Potential Range Expansion of Japanese Honeysuckle (Lonicera japonica Thunb.) in Southern U.S. Forestlands



Abstract
:
1. Introduction
2. Experimental Section
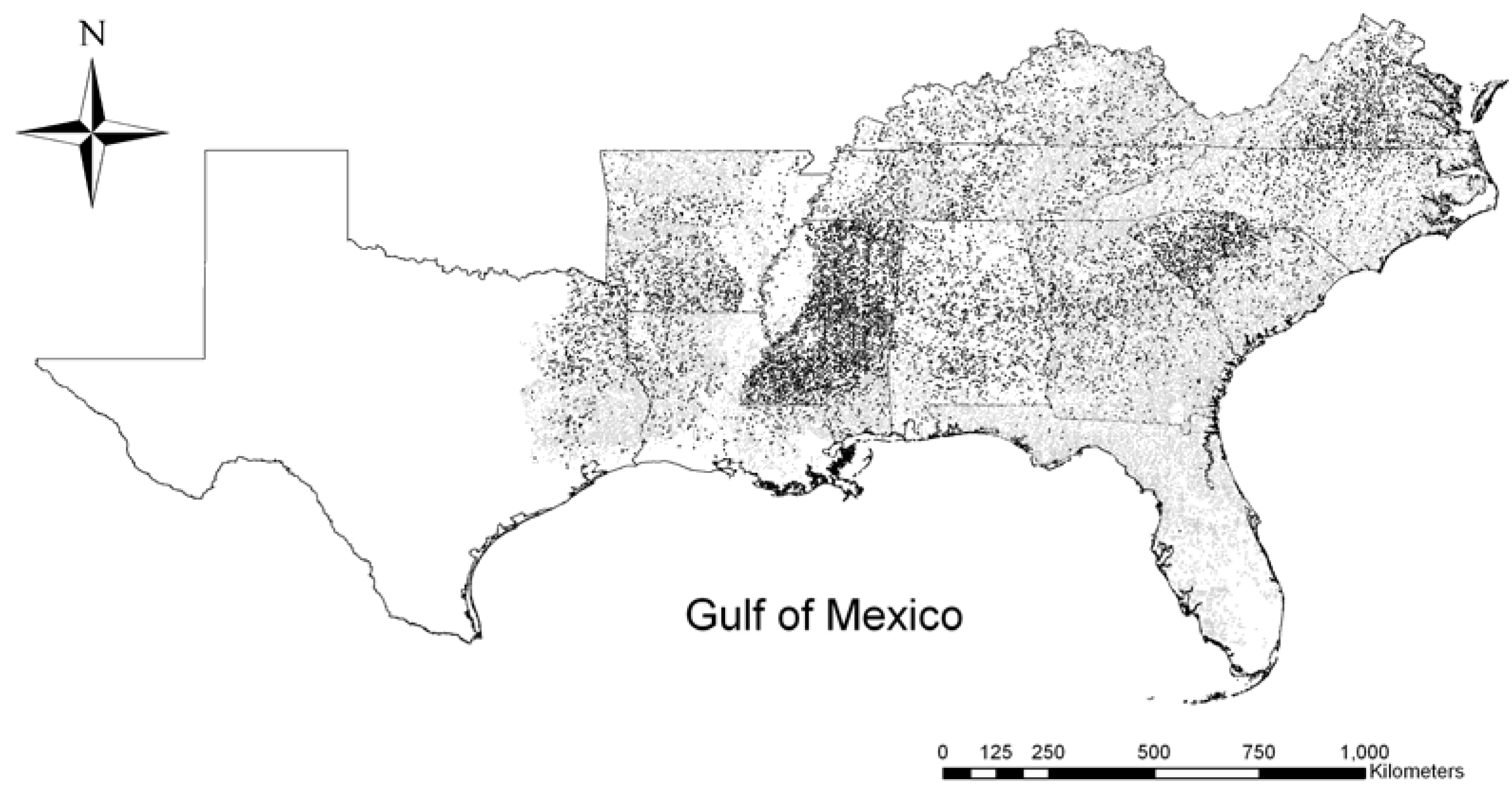
2.1. Study Area and Data Sources
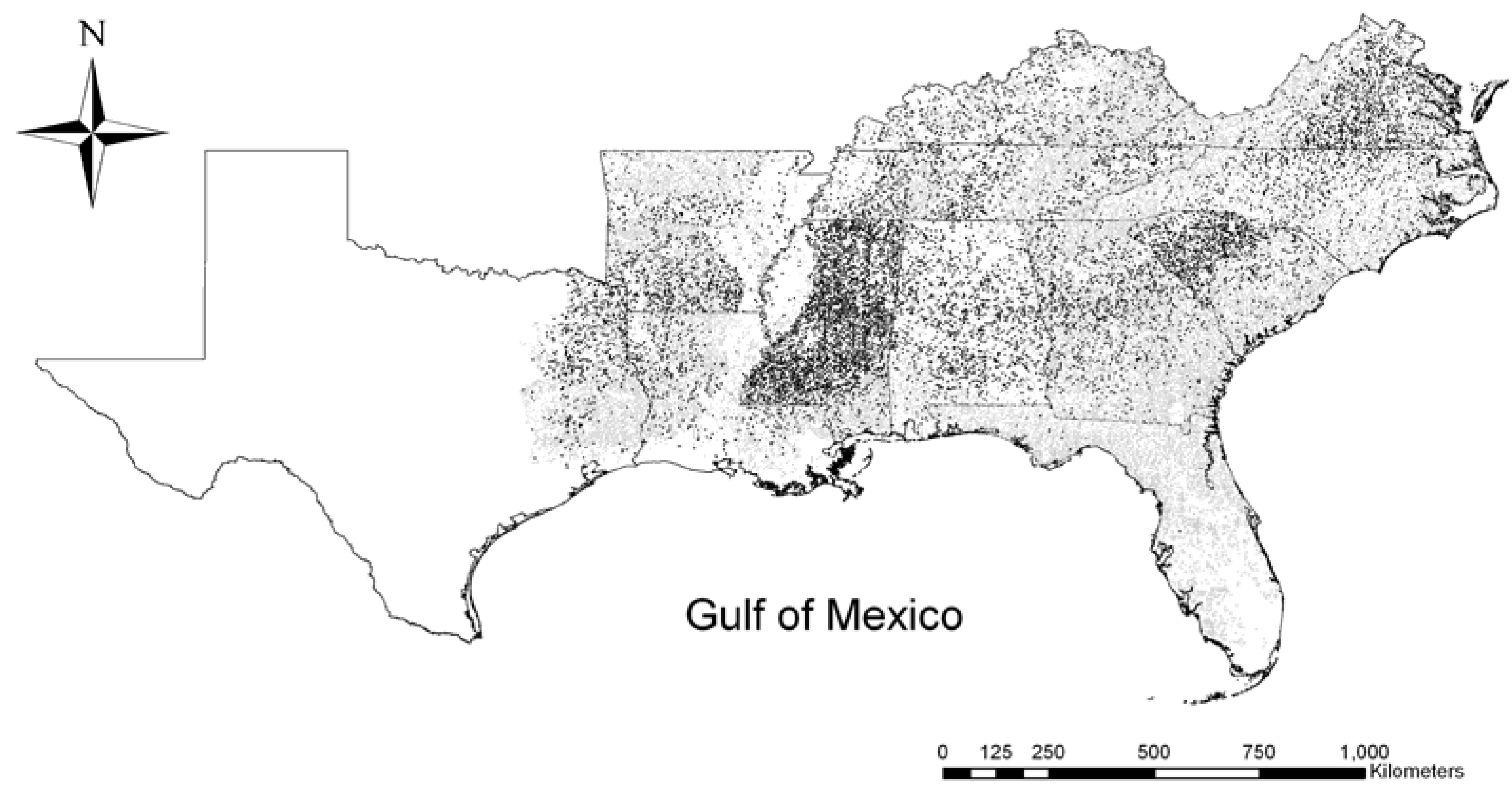
2.2. Potential Predictors of Invasion
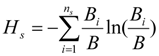

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable description | Value or unit of measure | Mean (range) or frequency |
|---|---|---|
| Landscape features | ||
| Elevation | m | 139 (−36~1524) a |
| Slope | degree | 5.39 (0~77.5) a |
| Adjacency to water bodies within 300 m | 0: no | 0: 27,940 |
| 1: yes | 1: 6,731 | |
| x | Easting of plot UTM coordinates (m) | 3.62 × 105 (2.29 × 105~5.28 × 105) a |
| y | Northing of plot UTM coordinates (m) | 3.75 × 106 (2.72 × 106~4.36 × 106) a |
| Climatic conditions | ||
| Mean daily minimum temperature | °C | 8.44 (−6.61~21.11) a |
| Mean daily maximum temperature | °C | 28.28 (21.17~29.44) a |
| Forest conditions | ||
| Stand age | years | 44.27 (1~184) a |
| Site productivity (height-age curve categories) | 1: 0–1.39 | 1: 139 |
| 2: 1.40–3.49 | 2: 5,912 | |
| 3: 3.50–5.94 | 3: 17,270 | |
| 4: 5.95–8.39 | 4: 12,112 | |
| 5: 8.40–11.54 | 5: 5,609 | |
| 6: 11.55–15.74 | 6: 1,483 | |
| 7: >15.74 m3/ha/yr | 7: 112 | |
| Species diversity | Shannon’s species diversity | 1.48 (0~3.02) a |
| Management activities and disturbances | ||
| Timber harvest b | 0: no | 0: 36,216 |
| 1: yes | 1: 6,421 | |
| Site preparation b | 0: no | 0: 41,181 |
| 1: yes | 1: 1,456 | |
| Artificial regeneration b | 0: no | 0: 31,545 |
| 1: yes | 1: 11,092 | |
| Natural regeneration b | 0: no | 0: 41,663 |
| 1: yes | 1: 974 | |
| Distance to the nearest road | 1: <30 | 1: 2,846 |
