1. Introduction
To date, poplar plantations in Sweden have been managed with the aim of promoting the establishment of seedlings (cuttings), protecting them against damage caused by wild animals (moose, deer and roe deer) and ensuring that stands reach an age that makes them suitable for thinning and final cutting. The total area planted with poplar in Sweden is small (<1000 ha), and Swedish demand for poplar products (timber and pulp wood) is uncertain. At present, most harvested poplars are sold to thermal power stations as biomass, with a smaller quantity going to pulp mills as pulp wood. Most current poplar stands are around 15–25 years old and their owners are interested in harvesting them.
The most common methods for establishing new poplar stands under conditions such as those found in Sweden are:
Poplar reproduces vegetatively by sprouting from its stumps and also by growing suckers from its roots. Black cottonwood (
Populus trichocarpa Torr. and Gray) commonly produces sprouts while balsam poplar produces both sprouts and suckers [
2]. Suckers are less common on gravel soil types than on silty soils. Sprout biomass production is dependent on a range of factors, including soil conditions, the overall growth rate, and the number of sprouts per stump. In a study on a 6-year-old coppice that included 17 clones, the observed rates of biomass production ranged from 2 to 11 tons ha
−1 year
−1d. w. [
3]. However clones with high biomass production could be sensitive to different abiotic factors. It was therefore proposed that a mixture of clones should be planted to minimize the risk of generating a stand with a low sprouting capacity. When growing sprouts for pulp wood or timber production, it is necessary to ensure that the number of sprouts per stump does not exceed one or two; if this is not done, competition among the sprouts will reduce stem dimensions and the harvested trees will be too small for pulp wood or timber production [
4]. According to Stanturf
et al. [
4], the level of management needed to accomplish this is not cost effective for small landowners because the stand must be entered twice.
Previous studies on poplar sprouting have mostly focused on young poplars (2–10 years old). The rotation period might contain two or three coppice during 3–4 years each. However, despite demand from Swedish forest owners, there is little information on the management of poplar sprout growth for biomass production when working with older (15–20 years) trees that have recently been harvested. Coppicing is not widely used in Sweden for tree species other than Salix grown using short rotations, and there is little collective practical experience with or knowledge of sprout management.
Many factors influence a stump’s ability to sprout. According to Lust and Mohammady [
5], these include the tree species in question, the dimensions (and thus age) of the stump, the season in which the tree was cut, and the site conditions. The most important factors in terms of coppice regeneration are the age of the harvested stand and the time of year in which they were harvested. According to Stanturf
et al. [
4], trees should be harvested before they are 10 years old in order to maximize the extent of sprouting. In most studies on the relationship between felling time and sprouting ability, it has been found that the stumps of trees felled during the growing season produce the shortest and fewest sprouts [
6]. This may be because the levels of starch and soluble carbohydrates in the roots are low during the growing season [
7]. Harvesting should therefore be conducted in the winter or spring if the aim is to maximize the rate of sprouting [
8,
9]. After harvesting, the stumps of the poplar stand may start to produce sprouts. The frequency of sprouting depends on the clone in question, the season during which harvesting was conducted, and the stump height—taller stumps (30 cm) have been reported to produce more sprouts per living stump than shorter ones (10 cm) [
9]. These findings were confirmed in a study reported by Crist
et al. [
10], who noted that 46 cm tall stumps produced more sprouts than those that were 8 or 15 cm tall.
Coppicing has been widely adopted as a management technique for biomass production because the early growth rate of sprouts is greater than that of seedlings or cuttings [
11,
12]. Sprouts develop from dormant buds, which are released following hormonal suppression when a tree is cut [
13,
14]. Sprouts emerge on the vertical side of the stump above ground level and to some extent below ground level <5 cm [
15]. However sprouts that originate too far above ground level are subject to breakage [
16]. While the literature contains few discussions of coppicing harvested poplars, Dryck and Stroble [
17] report that a paper mill company once subjected poplar stands to two coppice rotations following an initial harvest at 12 years of age.
