Carbon Content of Tree Tissues: A Synthesis



Abstract
:1. Introduction
2. Methods
2.1. Data Collection
2.2. Data Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Biome | Location | Species ( N) |
|---|---|---|---|
| Arias et al. 2011* [45] | tropical | Costa Rica | 6 |
| Becker et al. 2012 [35] | tropical | Uganda, Kenya | 17 |
| Bert and Danjon 2006* [38] | temperate/boreal | France | 1 |
| Castaño-Santamaria and Bravo 2012* [46] | subtropical/Mediterranean | Spain | 2 |
| Correia et al. 2010* [47] | subtropical/Mediterranean | Portugal | 1 |
| Elias and Potvin 2003 [4] | tropical | Panama | 32 |
| Fang et al. 2007* [48] | temperate/boreal | China | 1 |
| Fukatsu et al. 2008 [49] | temperate/boreal | Japan | 3 |
| Herrero de Aza et al. 2011* [50] | subtropical/Mediterranean | Spain | 3 |
| Huet et al. 2004* [51] | temperate/boreal | France | 1 |
| Jacobs et al. 2009 [52] | temperate/boreal | United States | 3 |
| Janssens et al. 1999* [53] | temperate/boreal | Belgium | 1 |
| Jones and O’Hara 2012 [29] | temperate/boreal | United States | 1 |
| Joosten and Shulte 2002 [54] | temperate/boreal | Germany | 1 |
| Joosten et al. 2004 [55] | temperate/boreal | Germany | 1 |
| Kort and Turnock 1998 [56] | temperate/boreal | Canada | 4 |
| Kraenzel et al. 2003* [36] | tropical | Panama | 1 |
| Laiho and Laine 1997* [40] | temperate/boreal | Finland | 3 |
| Lamlom and Savidge 2003 [1] | temperate/boreal | Canada | 41 |
| Li et al. 2011* [57] | temperate/boreal | South Korea | 1 |
| Martin and Thomas 2011 [2] | tropical | Panama | 59 |
| Peri et al. 2010* [58] | temperate/boreal | Argentina | 1 |
| Rana et al. 2010 [59] | tropical | Philippines | 5 |
| Telmo et al. 2010 [39] | subtropical/Mediterranean | Portugal | 17 |
| Thomas and Malczewski 2007 [3] | temperate/boreal | China | 14 |
| Tolunary 2009 [60] | temperate/boreal | Turkey | 1 |
| van Geffen et al. 2010 [61] | tropical | Bolivia | 15 |
| Xing et al. 2005* | temperate/boreal | Canada | 1 |
| Zabek and Prescott 2006* [62] | temperate/boreal | Canada | 1 |
| Zhang et al. 2009* [28] | temperate/boreal | China | 10 |
| Zheng et al. 2008* [63] | subtropical/Mediterranean | China | 5 |
