1. Introduction
Drought stress not only restricts the growth and yield of plants but also causes changes in plant structure and function [
1]. The root system is the first part of the plant to perceive drought stress. Drought stress directly causes root cells to lose water, which damages the cell membrane structure, disintegrates organelles, hinders cell metabolism, accelerates the aging process, and eventually leads to plant death [
2]. Mitochondria in plant root cells are sensitive to drought stress. Maintaining the normal structure and function of mitochondria plays an important role in plant drought resistance [
3,
4]. Under a drought stress environment, with the aggravation of drought stress, the mitochondrial matrix gradually becomes thinner and the internal cristae decrease; then, the mitochondrial membrane breaks down and disintegrates, small vacuoles and medullary structures appear in the cells, and the cells die [
4]. Under drought stress conditions, plants activate the expression of drought-resistant regulatory and functional genes, which encode functional proteins and organic molecular synthases, regulate drought signal transduction, and participate in the synthesis and accumulation of hormones and osmotic substances to help plants respond to drought stress. Abscisic acid (ABA) is a signal molecule that mediates the activation of these genes and some related molecular reactions [
5]. Under drought stress conditions, plant roots first sense stress signals, which stimulate the activity of ABA synthase. ABA is synthesized rapidly in root cap cells and actively transported to the growth site, reducing stomatal conductance and water loss of leaves [
6]. At the same time, under drought stress conditions, ABA can also improve root hydraulic conductivity by promoting the expression and protein activity of aquaporins (AQPs) [
7]. Besides ABA, gibberellic acid (GA), indole-3-acetic acid (IAA), and zeatin riboside (ZR) also respond synergistically to drought stress conditions by enhancing or inhibiting their synthetic ability, thereby increasing or decreasing the content levels of these hormones, changing the balance and ratio of hormones, slowing down plant growth and development to reduce excessive water consumption, maintaining normal cell swelling pressure in order to maintain normal water balance, promoting stomatal closure to reduce water transpiration, and increasing cell solute concentration to enhance water absorption capacity, thereby enhancing plant drought resistance [
8]. The response of plant root hormones to drought stress is ultimately reflected by changes in the root morphological structure and function. Under water stress conditions, in order to minimize root metabolism and maximize resource acquisition, the root morphology and configuration will change according to soil, water, and other factors [
9]. Drought stress can change the morphology and configuration of host roots, including the number of lateral roots, projected area, surface area, volume, and branch structure, and these changes are closely related to the balance of endogenous hormones [
10,
11].
Beneficial microorganisms (e.g., endophytic fungi, mycorrhizal fungi, and plant rhizosphere growth-promoting bacteria) in the plant rhizosphere can promote plant growth and increase plant tolerance to abiotic stresses [
12]. Beneficial microorganisms can enhance plant drought tolerance by either promoting drought stress avoidance by increasing water uptake or decreasing the transpiration rate, or by promoting tolerance by enhancing osmotic regulation [
13,
14]. Under drought stress, the colonization of plants by beneficial endophytic fungi can increase plant biomass, regulate osmotic substances, stabilize photosynthetic parameters, reduce stomatal conductance, and/or reduce overall water loss [
14,
15,
16]. In addition to changes to the aboveground parts, endophytic fungi can also change the growth of host roots, making the host better adapted to the drought environment [
9,
17]. In recent years, interest in dark septate endophytes (DSEs) has increased. DSEs are not a scientific taxonomic unit but generally refer to a group of endophytic fungi (Ascomycetes or Deuteromycetes) with dark (melanized), septate hyphae that form “microsclerotia” in plant root cells and reside endophytically in plant roots [
18,
19]. DSEs lack the typical structure of mycorrhizal fungi and colonize plant roots without forming a pathogenic association with the host. DSEs are widely distributed and have been found in almost all habitats. Recently, DSEs have been found to play an active role in improving host drought resistance [
20,
21]; however, our understanding of their role in drought resistance is still limited.
Ormosia hosiei Hemsley & E. H. Wilson is a precious timber species that is specially produced in China [
22].
O. hosiei has many important ecological and economic values. It is a kind of redwood tree species that is not only an excellent material for high-grade furniture but also for landscaping. However, a previous study found that
O. hosiei could not tolerate drought during its initial growth stages [
23]. Therefore, it is important to increase the drought resistance at the early stage, which is important to improve plant growth and productivity. A previous study found that DSEs increased the growth of
O. hosiei [
24]. However, other ecological functions, such as the drought resistance of DSE fungi for
O. hosiei, are still not well understood, and the mechanism by which DSEs improve plant growth under drought stress remains unclear. To date, the role of DSEs in improving host drought tolerance has only been reported sporadically in sorghum [
20] and
Ammopiptanthus mongolicus [
21]; both of these studies focused on descriptions of aboveground biomass or photosynthetic characteristics. Whether DSEs, like arbuscular mycorrhizal fungi, can change the morphology and configuration of roots to adapt the host to a drought environment, regulate the balance of endogenous hormones to improve plant drought resistance, and alleviate root ultrastructural damage under drought stress has yet to be determined.