| 2: 30–91 | 2: 4,732 | |
| 3: 91–152 | 3: 4,218 | |
| 4: 152–305 | 4: 7,409 | |
| 5: 305–805 | 5: 10,424 | |
| 6: 805–1,609 | 6: 4,339 | |
| 7: 1,609–4,828 | 7: 1,349 | |
| 8: 4,828–8,047 | 8: 119 | |
| 9: >8047 m | 9: 84 | |
| Insect disturbance b | 0: no | 0: 42,271 |
| 1: yes | 1: 366 | |
| Disease disturbance b | 0: no | 0: 42,480 |
| 1: yes | 1: 157 | |
| Fire disturbance b | 0: no | 0: 41,551 |
| 1: yes | 1: 1,086 | |
| Animal disturbance b | 0: no | 0: 42,069 |
| 1: yes | 1: 568 | |
| Wind disturbance b (including hurricanes and tornados) | 0: no | 0: 41,269 |
| 1: yes | 1: 1,368 | |
| Ownership | ||
| Forestland ownership | 0: public | 0: 5,349 |
| 1: private | 1: 37,288 |
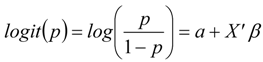
2.3. Likelihood of Further Invasion
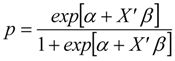
3. Results
3.1. Potential Determinants of Invasion
| Variable | Estimated coefficient | Estimated standard error | Estimated odds ratio a | 95% confidence intervals for the odds ratio | |
|---|---|---|---|---|---|
| Lower | Upper | ||||
| Landscape features | |||||
| Slope | −0.0219 | 0.0021 | 0.978 | 0.974 | 0.982 |
| Adjacency to water bodies within 300 m | 0.1604 | 0.0327 | 1.174 | 1.101 | 1.252 |
| Climatic conditions | |||||
| Mean daily maximum temperature | 0.4626 | 0.0353 | 1.588 | 1.482 | 1.702 |
| Forest conditions | |||||
| Stand age | −0.0144 | 0.0006 | 0.986 | 0.985 | 0.987 |
| Site productivity | 0.2435 | 0.0127 | 1.276 | 1.244 | 2.308 |
| Species diversity | 0.3340 | 0.0222 | 1.397 | 1.337 | 1.459 |
| Management activities and disturbances | |||||
| Artificial regeneration | −0.5511 | 0.0686 | 0.576 | 0.504 | 0.659 |
| Distance to the nearest road | −0.0653 | 0.0080 | 0.937 | 0.922 | 0.952 |
| Fire disturbance | −0.3494 | 0.0880 | 0.705 | 0.593 | 0.838 |