3. Results
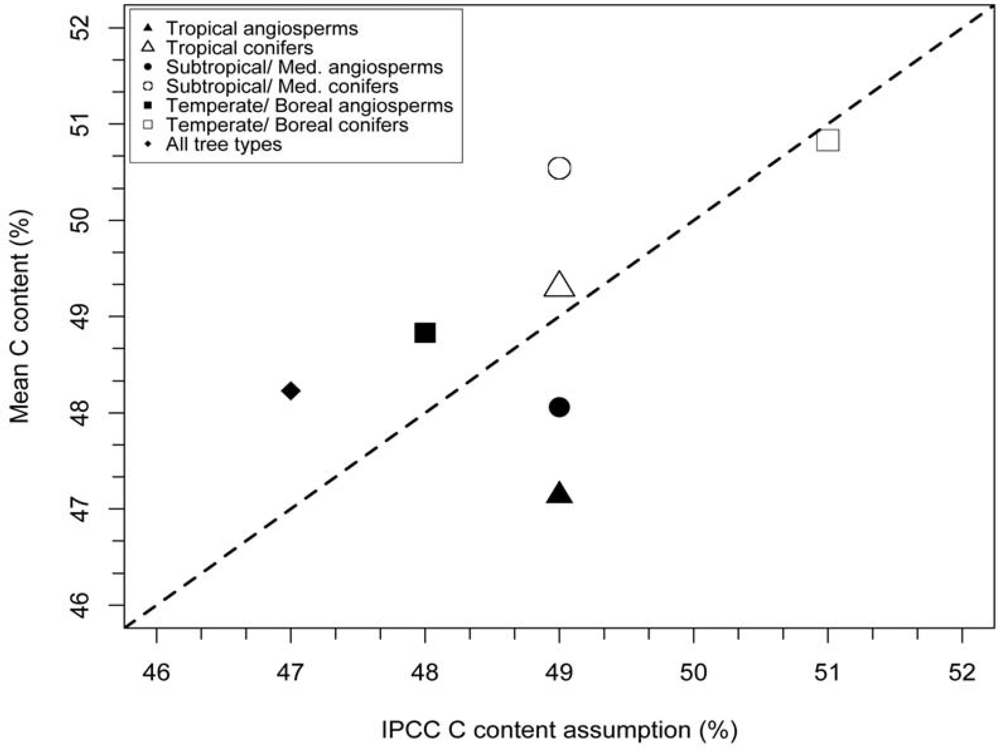
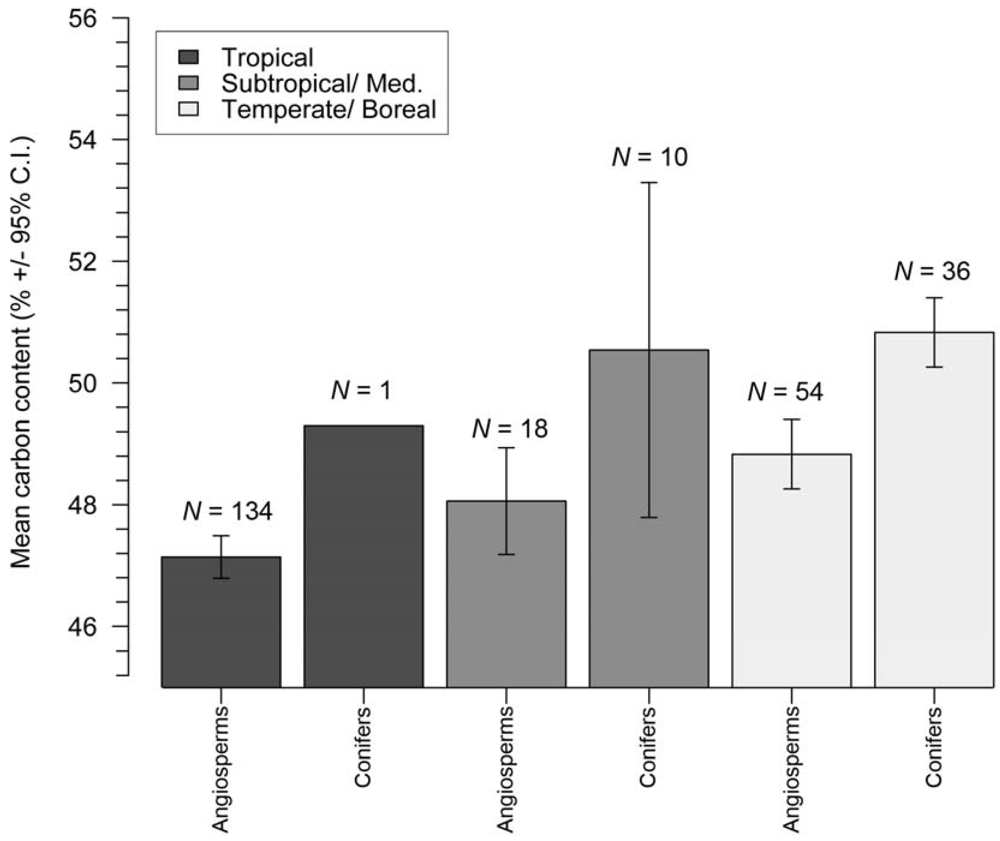
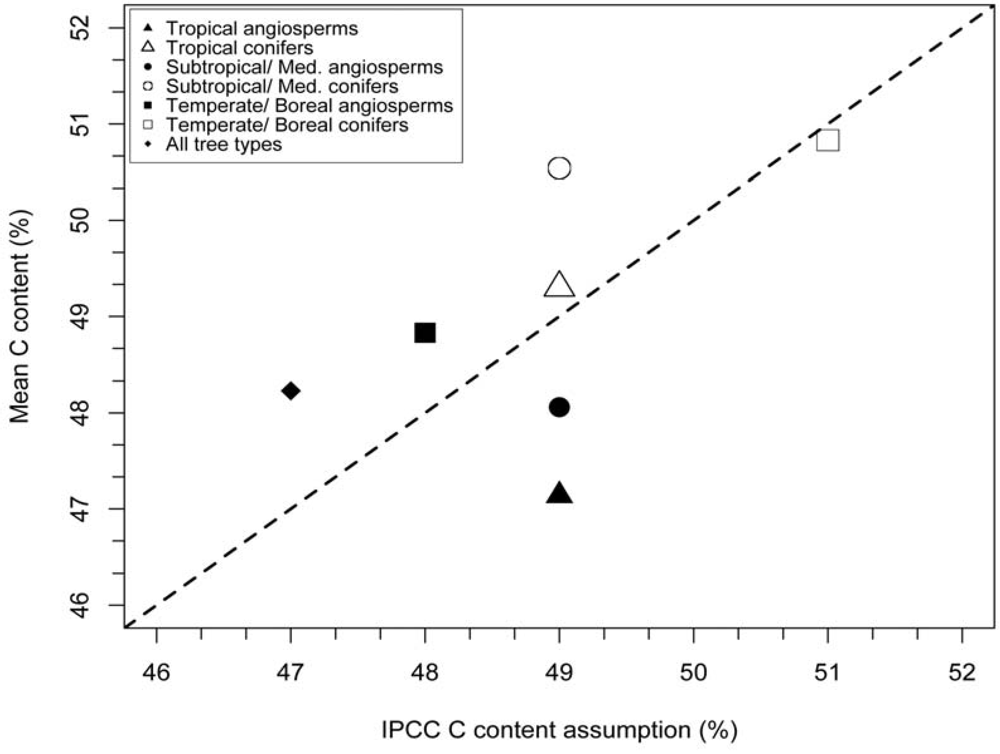
3.1. Stem Wood Carbon Content
| Biome | Type | N (References) | N (Species) | Observed mean C fraction (%) | IPCC (2006) C fraction (%) | Cvol (%) |
|---|---|---|---|---|---|---|
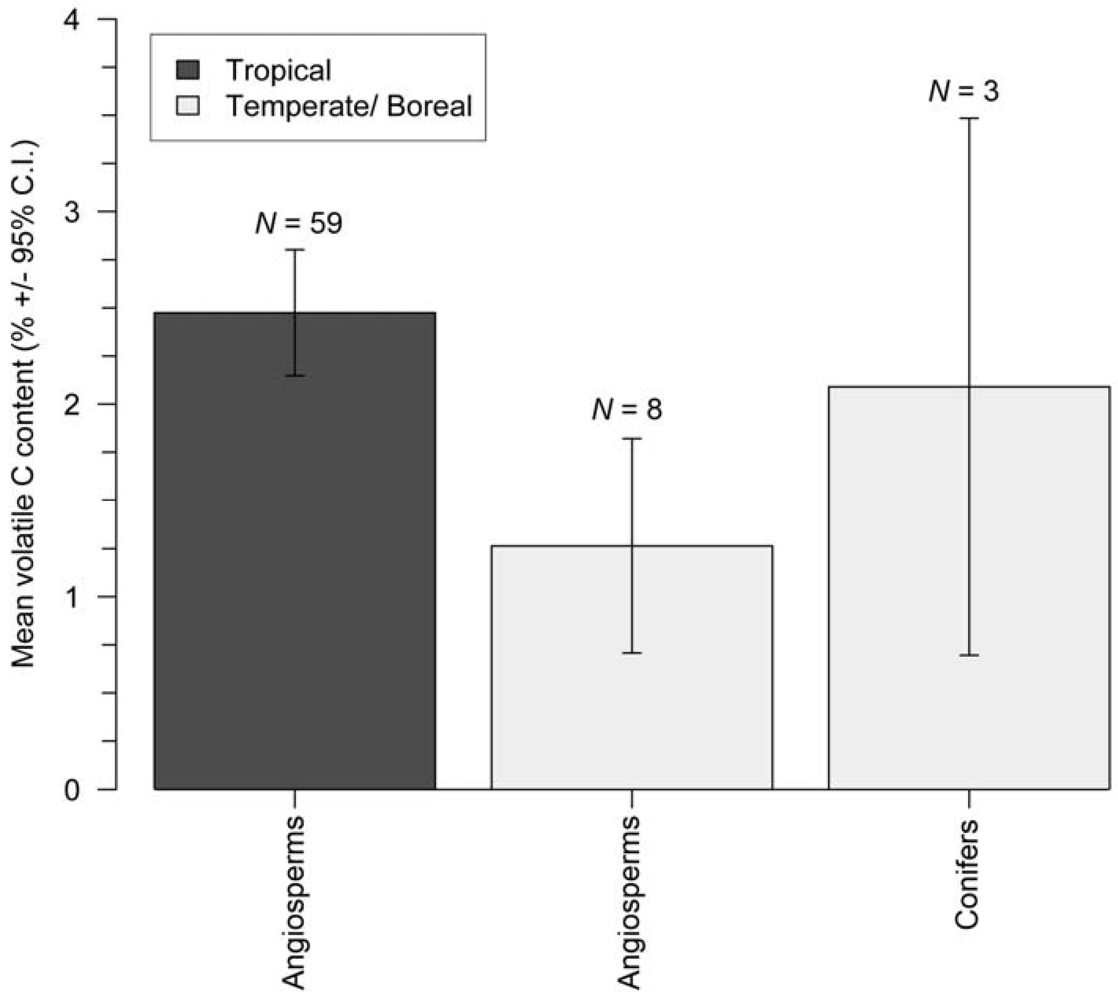
| Tropical | angiosperm | 7 | 134 | 47.1 ± 0.4 | 49 | 2.5 ± 0.3 |
| Tropical | conifer | 1 | 1 | 49.3 | 49 | N.A. |
| Subtropical/Mediterranean | angiosperm | 3 | 18 | 48.1 ± 0.9 | 49 | N.A. |
| Subtropical/Mediterranean | conifer | 3 | 10 | 50.54 ± 2.8 | 49 | N.A. |
| Temperate/Boreal | angiosperm | 10 | 54 | 48.8 ± 0.6 | 48 ± 2 | 1.3 ± 0.6 |
| Temperate/Boreal | conifer | 13 | 36 | 50.8 ± 0.6 | 51 ± 4 | 2.1 ± 1.4 |
| All biomes | angiosperm | N.A. | 206 | 47.7 ± 0.3 | N.A. | 2.3 ± 0.3 |
| All biomes | conifer | N.A. | 47 | 50.8 ± 0.8 | N.A. | 2.1 ± 1.4 |
| Complete dataset | N.A. | 31 | 253 | 48.3 ± 0.3 | 47 | 2.3 ± 0.3 |
| Term | Type III SS | D.F. | F value | p value | |
|---|---|---|---|---|---|
| Wood C content | Intercept | 57864 | 1 | 12455.66 | < 0.0001 |
| Biome | 112 | 2 | 12.02 | < 0.0001 | |
| Type | 136 | 1 | 29.21 | < 0.0001 | |
| Residuals | 1157 | 249 | - | - | |
| Cvol | Intercept | 12.78 | 1 | 8.34 | 0.005 |
| Biome | 10.33 | 1 | 6.74 | 0.012 | |
| Type | 1.49 | 1 | 0.97 | 0.328 | |
| Residuals | 102.67 | 67 | - | - | |
| Term | Estimate | Std. Error | t value | p value | |
|---|---|---|---|---|---|
| Wood C content | Intercept | 48.18 | 0.43 | 111.61 | < 0.0001 |
| Temperate | 0.59 | 0.47 | 1.26 | 0.209 | |
| Tropical | −1.04 | 0.47 | −2.21 | 0.028 | |
| Conifer | 2.16 | 0.40 | 5.41 | < 0.0001 | |
| Cvol | Intercept | 1.26 | 0.44 | 2.89 | 0.005 |
| Tropical | 1.21 | 0.47 | 2.59 | 0.012 | |
| Conifer | 0.83 | 0.84 | 0.99 | 0.328 | |
3.2. Volatile Carbon Content

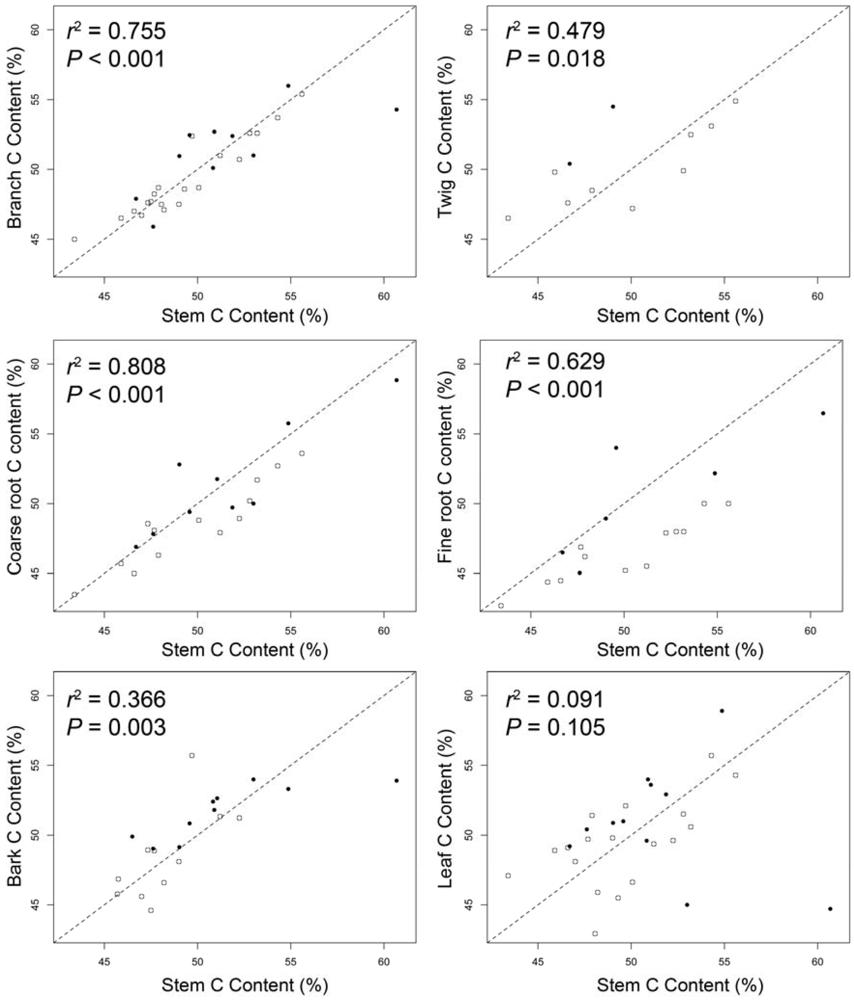
3.3. Tissue Comparisons

4. Discussion
4.1. Tissue-Specific Wood C Values
4.2. Incorporating Wood C Data into Forest C Accounting
4.3. Methodological Considerations for Wood C Determinations
 (1)
(1)  (2)
(2)  (3)
(3)  (4)
(4) 4.4. Future Research and Data Needs
4.5. Conclusions
