

Redbay (Persea borbonia L. Spreng) Seedling and Sapling Growth and Recruitment Near Trees with and without Putative Resistance to Laurel Wilt Disease

Abstract
1. Introduction
2. Materials and Methods
2.1. Survey Sites and Initial Data Collection
2.2. Survey Design
2.2.1. Redbay Sapling Survey
2.2.2. Redbay Seedling Survey
2.3. Statistical Analysis
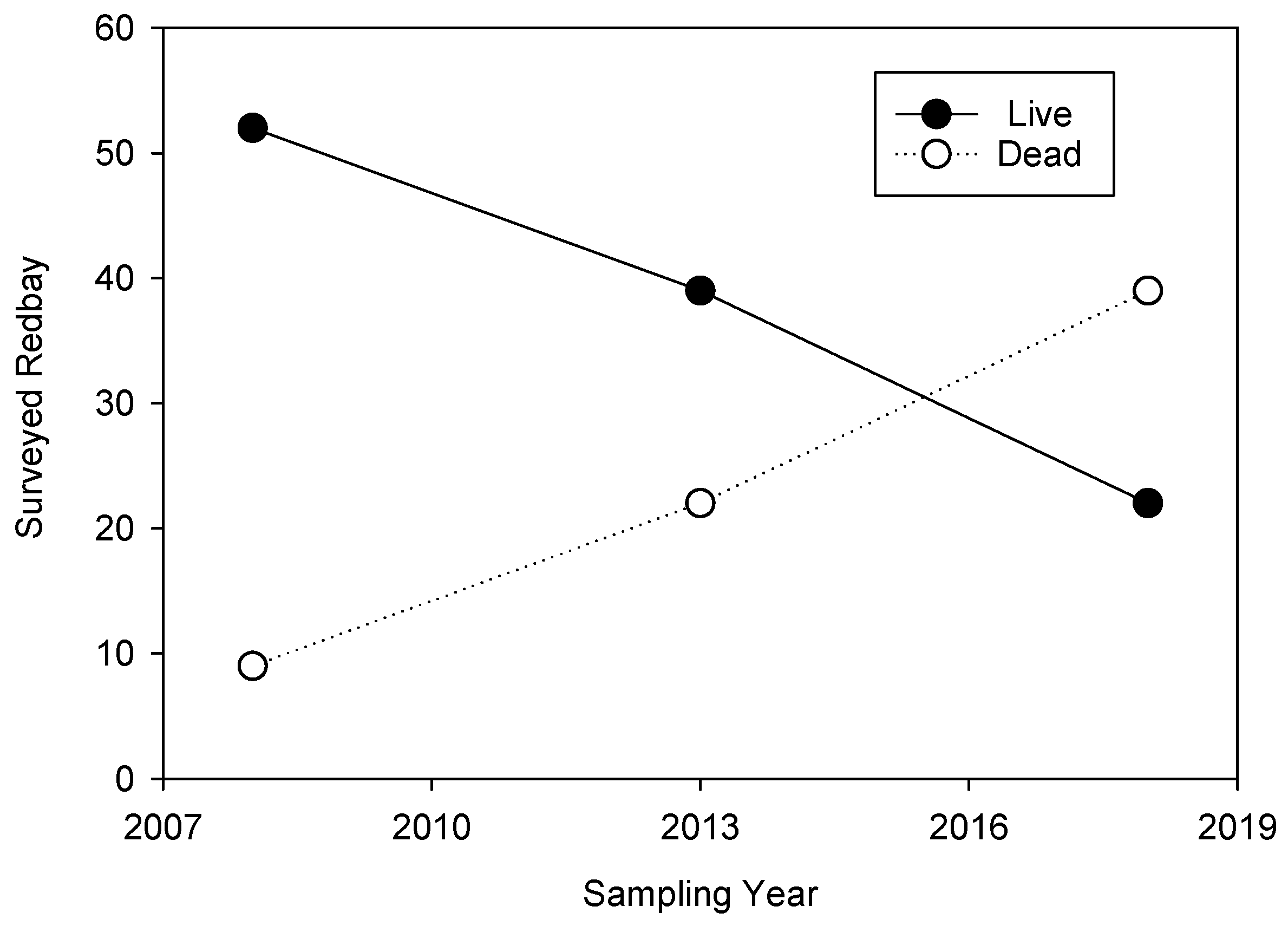
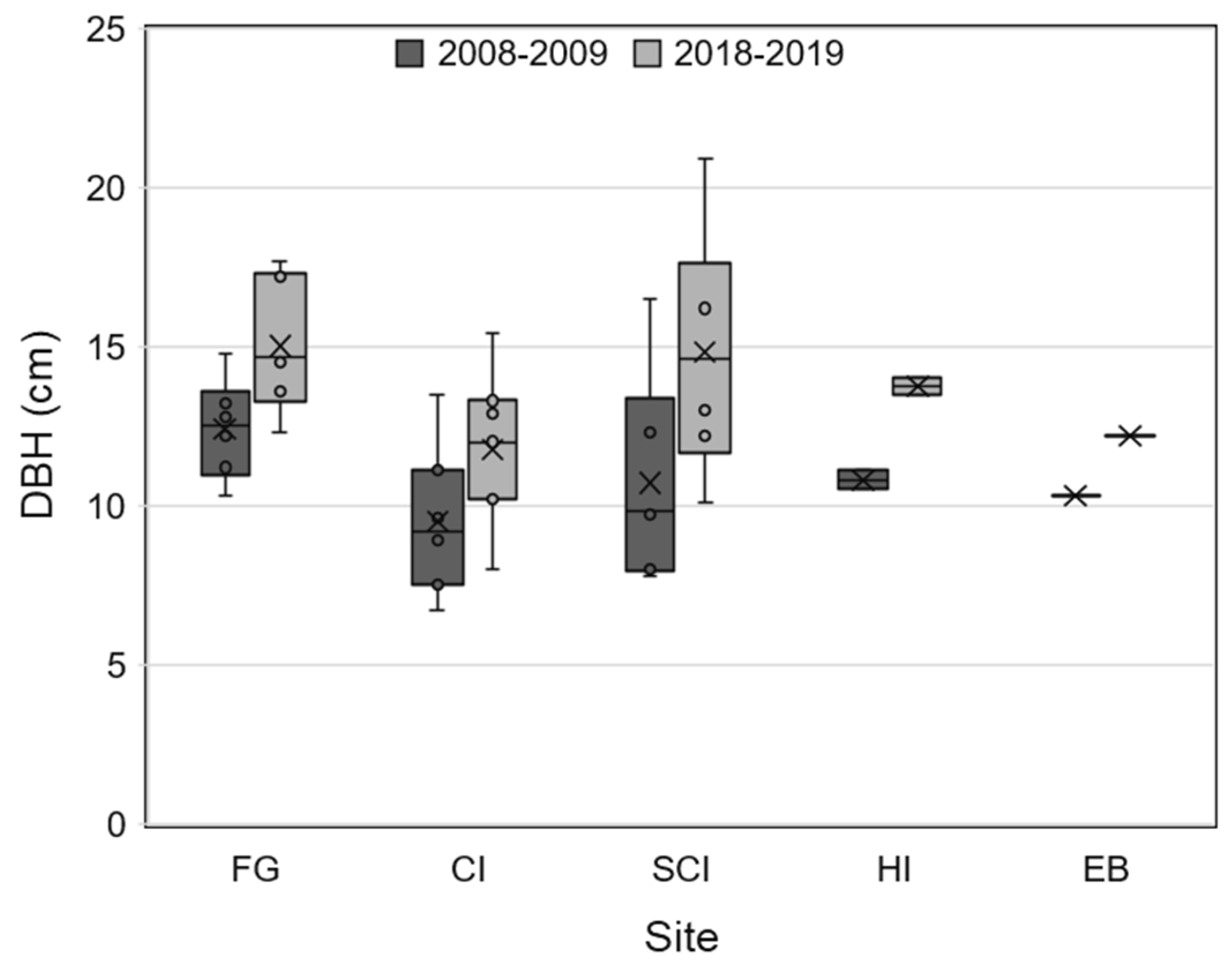
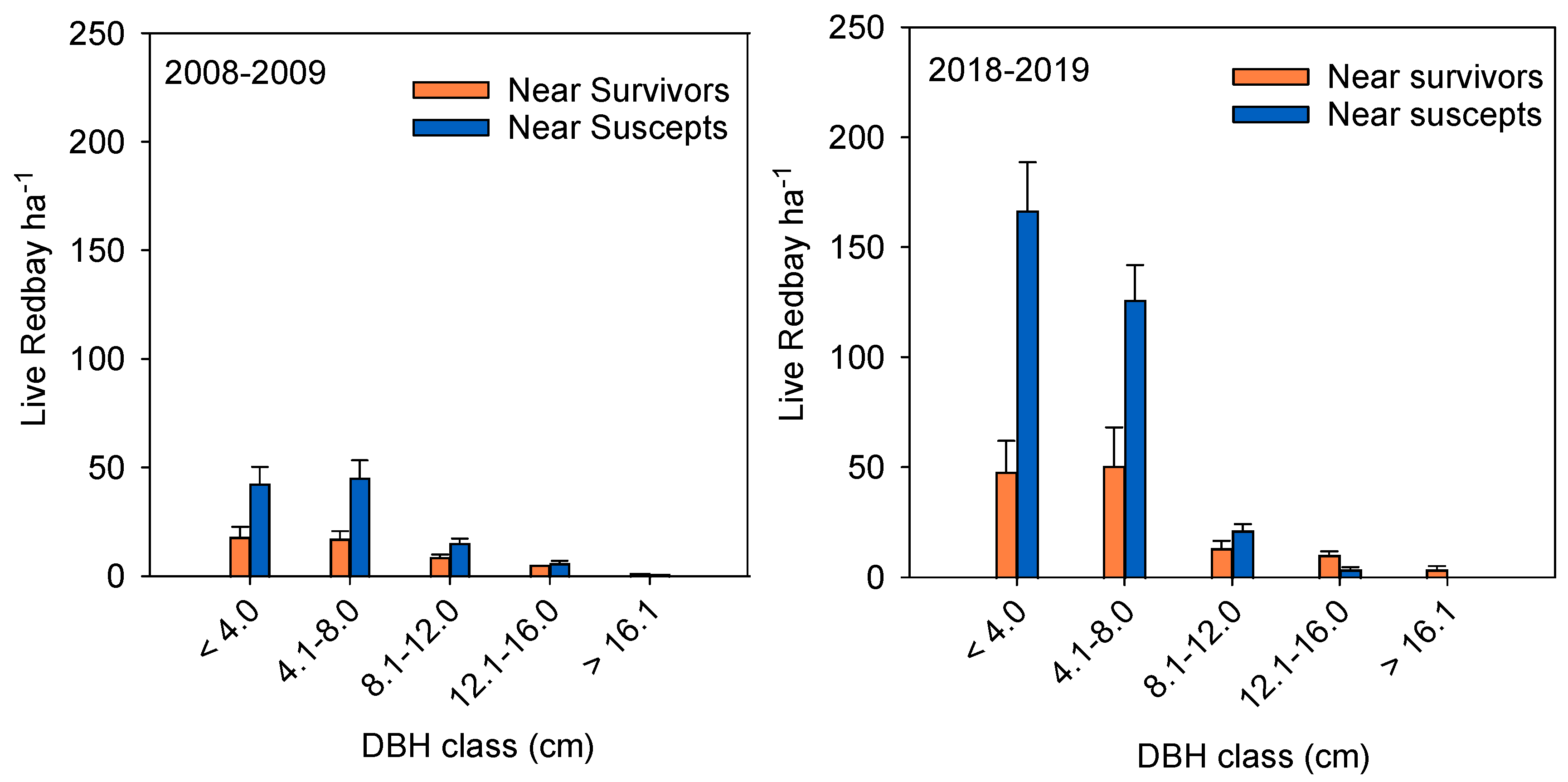
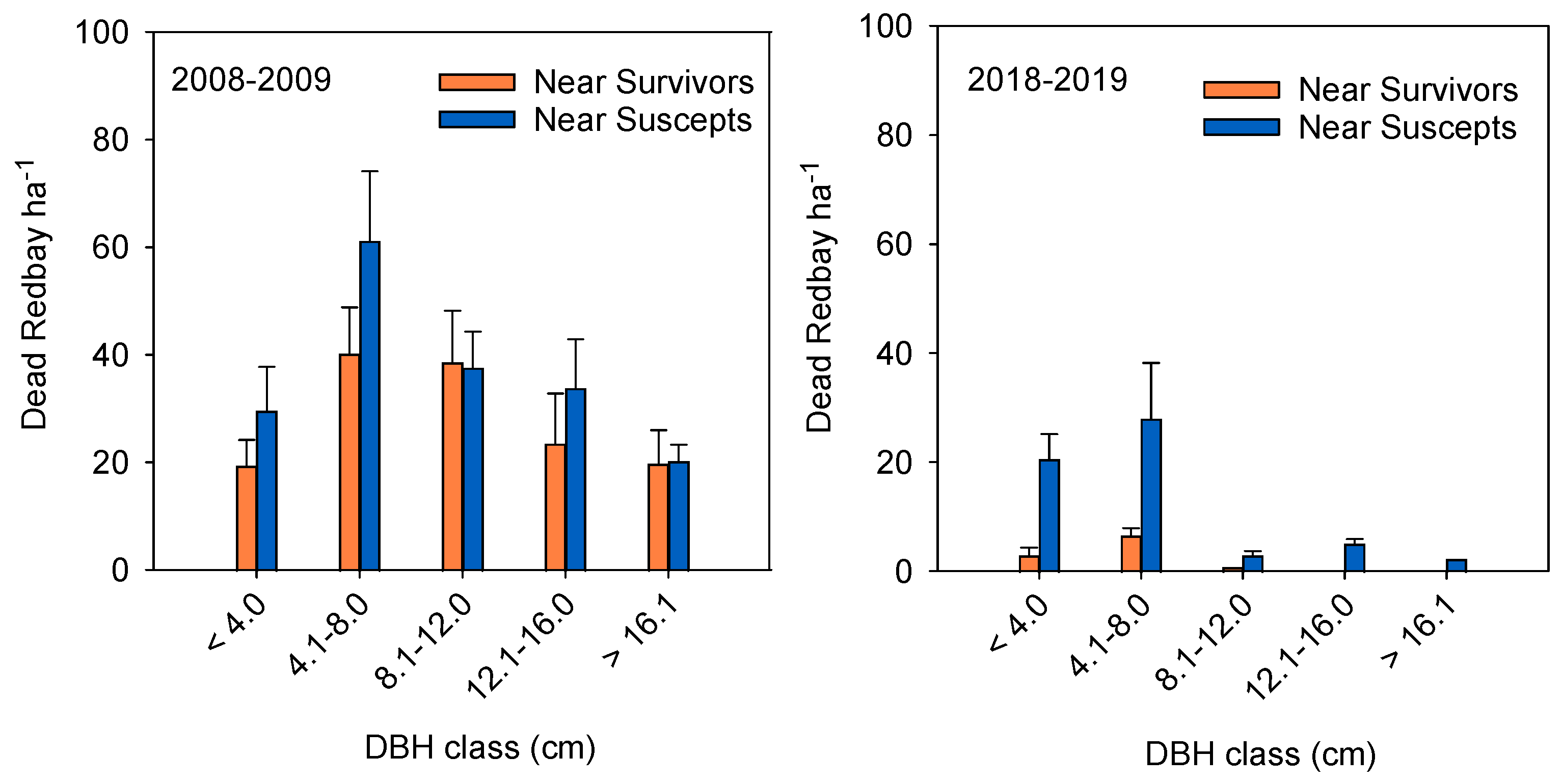
3. Results
3.1. Field Surveys
3.1.1. Redbay Sapling Survey
Understory Redbay Survey
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liebhold, A.M.; MacDonald, W.L.; Bergdahl, D.; Mastro, V.C. Invasion by Exotic Forest Pests: A Threat to Forest Ecosystems. For. Sci. 1995, 41, a0001–z0001. [Google Scholar] [CrossRef]
- Lovett, G.M.; Canham, C.D.; Arthur, M.A.; Weathers, K.C.; Fitzhugh, R.D. Forest Ecosystem Responses to Exotic Pests and Pathogens in Eastern North America. Bioscience 2006, 56, 395–405. [Google Scholar] [CrossRef]
- Meyer, S.E.; Callaham, M.A., Jr.; Stewart, J.E.; Warren, S.D. Invasive Species Response to Natural and Anthropogenic Disturbance. In Invasive Species in Forests and Rangelands of the United States; Poland, T.M., Patel-Weynand, T., Finch, D.M., Miniat, C.F., Hayes, D.C., Lopez, V.M., Eds.; Springer International Publishing: Cham, Switzerland, 2021; pp. 85–110. ISBN 978-3-030-45366-4. [Google Scholar]
- Prakash, S.; Verma, A.K. Anthropogenic Activities and Biodiversity Threats. Int. J. Biol. Innov. 2022, 4, 94–103. [Google Scholar] [CrossRef]
- Loehle, C.; Hulcr, J.; Smith, J.A.; Munro, H.L.; Fox, T. Preventing the Perfect Storm of Forest Mortality in the United States Caused by Invasive Species. J. For. 2023, 121, 104–117. [Google Scholar] [CrossRef]