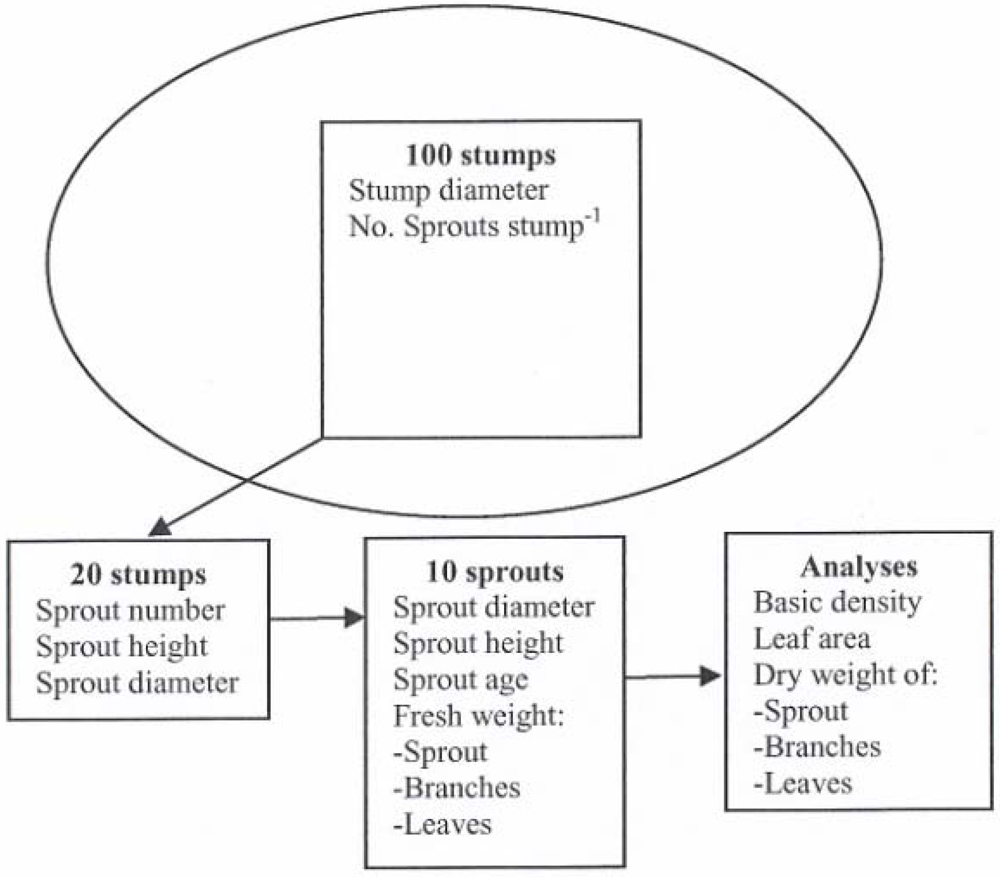
The aim of the study reported herein was to investigate the sprouting potential and biomass production of poplar stumps with one to seven year old sprouts. A biomass equation for estimating sprouts’ dry mass based on their diameter 10 cm above the ground (D10) was developed. The studied poplar stands were 8 to 21 years old. In addition to stump survival, the number of sprouts per living stump, the heights and diameters of sprouts with different ages, and the sprouts’ biomass were recorded.
4. Discussion
The first generation of poplar plantations in Sweden are about 20 years old and most of them are due to be cut in the near future. Their owners are interested in identifying options for the future management of the plantations following cutting. As such, information on the sprouting ability of poplar stumps, the number of living stumps and the dry mass production per stump and hectare are important factors in determining the owners’ optimal course of action. One option for the plantations’ future management will be to encourage stump sprouting. The sprouts can be used for biomass production or, after removing most of the sprouts from each stump, for pulp wood production. In the future, sprout biomass could represent a valuable source of bioenergy and become an economically important component of poplar forestry operations.