Acknowledgments
Conflict of Interest
References
- Lamlom, S.H.; Savidge, R.A. A reassessment of carbon content in wood: Variation within and between 41 North American species. Biomass Bioenergy 2003, 25, 381–388. [Google Scholar] [CrossRef]
- Martin, A.R.; Thomas, S.C. A reassessment of carbon content in tropical trees. PLoS One 2011, 6, e23533:1–e23533:9. [Google Scholar]
- Thomas, S.C.; Malczewski, G. Wood carbon content of tree species in eastern China: Interspecific variability and the importance of the volatile fraction. J. Environ. Manag. 2007, 85, 659–662. [Google Scholar] [CrossRef]
- Elias, M.; Potvin, C. Assessing inter- and intra-specific variation in trunk carbon concentration for 32 neotropical tree species. Can. J. For. Res. 2003, 33, 1039–1045. [Google Scholar] [CrossRef]
- Qureshi, A.; Pariva, R.B.; Hussain, S.A. A review of protocols used for assessment of carbon stock in forested landscapes. Environ. Sci. Policy 2012, 16, 81–89. [Google Scholar]
- Brown, S. Measuring carbon in forests: Current status and future challenges. Environ. Pollut. 2002, 116, 363–372. [Google Scholar] [CrossRef]
- Gibbs, H.K.; Brown, S.; Niles, J.O.; Foley, J.A. Monitoring and estimating tropical forest carbon stocks: Making REDD a reality. Environ. Res. Lett. 2007, 2, 1–13. [Google Scholar]
- Feldpausch, T.R.; Rondon, M.A.; Fernandes, E.C.M.; Riha, S.J.; Wandelli, E. Carbon and nutrient accumulation in secondary forests regenerating on pastures in central Amazonia. Ecol. Appl. 2004, 14, 164–176. [Google Scholar]
- Hughes, R.F.; Kauffman, J.B.; Jaramillo, V.J. Ecosystem-scale impacts of deforestation and land use in a humid tropical region of Mexico. Ecol. Appl. 2000, 10, 515–527. [Google Scholar] [CrossRef]
- Ruiz-Jaen, M.C.; Potvin, C. Can we predict carbon stocks in tropical ecosystems from tree diversity? Comparing species and functional diversity in a plantation and a natural forest. New Phytol. 2011, 189, 978–987. [Google Scholar] [CrossRef]
- Bradford, J.B.; Fraver, S.; Milo, A.M.; D’Amato, A.W.; Palik, B.; Shinneman, D.J. Effects of multiple interacting disturbances and salvage logging on forest carbon stocks. For. Ecol. Manag. 2012, 267, 209–214. [Google Scholar] [CrossRef]
