American Chestnut Growth and Survival Five Years after Planting in Two Silvicultural Treatments in the Southern Appalachians, USA

Abstract
:1. Introduction
2. Methods
2.1. Study Site and Experimental Material
2.2. Study Sites and Treatments

{kind=link}
{kind=link}
| Moisture regime | Elevation (m) | Landform | Aspect (degrees) | Steepness (percent) | Surface shape | Site index for Q. rubra (m, base age 50) |
|---|---|---|---|---|---|---|
| Mesic | 880 | Cove | 45–74 | 45–74 | Linear-concave | 26.2 |
| Subxeric | 780 | Slope | 195–282 | 38–43 | Linear-convex | 20.1 |
| Xeric | 760 | Ridge | 329–345 | 39–46 | Linear-convex | 17.7 |
2.3. Data Collection
2.4. Statistical Analysis
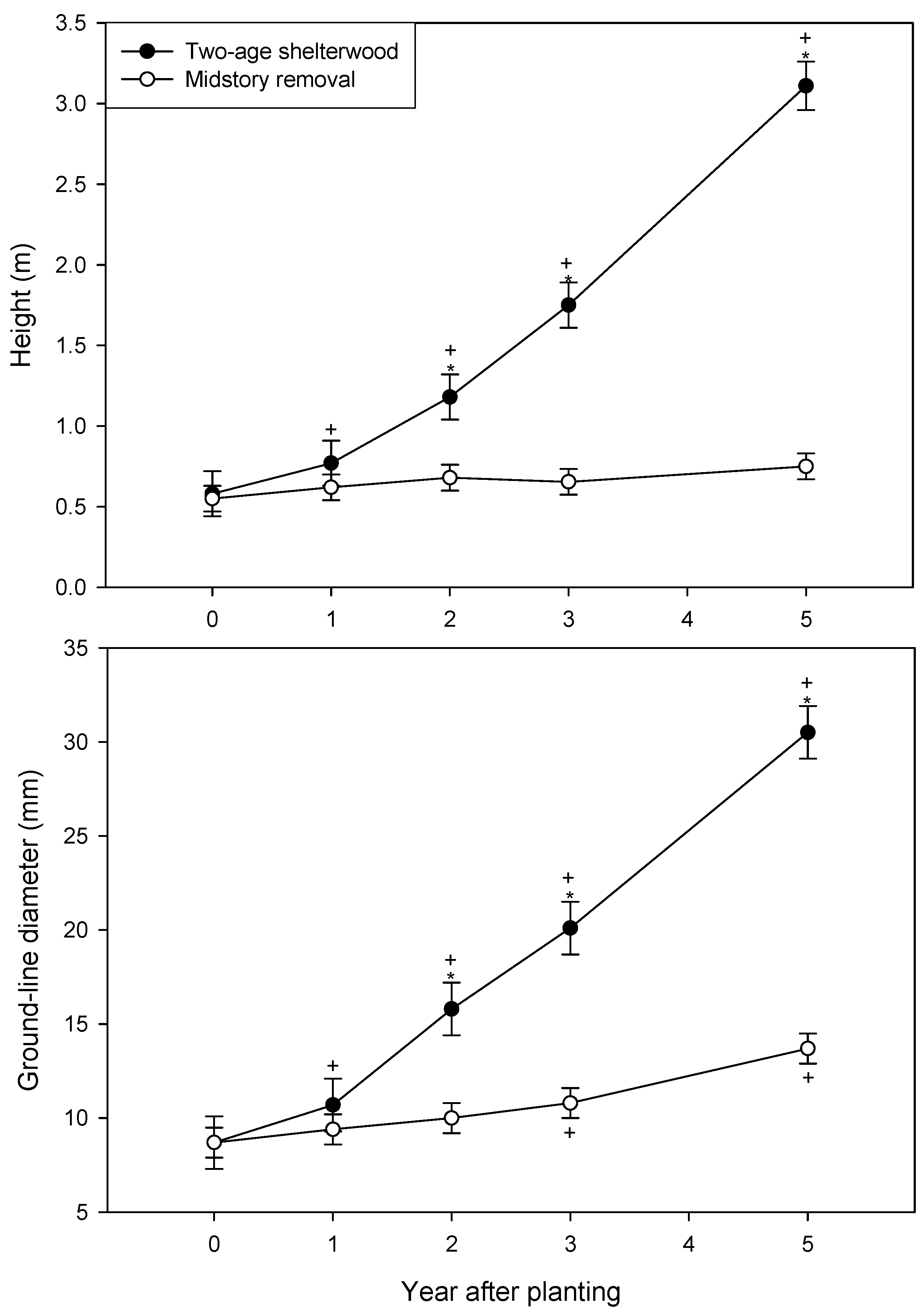
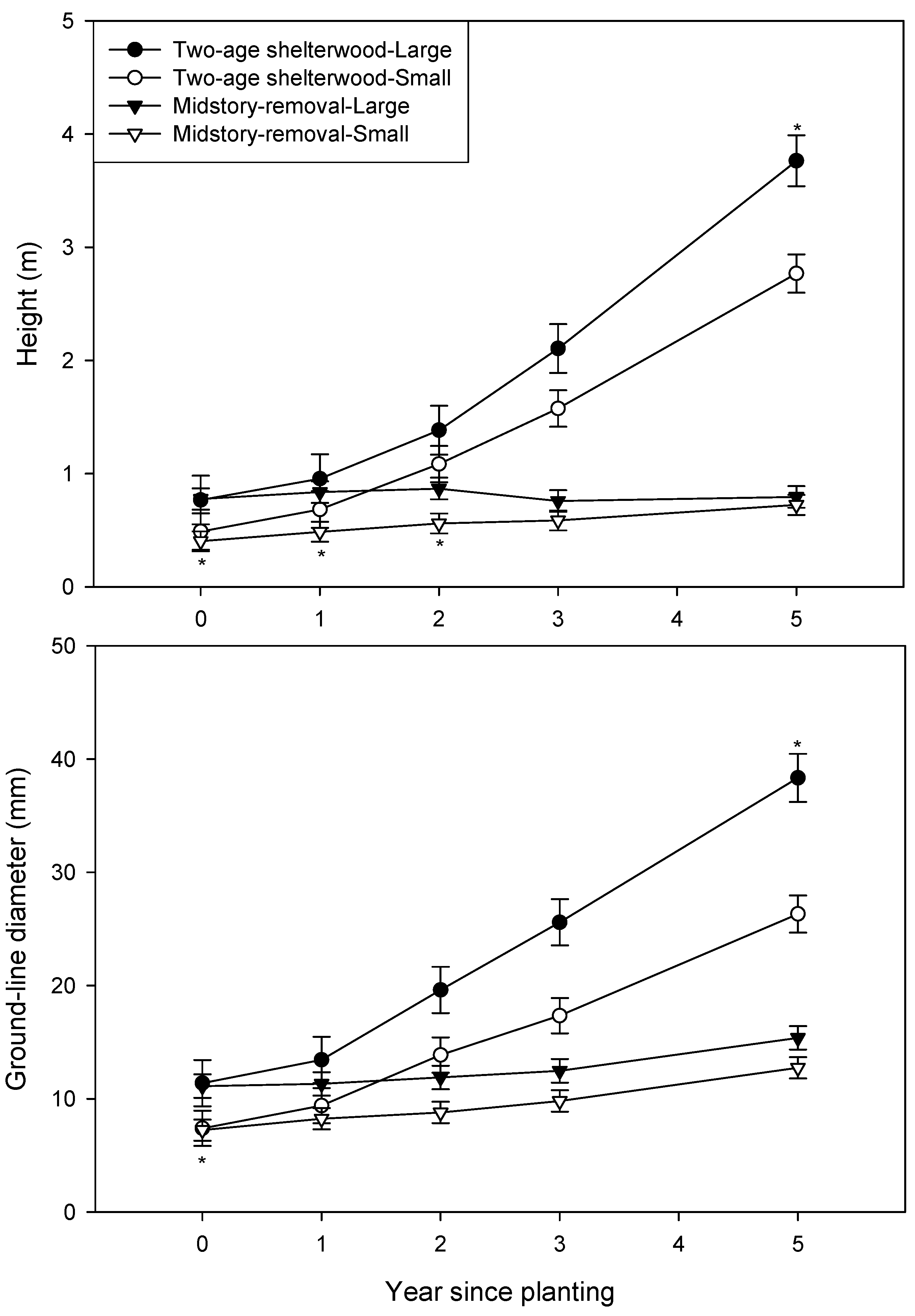
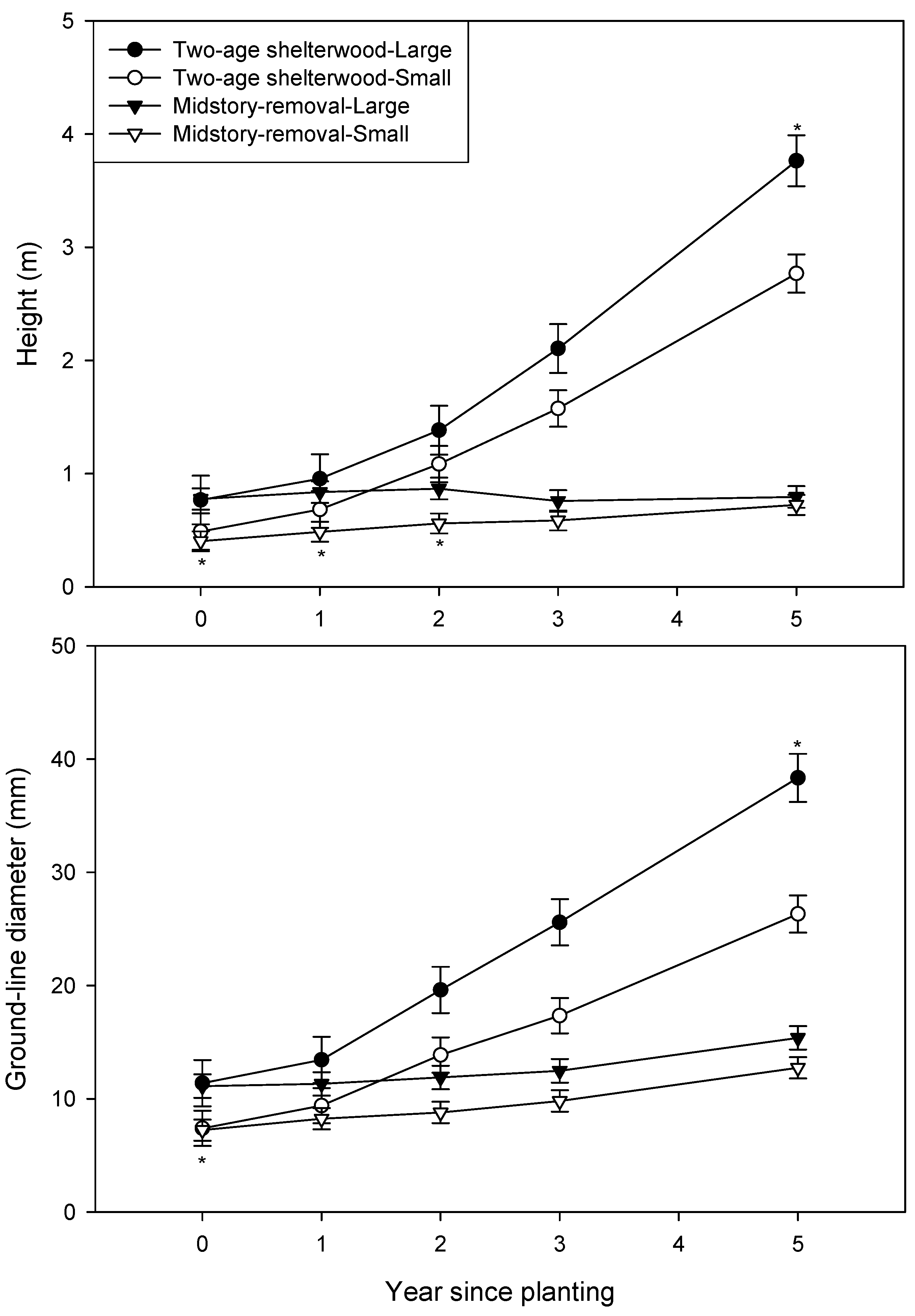
3. Results
| Year after planting | Two-age shelterwood | MidstoryRemoval | All |
|---|---|---|---|
| 1 | 84 (8) | 87 (8) | 86 (2) |
| 2 | 78 (11) | 81 (11) | 79 (3) |
| 3 | 75 (10) | 75 (11) | 75 (3) |
| 5 | 63 (11) | 72 (11) | 67 (3) |

| Year after planting | Two-age shelterwood | Midstory | Across treatments | Number of trees examined for blight |
|---|---|---|---|---|
| 1 | 0 (3) b | 1 (3) b | 0 (2) C | 213 |
| 2 | 10 (3) b | 11 (3) a,b | 11 (2) B | 181 |
| 3 | 15 (3) a,b | 9 (3) b | 12 (2) A,B | 169 |
| 5 | 28 (3) a | 13 (3) a,b | 21 (2) A | 160 |
| Year after planting | Height (m) | Ground-line diameter (mm) | ||
