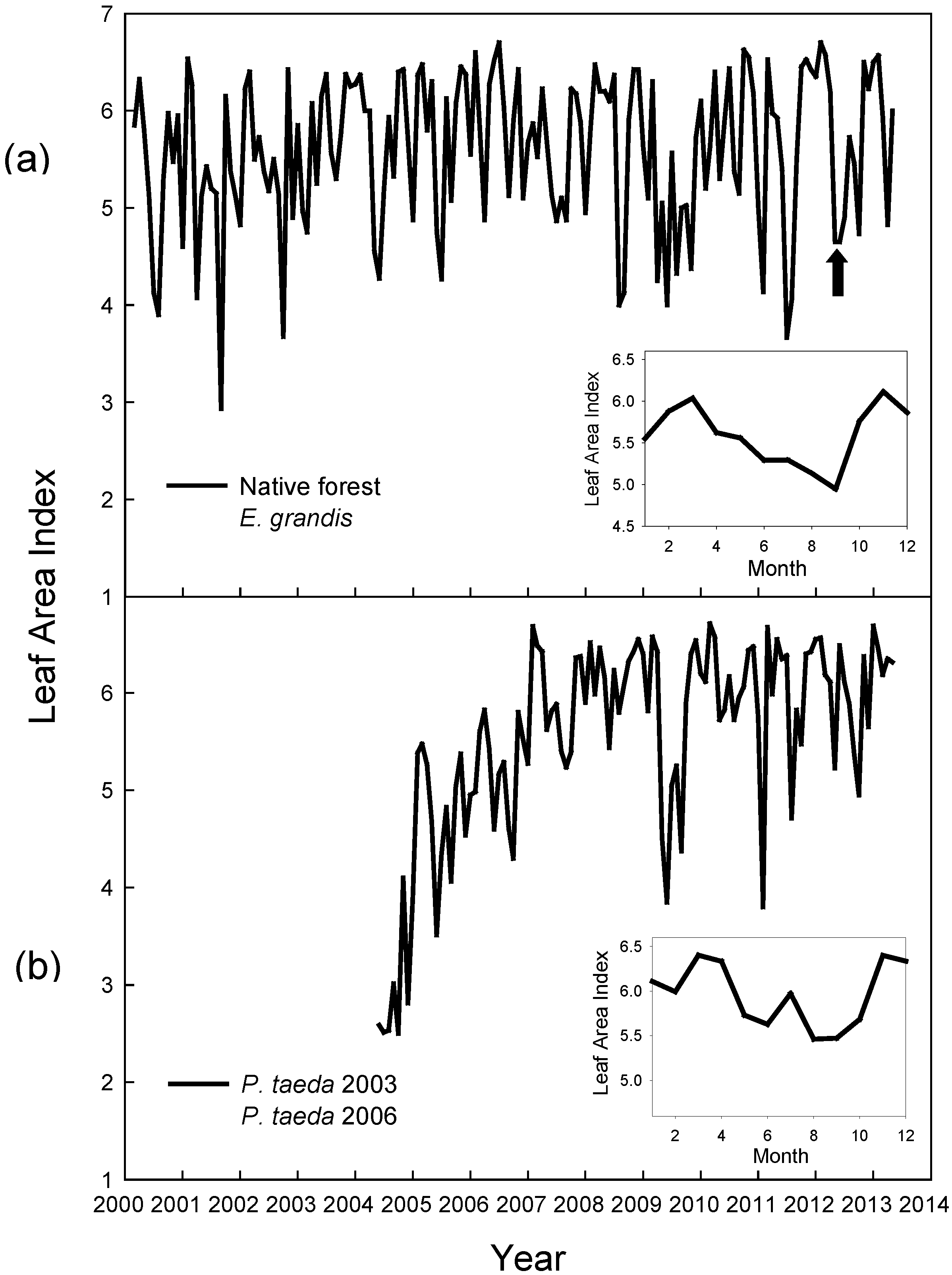
2.4. Data Analysis
3. Results and Discussion
3.1. Results

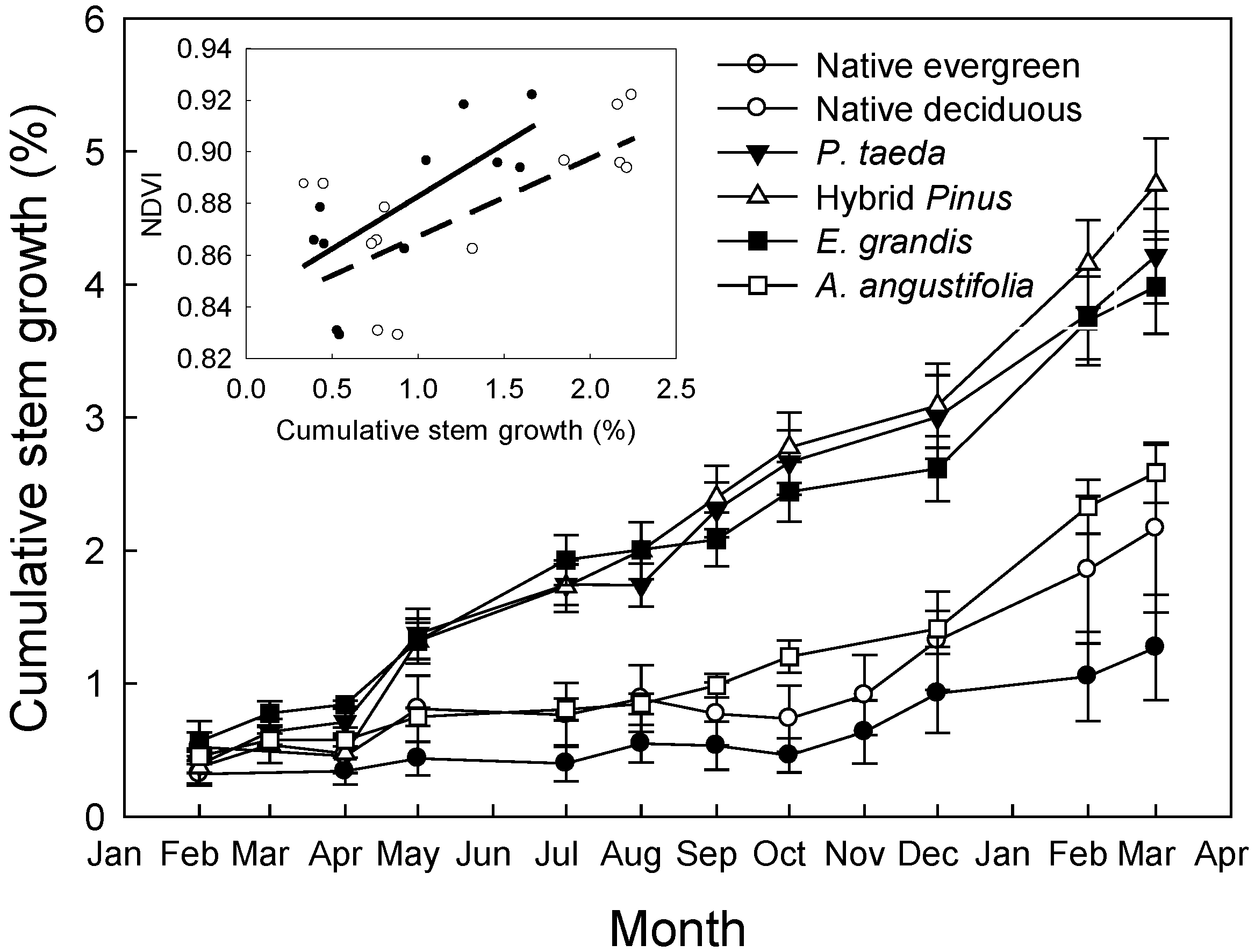
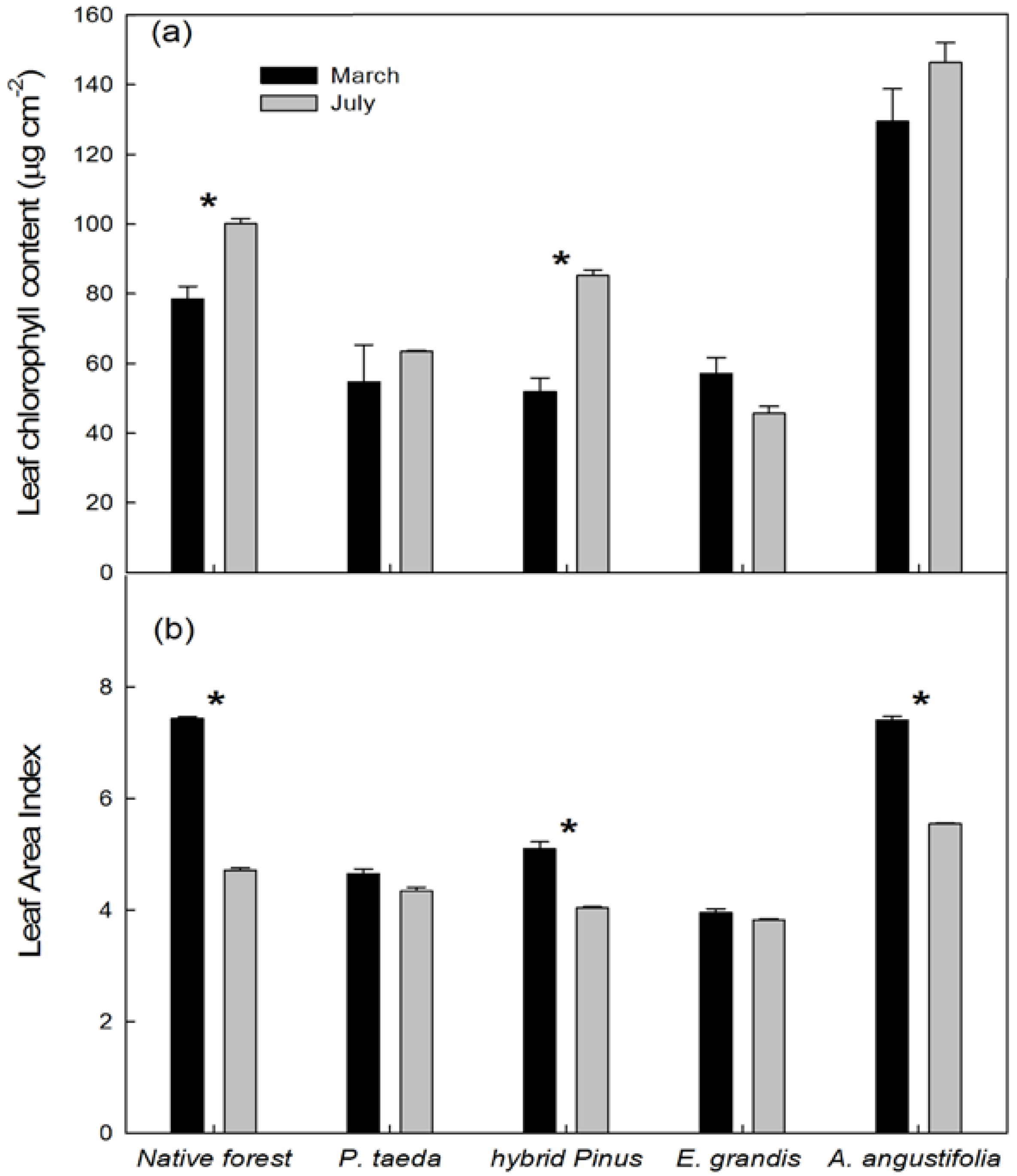
| Study sites | Native forests | E. grandis | A. angustifolia | P. taeda | ||||
|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 1 | 2 | 1 | 1 | 2 | |
| Mean NDVI | 0.88 | 0.88 | 0.89 | 0.85 | 0.87 | 0.83 | 0.86 | 0.87 |
| Intra-annual CV | 2.2 | 2.0 | 1.6 | 1.8 | 2.6 | 2.2 | 2.1 | 2.0 |
| Inter-annual CV | 0.8 | 0.9 | 0.9 | 2.9 | 7.3 | 4.5 | 14.6 | 13.6 |
| Mean LAI | 5.59 | 5.58 | 5.60 | 5.67 | 5.62 | 5.21 | 5.96 | 6.09 |
| Intra-annual CV | 6.5 | 5.8 | 5.8 | 9.0 | 6.4 | 14.4 | 6.0 | 4.7 |
| Inter-annual CV | 4.9 | 3.8 | 3.3 | 2.9 | 13.7 | 6.5 | 3.6 | 2.8 |


| Study Sites | Native Forests | E. grandis | P. taeda | A. angustifolia | ||||
|---|---|---|---|---|---|---|---|---|
| NDVI | LAI | NDVI | LAI | NDVI | LAI | NDVI | LAI | |
| Initial date | 1 January | 1 January | 1 January | 1 January | 7 January | 7 July | 1 January | 1 January |
| Ending date | 12 December | 12 December | 11 December | 11 December | 12 December | 12 December | 11 December | 11 December |
| Period | 12 | 12 | 12 | 12 | 12 | 12 | 12 | 12 |
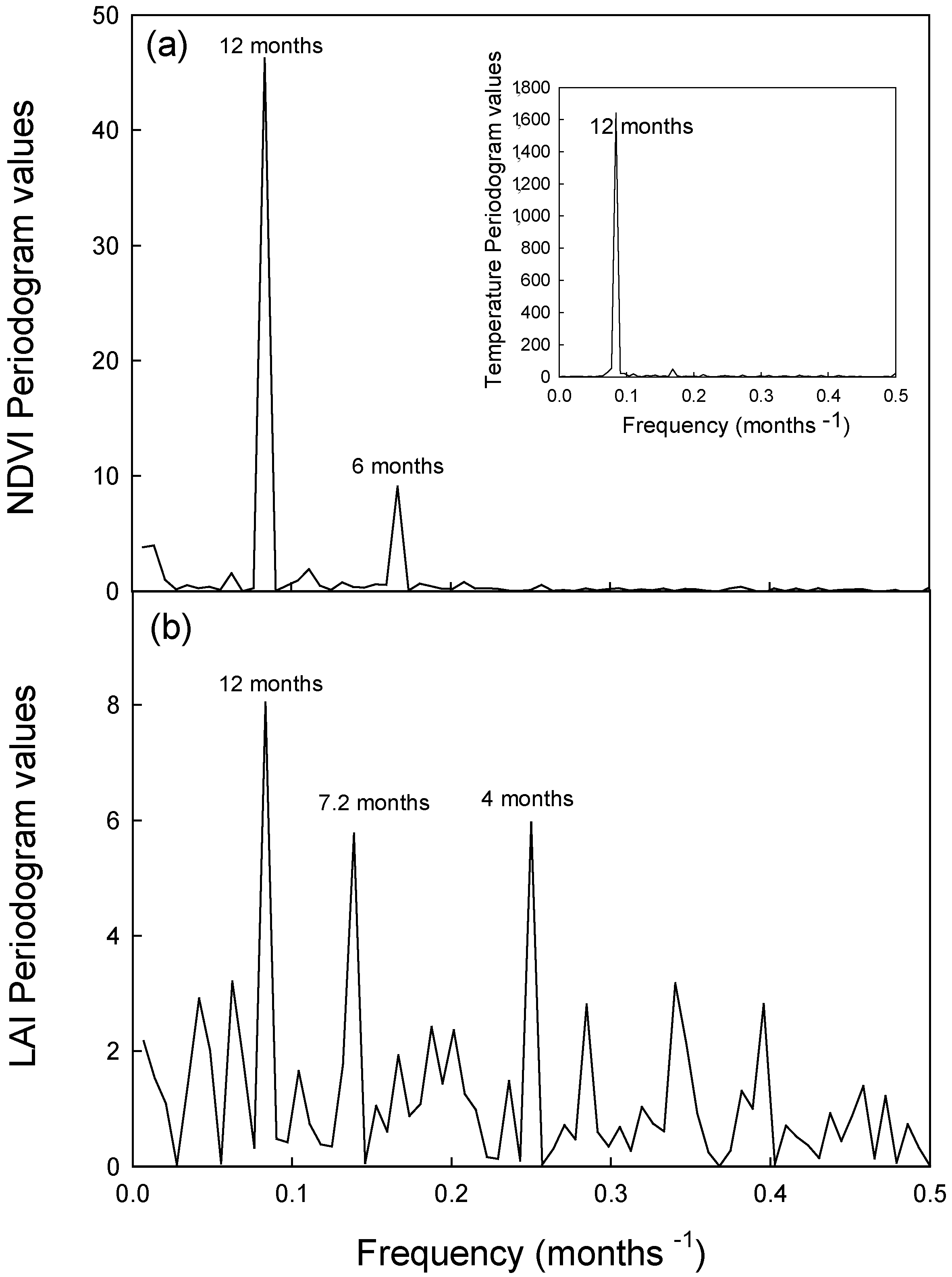
| Var. Explained | 46.2 | 8.0 | 37.7 | 20.8 | 49.31 | 5.86 | 50.8 | 26.2 |
| Period | 6 | 4 | 132 | 6 | 72 | 10 and 7 | 120 | 11 |
| Var. Explained | 9.1 | 6.0 | 29.4 | 5.0 | 6.6 | 3.6 | 14.7 | 14.4 |

3.2. Discussion