When analyzing the results obtained in this work, it is important to note that we examined relatively old stumps (8–21 years) whereas previously published works on poplar sprouting have focused on seedlings cut at 1–3 years of age. Poplar sprouting is generally vigorous [
27], but most of the published results in this area relate to young poplars (2–6 years). In most species, older stumps have lower numbers of dormant buds [
14,
28]. However it has also been reported that the abundance of bud clusters increases with stump age [
28]. In this work, few visible buds were observed on the poplar stumps, and the sprouts that were observed appeared to have initially developed between the cambium and the bark. Many sprouts emerged in the upper part of the stump. However, many of these sprouts were unstable and fell after 1–2 vegetation periods. Sprouts emerging from the lower part of the stump were more stable.
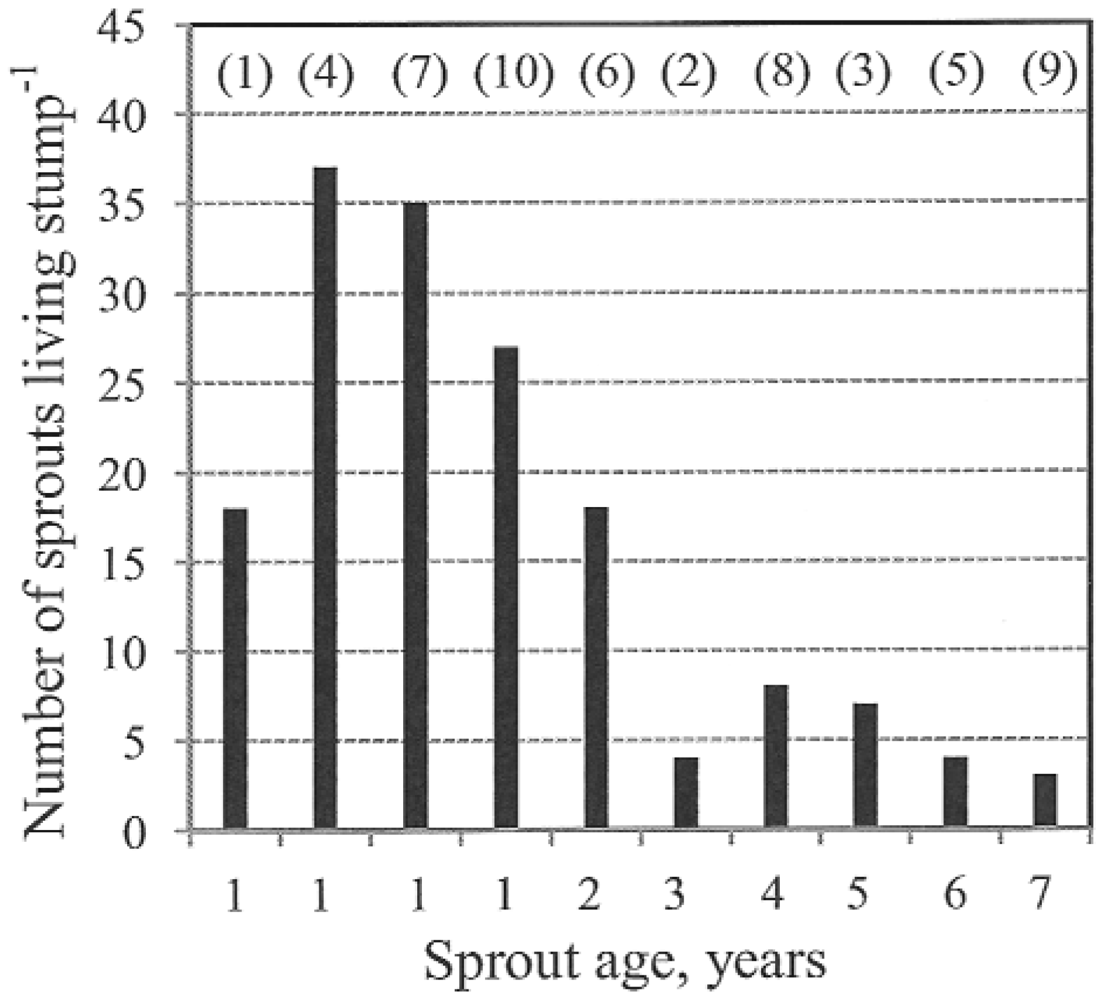
In a study of an Eastern cottonwood stand harvested monthly during a year, the harvest between January and March yielded the greatest number of sprouts [
8]. The lowest number of sprouts was found after harvest between April and August. Stumps cut during this period yielded the greatest sprout heights and diameters. In our study, the number of sprouts per living stump