| Ownership | |||||
| Forestland ownership | 0.7814 | 0.0481 | 2.185 | 1.988 | 2.401 |
| Constant | −2.5040 | 0.2463 | − | − | − |
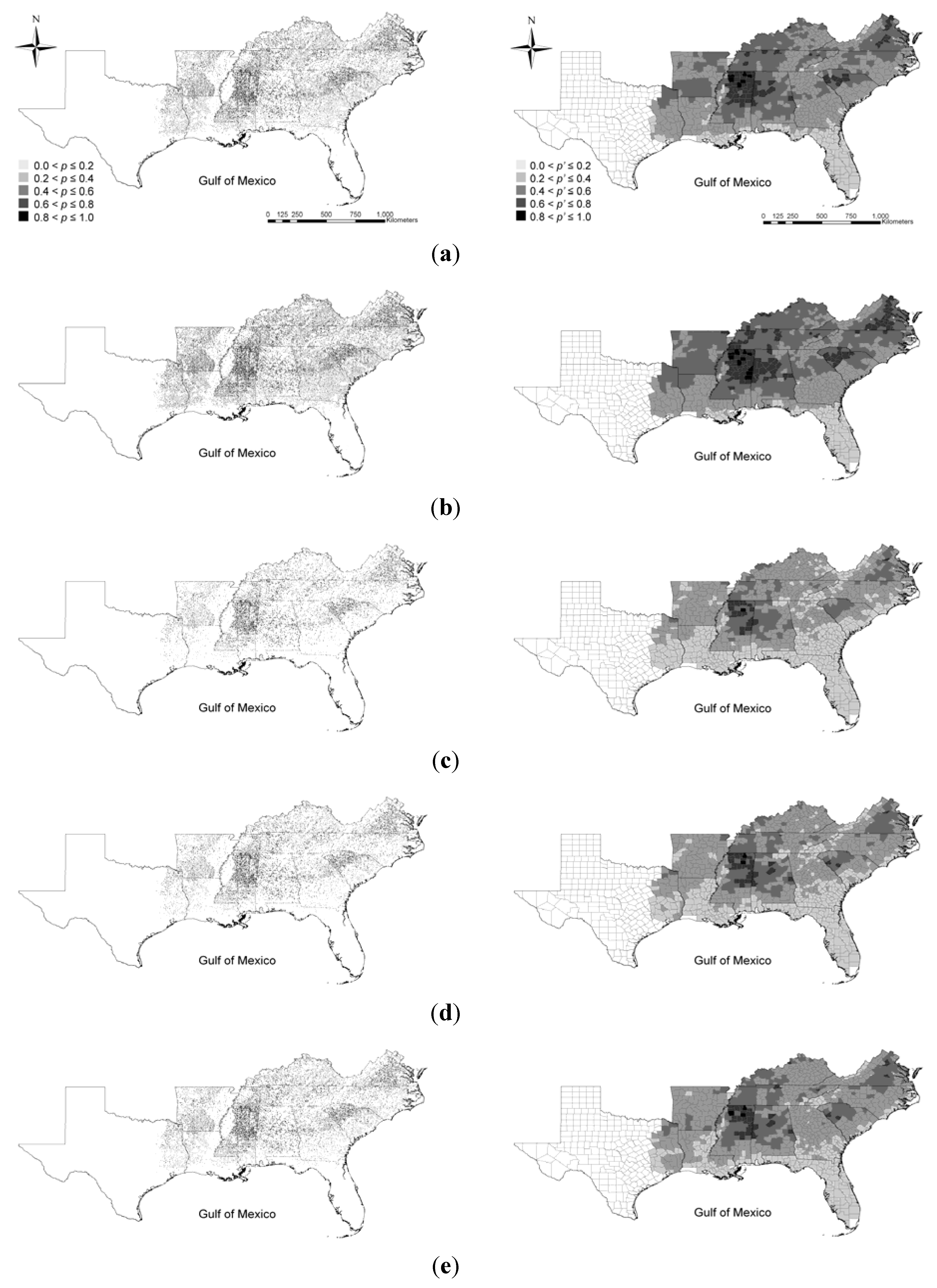
3.2. Likelihood of Further Invasion
3.2.1. Under Current Conditions
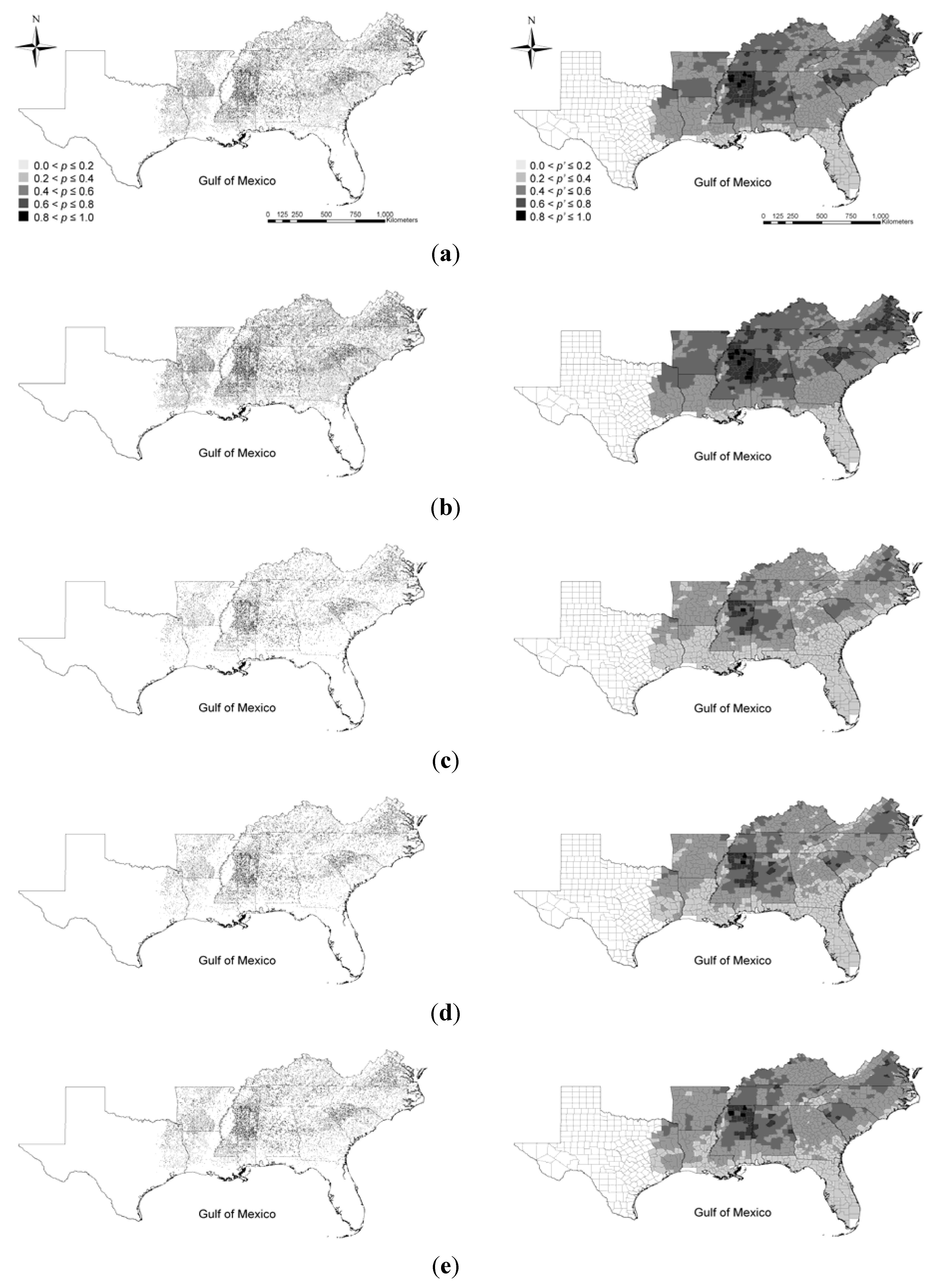
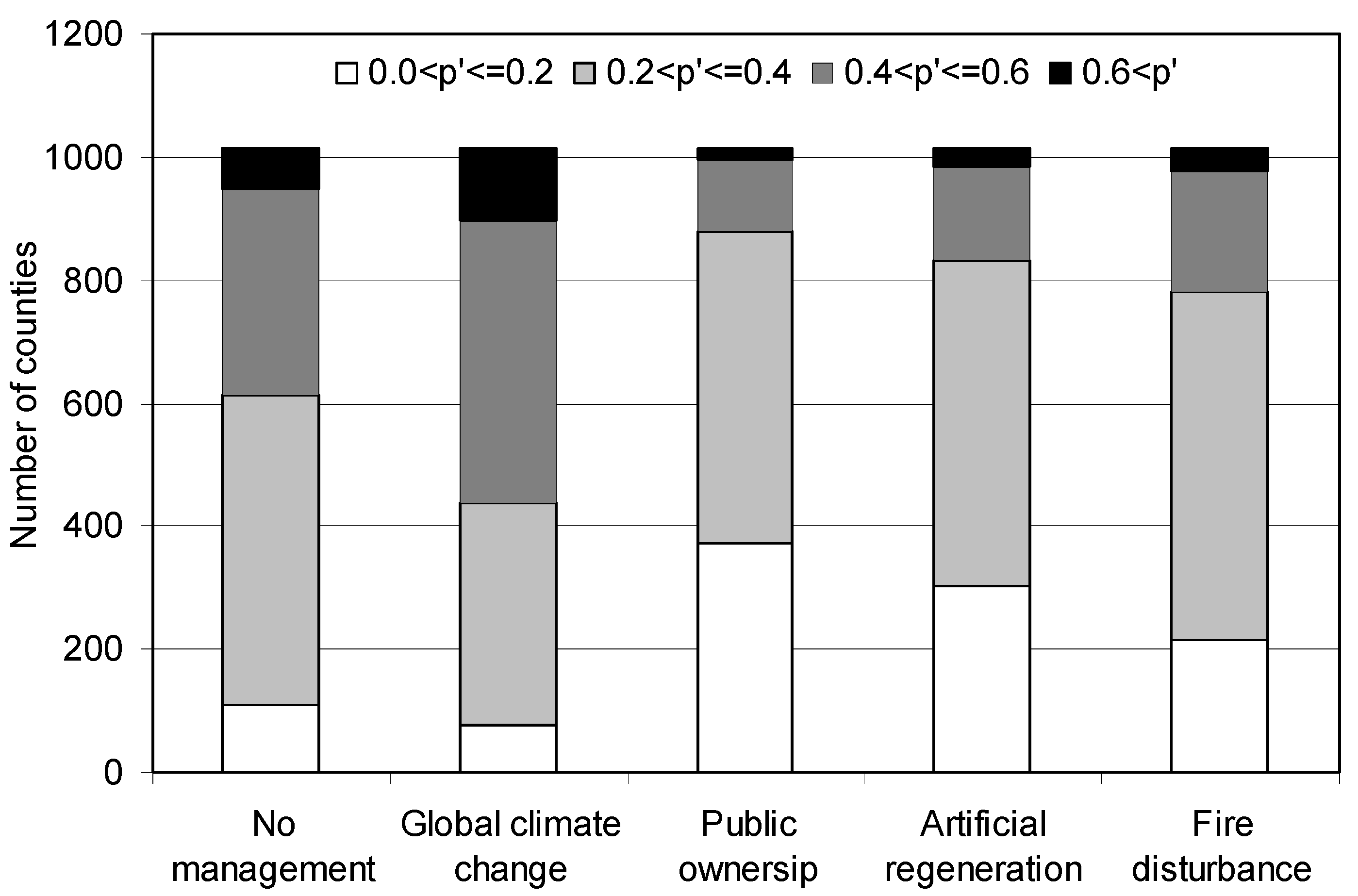
3.2.2. Under Global Climate Change and Alternative Management Strategy
4. Discussion
Conflict of Interest
References
- Mack, R.N.; Simberloff, D.; Lonsdale, W.M.; Evans, H.; Clout, M.; Bazzaz, F.A. Biotic invasions: Causes, epidemiology, global consequences, and control. Ecol. Appl. 2000, 10, 689–710. [Google Scholar]
- Rhymer, J.M.; Simberloff, D. Extinction by hybridization and introgression. Annu. Rev. Ecol. Syst. 1996, 27, 83–109. [Google Scholar]
- Vitousek, P.M.; D’Antonio, C.M.; Loope, L.L.; Westbrooks, R. Biological invasions as global environmental change. Am. Sci. 1996, 84, 468–478. [Google Scholar]
- Walker, B.H.; Steffen, W. An overview of the implications of global change for natural and managed terrestrial ecosystems. Conserv. Ecol. 1997, 1, 2. [Google Scholar]
- Lodge, D.M.; Williams, S.; MacIsaac, H.J.; Hayes, K.R.; Leung, B.; Reichard, S.; Mack, R.N.; Moyle, P.B.; Smith, M.; Andow, D.A.; et al. Biological invasions: Recommendations for U.S. policy and management. Ecol. Appl. 2006, 16, 2035–2054. [Google Scholar] [CrossRef]
- Sandlund, O.T.; Schei, P.J.; Viken, Å. Invasive Species and Biodiversity Management; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2001; p. 437. [Google Scholar]
- Pimentel, D. Environmental and economic costs of the application of pesticides primarily in the United States. Environ. Dev. Sustain. 2005, 2, 229–252. [Google Scholar] [CrossRef]