|---|---|---|---|---|
| Free-to-grow | Overtopped | Free-to-grow | Overtopped | |
| 2 | 1.08 d | 1.24 d | 15.2 c | 16.3 c |
| 3 | 1.80 b,c | 1.44 c,d | 21.1 b | 16.3 c |
| 5 | 3.52 a | 1.99 b | 34.5 a | 20.0 b |
4. Discussion
5. Conclusions
Acknowledgments
Conflict of Interest
References
- Delcourt, H.R.; Delcourt, P.A. Late-quaternary vegetational dynamics and community stability reconsidered. Quat. Res. 1983, 19, 265–271. [Google Scholar] [CrossRef]
- Burnham, C.R. The restoration of the American chestnut. Am. Sci. 1988, 76, 478–487. [Google Scholar]
- Russell, E.W.B. Pre-blight distribution of Castanea dentata (Marsh.) Borkh. Bull. Torrey Bot. Club 1987, 114, 183–190. [Google Scholar]
- Hepting, G.H. Death of the American chestnut. J. For. Hist. 1974, 18, 60–67. [Google Scholar]
- Ashe, W.W. Chestnut in Tennessee; Bulletin Number 10-B; Tennessee Geological Survey: Nashville, TN, USA, 1911. [Google Scholar]
- Emerson, A.I.; Weed, C.M. Our Trees and How to Know Them; J. B. Lippincott Company: Philadelphia, PA, USA, 1908. [Google Scholar]
- Anagnostakis, S.L. The pathogens and pests of chestnuts. Adv. Bot. Res. 1995, 21, 125–145. [Google Scholar] [CrossRef]
- Paillet, F.L. Chestnut: History and ecology of a transformed species. J. Biogeogr. 2002, 29, 1517–1530. [Google Scholar] [CrossRef]