- D’Amato, A.W.; Orwig, D.A.; Siegert, N.W.; Mahaffey, A.; Benedict, L.; Everett, T.; Daigle, J.; Johnson, L.; Catanzaro, P.; Cusack, C. Species Preservation in the Face of Novel Threats: Cultural, Ecological, and Operational Considerations for Preserving Tree Species in the Context of Non-Indigenous Insects and Pathogens. J. For. 2023, 121, 470–479. [Google Scholar] [CrossRef]
- Grittinger, T.F. Loss of Elm from Some Lowland Forests in Eastern Wisconsin. Trans. Wis. Acad. Sci. Arts Lett. 1978, 66, 195–205. [Google Scholar]
- Jacobs, D.F. Toward Development of Silvical Strategies for Forest Restoration of American Chestnut (Castanea dentata) Using Blight-Resistant Hybrids. Biol. Conserv. 2007, 137, 497–506. [Google Scholar] [CrossRef]
- Weed, A.S.; Ayres, M.P.; Hicke, J.A. Consequences of Climate Change for Biotic Disturbances in North American Forests. Ecol. Monogr. 2013, 83, 441–470. [Google Scholar] [CrossRef]
- Cobb, R.C.; Haas, S.E.; Kruskamp, N.; Dillon, W.W.; Swiecki, T.J.; Rizzo, D.M.; Frankel, S.J.; Meentemeyer, R.K. The Magnitude of Regional-Scale Tree Mortality Caused by the Invasive Pathogen Phytophthora ramorum. Earth’s Future 2020, 8, e2020EF001500. [Google Scholar] [CrossRef]
- Fraedrich, S.W.; Harrington, T.C.; Rabaglia, R.J.; Ulyshen, M.D.; Mayfield, A.E.; Hanula, J.L.; Eickwort, J.M.; Miller, D.R. A Fungal Symbiont of the Redbay Ambrosia Beetle Causes a Lethal Wilt in Redbay and Other Lauraceae in the Southeastern United States. Plant Dis. 2008, 92, 215–224. [Google Scholar] [CrossRef]
- Harrington, T.; Fraedrich, S.W.; Aghayeva, D.N. Raffaelea lauricola, a New Ambrosia Beetle Symbiont and Pathogen on the Lauraceae. Mycotaxon 2008, 104, 399–404. [Google Scholar]