- McNeely, J.A. Human Dimensions of Invasive Alien Species. In Invasive Alien Species: A New Synthesis; Mooney, H.A., Mack, R.N., McNeely, J.A., Neville, L.E., Schei, P.J., Waage, J.K., Eds.; Island Press: Washington, DC, USA, 2005; pp. 285–309. [Google Scholar]
- Henderson, L.; Wells, M.J. Alien Plant Invasions in the Grassland and Savanna Biomes. In The Ecology and Management of Biological Invasions in Southern Africa; Macdonald, I.A.W., Kruger, F.J., Ferrar, A.A., Eds.; Oxford Univeristy Press: Capetown, South Africa, 1986; pp. 109–117. [Google Scholar]
- Di Castri, F. History of Biological Invasions with Special Emphasis on the Old World. In Biological Invasions: A Global Perspective; Drake, J., di Castri, F., Groves, R., Kruger, F., Mooney, H.A., Rejmanek, M., Williamson, M., Eds.; Wiley: New York, NY, USA, 1989; pp. 1–30. [Google Scholar]
- Simberloff, D. Global climate change and introduced species in United States forests. Sci. Total Environ. 2000, 262, 253–261. [Google Scholar] [CrossRef]
- McAusland, C.; Costello, C. Avoiding invasives: Trade-related policies for controlling unintentional exotic species introductions. J. Environ. Econ. Manag. 2004, 48, 954–977. [Google Scholar] [CrossRef]
- McNulty, S.G.; Moore, J.A.; Iverson, L.; Prasad, A.; Abt, R.; Smith, B.; Sun, G.; Gavazzi, M.; Bartlett, J.; Murray, B.; et al. Application of linked regional scale growth, biogeography, and economic models for southeastern united states pine forests. World Resour. Rev. 2000, 12, 298–320. [Google Scholar]
- USDA, The Forest Inventory and Analysis Database: Database Description and Users Manual Version 5.1; U.S. Department of Agriculture Forest Service: Arlington, VA, USA, 2011.
- Hayes, D. Earth day 1990: Threshold of the green decade. World Policy J. 1990, 7, 289–304. [Google Scholar]