- Chave, J.; Condit, R.; Muller-Landau, H.C.; Thomas, S.C.; Ashton, P.S.; Bunyavejchewin, S.; Co, L.L.; Dattaraja, H.S.; Davies, S.J.; Esufali, S.; et al. Assessing evidence for a pervasive alteration in tropical tree communities. PLoS Biol. 2008, 6, 455–462. [Google Scholar]
- Lewis, S.L.; Lopez-Gonzalez, G.; Sonke, B.; Affum-Baffoe, K.; Baker, T.R.; Ojo, L.O.; Phillips, O.L.; Reitsma, J.M.; White, L.; Comiskey, J.A.; et al. Increasing carbon storage in intact African tropical forests. Nature 2009, 457, 1003–1006. [Google Scholar]
- Pyle, E.H.; Santoni, G.W.; Nascimento, H.E.M.; Hutyra, L.R.; Vieira, S.; Curran, D.J.; van Haren, J.; Saleska, S.R.; Chow, V.Y.; Carmago, P.B.; et al. Dynamics of carbon, biomass, and structure in two Amazonian forests. J. Geophys. Res. 2008, 113, G00B08:1–G00B08:20. [Google Scholar]
- Saatchi, S.S.; Harris, N.L.; Brown, S.; Lefsky, M.; Mitchard, E.T.A.; Salas, W.; Zutta, B.R.; Buermann, W.; Lewis, S.L.; Hagen, S.; et al. Benchmark map of forest carbon stocks in tropical regions across three continents. Proc. Natl. Acad. Sci. USA 2011, 108, 9899–9904. [Google Scholar]
- Fang, J.Y.; Chen, A.P.; Peng, C.H.; Zhao, S.Q.; Ci, L. Changes in forest biomass carbon storage in China between 1949 and 1998. Science 2001, 292, 2320–2322. [Google Scholar]
- Kurz, W.A.; Dymond, C.C.; White, T.M.; Stinson, G.; Shaw, C.H.; Rampley, G.J.; Smyth, C.; Simpson, B.N.; Neilson, E.T.; Trofymow, J.A.; et al. CBM-CFS3: A model of carbon-dynamics in forestry and land-use change implementing IPCC standards. Ecol. Model. 2009, 220, 480–504. [Google Scholar]
- Kauppi, P.E.; Tomppo, E.; Ferm, A. C and N storage in living trees within Finland since 1950s. Plant Soil 1995, 168, 633–638. [Google Scholar] [CrossRef]
- Fang, J.Y.; Oikawa, T.; Kato, T.; Mo, W.H.; Wang, Z.H. Biomass carbon accumulation by Japan’s forests from 1947 to 1995. Glob. Biogeochem. Cycles 2005, 19, GB2004:1–GB2004:10. [Google Scholar]
- Blanc, L.; Echard, M.; Herault, B.; Bonal, D.; Marcon, E.; Chave, J.; Baraloto, C. Dynamics of aboveground carbon stocks in a selectively logged tropical forest. Ecol. Appl. 2009, 19, 1397–1404. [Google Scholar] [CrossRef]