- Inch, S.; Ploetz, R.; Held, B.; Blanchette, R. Histological and Anatomical Responses in Avocado, Persea americana, Induced by the Vascular Wilt Pathogen, Raffaelea lauricola. Botany 2012, 90, 627–635. [Google Scholar] [CrossRef]
- Loyd, A.L.; Chase, K.D.; Nielson, A.; Hoover, N.; Dreaden, T.J.; Mayfield, A.E.; Crocker, E.; Fraedrich, S.W. First Report of Laurel Wilt Caused by Raffaelea Lauricola on Sassafras Albidum in Tennessee and Kentucky. Plant Dis. 2020, 104, 567. [Google Scholar] [CrossRef]
- Eaton, M.J.; Beale, J.; Long, S.; Dreaden, T.J.; Blevins, A.; Mayfield, A.; Buland, M.; Hadziabdic, D.; Crocker, E.V. First Report of Laurel Wilt Caused by Harringtonia lauricola (Previously Raffaelea lauricola) on Northern Spicebush in Kentucky and Tennessee. Plant Dis. 2023, 107, 1221. [Google Scholar] [CrossRef] [PubMed]
- Inch, S.A.; Ploetz, R.C. Impact of Laurel Wilt, Caused by Raffaelea Lauricola, on Xylem Function in Avocado, Persea Americana. For. Pathol. 2012, 42, 239–245. [Google Scholar] [CrossRef]
- Kendra, P.E.; Montgomery, W.S.; Niogret, J.; Epsky, N.D. An Uncertain Future for American Lauraceae: A Lethal Threat from Redbay Ambrosia Beetle and Laurel Wilt Disease (A Review). Am. J. Plant. Sci. 2013, 04, 727–738. [Google Scholar] [CrossRef]
- Castillo-Argaez, R.; Vazquez, A.; Konkol, J.L.; Vargas, A.I.; Ploetz, R.C.; Etxeberria, E.; Schaffer, B. Sap Flow, Xylem Anatomy and Photosynthetic Variables of Three Persea Species in Response to Laurel Wilt. Tree Physiol. 2021, 41, 1004–1018. [Google Scholar] [CrossRef] [PubMed]
- Shields, J.; Jose, S.; Freeman, J.; Bunyan, M.; Celis, G.; Hagan, D.; Morgan, M.; Pieterson, E.C.; Zak, J. Short-Term Impacts of Laurel Wilt on Redbay (Persea borbonia [L.] Spreng.) in a Mixed Evergreen-Deciduous Forest in Northern Florida. J. For. 2011, 109, 82–88. [Google Scholar] [CrossRef]