- Anagnostakis, S.L. The effect of multiple importations of pests and pathogens on a native tree. Biol. Invasions 2001, 3, 245–254. [Google Scholar] [CrossRef]
- Burnham, C.R.; Rutter, P.A.; French, D.W. Breeding blight-resistant chestnuts. Plant Breed. Rev. 1986, 4, 347–397. [Google Scholar]
- Clark, S.L.; Schlarbaum, S.E.; Saxton, A.M.; Hebard, F.V. Making history: Field testing of blight-resistant American chestnut (Castanea dentata) in the southern region. In Proceedings of the 17th Central Hardwood Forest Conference, Lexington, KY, USA, 5–7 April 2010; Fei, S., Lhotka, J.M., Stringer, J.W., Gottschalk, K.W., Miller, G.W., Eds.; USDA Forest Service: Newtown Square, PA, USA, 2011; pp. 656–657. [Google Scholar]
- Jacobs, D.F. Toward development of silvical strategies for forest resoration of American chestnut (Castanea dentata) using blight-resistant hybrids. Biol. Conserv. 2007, 137, 497–506. [Google Scholar] [CrossRef]
- Griffin, G.J. Blight control and restoration of the American chestnut. J. For. 2000, 98, 22–27. [Google Scholar]
- Clark, S.L.; Schlarbaum, S.E.; Saxton, A.M.; Hebard, F.V.; Blanton, J.; Casey, D.; Crane, B.; MacFarlane, R.; Rodrigue, J.; Stelick, J. Lessons from the field: The first tests of restoration American chestnut (Castanea dentata) seedlings planted in the Southern Region. In Proceedings of the 16th Biennial Southern Silvicultural Research Conference, Charleston, SC, USA, 15–16 February 2011; Butner, J.R., Ed.; USDA Forest Service: Asheville, NC ,USA, 2012; pp. 69–70. [Google Scholar]