- Soto-Pinto, L.; Anzueto, M.; Mendoza, J.; Ferrer, G.J.; de Jong, B. Carbon sequestration through agroforestry in indigenous communities of Chiapas, Mexico. Agrofor. Syst. 2010, 78, 39–51. [Google Scholar] [CrossRef]
- Beets, P.N.; Brandon, A.M.; Goulding, C.J.; Kimberley, M.O.; Paul, T.S.H.; Searles, N. The inventory of carbon stock in New Zealand’s post-1989 planted forest for reporting under the Kyoto protocol. For. Ecol. Manag. 2011, 262, 1119–1130. [Google Scholar] [CrossRef]
- Redondo-Brenes, A.; Montagnini, F. Growth, productivity, aboveground biomass, and carbon sequestration of pure and mixed native tree plantations in the Caribbean lowlands of Costa Rica. For. Ecol. Manag. 2006, 232, 168–178. [Google Scholar] [CrossRef]
- Redondo-Brenes, A. Growth, carbon sequestration, and management of native tree plantations in humid regions of Costa Rica. New For. 2007, 34, 253–268. [Google Scholar] [CrossRef]
- Fahey, T.J.; Siccama, T.G.; Driscoll, C.T.; Likens, G.E.; Campbell, J.; Johnson, C.E.; Battles, J.J.; Aber, J.D.; Cole, J.J.; Fisk, M.C.; et al. The biogeochemistry of carbon at Hubbard Brook. Biogeochemistry 2005, 75, 109–176. [Google Scholar] [CrossRef]
- Saner, P.; Loh, Y.Y.; Ong, R.C.; Hector, A. Carbon stocks and fluxes in tropical lowland Dipterocarp rainforests in Sabah, Malaysian Borneo. PLoS ONE 2012, 7, e29642:1–e29642:11. [Google Scholar]
- Melson, S.L.; Harmon, M.E.; Fried, J.S.; Domingo, J.B. Estimates of live-tree carbon stores in the Pacific Northwest are sensitive to model selection. Carbon Balance Manag. 2011, 6, 1–16. [Google Scholar] [CrossRef]
- Zhang, Q.Z.; Wang, C.K.; Wang, X.C.; Quan, X.K. Carbon concentration variability of 10 Chinese temperate tree species. For. Ecol. Manag. 2009, 258, 722–727. [Google Scholar] [CrossRef]
- Jones, D.A.; O’Hara, K.L. Carbon density in managed coast redwood stands: implications for forest carbon estimation. Forestry 2012, 85, 99–110. [Google Scholar]
- IPCC, Forest lands. In Intergovernmental Panel on Climate Change Guidelines for National Greenhouse Gas Inventories; Institute for Global Environmental Strategies (IGES): Hayama, Japan, 2006; 4, p. 83.