- Skeen, J.N.; Doerr, P.D.; Lear, D.H.V.; van Lear, D.H.; Quarterman, E.; Burbanck, M.P.; Shure, D.J.; DeSelm, H.R.; Murdock, N.; Bryant, W.S.; et al. Biodiversity of the Southeastern United States: Upland Terrestrial Communities; John Wiley & Sons: New York, NY, USA, 1993; p. 400. [Google Scholar]
- Miller, J.H. Pine Plantation Communities: How Do We Begin to Manage for Plant Diversity. In New Century: New Opportunities: Proceedings, 54th Annual Southern Weed Science Society Meeting, Biloxi, MS, USA, 22–24 January 2001; Southern Weed Science Society: Champaign, IL, USA, 2011; pp. 215–219. [Google Scholar]
- Linder, E.T. Biodiversity and Southern Forests. In Southern Forest Science: Past, Present, and Future; Gen. Tech. Rep. SRS 75; U.S. Department of Agriculture, Forest Service, Southern Research Station: Asheville, NC, USA, 2004; Volume 25, pp. 303–306. [Google Scholar]
- Hardt, R.A. Japanese honeysuckle: From “one of the best” to ruthless pest. Arnoldia USA 1986, 46, 27–34. [Google Scholar]
- Schierenbeck, K.A. Japanese honeysuckle (Lonicera japonica) as an invasive species: History, ecology, and context. Crit. Rev. Plant Sci. 2004, 23, 391–400. [Google Scholar] [CrossRef]
- Liu, D.; Jiang, H.; Zhang, R.; He, K.S. Predicting the Spatial Distribution of Lonicera japonica, Based on Species Occurrence Data from Two Watersheds in Western Kentucky and Tennessee. In Proceedings of 17th Central Hardwood Forest Conference, Lexington, KY, USA, 5–7 April 2010; Fei, S., Lhotka, J.M., Stringer, J.W., Gottschalk, K.W., Miller, G.W., Eds.; U.S. Department of Agriculture, Forest Service, Northern Research Station: Lexington, KY, USA, 2011; pp. 418–424. [Google Scholar]
- Leatherman, A.D. Ecological Life-History of Lonicera japonica Thunb.