3.2.1. Seasonal Variation in NDVI and Potential Canopy Photosynthesis
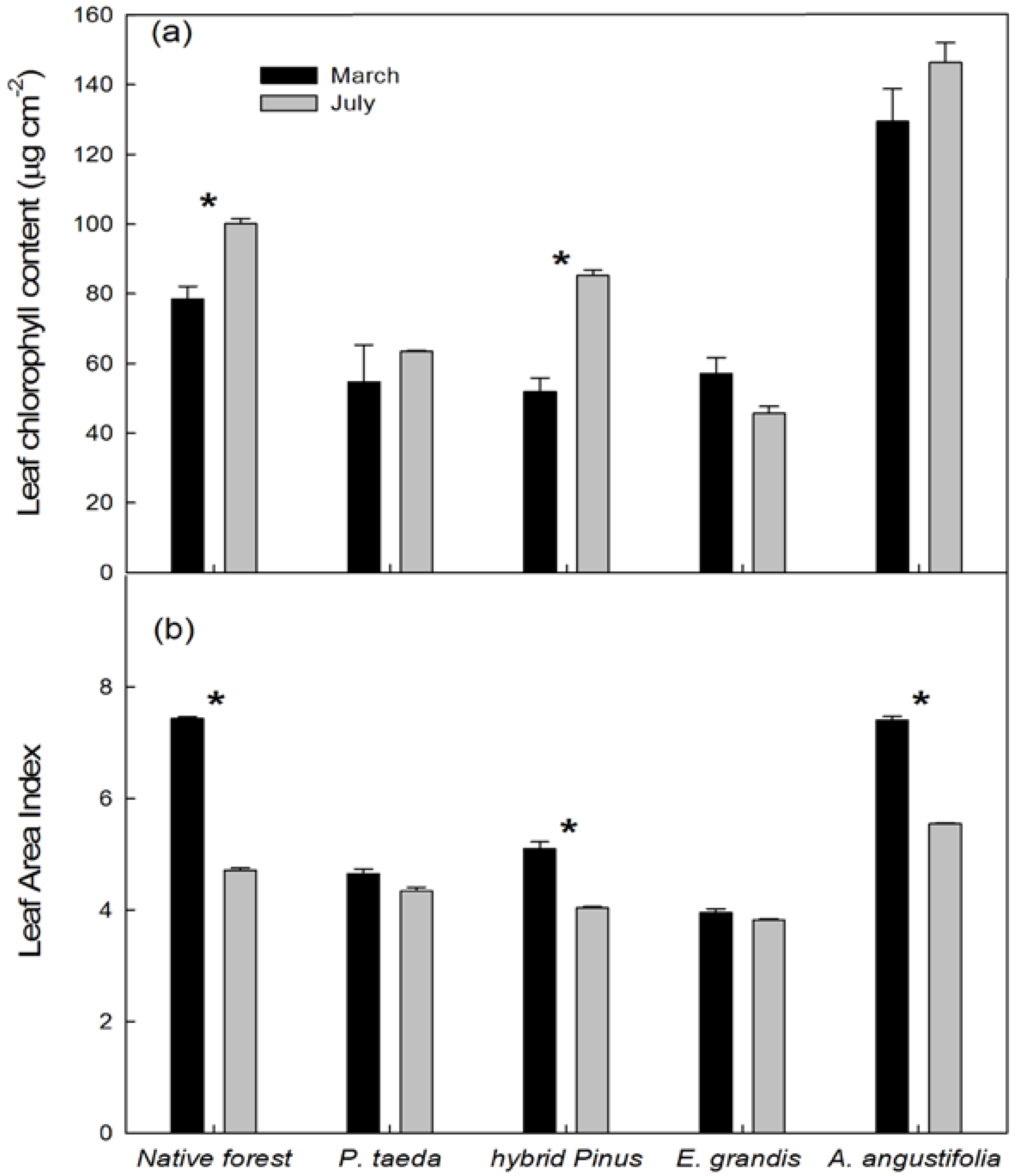
3.2.2. NDVI in Relation to Leaf Area Index, Chlorophyll Content, and Stem Growth Rates
4. Conclusions
Acknowledgments
Conflict of interest
References
- Tan, Z.H.; Zhang, Y.P.; Liang, N.; Hsia, Y.J.; Zhang, Y.J.; Zhou, G.Y.; Li, Y.L.; Juang, J.Y.; Chu, H.S.; Yan, J.H.; et al. An observational study of the carbon-sink strength of East Asian subtropical evergreen forests. Environ. Res. Lett. 2012, 7, 1–12. [Google Scholar]
- Zhang, Y.J.; Yang, Q.Y.; Lee, D.W.; Goldstein, G.; Cao, K.F. Extended leaf senescence promotes carbon gain and nutrient resorption: Importance of maintaining winter photosynthesis in subtropical forests. Oecologia 2013, 173, 721–730. [Google Scholar] [CrossRef]
- Zhang, Y.J. Water and Carbon Balances of Evergreen and Deciduous Broadleaf Trees from a Subtropical Cloud Forest in Southwest China. Ph.D. Thesis, University of Miami, Coral Gables, FL, USA, 7 May 2012. [Google Scholar]
- Zhang, Y.J.; Cao, K.F.; Goldstein, G. Winter Photosynthesis of Evergreen Broadleaf Trees from a Montane Cloud Forest in Subtropical China. In Photosynthesis Research for Food, Fuel and the Future; Springer Berlin: Heidelberg, Germany, 2013; pp. 812–817. [Google Scholar]
- Galindo-Leal, C.; Gusmão Câmara, I. The Status of the Atlantic Forest Hotspot: The Dynamics of Biodiversity Loss; Island Press: Washington, DC, USA, 2003; p. 85. [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; da Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Izquierdo, A.E.; De