- Mascaro, J.; Asner, G.P.; Muller-Landau, H.C.; van Breugel, M.; Hall, J.; Dahlin, K. Controls over aboveground forest carbon density on Barro Colorado Island, Panama. Biogeosciences 2011, 8, 1615–1629. [Google Scholar] [CrossRef]
- Pan, Y.D.; Birdsey, R.A.; Fang, J.Y.; Houghton, R.; Kauppi, P.E.; Kurz, W.A.; Phillips, O.L.; Shvidenko, A.; Lewis, S.L.; Canadell, J.G.; et al. A large and persistent carbon sink in the world’s forests. Science 2011, 333, 988–993. [Google Scholar]
- Clark, D.A.; Brown, S.; Kicklighter, D.W.; Chambers, J.Q.; Thomlinson, J.R.; Ni, J. Measuring net primary production in forests: Concepts and field methods. Ecol. Appl. 2001, 11, 356–370. [Google Scholar] [CrossRef]
- Martin, A.R.; Thomas, S.C.; Zhao, Y. Size-dependent changes in wood chemical traits: A comparison of neotropical saplings and large trees. Oikos 2012. in review.. [Google Scholar]
- Becker, G.S.; Braun, D.; Gliniars, R.; Dalitz, H. Relations between wood variables and how they relate to tree size variables of tropical African tree species. Trees Struct. Funct. 2012, in press. [Google Scholar]
- Kraenzel, M.; Castillo, A.; Moore, T.; Potvin, C. Carbon storage of harvest-age teak (Tectona grandis) plantations, Panama. For. Ecol. Manag. 2003, 173, 213–225. [Google Scholar] [CrossRef]
- Lamlom, S.H.; Savidge, R.A. Carbon content variation in boles of mature sugar maple and giant sequoia. Tree Physiol. 2006, 26, 459–468. [Google Scholar] [CrossRef]
- Bert, D.; Danjon, F. Carbon concentration variations in the roots, stem and crown of mature Pinus pinaster (Ait.). For. Ecol. Manag. 2006, 222, 279–295. [Google Scholar] [CrossRef]
- Telmo, C.; Lousada, J.; Moreira, N. Proximate analysis, backwards stepwise regression between gross calorific value, ultimate and chemical analysis of wood. Bioresour. Technol. 2010, 101, 3808–3815. [Google Scholar]
- Laiho, R.; Laine, J. Tree stand biomass and carbon content in an age sequence of drained pine mires in southern Finland. For. Ecol. Manag. 1997, 93, 161–169. [Google Scholar]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 2nd ed; SAGE: Thousand Oaks, CA, USA, 2011. [Google Scholar]
- Searle, S.R.; Speed, F.M.; Milliken, G.A. Population marginal means in the linear-model—An alternative to least-squares means. Am. Stat. 1980, 34, 216–221. [Google Scholar]
- Nlme: Linear and Nonlinear Mixed Effects Models, version 3.3.1; Software for mixed-effects models, José Pinheiro and Douglas Bates: Murray Hill, NJ, USA, 2012.