- Rhoades, C.; Loftis, D.; Lewis, J.; Clark, S. The influence of silvicultural treatments and site conditions on American chestnut (Castanea dentata) seedling establishment in eastern Kentucky, USA. For. Ecol. Manag. 2009, 258, 1211–1218. [Google Scholar] [CrossRef]
- Joesting, H.M.; McCarthy, B.C.; Brown, K.J. The photosynthetic response of American chestnut seedlings to differing light conditions. Can. J. For. Res. 2007, 37, 1714–1722. [Google Scholar] [CrossRef]
- Wang, G.G.; Bauerle, W.L.; Mudder, B.T. Effects of light acclimation on the photosynthesis, growth, and biomass allocation in American chestnut (Castanea dentata) seedlings. For. Ecol. Manag. 2006, 226, 173–180. [Google Scholar] [CrossRef]
- Latham, R.E. Co-occurring tree species change rank in seedling performance with resources varied experimentally. Ecology 1992, 73, 2129–2144. [Google Scholar] [CrossRef]
- McCament, C.L.; McCarthy, B.C. Two-year response of American chestnut (Castanea dentata) seedlings to shelterwood harvesting and fire in a mixed-oak forest ecosystem. Can. J. For. Res. 2005, 35, 740–749. [Google Scholar] [CrossRef]
- McNab, W.H.; Patch, S.; Nutter, A.A. Early results from a pilot test of planting small American chestnut seedlings under a forest canopy. J. Am. Chestnut Found. 2003, 16, 32–41. [Google Scholar]