Angelo, C.D.; Aide, T.M. Thirty years of human demography and land-use change in the Atlantic Forest of Misiones, Argentina: An evaluation of the forest transition model. Ecol. Soc. 2008, 13, 3. [Google Scholar]
- Gatti, M.G.; Campanello, P.I.; Montti, L.F.; Goldstein, G. Frost resistance in the tropical palm Euterpe edulis and its pattern of distribution in the Atlantic Forest of Argentina. For. Ecol. Manag. 2008, 256, 633–640. [Google Scholar] [CrossRef]
- Tucker, C.J. Remote sensing of leaf water content in the near infrared. Remote Sens. Environ. 1980, 10, 23–32. [Google Scholar] [CrossRef]
- Gower, S.T.; Kucharik, C.J.; Norman, J.M. Direct and indirect estimation of leaf area index, fPAR and net primary production of terrestrial ecosystems. Remote Sens. Environ. 1999, 70, 29–51. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Oesterheld, M.; Di Bella, C.; Arzadum, M.; Lafountaine, J.; Cahuepé, M.; Rebella, C.M. Estimation of primary production of subhumid rangelands from remote sensing data. Appl. Veg. Sci. 2000, 3, 189–195. [Google Scholar] [CrossRef]
- Oliveira, T.; Carvalho, L.; Oliveira, L.; Lacerda, W.; Acerbi, F., Jr. NDVI Time Series for Mapping Phonological Variability of Forests Across the Cerrado Biome in Minas Gerais, Brazil. In Phenology and Climate Change; Zhang, X., Ed.; In Tech: Shanghai, China, 2012; pp. 253–272. [Google Scholar]