decreased as the length of time since cutting increased. Within one year of cutting, the mean number of sprouts per living stump was 18–37 with a range of 2 to 68. Conversely, trees that had been cut 7 years ago had only 3 (1–12) sprouts per living stump. The increasing sprout size and the larger leaf area increase the interior competition with suppressed and dying sprouts [
29]. There are indications that sprout suppression might be clone specific. In a 6-year old coppice culture of 17 different clones the authors reported that the number of sprouts per stool was highly clone specific. Within a clone the number of sprouts after cutting was positively correlated with the stool size [
29].
Table 8 presents the per-stump sprout numbers reported in previous studies on poplar coppicing. As shown by the table, most previous authors observed fewer sprouts on 1-year-old stumps than were seen in our study.
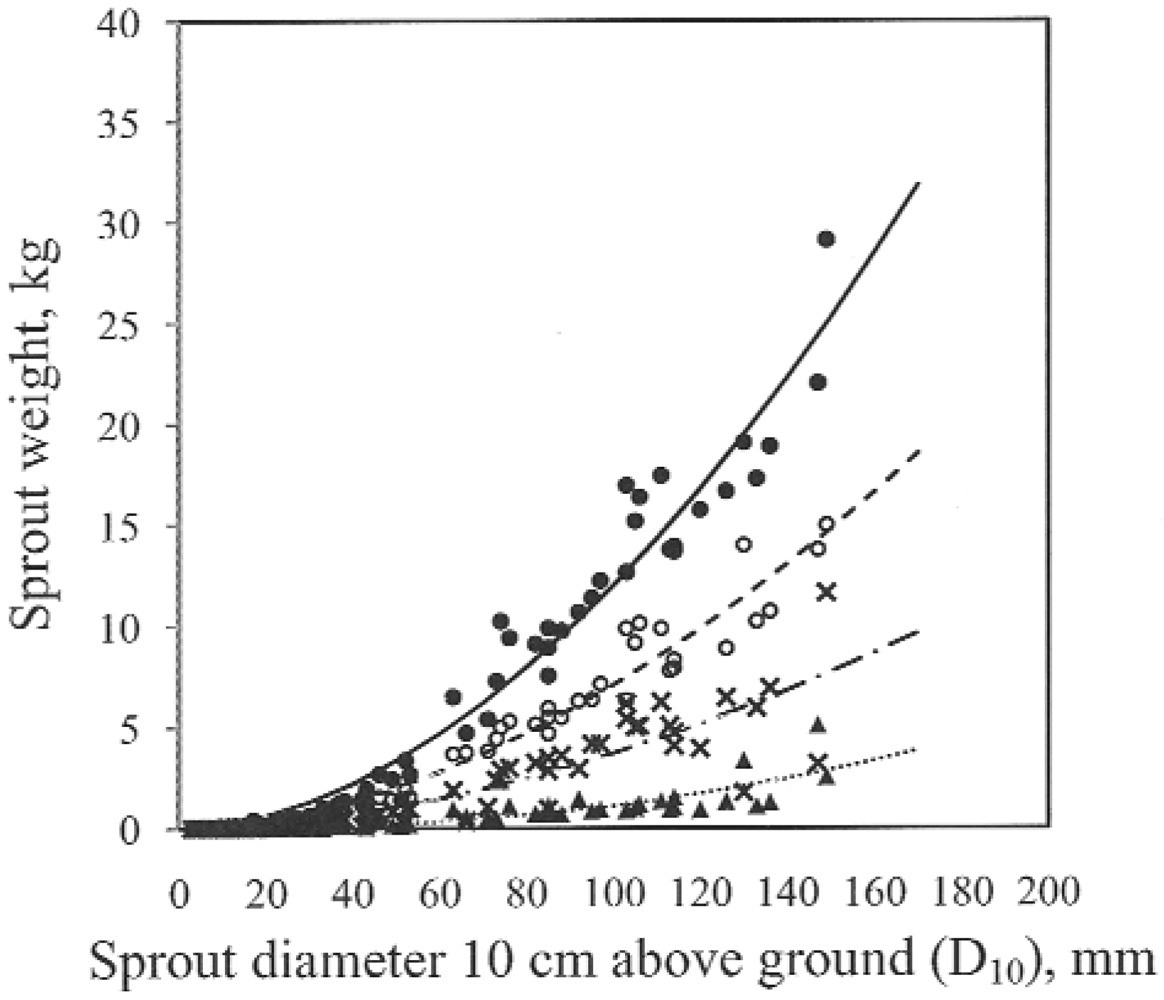
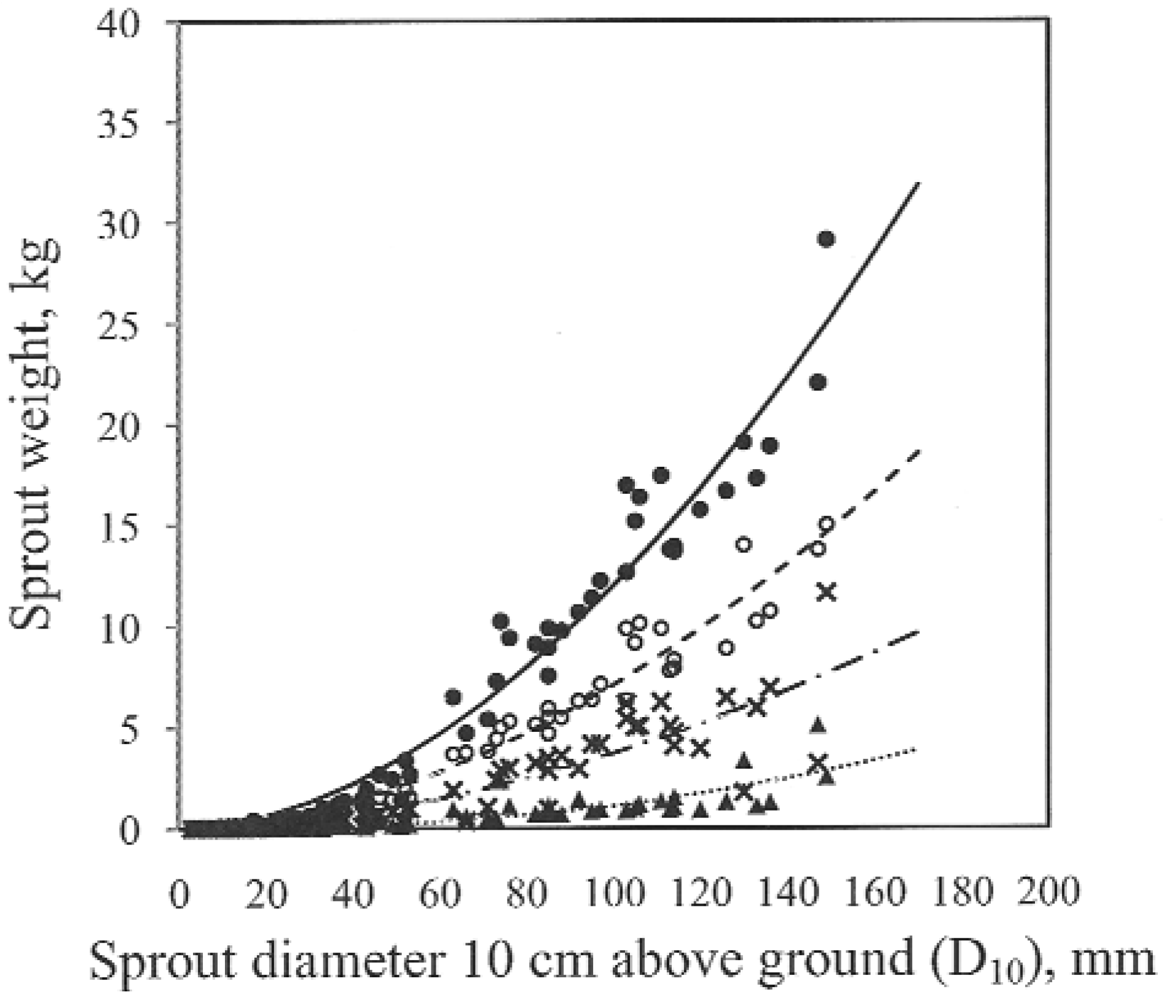
The power model fit the data well. In our study the biomass was expressed as depending on diameter of sprouts at 0.1 m above ground level. An application of biomass vs. sprout age is another way but in practice you may have to know the age of the sprouts. Diameter of the sprouts is easy to measure and in practice a convenient way to estimate the sprout biomass.