- Hughes, M.A.; Riggins, J.J.; Koch, F.H.; Cognato, A.I.; Anderson, C.; Formby, J.P.; Dreaden, T.J.; Ploetz, R.C.; Smith, J.A. No Rest for the Laurels: Symbiotic Invaders Cause Unprecedented Damage to Southern USA Forests. Biol. Invasions 2017, 19, 2143–2157. [Google Scholar] [CrossRef]
- Brendemuehl, R.H. Persea borbonia (L.) Spreng. Redbay. In Silvics of North America; Burns, R.M., Honkala, B.H., Eds.; Forest Service Agricultural Handbook 654; U.S. Department of Agriculture: Washington, DC, USA, 1990; Volume 2, pp. 503–506. [Google Scholar]
- Chupp, A.D.; Battaglia, L.L. Potential for Host Shifting in Papilio Palamedes Following Invasion of Laurel Wilt Disease. Biol. Invasions 2014, 16, 2639–2651. [Google Scholar] [CrossRef]
- Riggins, J.J.; Chupp, A.D.; Formby, J.P.; Dearing, N.A.; Bares, H.M.; Brown, R.L.; Oten, K.F. Impacts of Laurel Wilt Disease on Arthropod Herbivores of North American Lauraceae. Biol. Invasions 2019, 21, 493–503. [Google Scholar] [CrossRef]
- Spiegel, K.S.; Leege, L.M. Impacts of Laurel Wilt Disease on Redbay (Persea borbonia (L.) Spreng.) Population Structure and Forest Communities in the Coastal Plain of Georgia, USA. Biol. Invasions 2013, 15, 2467–2487. [Google Scholar] [CrossRef]
- Evans, J.P.; Scheffers, B.R.; Hess, M. Effect of Laurel Wilt Invasion on Redbay Populations in a Maritime Forest Community. Biol. Invasions 2014, 16, 1581–1588. [Google Scholar] [CrossRef]
- Ward, S.F.; Riggins, J.J. Drivers of Invasion by Laurel Wilt of Redbay and Sassafras in the Southeastern US. Landsc. Ecol. 2023, 38, 567–581. [Google Scholar] [CrossRef] [PubMed]
- Parra, P.P.; Dantes, W.; Sandford, A.; de la Torre, C.; Pérez, J.; Hadziabdic, D.; Schaffer, B.; Gazis, R. Rapid Detection of the Laurel Wilt Pathogen in Sapwood of Lauraceae Hosts. Plant Health Prog. 2020, 21, 356–364. [Google Scholar] [CrossRef]