- Griffin, G.J. Incidence of chestnut blight and survival of American chestnut in forest clear-cut and neighboring understory sites. Plant Dis. 1989, 73, 123–127. [Google Scholar] [CrossRef]
- Spetich, M.A.; Dey, D.C.; Johnson, P.S.; Graney, D.L. Competitive capacity of Quercus rubra L. planted in Arkansas Boston Mountains. For. Sci. 2002, 48, 504–517. [Google Scholar]
- Schweitzer, C.J.; Dey, D.C. Forest structure, composition, and tree diversity response to a gradient of regeneration harvests in the mid-Cumberland Plateau escarpment region, USA. For. Ecol. Manag. 2011, 262, 1729–1741. [Google Scholar] [CrossRef]
- Clark, S.L.; Schweitzer, C.J.; Schlarbaum, S.E.; Dimov, L.D.; Hebard, F.V. Nursery quality and first-year response of American chestnut (Castanea dentata) seedlings planted in the southeastern United States. Tree Planters Notes 2009, 53, 13–21. [Google Scholar]
- Loftis, D.L. A shelterwood method for regenerating red oak in the southern Appalachians. For. Sci. 1990, 36, 917–929. [Google Scholar]
- Jones, C.; Griffin, G.J.; Elkins, J.R. Association of climatic stress with blight on Chinese chestnut in the eastern United States. Plant Dis. 1980, 64, 1001–1004. [Google Scholar]
- United State Department of Agriculture; Forest Service. In Land and Resource Management Plan, Cherokee National Forest; Management Bulletin 8-MB-114A; USDA Forest Service: Southern Region, Atlanta, GA, USA, 2004.