- Tucker, C.J.; van Praet, C.L.; Sharman, M.J.; van Ittersum, G. Satellite remote sensing of total herbaceous biomass production in the Senegalese Sahel. Remote Sens. Environ. 1985, 17, 233–249. [Google Scholar] [CrossRef]
- Baret, F.; Guyot, G. Potentials and limits of vegetation indices for LAI and APAR assessment. Remote Sens. Environ. 1991, 35, 161–173. [Google Scholar] [CrossRef]
- Gamon, J.A.; Field, C.B.; Goulden, M.L.; Griffin, K.L.; Hartley, A.E.; Joel, G.; Valentini, R. Relationships between NDVI, canopy structure and photosynthesis in three Californian vegetation types. Ecol. Appl. 1995, 5, 28–41. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.Q.; Batchily, K.; van Leeuwen, W. A comparison of vegetation indices global set of TM images for EOSMODIS. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- Paruelo, J.M.; Epstein, H.E.; Lauenroth, W.K.; Burke, I.C. ANPP estimates from NDVI for the central grassland region of the US. Ecology 1997, 78, 953–958. [Google Scholar] [CrossRef]
- Gamon, J.; Field, C.; Goulden, M.; Griffin, K.; Hartley, A.; Joel, G.; Peñuelas, J.; Valentini, R. Relationship between NDVI, canopy structure, and photosynthesis in three Californian vegetation types. Ecol. Appl. 1995, 5, 28–41. [Google Scholar] [CrossRef]
- Muraoka, H.; Noda, H.; Nagai, S.; Motohka, T.; Saitoh, T.; Nasahara, K.; Saigusa, N. Spectral vegetation indices as the indicator of canopy photosynthetic productivity in a deciduous broadleaf forest. J. Plant Ecol. Adv. Access 2013, 6, 393–407. [Google Scholar] [CrossRef]