In this study, aseptic O. hosiei seedlings inoculated with the DSE strain Acrocalymma vagum were cultivated for one growing season under three water stress conditions. The root cell ultrastructure, root morphology, and endogenous hormones of DSE-inoculated seedlings and noninoculated seedlings were investigated to further clarify the relationship between the effects of DSE on host root growth and the improvement of host drought tolerance.
2. Materials and Methods
2.1. Materials
In November 2017, the seeds of O. hosiei were collected from Guanling County, Guizhou Province, China. Healthy, pest-free seeds of the same size and shape and with smooth skin were selected, sterilized with 0.3% potassium permanganate for 1 h, washed with water several times, and then soaked for 12 h in hot water that was cooled to 90 °C after boiling. Swollen seeds were selected and washed with sterile water several times and then germinated in sterilized river sand for three weeks. After the radical emerged from the seeds, seedlings exhibiting similar growth were selected and transplanted to experimental pots.
The experimental fungal strain of
A. vagum used was isolated from the roots of a wild
O. hosiei tree [
24]. Potato dextrose liquid medium was inoculated with a small piece of mycelium of
A. vagum in a flask and cultured on a rotary shaker at 120 rpm for 15 days at 28 °C to obtain the mycelial mass. The mycelial mass was collected by filtration through sterile gauze and rinsed several times with sterile water. The mycelial mass was broken up and diluted to form a concentration of 1 × 10
5 cfu/mL.
Yellow loam soil from the nursery of the Guizhou Institute of Biology was used as the experimental soil. The soil was sieved through a 2 mm mesh and then mixed with river sand, perlite, and vermiculite in a volume ratio of 1:1:1:1 before sterilization at 121 °C and 0.05 MPa for 2 h. The soil was sterilized again the following day and then dried for use. The experimental soil contained 13.54 g/kg of organic substances, 48.39 mg/kg of alkali nitrogen, and 36.15 mg/kg of available phosphorus, resulting in soil with a pH of 5.6. The seedlings were grown in plastic flowerpots (width, 31 cm; length, 57 cm; height, 13.5 cm) that were rinsed with tap water, wiped with 75% alcohol, and then dried before use.
2.2. Methods
This study adopted a two-factor experimental design consisting of drought stress and inoculation treatments. Drought stress levels were established using three controlled water contents: low water (LW), moderate water (MW), and well watered (WW), corresponding to 35%, 55%, and 75% of field water capacity, respectively. Inoculation treatments comprised A. vagum (AV) and non- A. vagum (Non-AV) groups. Fifteen seedlings were planted in each pot and three pots were used in each treatment (i.e., 45 seedlings were used in each of the six treatments and 270 seedlings were used in total). Seedlings were inoculated by root irrigation with 50 mL of either a viable fungal solution (AV group) or a nonviable fungal solution (Non-AV group) that had been subjected to high-temperature sterilization (at 121 °C and 0.05 MPa for 2 h), which acted as the control. The root irrigation procedure was repeated 7 days later. All seedlings were watered with sterile water in 75% of field water capacity for 1 month. Seedlings were irrigated with 1/2 Hoagland’s nutrient solution once every 15 days. The study began on April 2, 2018 and ended on July 25, 2018. All experiments were conducted in a greenhouse at the Guizhou Institute of Biology. The temperature and humidity conditions in the shed were consistent with the local meteorological conditions in Guiyang City, Guizhou Province. The annual sunshine hours are about 1148.3 h, the annual average temperature is about 15.3 °C, the annual rainfall is about 1129.5 mm, and the annual average relative humidity is 77%.