- United State Department of Agriculture; Forest Service Southern Region. In Land and Resource Management Plan, Jefferson National Forest; Management Bulletin R8-MB-115A; USDA Forest Service: Southern Region, Atlanta, GA, USA, 2004.
- United State Department of Agriculture; Forest Service Southern Region. In Land and Resource Management Plan for the Daniel Boone National Forest; Management Bulletin R8-MB-117A; USDA Forest Service: Southern Region, Atlanta, GA, USA, 2004.
- United State Department of Agriculture; Forest Service Southern Region. Environmental Assessment, Upper Santeetlah Project. Cheoah Ranger District, Nantahala National Forest. USDA Forest Service Nantahala National Forest: Robbinsville, NC, USA. Available online: http://www.fs.fed.us/nepa/fs-usda-pop.php/?project=23585 (assessed on 10 August 2012).
- Gravatt, G.F.; Gill, L.S. Chestnut Blight; United States Department of Agriculture Farmers Bulletin 1641; United States Government Printing Office: Washington, DC, USA, 1930. [Google Scholar]
- Braun, E.L. Deciduous Forests of Eastern North America; Blakiston Co.: Philadelphia, PA, USA, 1950. [Google Scholar]
- Keever, C. Present composition of some stands of the former oak-chestnut forest in the Southern Blue Ridge Mountains. Ecology 1953, 34, 44–54. [Google Scholar] [CrossRef]
- Woods, F.W.; Shanks, R.E. Replacement of chestnut by other species in the Great Smoky Mountains National Park. Ecology 1959, 40, 349–361. [Google Scholar] [CrossRef]
- Beck, D.E. Management options for southern Appalachian hardwoods: The two-aged stand. In Proceedings of the Fourth Biennial Southern Silvicultural Research Conference, Atlanta, GA, USA, 4–6 November 1986; Phillips, D.R., Ed.; USDA Forest Service: Asheville, NC, USA, 1986; pp. 451–454. [Google Scholar]
- Griffin, G.J.; Elkins, J.R. Chestnut Blight. In Chestnut Blight, Other Endothia Diseases, and the Genus Endothia; Roane, M.K., Griffin, G.J., Elkins, J.R., Eds.; APS Press: St. Paul, MN, USA, 1986; pp. 1–26. [Google Scholar]
- SAS Institute Inc. SAS/STAT User’s Guide, Version 9, 2nd ed; SAS Institute Inc.: Cary, NC, USA, 2009.
- Littell, R.C.; Henry, P.R.; Ammerman, C.B. Statistical analysis of repeated measures data using SAS procedures. J. Anim. Sci. 1998, 76, 1216–1231. [Google Scholar]
- Design and Analysis Web Guide. DAWG Web site. Available online: http://dawg.utk.edu/ (accessed on 15 June 2011).