- Schierenbeck, K.A. Comparative Ecological and Genetic Studies between a Native (Lonicera sempervirens L.) and an Introduced Congener (L. japonica Thunb.).
- West, N.M.; Gibson, D.J.; Minchin, P.R. Microhabitat analysis of the invasive exotic liana Lonicera japonica Thunb. J. Torrey Bot. Soc. 2010, 137, 380–390. [Google Scholar] [CrossRef]
- Faulkner, J.L.; Clebsch, E.E.C.; Sanders, W.L. Use of prescribed burning for managing natural and historic resources in Chickamauga and Chattanooga National Military Park, USA. Environ. Manag. 1989, 13, 603–612. [Google Scholar] [CrossRef]
- Cain, M.D.; Wigley, T.B.; Reed, D.J. Prescribed fire effects on structure in uneven-aged stands of loblolly and shortleaf pines. Wildl. Soc. Bull. 1998, 26, 209–218. [Google Scholar]
- Casper, B.B.; Jackson, R.B. Plant competition underground. Annu. Rev. Ecol. Syst. 1997, 28, 545–570. [Google Scholar]
- Oosting, H.J.; Livingston, R.B. A resurvey of a loblolly pine community twenty-nine years after ground and crown fire. Bull. Torrey Bot. Club 1964, 91, 387–395. [Google Scholar] [CrossRef]
- Bell, D.J.; Forseth, I.N.; Teramura, A.H. Field water relations of three temperate vines. Oecologia 1988, 74, 537–545. [Google Scholar] [CrossRef]
- Yurkonis, K.A.; Meiners, S.J. Invasion impacts local species turnover in a successional system. Ecol. Lett. 2004, 7, 764–769. [Google Scholar]
- Nyboer, R. Vegetation management guideline: Japanese honeysuckle (Lonicera japonica Thunb.). Nat. Areas J. 1992, 12, 217–218. [Google Scholar]
- Thomas, L.K. The impact of three exotic plant species on a Potomac Island. Natl. Park Serv. Sci. Monogr. Ser. 1980, 13, 1–179. [Google Scholar]
- Myster, R.W.; Pickett, S.T.A. Dynamics of associations between plants in ten old fields during 31 years of succession. J. Ecol. 1992, 80, 291–302. [Google Scholar] [CrossRef]
- Dillenburg, L.R.; Whigham, D.F.; Teramura, A.H.; Forseth, I.N. Effects of below- and aboveground competition from the vines Lonicera japonica and Parthenocissus quinquefolia on the growth of the tree host Liquidambar styraciflua. Oecologia 1993, 93, 48–54. [Google Scholar]
- Thuiller, W.; Albert, C.; Araújo, M.B.; Berry, P.M.; Cabeza, M.; Guisan, A.; Hickler, T.; Midgley, G.F.; Paterson, J.; Schurr, F.M.; et al. Predicting global change impacts on plant species’ distributions: Future challenges. Perspect. Plant Ecol. Evol. Syst. 2008, 9, 137–152. [Google Scholar] [CrossRef]
- Bradley, B.A.; Blumenthal, D.M.; Wilcove, D.S.; Ziska, L.H. Predicting plant invasions in an era of global change. Trends Ecol. Evol. 2010, 25, 310–318. [Google Scholar] [CrossRef]
- Bradley, B.A. Assessing ecosystem threats from global and regional change: Hierarchical modeling of risk to sagebrush ecosystems from climate change, land use and invasive species in Nevada, USA. Ecography 2010, 33, 198–208. [Google Scholar]
- Westerling, A.L.; Hidalgo, H.G.; Cayan, D.R.; Swetnam, T.W. Warming and earlier spring increase western U.S. forest wildfire activity. Science 2006, 313, 940–943. [Google Scholar]
- Hansen, M.; Clevenger, A. The influence of disturbance and habitat on the presence of non-native plant species along transport corridors. Biol. Conserv. 2005, 125, 249–259. [Google Scholar] [CrossRef]
- USDA Agricultural productivity in the United States. Available online: http://www.ers.usda.gov/Data/AgProductivity (accessed on 13 January 2011).
- Rudis, V.A.; Gray, A.; McWilliams, W.; O’Brien, R.; Olson, C.; Oswalt, S.; Schulz, B. Regional Monitoring of Nonnative Plant Invasions with the Forest Inventory and Analysis Program. In Proceedings of the Sixth Annual FIA Symposium; Gen. Tech. Rep. WO-70, Denver, CO, USA, 21–24 September 2006; McRoberts, R.E., Reams, G.A., Deusen, P.C.V., McWilliams, W.H., Eds.; USDA Forest Service: Washington, DC, USA, 2006; pp. 49–64. [Google Scholar]
- USDA (United States Department of Agriculture). FIA data and tools. Available online: http://fia.fs.fed.us/tools-data (accessed on 4 November 2010).
- USDA (United States Department of Agriculture). Nonnative invasive plant data tool. Available online: http://srsfia2.fs.fed.us/data_center/index.shtml (accessed on 10 August 2011).
- NOAA (United States Department of Agriculture). Climate maps of the United States. Temperature maps. Available online: http://cdo.ncdc.noaa.gov/cgi-bin/climaps/climaps.pl (accessed on 15 June 2010).