The timing of harvesting is important in determining a stump’s sprouting ability [
16]. All of the stands examined in this work had been cut between November and March. A study conducted in Wisconsin on 1-year-old sprouts of the clone NE-299 (
P. nigra var.
betulifolia ×
P. trichocarpa) indicated that stump survival rates were 65%, 9% and 5% for poplars cut in June, July and August, respectively, compared to 92% and 93% for trees harvested in September-May, respectively [
9]. The percentage of surviving stumps observed in this work ranged from 57% to 100%. The percentages for 1-year-old sprouts were 79–100% while that for the oldest 7-year-old sprouts was 70%. Few or none of the sprouts had been damaged by wild animals, pathogens or insects. As shown in
Table 8, previous studies on young poplar stumps have produced results consistent with those observed in this work. It has previously been shown that extensive coppice management (planting seedlings directly on the bare soil without herbicides or fertilizers) yielded lower stump survival rates than intensive management involving adding fertilizers and other beneficial chemicals [
37]. In a study by Laureysens
et al. [
29], the number of sprouts per living stump varied depending on the clone used, with
Populus tricoharpa and
P. trichocarpa ×
P. deltoides clones giving the greatest number of sprouts.
Many previous studies have demonstrated that when using common short rotations (≤3 years), coppiced stands are more productive than their planted counterparts [
27,
38,
39,
40]. Sprouts grow more rapidly than seedlings because they can use the stump’s pre-existing root system. They also rapidly develop a high leaf area index (LAI) [
41]. Herve and Ceulemans [
27] studied coppiced and planted stands of five different poplar clones in Belgium and France, and observed that the height and diameter of 3-year-old sprouts were 1.77–3.49 m. and 17.8–30.0 mm, respectively, whereas those of 3-year-old seedlings were 1.21–1.94 m. and 10.5–16.8 mm, respectively. After three years the volume index (d
2h) dm
3 for the coppiced stands was lower than for the planted stands [
27]. The diameters and heights of 3-, 4-, 5- and 7-year-old sprouts examined in our study were compared to the corresponding values for planted poplar seedlings of the same age. Most of the planted poplars used in this comparison were grown in Sweden and were of different clones to the stumps examined in this work; however, the 3 year old planted seedlings were of the same clone and were grown in the Czech Republic. As shown in
Table 9, the older seedlings (7 years) were taller than the sprouts.
Table 8.
Reported sprout characteristics for poplar (Populus sp.).
Table 8.
Reported sprout characteristics for poplar (Populus sp.).
| Reference | Sprout age years | No. sprouts (stump−1) | Living stumps (%) | Total production (tons ha−1) | Sprout | Remarks |
|---|
| Height (m) | Diam. (mm) |
|---|
| 30 | 3 | - | - | 0.61–4.40 | 2.8–4.1 | 20–43 1 | Populus nigra spp. nigra Czech Republic. |
| 3 | - | - | 3.18–4.23 | 3.4–4,7 | 36–45 1 | NE-42 (
P. maximowisczii × P. trichocarpa). |
| 31 | 4 | 7.5–16.8 | 50–87 | 6.6–7.9 | - | - | Populus nigra spp. nigra Czech Republic |
| 4 | 9.3 | 68 | 9.4–9.8 | - | - | NE-42 (
P. maximowisczii × P. trichocarpa). |
| 13 | 3 | 1 | 85–90 | 2.0–3.9 | 1.5 | 20 2 | “Beaupré” (
P. trichocarpa × P. deltoides France. |
| 32 | 1 | - | - | 2.9–8.3 | - | - | Belgium. |
| 33 | 4 | 8.0 | - | 26 | 6.6 | 44 3 | P. trichocarpa Mount Vernon, Virginia. |
| 8 | 1.4 | - | 52 | 10.9 | 69 3 | P. trichocarpa Mount Vernon, Virginia. |
| 29 | 1 | 3–7 | 3 | - | - | - | 17 clones. First coppicing. Belgium and France. |
| 4 | 8–19 | 7–65 | 6.4–43.2 | - | 20–68 4 | Second coppicing after 4 years growth. |
| 34 | 1 | - | - | 2.8 | - | - | Balsam spire (
P. balsamifera × P. trichocarpa ) |
| 2 | - | - | 13.9 | - | - | Scotland. |
| 3 | - | - | 33.6 | - | - | - |
| 4 | - | - | 27.1 | - | - | - |
| 6 | - | - | 55.9 | - | - | - |
| 10 | 1 | 8.3–20.1 | - | - | 1.0–1.3 | 8–10 2 | “Tristis” (
P. tristis × P. balsamifera) Wisconsin. |
| 2 | 6.0–14.9 | - | - | 1.6–2.4 | 10–17 2 | - |
| 3 | 2.2–4.2 | - | - | 3.4–4.2 | 21–28 2 | - |
| 35 | 4 | 2.6–4.4 | 22–94 | 12–30 | - | 26 2 | 17 clones. First coppicing. Belgium. |
| 9 | 1 | 3.4–7.5 | 92–93 | - | 1.5–2.3 | 5–9 2 | NE-299 (
P. nigra × P. trichocarpa) Wisconsin. |
| 27 | 3 | - | - | - | 1.8–3.5 | 18–30 2 | 5 poplar clones. Belgium. |
| 36 | 3 | 10 | - | - | 9.0 | 71 3 | Pennsylvania. |
Table 9.