- Hughes, M.A.; Smith, J.A.; Ploetz, R.C.; Kendra, P.E.; Mayfield, A.E.; Hanula, J.L.; Hulcr, J.; Stelinski, L.L.; Cameron, S.; Riggins, J.J.; et al. Recovery Plan for Laurel Wilt on Redbay and Other Forest Species Caused by Raffaelea lauricola and Disseminated by Xyleborus glabratus. Plant Health Prog. 2015, 16, 173–210. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Kendra, P.E.; Choudhury, R.A.; Rollins, J.A.; Campbell, A.; Garrett, K.; Hughes, M.; Dreaden, T. Laurel Wilt in Natural and Agricultural Ecosystems: Understanding the Drivers and Scales of Complex Pathosystems. Forests 2017, 8, 48. [Google Scholar] [CrossRef]
- Hamilton, J.L.; Fraedrich, S.W.; Nairn, C.J.; Mayfield, A.E.; Villari, C. A Field-Portable Diagnostic Approach Confirms Laurel Wilt Disease Diagnosis in Minutes Instead of Days. Arboric. Urban For. 2021, 47, 98–109. [Google Scholar] [CrossRef]
- Hughes, M.A.; Smith, K.E.; Sims, A.; Zhang, J.; Held, B.W.; Blanchette, R.A.; Smith, J.A. Screening of Persea Borbonia Clones for Resistance to the Laurel Wilt Pathogen, Raffaelea lauricola. For. Pathol. 2022, 52, e12776. [Google Scholar] [CrossRef]
- Schlarbaum, S.E.; Hebard, F.; Spaine, P.C.; Kamalay, J.C. Three American Tragedies: Chestnut Blight, Butternut Canker, and Dutch Elm Disease. In Proceedings of the Exotic Pests of Eastern Forests Conference Proceedings, Nashville, TN, USA, 8–10 April 1997; Britton, K.O., Ed.; U.S. Forest Service and Tennessee Exotic Pest Plant Council: Nashville, TN, USA, 1998; pp. 45–54. [Google Scholar]
- Sniezko, R.A.; Johnson, J.S.; Reeser, P.; Kegley, A.; Hansen, E.M.; Sutton, W.; Savin, D.P. Genetic Resistance to Phytophthora Lateralis in Port-Orford-Cedar (Chamaecyparis lawsoniana)—Basic Building Blocks for a Resistance Program. Plants People Planet 2020, 2, 69–83. [Google Scholar] [CrossRef]