- De Kauwe, M.G.; Disney, M.I.; Quaife, T.; Lewis, P.; Williams, M. An assessment of the MODIS collection 5 leaf area index product for a region of mixed coniferous forest. Remote Sens. Environ. 2011, 115, 767–780. [Google Scholar] [CrossRef]
- Rautiainen, M.; Heiskanen, J.; Korhonen, L. Seasonal changes in canopy leaf area index and MODIS vegetation products for a boreal forest site in central Finland. Boreal Environ. Res. 2011, 17, 71–84. [Google Scholar]
- Ligier, H.D.; Matteio, H.R.; Polo, H.L.; Rosso, J.R. Provincia de Misiones. In Atlas de suelos de la República Argentina, Volumen II; Centro de Investigaciones de Recursos Naturales, INTA: Buenos Aires, Argentina, 1990; pp. 111–154. [Google Scholar]
- Soil Survey Staff. Keys to Soil Taxonomy, SMSS Technical Monograph No. 19, 5th ed.; Pocahontas Press Inc.: Blacksburg, VA, USA, 1992; p. 556. [Google Scholar]
- Pinkard, E.A.; Patel, V.; Mohammed, C. Chlorophyll and nitrogen determination for plantation-grown Eucalyptus nitens and E. globulus using a non-destructive meter. For. Ecol. Manag. 2006, 223, 211–217. [Google Scholar] [CrossRef]
- Coste, S.; Baraloto, C.; Leroy, C.; Marcon, E.; Renaud, A.; Richardson, A.D.; Roggy, J.C.; Schimann, H.; Uddling, J.; Herault, B. Assessing foliar chlorophyll contents with the SPAD-502 chlorophyll meter: A calibration test with thirteen tree species of tropical rainforest in French Guiana. Ann. For. Sci. 2010, 67, 607. [Google Scholar] [CrossRef]