- Thomas, S.C.; Martin, A.R. Dryad wood carbon content database. Dryad. 2012. Available online: http://dx.doi.org/10.5061/dryad.69sg2 (accessed on 18 June 2012). [CrossRef]
- Arias, D.; Calvo-Alvarado, J.; Richter, D.D.; Dohrenbusch, A. Productivity, aboveground biomass, nutrient uptake and carbon content in fast-growing tree plantations of native and introduced species in the southern region of Costa Rica. Biomass Bioenergy 2011, 35, 1779–1788. [Google Scholar] [CrossRef]
- Castaño-Santamaría, J.; Bravo, F. Variation in carbon concentration and basic density along stems of sessile oak (Quercus petraea (Matt.) Liebl.) and Pyrenean oak (Quercus pyrenaica Willd.) in the Cantabrian Range (NW Spain). Ann. For. Sci. 2012, in press. [Google Scholar] [CrossRef]
- Correia, A.C.; Tome, M.; Pacheco, C.A.; Faias, S.; Dias, A.C.; Freire, J.; Carvalho, P.O.; Pereira, J.S. Biomass allometry and carbon factors for a Mediterranean pine (Pinus pinea L.) in Portugal. For. Syst. 2010, 19, 418–433. [Google Scholar]
- Fang, S.; Xue, J.; Tang, L. Biomass production and carbon sequestration potential in poplar plantations with different management patterns. J. Environ. Manag. 2007, 85, 672–679. [Google Scholar] [CrossRef]
- Fukatsu, E.; Fukuda, Y.; Takahashi, M.; Nakada, R. Clonal variation of carbon content in wood of Larix kaempferi (Japanese larch). J. Wood Sci. 2008, 54, 247–251. [Google Scholar] [CrossRef]
- De Aza, C.H.; Turrion, M.B.; Pando, V.; Bravo, F. Carbon in heartwood, sapwood and bark along the stem profile in three Mediterranean Pinus species. Ann. For. Sci. 2011, 68, 1067–1076. [Google Scholar] [CrossRef]
- Huet, S.; Forgeard, F.; Nys, C. Above- and belowground distribution of dry matter and carbon biomass of Atlantic beech (Fagus sylvatica L.) in a time sequence. Ann. For. Sci. 2004, 61, 683–694. [Google Scholar] [CrossRef]
- Jacobs, D.F.; Selig, M.F.; Severeid, L.R. Aboveground carbon biomass of plantation-grown American chestnut (Castanea dentata) in absence of blight. For. Ecol. Manag. 2009, 258, 288–294. [Google Scholar] [CrossRef]
- Janssens, I.A.; Sampson, D.A.; Cermak, J.; Meiresonne, L.; Riguzzi, F.; Overloop, S.; Ceulemans, R. Above- and belowground phytomass and carbon storage in a Belgian Scots pine stand. Ann. For. Sci. 1999, 56, 81–90. [Google Scholar]
- Joosten, R.; Schulte, A. Possible effects of altered growth behaviour of Norway spruce (Picea abies) on carbon accounting. Clim. Change 2002, 55, 115–129. [Google Scholar] [CrossRef]
- Joosten, R.; Schumacher, J.; Wirth, C.; Schulte, A. Evaluating tree carbon predictions for beech (Fagus sylvatica L.) in western Germany. For. Ecol. Manag. 2004, 189, 87–96. [Google Scholar] [CrossRef]
- Kort, J.; Turnock, R. Carbon reservoir and biomass in Canadian prairie shelterbelts. Agrofor. Syst. 1998, 44, 175–186. [Google Scholar] [CrossRef]
- Li, X.; Yi, M.J.; Son, Y.; Park, P.S.; Lee, K.H.; Son, Y.M.; Kim, R.H.; Jeong, M.J. Biomass and carbon storage in an age sequence of Korean Pine (Pinus koraiensis) plantation forests in central Korea. J. Plant Biol. 2011, 54, 33–42. [Google Scholar] [CrossRef]
- Peri, P.L.; Gargaglione, V.; Pastur, G.M.; Lencinas, M.V. Carbon accumulation along a stand development sequence of Nothofagus antarctica forests across a gradient in site quality in Southern Patagonia. For. Ecol. Manag. 2010, 260, 229–237. [Google Scholar] [CrossRef]
- Rana, R.; Langenfeld-Heyser, R.; Finkeldey, R.; Polle, A. FTIR spectroscopy, chemical and histochemical characterisation of wood and lignin of five tropical timber wood species of the family of Dipterocarpaceae. Wood Sci. Technol. 2010, 44, 225–242. [Google Scholar] [CrossRef]
- Tolunay, D. Carbon concentrations of tree components, forest floor and understorey in young Pinus sylvestris stands in north-western Turkey. Scand. J. For. Res. 2009, 24, 394–402. [Google Scholar] [CrossRef]
- Van Geffen, K.G.; Poorter, L.; Sass-Klaasssen, U.; van Logtestijn, R.S.P.; Cornelissen, H.C. The trait contribution to wood decomposition rates of 15 neotropical tree species. Ecology 2010, 91, 3686–3697. [Google Scholar] [CrossRef]
- Zabek, L.M.; Prescott, C.E. Biomass equations and carbon content of aboveground leafless biomass of hybrid poplar in coastal British Columbia. For. Ecol. Manag. 2006, 223, 291–302. [Google Scholar] [CrossRef]
- Zheng, H.; Ouyang, Z.Y.; Xu, W.H.; Wang, X.K.; Miao, H.; Li, X.Q.; Tian, Y.X. Variation of carbon storage by different reforestation types in the hilly red soil region of southern China. For. Ecol. Manag. 2008, 255, 1113–1121. [Google Scholar] [CrossRef]
- Minami, E.; Saka, S. Comparison of the decomposition behaviors of hardwood and softwood in supercritical methanol. J. Wood Sci. 2003, 49, 73–78. [Google Scholar] [CrossRef]