- Luiz, B.C.; Giardina, C.P.; Keith, L.M.; Jacobs, D.F.; Sniezko, R.A.; Hughes, M.A.; Friday, J.B.; Cannon, P.; Hauff, R.; Francisco, K.; et al. A Framework for Establishing a Rapid ‘Ōhi‘a Death Resistance Program. New For. 2023, 54, 637–660. [Google Scholar] [CrossRef]
- Hughes, M.A. The Evaluation of Natural Resistance to Laurel Wilt Disease in Redbay (Persea borbonia). Doctoral Dissertation, University of Florida, Gainesville, FL, USA, 2013. [Google Scholar]
- Gonzalez-Akre, E.; Meakem, V.; Eng, C.Y.; Tepley, A.J.; Bourg, N.A.; McShea, W.; Davies, S.J.; Anderson-Teixeira, K. Patterns of Tree Mortality in a Temperate Deciduous Forest Derived from a Large Forest Dynamics Plot. Ecosphere 2016, 7, e01595. [Google Scholar] [CrossRef]
- Eicholtz, M. An Assessment of Redbay Regeneration (Persea borbonia) Regeneration Potential Post-Laurel Wilt Disease Epidemic. Master’s Thesis, University of Florida, Gainesville, FL, USA, 2019. [Google Scholar]
- Cameron, R.S.; Hanula, J.; Fraedrich, S.; Bates, C. Progression and Impact of Laurel Wilt Disease within Redbay and Sassafras Populations in Southeast Georgia. Southeast. Nat. 2015, 14, 650–674. [Google Scholar] [CrossRef]
- Choudhury, R.A.; Er, H.L.; Hughes, M.A.; Smith, J.A.; Pruett, G.E.; Konkol, J.; Ploetz, R.C.; Marois, J.J.; Garrett, K.A.; van Bruggen, A.H.C. Host Density Dependence and Environmental Factors Affecting Laurel Wilt Disease Incidence. Plant Pathol. 2021, 70, 676–688. [Google Scholar] [CrossRef]
- Mcdonald, B.A.; Linde, C. The Population Genetics of Plant Pathogens and Breeding Strategies for Durable Resistance. Euphytica 2002, 124, 163–180. [Google Scholar] [CrossRef]
- Kinloch, B.B. Race of Cronartium Ribicola Virulent to Major Gene Resistance in Sugar Pine. Plant Dis. 1981, 65, 604. [Google Scholar] [CrossRef]