- Cattelino, P.J.; Becker, C.A.; Fuller, L.G. Construction and installation of manmade dendrometer bands. North. J. Appl. For. 1986, 3, 73–75. [Google Scholar]
- Running, S.W.; Justice, C.O.; Salomonson, V.; Hall, D.; Barker, J.; Kaufmann, Y.J.; Strahler, A.H.; Huete, A.R.; Muller, J.P.; Vanderbilt, V.; et al. Terrestrial remote sensing science and algorithms planned for EOS/MODIS. Int. J. Remote Sens. 1994, 15, 3587–3620. [Google Scholar] [CrossRef]
- Tedrow, L.; Weber, K.T. NDVI Changes Over a Calendar Year in the Rangelands of Southeast Idaho. In Final Report: Assessing Post-Fire Recovery of Sagebrush-Steppe Rangelands in Southeastern Idaho (NNX08AO90G); Weber, K.T., Davis, K., Eds.; Idaho State University: Pocatello, ID, USA, 2011; pp. 105–116. [Google Scholar]
- Gessner, U.; Niklaus, M.; Kuenzer, C.; Dech, S. Intercomparison of leaf area index products for a Gradient of sub-humid to arid environments in West Africa. Remote Sens. 2013, 5, 1235–1257. [Google Scholar] [CrossRef]
- Myneni, R.B.; Hoffman, S.; Knyazikhin, Y.; Privette, J.L.; Glassy, J.; Tian, Y.; Wang, Y.; Song, X.; Zhang, Y.; Smith, G.R. Global products of vegetation leaf area and fraction absorbed PAR from year one of MODIS data. Remote Sen. Environ. 2002, 83, 214–231. [Google Scholar] [CrossRef]
- Jacquemoud, S.; Verhoef, W.; Baret, F.; Bacour, C.; Zarco-Tejada, P.J.; Asner, G.P.; Francois, C.; Ustin, S.L. PROSPECT+SAIL models: A review of use for vegetation characterization. Remote Sens. Environ. 2009, 113, 556–566. [Google Scholar]