2.3. Index Determination and Sample Analysis
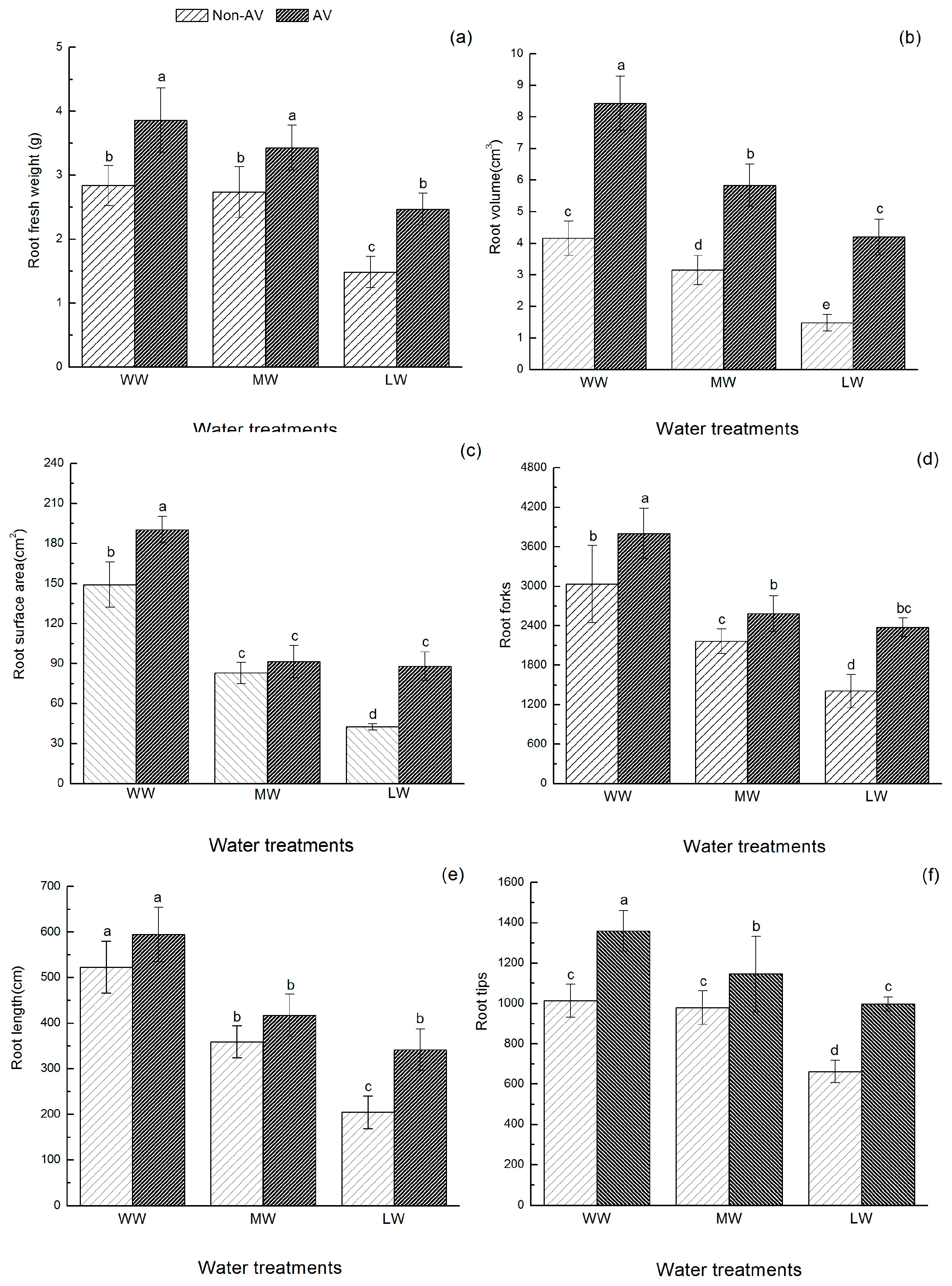
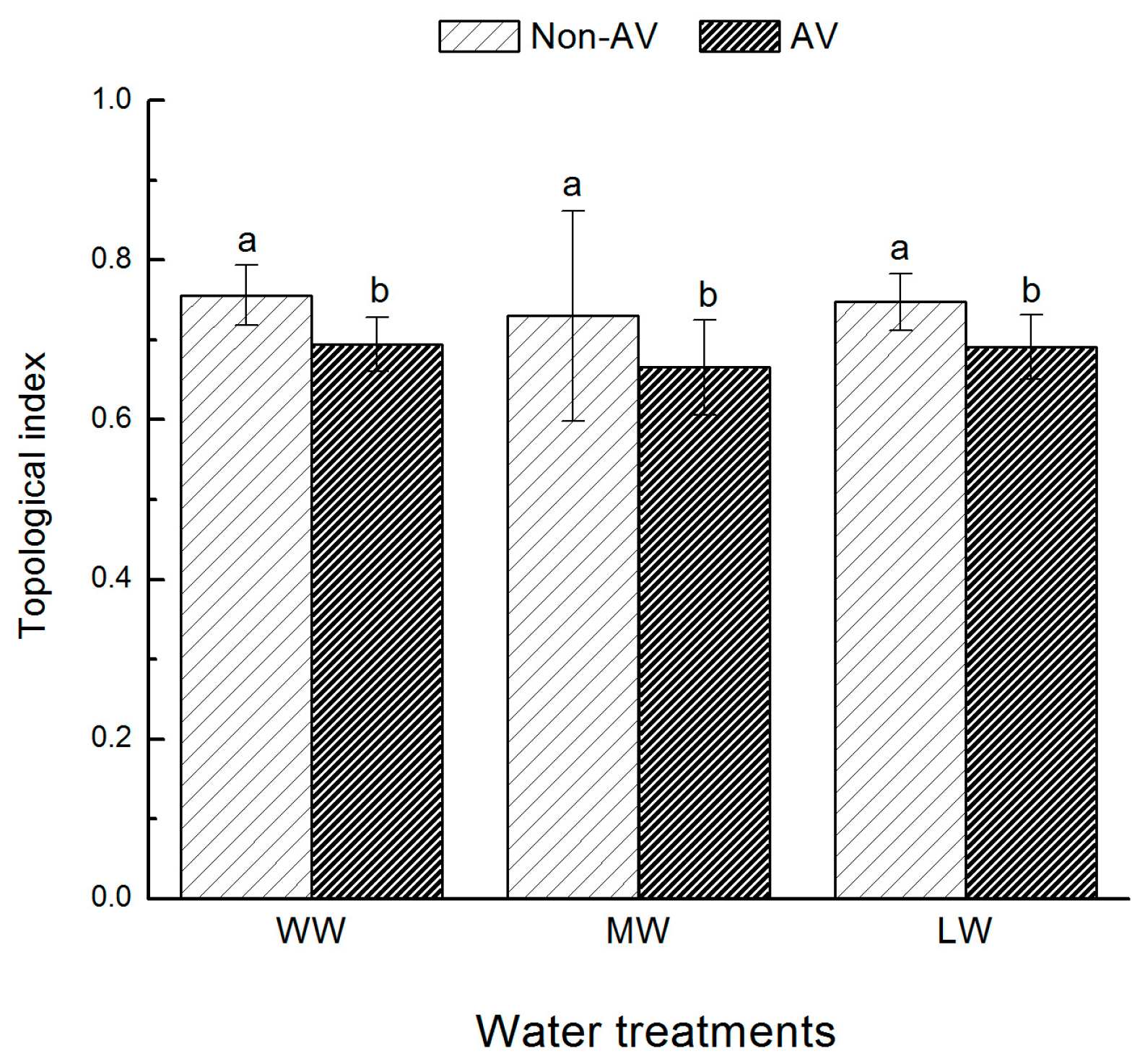
Five seedlings were randomly selected from each treatment. Roots were cleaned with distilled water and the fresh weight was determined using a 1/10,000-scale analytical balance. Roots were scanned (MicrotekScan Marker i800 plus, Microtek, Shanghai, China) and analyzed using Microtek Scan Wizard E2 software (Microtek, Shanghai, China) to determine total root length, root surface area, root volume, number of root tips, and root forks. The branching pattern of the root systems was analyzed using the topological index (TI) (i.e., TI = log A/log M, where A is the maximum number of paths of the root channel with maximum length, and M is the total number of all external root connections). Fitter et al. [
25] proposed two typical root topology structures: herringbone branching and dichotomous branching (
Figure 1, I: herringbone branching (A = 7, M = 7); II: dichotomous branching (A = 4, M = 8)). For typical herringbone branching, TI = 1, and for typical dichotomous branching, TI = 0.5. Studies have shown that the branch structures of plant roots are somewhere between herringbone architecture and dichotomous architecture, so the closer TI is to 1, the closer the roots are to a herringbone architecture. By contrast, the closer TI is to 0.5, the closer the roots are to a dichotomous architecture [
26].
GA, ZR, IAA, and ABA were measured using ELISA kits from Beijing Epsilon Biotechnology Co., Ltd. Fresh roots (0.5 g) were washed with 2 mL of phosphate buffer (pH 7.5) and then ground using a small amount of quartz sand in an ice-cold mortar. The mortar was rinsed with 2 mL of PBS and the slurry was collected in a 10 mL centrifuge tube and incubated for 4 h at 4 °C. After centrifugation at 4 °C and 10,000 rpm for 15 min, 2 mL of the supernatant was transferred to a centrifuge tube and GA, IAA, ZT, and ABA levels were determined using a Tecan Infinite Pro full-wavelength multifunction microplate reader.