- Ennos, R.A. Resilience of Forests to Pathogens: An Evolutionary Ecology Perspective. Forestry 2015, 88, 41–52. [Google Scholar] [CrossRef]
- Martín, J.A.; Domínguez, J.; Solla, A.; Brasier, C.M.; Webber, J.F.; Santini, A.; Martínez-Arias, C.; Bernier, L.; Gil, L. Complexities Underlying the Breeding and Deployment of Dutch Elm Disease Resistant Elms. New For. 2023, 54, 661–696. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, J.; Vanderpool, D.; Smith, J.A.; Rollins, J.A. Genomic and Transcriptomic Insights into Raffaelea lauricola Pathogenesis. BMC Genom. 2020, 21, 1–23. [Google Scholar] [CrossRef]
- Boshier, D.; Buggs, R.J.A. The Potential for Field Studies and Genomic Technologies to Enhance Resistance and Resilience of British Tree Populations to Pests and Pathogens. Forestry 2015, 88, 27–40. [Google Scholar] [CrossRef]
- Myers, A.L.; Storer, A.J.; Dickinson, Y.L.; Bal, T.L. A Review of Propagation and Restoration Techniques for American Beech and Their Current and Future Application in Mitigation of Beech Bark Disease. Sustainability 2023, 15, 7490. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Survivor Plots | Suscept Plots | Survivor Effect | Site Effect | Survivor × Site Interaction | |
|---|---|---|---|---|---|
| ΔTPH | 58.4 ± 21.7 | 60.8 ± 14.1 | p = 0.758 | p = 0.00599 * | p = 0.01050 * |
| ΔQMD | 1.36 ± 0.394 | −0.675 ± 0.448 | p = 0.00269 * | p = 0.95495 | p = 0.87504 |
| Site | Contrast | Estimate | SE | df | t-Value | p-Value | |
|---|---|---|---|---|---|---|---|
| ΔTPH | CINS | No–Yes | 1.62 | 2.28 | 50 | 0.707 | 0.4826 |
| EB | No–Yes | 2.46 | 3.47 | 50 | 0.708 | 0.4825 | |
| FG | No–Yes | 1.95 | 2.14 | 50 | 0.914 | 0.3649 | |
| HI | No–Yes | −9.48 | 2.62 | 50 | −3.622 | 0.0007 * | |
| SCI | No–Yes | −1.41 | 2.14 | 50 | −0.662 | 0.5111 |
| Survivor Plots | Suscept Plots | Survivor Effect | Site Effect | Survivor × Site Interaction | |