- Land Processes Distributed Active Archive Center. Available online: http://lpdaac.usgs.gov (accessed on Day September 2013).
- Bloomfield, P. Fourier Analysis of Time Series: An Introduction, 2nd ed.; Wiley: New York, NY, USA, 1976; p. 257. [Google Scholar]
- Elliott, D.F.; Rao, K.R. Fast Transforms: Algorithms, Analyses, Applications; Academic Press: New York, NY, USA, 1982; p. 448. [Google Scholar]
- Wei, W.W. Time Series Analysis: Univariate and Multivariate Methods; Addison-Wesley: New York, NY, USA, 1989; p. 614. [Google Scholar]
- Hashemi, S.A.; Chenani, S.K. Investigation of NDVI index in Relation to Chlorophyll Content Change and Phenological Event. In Recent Advances in Environment, Energy Systems and Naval Science, Proceedings of the 4th International Conference on Environment, Barcelona, Spain, 15–17 September 2011; Wseas LLC: City, State Abbr., USA, 2011; pp. 22–28. [Google Scholar]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Holaday, A.S.; Martindale, W.; Aired, R.; Brooks, A.; Leegood, R.C. Changes in activities of enzymes of carbon metabolism in leaves during exposure to low temperature. Plant Physiol. 1992, 98, 1105–1114. [Google Scholar] [CrossRef]
- Campanello, P.I.; Gatti, M.G.; Goldstein, G. Coordination between water-transport efficiency and photosynthetic capacity in canopy tree species at different growth irradiances. Tree Physiol. 2008, 28, 85–94. [Google Scholar] [CrossRef]
- Zhang, Y.J.; Harvard University, Cambridge, MA, USA. Carbon balance of an Asian subtropical cloud forest. Unpublished work, 2014.