- Hebard, F.V. Backcross breeding program produces blight-resistant American chestnuts (Virginia). Ecol. Restor. 2001, 19, 252–254. [Google Scholar]
- Joesting, H.M.; McCarthy, B.C.; Brown, K.J. Determining the shade tolerance of American chestnut using morphological and physiological leaf parameters. For. Ecol. Manag. 2009, 257, 280–286. [Google Scholar] [CrossRef]
- Paillet, F.L. Growth form and life histories of American chestnut and Allegheny and Ozark chinquapin at various North American sites. Bull. Torrey Bot. Club 1993, 120, 257–268. [Google Scholar] [CrossRef]
- Oswalt, C.M.; Clatterbuck, W.K.; Houston, A.E. Impacts of deer herbivory and visual grading on the early performance of high-quality oak planting stock in Tennessee, USA. For. Ecol. Manage. 2006, 229, 128–135. [Google Scholar] [CrossRef]
- Loftis, D.L. Northern Red Oak Performs Poorly in North Carolina Planting; Research Note SE-277; USDA Forest Service: Asheville, NC, USA, 1979. [Google Scholar]
- Schuler, J.L.; Robison, D.J. Performance of northern red oak enrichment plantings in naturally regenerating Southern Appalachian hardwood stands. New For. 2010, 40, 119–130. [Google Scholar] [CrossRef]
- Jacobs, D.F.; Severeid, L.R. Dominance of interplanted American chestnut (Castanea dentata) in southwestern Wisconsin, USA. For. Ecol. Manag. 2004, 191, 111–120. [Google Scholar] [CrossRef]
- Clark, S.L.; Schlarbaum, S.E.; Kormanik, P.P. Visual grading and quality of 1-0 northern red oak seedlings. South. J. Appl. For. 2000, 24, 93–97. [Google Scholar]
- Paillet, F.L. Growth form and ecology of American chestnut (Castanea dentate) sprout clones in northeastern Massachusetts. Bull. Torrey Bot. Club 1984, 111, 316–328. [Google Scholar] [CrossRef]
- Paillet, F.L. Character and distribution of American chestnut sprouts in southern New England woodlands. Bull. Torrey Bot. Club 1988, 115, 32–44. [Google Scholar] [CrossRef]
- Dey, D.C.; Jacobs, D.F.; McNabb, K.; Miller, G.; Baldwin, V.; Foster, G. Artificial regeneration of major oak (Quercus) species in the eastern United States—A review of the literature. For. Sci. 2008, 54, 77–106. [Google Scholar]
- Kormanik, P.P.; Kormanik, T.L.; Sung, S.J.S.; Zarnoch, S.J.; Possee, C. Artificial regeneration of multiple hardwood species to develop specific forest communities. In Proceedings of the 10th Biennial Southern Silvicultural Research Conference, Shreveport, LA, USA, 16–18 February 1999; Haywood, J.D., Ed.; USDA Forest Service: Asheville, NC, USA, 1999; pp. 132–135. [Google Scholar]
- Johnson, P.S. Growing hardwood nursery stock for planting on forest sites with special reference to northern red oak. In Proceedings of the 1989 Northeastern Area Nurseryman’s Conference, Peoria, IL, USA, 24–27 July 1989; Mason State Nursery: Mason County, IL, USA, 1989; pp. 46–62. [Google Scholar]
- Clark, S.L.; Schlarbaum, S.E.; Saxton, A.M.; Hebard, F.V. Nursery performance of American and Chinese chestnuts and backcross generations in commercial tree nurseries. Forestry 2012, 85, 589–600. [Google Scholar] [CrossRef]
- Reynolds, D.L.; Burke, K.L. The effect of growth rate, age, and chestnut blight on American chestnut mortality. Castanea 2011, 76, 129–139. [Google Scholar] [CrossRef]
- Milgroom, M.G.; Cortesi, P. Biological control of chestnut blight with hypovirulence: A critical analysis. Ann. Rev. Phytopathol. 2004, 42, 311–338. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Clark, S.; McNab, H.; Loftis, D.; Zarnoch, S. American Chestnut Growth and Survival Five Years after Planting in Two Silvicultural Treatments in the Southern Appalachians, USA. Forests 2012, 3, 1017-1033. https://doi.org/10.3390/f3041017
Clark S, McNab H, Loftis D, Zarnoch S. American Chestnut Growth and Survival Five Years after Planting in Two Silvicultural Treatments in the Southern Appalachians, USA. Forests. 2012; 3(4):1017-1033. https://doi.org/10.3390/f3041017
Chicago/Turabian StyleClark, Stacy, Henry McNab, David Loftis, and Stanley Zarnoch. 2012. "American Chestnut Growth and Survival Five Years after Planting in Two Silvicultural Treatments in the Southern Appalachians, USA" Forests 3, no. 4: 1017-1033. https://doi.org/10.3390/f3041017