|---|---|---|---|---|---|
| Seedling TPH | 53.4 ± 21.6 | 90.3 ± 14.1 | p = 0.00264 * | p = 8.47×10−6 * | p = 0.01844 * |
| Sprout TPH | 30.3 ± 10.9 | 46.3 ± 7.9 | p = 0.0185 * | p = 1.31×10−8 * | p = 0.1118 |
| Sdu | 4.34 ± 0.99 | 12.00 ± 2.51 | p = 0.00755 * | p = 1.17×10−5 * | p = 0.23206 |
| Site | Contrast | Estimate | SE | df | t-Value | p-Value | |
|---|---|---|---|---|---|---|---|
| Seedling TPH | CINS | No–Yes | −0.583 | 2.59 | 50 | −0.225 | 0.8230 |
| EB | No–Yes | 4.879 | 3.94 | 50 | 1.237 | 0.2217 | |
| FG | No–Yes | 5.689 | 2.43 | 50 | 2.344 | 0.0231 * | |
| HI | No–Yes | −7.285 | 2.97 | 50 | −2.451 | 0.0178 * | |
| SCI | No–Yes | −0.757 | 2.43 | 50 | −0.312 | 0.7563 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eicholtz, M.; Smith, J.; Vogel, J. Redbay (Persea borbonia L. Spreng) Seedling and Sapling Growth and Recruitment Near Trees with and without Putative Resistance to Laurel Wilt Disease. Forests 2024, 15, 817. https://doi.org/10.3390/f15050817
Eicholtz M, Smith J, Vogel J. Redbay (Persea borbonia L. Spreng) Seedling and Sapling Growth and Recruitment Near Trees with and without Putative Resistance to Laurel Wilt Disease. Forests. 2024; 15(5):817. https://doi.org/10.3390/f15050817
Chicago/Turabian StyleEicholtz, Matthew, Jason Smith, and Jason Vogel. 2024. "Redbay (Persea borbonia L. Spreng) Seedling and Sapling Growth and Recruitment Near Trees with and without Putative Resistance to Laurel Wilt Disease" Forests 15, no. 5: 817. https://doi.org/10.3390/f15050817
APA StyleEicholtz, M., Smith, J., & Vogel, J. (2024). Redbay (Persea borbonia L. Spreng) Seedling and Sapling Growth and Recruitment Near Trees with and without Putative Resistance to Laurel Wilt Disease. Forests, 15(5), 817. https://doi.org/10.3390/f15050817