- Spittlehouse, D.L.; Stathers, R.J. Seedling Microclimate; Land Management Report; Ministry of Forests: Victoria, BC, Canada, 1990; pp. 28–36. [Google Scholar]
- Jones, R.H.; McLeod, K.W. Shade tolerance in seedlings of Chinese tallow tree, American sycamore, and cherrybark oak. Bull. Torrey Bot. Club 1989, 116, 371–377. [Google Scholar]
- Burns, J.H.; Miller, T.E. Invasion of Chinese tallow (Sapium sebiferum) in the Lake Jackson area, northern Florida. Am. Midl. Nat. 2004, 152, 410–417. [Google Scholar]
- Pattison, R.R.; Mack, R.N. Potential distribution of the invasive tree Triadica sebifera (Euphorbiaceae) in the United States: Evaluating CLIMEX predictions with field trials. Glob. Chang. Biol. 2008, 14, 813–826. [Google Scholar] [CrossRef]
- Wills, C.; Condit, R.; Foster, R.B.; Hubbell, S.P. Strong density- and diversity-related effects help to maintain tree species diversity in a neotropical forest. Proc. Nat. Acad. Sci. USA 1997, 94, 1252–1257. [Google Scholar]
- Filipescu, C.N.; Comeau, P.G. Competitive interactions between aspen and white spruce vary with stand age in boreal mixedwoods. For. Ecol. Manag. 2007, 247, 175–184. [Google Scholar] [CrossRef]
- Lombardo, K.; Fehmi, J.S.; Rice, K.J.; Laca, E.A. Nassella pulchra survival and water relations depend more on site productivity than on small-scale disturbance. Restor. Ecol. 2007, 15, 177–178. [Google Scholar]
- Huebner, C.D.; Tobin, P.C. Invasibility of mature and 15-year-old deciduous forests by exotic plants. Plant Ecol. 2006, 186, 57–68. [Google Scholar] [CrossRef]
- Pyšek, P.; Jarošík, V.; Kučera, T. Patterns of invasion in temperate nature reserves. Biol. Conserv. 2002, 104, 13–24. [Google Scholar]
- Overmars, K.P.; de Koning, G.H.J.; Veldkamp, A. Spatial autocorrelation in multi-scale land use models. Ecol. Model. 2003, 164, 257–270. [Google Scholar]
- Gan, J.; Miller, J.H.; Wang, H.-H.; Taylor, J.W. Invasion of tallow tree into southern U.S. forests: Influencing factors and implications for mitigation. Can. J. For. Res. 2009, 39, 1346–1356. [Google Scholar]
- Martin, T.G.; Wintle, B.A.; Rhodes, J.R.; Kuhnert, P.M.; Field, S.A.; Low-Choy, S.J.; Tyre, A.J.; Possingham, H.P. Zero tolerance ecology: Improving ecological inference by modelling the source of zero observations. Ecol. Lett. 2005, 8, 1235–1246. [Google Scholar]
- Liang, J.; Buongiorno, J.; Monserud, R.A.; Kruger, E.L.; Zhou, M. Effects of diversity of tree species and size on forest basal area growth, recruitment, and mortality. For. Ecol. Manag. 2007, 243, 116–127. [Google Scholar]
- Agresti, A. An Introduction to Categorical Data Analysis, 2nd ed; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2007; p. 372. [Google Scholar]
- Akaike, H. Information Theory and an Extension of the Maximum Likelihood Principle. In Second International Symposium on Information Theory; Kotz, S., Johnson, N.L., Eds.; Academia Kiado: Budapest, Hungary, 1973; pp. 267–281. [Google Scholar]
- Hosmer, D.W.; Lemeshow, S. Applied Logistic Regression; John Wiley & Sons, Inc.: New York, NY, USA, 2000; p. 392. [Google Scholar]
- Bernstein, L.; Bosch, P.; Canziani, O.; Chen, Z.; Christ, R.; Davidson, O.; Hare, W.; Huq, S.; Karoly, D.; Kattsov, V.; et al. Climate Change 2007, the Fourth Assessment Report (AR4) of the United Nations Intergovernmental Panel on Climate Change (IPCC). In Proceedings of IPCC, UN, Valencia, Spain, 12–17 November 2007.
- Lemke, D.; Hulme, P.E.; Brown, J.A.; Tadesse, W. Distribution modelling of Japanese honeysuckle (Lonicera japonica) invasion in the Cumberland Plateau and mountain region, USA. For. Ecol. Manag. 2012, 262, 139–149. [Google Scholar]
- Beans, C.; Kilkenny, F.; Galloway, L. Climate suitability and human influences combined explain the range expansion of an invasive horticultural plant. Biol. Invasions 2012. [Google Scholar]
- Woodward, F.I. Climate and Plant Distribution; Cambridge University Press: Cambridge, UK, 1987; p. 174. [Google Scholar]
- Richardson, D.M.; Wilgen, B.W.V. Invasive alien plants in South Africa: How well do we understand the ecological impacts. S. Afr. J. Sci. 2004, 100, 45–55. [Google Scholar]
- Ibáñez, I.; Silander, J.A., Jr.; Wilson, A.M.; LaFleur, N.; Tanaka, N.; Tsuyama, I. Multivariate forecasts of potential distributions of incasive plant species. Ecol. Appl. 2009, 19, 359–375. [Google Scholar]
- Daubenmire, R.F. Plants and Environment: A Textbook of Plant Autecology; John Wiley & Sons: New York, NY, USA, 1974; p. 432. [Google Scholar]
- Cain, M.D. Japanese honeysuckle in uneven-aged pine stands: Problems with natural pine regeneration. Proc. South. Weed Sci. Soc. 1992, 45, 264–269. [Google Scholar]
- Smith, D.M.; Larson, B.C.; Kelty, M.J.; Ashton, P.M.S. The Practice of Silviculture: Applied Forest Ecology; John Wiley & Sons, Inc: New York, NY, USA, 1997; p. 560. [Google Scholar]
- Wang, Z.; Nyland, R.D. Changes in the condition and species composition of developing even-aged northern hardwood stands in central New York. North. J. Appl. For. 1996, 13, 189–194. [Google Scholar]
- Surrette, S.B.; Brewer, J.S. Inferring relationships between native plant diversity and Lonicera japonica in upland forests in north Mississippi, USA. Appl. Veg. Sci. 2008, 11, 205–214. [Google Scholar]
- Weber, J.S.; Gibson, K.D. Exotic plant species in old-growth forest in Indiana. Weed Sci. 2007, 55, 299–304. [Google Scholar]
- Barden, L.S.; Matthews, J.F. Change in abundance of honeysuckle (Lonicera japonica) and other ground flora after prescribed burning of a Piedmont pine forest. Castanea 1980, 45, 257–260. [Google Scholar]
- Cacek, T. The National Park Service Integrated Pest Management Manual. Identification, Biology, Management; National Park Services, U.S. Department of the Interior: Washington, DC, USA, 1998. Available online: http://www.nature.nps.gov/biology/ipm/manual/fleas.cfm (accessed on 1 May 2012).