Diameters (mm) and heights (m) for 4–8 year-old dominant sprouts and plants.
Table 9.
Diameters (mm) and heights (m) for 4–8 year-old dominant sprouts and plants.
| Stand age, years | Diameter DBH, mm | Height, m |
|---|
| Sprout | Plant | Sprout | Plant |
|---|
| 3 | 70 | 20–43 1,2 | 4.6 | 2.7–4.3 1,2 |
| 4 | 89 | 76 | 10.0 | 7.5 |
| 5 | 120 | 69 | 9.2 | 9.7 |
| 7 | 158 | 149 | 11.9 | 13.0–14.4 |
Dickmann
et al. [
39] studied the effects of coppicing on aboveground growth and other factors in two poplar clones—“Eugeni” (
Populus ×
euramericana (Dode) Guinier) and “Tristis I” (
Populus tristis Fisch ×
Populus balsamifera L.)—at Michigan State University’s Tree Research Center. The height of the 4-year-old poplar plants was 8–11 m. (“Eugene”) and 3–3.4 m. (“Tristis”). The trees were then cut and the sprouts’ heights were measured after one year, giving values of 2.6–3.8 m (“Eugene”) and 1.3–1.7 m (“Tristis”).
Power models are commonly used to estimate tree biomass from simple measurements of size. However the accumulation of biomass in different tree components changes over time [
42]. Al Afas
et al. [
35] have investigated whether a single biomass equation (power model) could be used for several years’ rotation and several poplar clones. They found that a single power model could indeed be used to estimate biomass production over rotation periods. The total potential for dry mass production varied between sites. Some previous studies on biomass production were conducted on experimental scales using small plots. Differences in yields between small plots and “field” stands with poorer average sites have been discussed by Hansen [
38]. Field conditions include areas with poor sites, depressions, knolls and drainages. The stands examined in this work were grown under field conditions without fertilizers, although it should be noted that they were growing on former farmland. The mean dry sprout mass production per living stump was 16.1 ± 14.0 (range: 3.3–37.2) kg; the average mass production for the ten sites examined was 17 ± 15 (1–41) tons ha
−1, with an MAI of 4.7 ± 1.4 (1.2–6.1) tons ha
−1 year
−1. As shown by
Table 8 previous authors observed yields of 3 to 50 tons ha
−1 depending on sprout age (3–8 years). Following a study conducted in the Czech Republic, Benetka
et al. [
30] reported that 29 clones of
Populus nigra L. ssp. n
igra) produced 0.61–4.40 tons ha
−1 after three years, while the control clone NE-42 (
Populus maximowiczii Henry ×
Populus trichocarpa Torr. and Gray) produced 3.18–4.23 tons ha
−1. At lower soil pH values, NE-42 had the highest production.
The stems, branches, and leaves accounted for 61 ± 8 (43–75), 8 ± 3 (2–23) and 31 ± 8 (7–53)%, respectively, of the sprouts’ overall dry mass. In a study of 17 poplar clones growing in Belgium, the corresponding proportions were found to be 71–75, 12–14 and 13–16 respectively [
41]. Proe
et al. [
34] reported that the proportion of the overall above ground dry mass that was attributable to leaves decreased year on year for the “Balsam spire” hybrid poplar (
P. balsamifera var. Michauxii (Henry) ×
P. trichocarpa var. Hastata (Dode) Farwell): at the end of the first year of coppicing, leaves accounted for 41% of the total dry mass, which fell to 25, 16, 10 and finally 9% over the next four years. Pellis
et al. [
32] reported that leaves constituted 32–44% of the total dry mass in seventeen 1-year-old poplar clones in Belgium.
Basic density (g cm−3) can be used to estimate dry weight if the volume is known. High densities indicate that would accounts for a large proportion of the tree or sprout’s mass. In our study the mean basic density of the sampled sprouts was 0.323 ± 0.012 (0.200–0.467) g cm−3. No reports dealing with basic density for poplar sprouts was found by us.