- Soudani, K.; Hmimina, G.; Delpierre, N.; Pontailler, J.Y.; Aubinet, M.; Bonal, D.; Caquet, B.; de Grandcourt, A.; Burban, B.; Flechard, C.; et al. Ground-Based Network of NDVI measurements for tracking temporal dynamics of canopy structure and vegetation phenology in different biomes. Remote Sens. Environ. 2012, 123, 234–245. [Google Scholar] [CrossRef]
- Kurek, I.; Chang, T.K.; Bertain, S.M.; Madrigal, A.; Liu, L.; Lassner, M.W.; Zhu, G. Enhanced thermostability of Arabidopsis Rubisco activase improves photosynthesis and growth rates under moderate heat stress. Plant Cell 2007, 19, 3230–3241. [Google Scholar] [CrossRef]
- Vargas, G.G.; Cordero, S.R.A. Photosynthetic responses to temperature of two tropical rainforest tree species from Costa Rica. Trees 2013, 27, 1261–1270. [Google Scholar] [CrossRef]
- Sharkey, T.D. Effects of moderate heat stress on photosynthesis: Importance of thylakoid reactions, rubisco deactivation, reactive oxygen species, and thermotolerance provided by isoprene. Plant Cell Environ. 2005, 28, 269–277. [Google Scholar] [CrossRef]
- Lloyd, J.; Farquhar, G.D. Effects of rising temperatures and CO2 on the physiology of tropical forest trees. Philos. Trans. R. Soc. Ser. B 2008, 363, 1811–1817. [Google Scholar] [CrossRef]
- Berry, J.A.; Bjorkman, O. Photosynthetic response and adaptation to temperature in higher plants. Ann. Rev. Plant Physiol. 1980, 31, 491–543. [Google Scholar] [CrossRef]
- Goldstein, G.; Meinzer, F.; Rada, F. Environmental biology of a tropical treeline species, Polylepis sericea. In Tropical Alpine Environments: Plants Form and Function; Rundel, P.W., Smith, A.P., Meinzer, F.C., Eds.; Cambridge University Press: Cambridge, UK, 1994; pp. 129–149. [Google Scholar]
- Larigauderie, A.; Korner, C. Acclimation to leaf dark respiration to temperature in Alpine and lowland plant species. J. Botan. 1995, 76, 245–252. [Google Scholar]
- Yang, W.; Shabanov, N.V.; Huang, D.; Wang, W.; Dickinson, R.E.; Nemani, R.R.; Knyazikhin, Y.; Myneni, R.B. Analysis of leaf area index products from combination of MODIS Terra and Aqua data. Remote Sens. Environ. 2006, 104, 297–312. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Cristiano, P.M.; Madanes, N.; Campanello, P.I.; Di Francescantonio, D.; Rodríguez, S.A.; Zhang, Y.-J.; Carrasco, L.O.; Goldstein, G. High NDVI and Potential Canopy Photosynthesis of South American Subtropical Forests despite Seasonal Changes in Leaf Area Index and Air Temperature. Forests 2014, 5, 287-308. https://doi.org/10.3390/f5020287
Cristiano PM, Madanes N, Campanello PI, Di Francescantonio D, Rodríguez SA, Zhang Y-J, Carrasco LO, Goldstein G. High NDVI and Potential Canopy Photosynthesis of South American Subtropical Forests despite Seasonal Changes in Leaf Area Index and Air Temperature. Forests. 2014; 5(2):287-308. https://doi.org/10.3390/f5020287
Chicago/Turabian StyleCristiano, Piedad M., Nora Madanes, Paula I. Campanello, Débora Di Francescantonio, Sabrina A. Rodríguez, Yong-Jiang Zhang, Laureano Oliva Carrasco, and Guillermo Goldstein. 2014. "High NDVI and Potential Canopy Photosynthesis of South American Subtropical Forests despite Seasonal Changes in Leaf Area Index and Air Temperature" Forests 5, no. 2: 287-308. https://doi.org/10.3390/f5020287
APA StyleCristiano, P. M., Madanes, N., Campanello, P. I., Di Francescantonio, D., Rodríguez, S. A., Zhang, Y.-J., Carrasco, L. O., & Goldstein, G. (2014). High NDVI and Potential Canopy Photosynthesis of South American Subtropical Forests despite Seasonal Changes in Leaf Area Index and Air Temperature. Forests, 5(2), 287-308. https://doi.org/10.3390/f5020287