- Kush, J.S.; Meldahl, R.S.; Boyer, W.D. Understory Plant Community Response to Season of Burn in Natural Longleaf Pine Forests. In Proceedings of Tall Timbers Fire Ecology Conference, Boston, MA, USA, 5–9 November 2000; Moser, W.K., Moser, C.E., Eds.; Tall Timbers Research Station: Tallahassee, FL, USA, 2000; pp. 32–39. [Google Scholar]
- Brewer, J.S.; Menzel, T. A method for evaluating outcomes of restoration when no reference sites exist. Restor. Ecol. 2009, 17, 4–11. [Google Scholar] [CrossRef]
- Moreno, J.M.; Oechel, W.C. Demography of Adenostoma fasciculatum after fires of different intensities in southern California chaparral. Oecologia 1993, 96, 95–101. [Google Scholar] [CrossRef]
- Higgins, S.I.; Lavorel, S.; Revilla, E. Estimating plant migration rates under habitat loss and fragmentation. Oikos 2003, 101, 354–366. [Google Scholar] [CrossRef]
- Holle, B.V.; Simberloff, D. Ecological resistance to biological invasion overwhelmed by propagule pressure. Ecology 2005, 86, 3212–3218. [Google Scholar]
- Kuhman, T.; Pearson, S.; Turner, M. Effects of land-use history and the contemporary landscape on non-native plant invasion at local and regional scales in the forest-dominated southern Appalachians. Landsc. Ecol. 2010, 25, 1433–1445. [Google Scholar] [CrossRef]
- Roberts, H.A. Periodicity of seedling emergence and seed survival in some Umbelliferae. J. Appl. Ecol. 1979, 16, 195–201. [Google Scholar]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Kühn, I.; Wild, J.; Arianoutsou, M.; Bacher, S.; Chiron, F.; Didžiulis, V.; Essl, F.; et al. Disentangling the role of environmental and human pressures on biological invasions across Europe. Proc. Natl. Acad. Sci. USA 2010, 107, 12157–12162. [Google Scholar]
- Baars, R.; Kelly, D.; Sparrow, A.D. Liane distribution within native forest remnants in two regions of the South Island, New Zealand. New Zeal. J. Ecol. 1998, 22, 71–85. [Google Scholar]
- Dukes, J.S.; Mooney, H.A. Does global change increase the success of biological invaders? Trends Ecol. Evol. 1999, 14, 135–139. [Google Scholar] [CrossRef]
- Richardson, D.M.; Iponga, D.M.; Roura-Pascual, N.; Krug, R.M.; Milton, S.J.; Hughes, G.O.; Thuiller, W. Accommodating scenarios of climate change and management in modelling the distribution of the invasive tree Schinus molle in South Africa. Ecography 2010, 33, 1049–1061. [Google Scholar]
- Walther, G.-R.; Roques, A.; Hulme, P.E.; Sykes, M.T.; Pyšek, P.; Kühn, I.; Zobel, M.; Bacher, S.; Botta-Dukát, Z.; Bugmann, H.; et al. Alien species in a warmer world: Risks and opportunities. Trends Ecol. Evol. 2009, 24, 686–693. [Google Scholar] [CrossRef]
- Peterson, A.T. Predicting the geography of species' invasions via ecological niche modeling. Q. Rev. Biol. 2003, 78, 419–433. [Google Scholar] [CrossRef]
- Ebeling, S.K.; Welk, E.; Auge, H.; Bruelheide, H. Predicting the spread of an invasive plant: Combining experiments and ecological niche model. Ecography 2008, 31, 709–719. [Google Scholar] [CrossRef]
- Soberón, J. Grinnellian and Eltonian niches and geographic distributions of species. Ecol. Lett. 2007, 10, 1115–1123. [Google Scholar]
- Rose, N.-A.; Burton, P.J. Using bioclimatic envelopes to identify temporal corridors in support of conservation planning in a changing climate. For. Ecol. Manag. 2009, 258, S64–S74. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, H.-H.; Wonkka, C.L.; Grant, W.E.; Rogers, W.E. Potential Range Expansion of Japanese Honeysuckle (Lonicera japonica Thunb.) in Southern U.S. Forestlands. Forests 2012, 3, 573-590. https://doi.org/10.3390/f3030573
Wang H-H, Wonkka CL, Grant WE, Rogers WE. Potential Range Expansion of Japanese Honeysuckle (Lonicera japonica Thunb.) in Southern U.S. Forestlands. Forests. 2012; 3(3):573-590. https://doi.org/10.3390/f3030573
Chicago/Turabian StyleWang, Hsiao-Hsuan, Carissa L. Wonkka, William E. Grant, and William E. Rogers. 2012. "Potential Range Expansion of Japanese Honeysuckle (Lonicera japonica Thunb.) in Southern U.S. Forestlands" Forests 3, no. 3: 573-590. https://doi.org/10.3390/f3030573