Projected Leaf Area (PLA) values are used in light interception studies and are useful indicators of biomass productivity in fast-growing trees [
43,
44]. Moreover, leaf area development and individual leaf size have been linked to productivity in
Populus. In our study the mean observed PLA was 50 (13–143) cm
2 and the mean SLA was 85 ± 25 (36–200) cm
2 g
−1 d. w. Our measurements of PLA and SLA mean values and ranges for poplar sprouts are compared with values for PLA of 47.4 ± 5.0 (range: 24–131) cm
2 and SLA of 85 (range: 36–200) cm
2 g
−1 d. w. for single stem poplars ranging from 4–73 years [
18] and for values measured in a study of seventeen different 1-year-old poplar clones aged, Pellis
et al. [
32] observed PLA values ranging from 45 to 174 cm
2 and SLA values of 160–211 cm
2 g
−1 d. w. This comparison is made as there, as we have found, no observations on leaf characteristics have been reported.

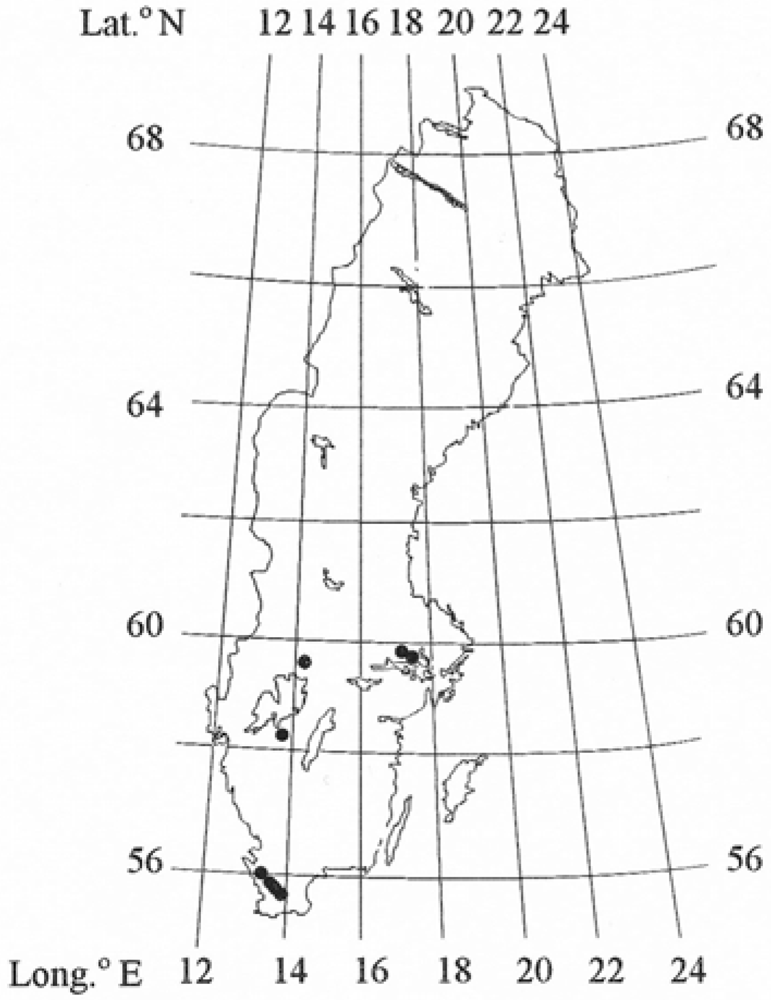

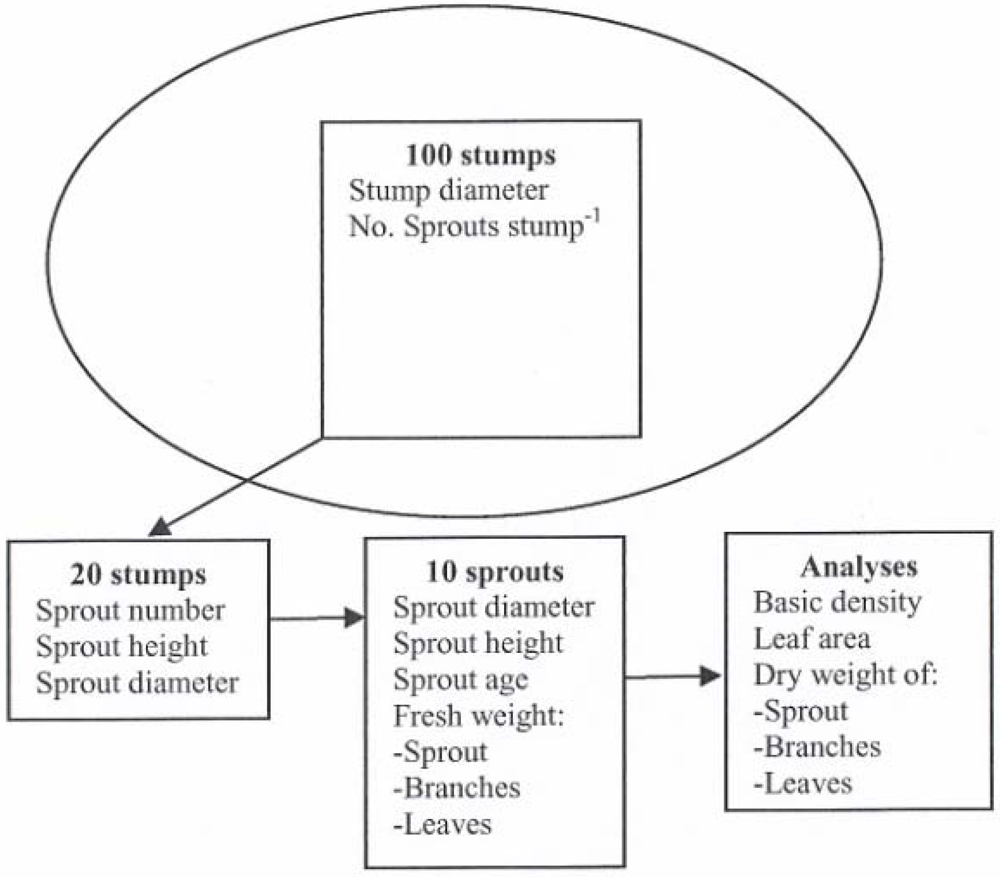
{kind=link}
{kind=link}
{kind=link}
{kind=link}
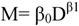
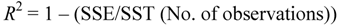
 and SST is
and SST is  .