- Campbell, M.M.; Sederoff, R.R. Variation in lignin content and composition—Mechanism of control and implications for the genetic improvement of plants. Plant Physiol. 1996, 110, 3–13. [Google Scholar]
- Hoch, G.; Richter, A.; Korner, C. Non-structural carbon compounds in temperate forest trees. Plant Cell Environ. 2003, 26, 1067–1081. [Google Scholar] [CrossRef]
- Daube, W. Chemische analysen des kernund splintholzes wichtiger waldbäume. Forstli. Blätter 1883, 20, 177–192. [Google Scholar]
- Lachenbruch, B.; Moore, J.R.; Evans, R. Radial variation in wood structure and function in woody plants, and hypotheses for its occurrence. In Size- and Age-Related Changes in Tree Structure and Function; Meinzer, F.C., Dawson, T., Lachenbruch, B.J., Eds.; Springer-Verlag: Berlin, Germany, 2011; pp. 121–164. [Google Scholar]
- Du, S.; Yamamoto, F. An overview of the biology of reaction wood formation. J. Integr. Plant Biol. 2007, 49, 131–143. [Google Scholar] [CrossRef]
- Potvin, C.; Mancilla, L.; Buchmann, N.; Monteza, J.; Moore, T.; Murphy, M.; Oelmann, Y.; Scherer-Lorenzen, M.; Turner, B.L.; Wilcke, W.; et al. An ecosystem approach to biodiversity effects: Carbon pools in a tropical tree plantation. For. Ecol. Manag. 2011, 261, 1614–1624. [Google Scholar] [CrossRef]
- Valencia, R.; Condit, R.; Foster, R.B.; Romoleroux, K.; Villa-Munoz, G.; Svenning, J.C.; Magard, E.; Bass, M.; Losos, E.C.; Balslev, H. Yasuni forest dynamics plot, Ecuador. In Tropical Forest Diversity and Dynamism: Findings from a Large-Scale Plot Network; Losos, E.C., Leigh, E.G., Eds.; University of Chicago Press: Chicago, IL, USA, 2004; pp. 609–620. [Google Scholar]
- Lee, H.S.; Tan, S.; Davies, S.J.; La-Frankie, J.V.; Ashton, P.S.; Yakamura, T.; Itoh, A.; Ohkubo, T.; Harrison, R. Lambir forest dynamics plot, Sarawak, Malaysia. In Tropical Forest Diversity and Dynamism: Findings from a Large-Scale Plot Network; Losos, E.C., Leigh, E.G., Eds.; University of Chicago Press: Chicago, IL, USA, 2004; pp. 527–539. [Google Scholar]
- Asner, G.P. Tropical forest carbon assessment: Integrating satellite and airborne mapping approaches. Environ. Res. Lett. 2009, 4, 034009:1–034009:11. [Google Scholar]
- Patenaude, G.; Hill, R.A.; Milne, R.; Gaveau, D.L.A.; Briggs, B.B.J.; Dawson, T.P. Quantifying forest above ground carbon content using LiDAR remote sensing. Remote Sens. Environ. 2004, 93, 368–380. [Google Scholar] [CrossRef]
- Brandtberg, T. Classifying individual tree species under leaf-off and leaf-on conditions using airborne lidar. ISPRS J. Photogramm. Remote Sens. 2007, 61, 325–340. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E. Airborne spectranomics: Mapping canopy chemical and taxonomic diversity in tropical forests. Front. Ecol. Environ. 2009, 7, 269–276. [Google Scholar] [CrossRef]
- Bond-Lamberty, B.; Wang, C.; Gower, S.T. Aboveground and belowground biomass and sapwood area allometric equations for six boreal tree species of northern Manitoba. Can. J. Forest Res. 2002, 32, 1441–1450. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.Q.; Eamus, D.; Folster, H.; Fromard, F.; Higuchi, N.; Kira, T.; et al. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Wang, C.K. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. For. Ecol. Manag. 2006, 222, 9–16. [Google Scholar] [CrossRef]
- Jenkins, J.C.; Chojnacky, D.C.; Heath, L.S.; Birdsey, R.A. National-scale biomass estimators for United States tree species. For. Sci. 2003, 49, 12–35. [Google Scholar]
- Zanne, A.E.; Lopez-Gonzalez, G.; Coomes, D.A.; Ilic, J.; Jansen, S.; Lewis, S.L.; Miller, R.B.; Swenson, N.G.; Wiemann, M.C.; Chave, J. Towards a worldwide wood economics spectrum. Dryad Digital Repository. 2009. Available online: http://hdl.handle.net/10255/dryad.235 (accessed on 11 June 2012). [CrossRef]
- Flores, O.; Coomes, D.A. Estimating the wood density of species for carbon stock assessments. Methods Ecol. Evol. 2011, 2, 214–220. [Google Scholar] [CrossRef]
- Schnitzer, S.A.; Bongers, F. Increasing liana abundance and biomass in tropical forests: Emerging patterns and putative mechanisms. Ecol. Lett. 2011, 14, 397–406. [Google Scholar] [CrossRef]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Thomas, S.C.; Martin, A.R. Carbon Content of Tree Tissues: A Synthesis. Forests 2012, 3, 332-352. https://doi.org/10.3390/f3020332
Thomas SC, Martin AR. Carbon Content of Tree Tissues: A Synthesis. Forests. 2012; 3(2):332-352. https://doi.org/10.3390/f3020332
Chicago/Turabian StyleThomas, Sean C., and Adam R. Martin. 2012. "Carbon Content of Tree Tissues: A Synthesis" Forests 3, no. 2: 332-352. https://doi.org/10.3390/f3020332