.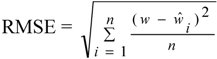
 and
and  are observed, mean and predicted weights (w).
are observed, mean and predicted weights (w).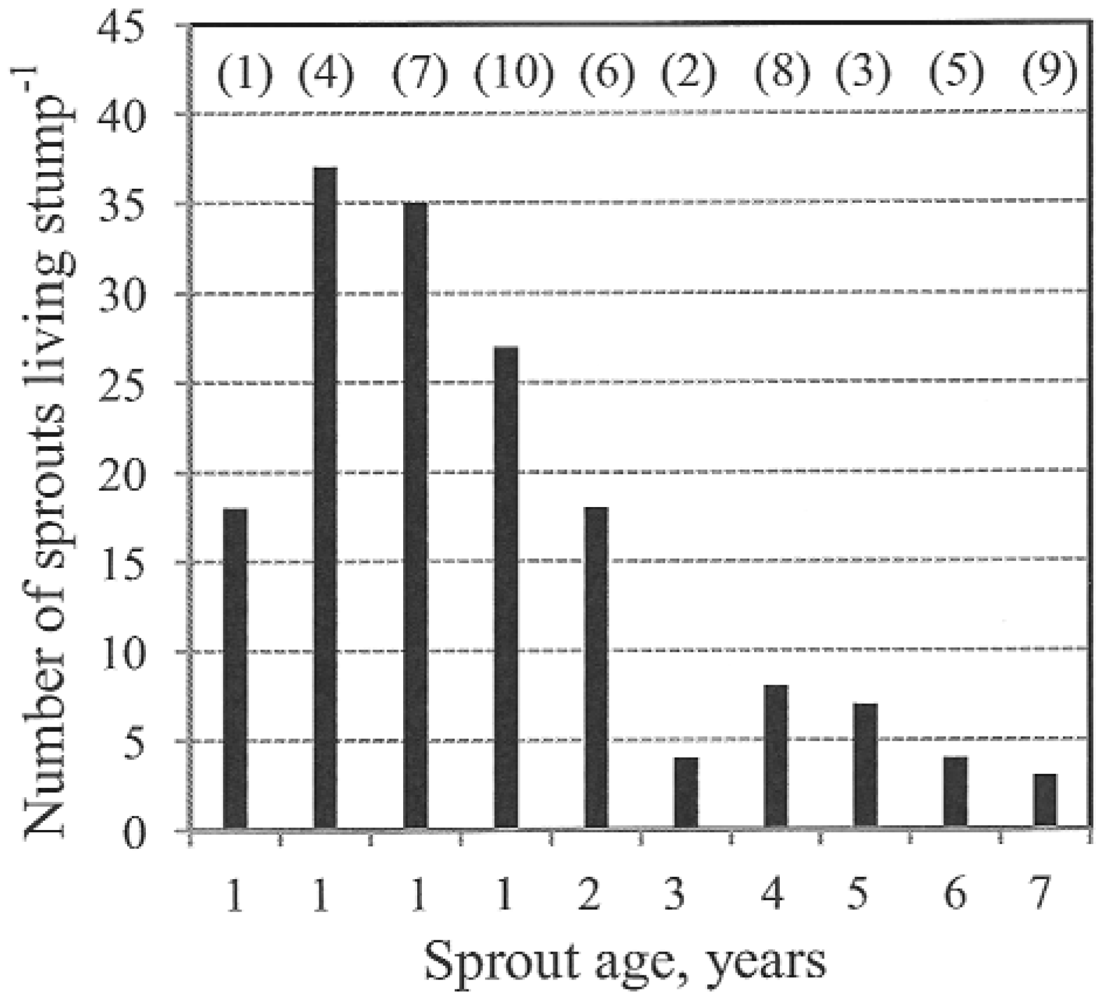
 -i008), stem (
-i008), stem (  -i009), branches (
-i009), branches (  -i010) and leaves (
-i010) and leaves (  -i011) for sampled poplar stump sprouts growing on abandoned farmland.
-i011) for sampled poplar stump sprouts growing on abandoned farmland.