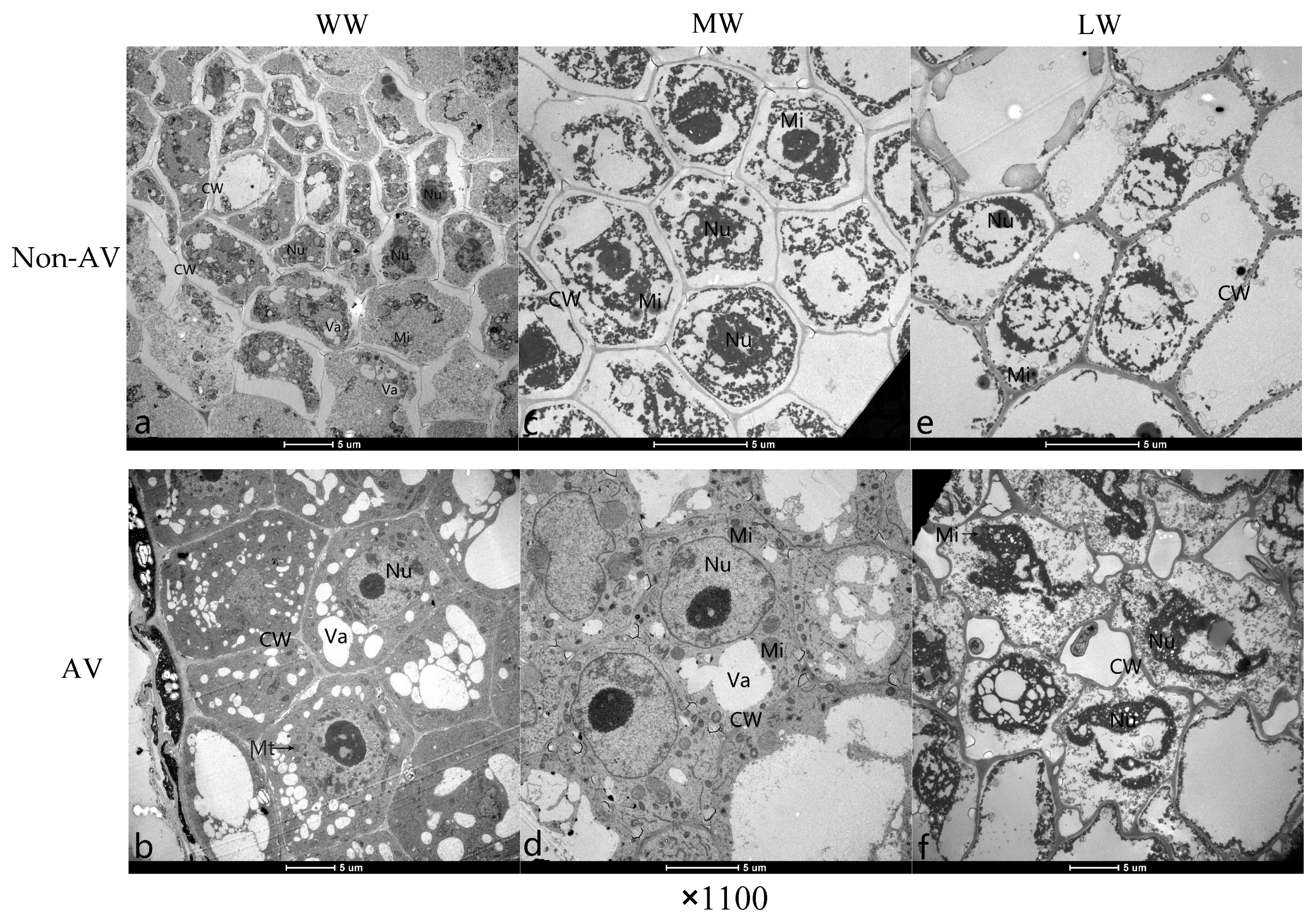
Root segments (0.2–0.3 cm) were cut from root tips with a sterile knife, quickly placed in a fixative (2.5% glutaraldehyde (0.1 mol·L−1 PBS, pH = 7.2)), and then fixed at 4 °C for 24 h in a refrigerator. Root segments were then rinsed with 0.1 mol·L−1 PBS, fixed with 1% citric acid solution for 4 h, rinsed with 0.1 mol·L−1 PBS, dehydrated in an ethanol series, embedded in Epon 812 epoxy resin, sectioned using an LKBV-type slicer, and stained with uranyl acetate and lead citrate. Sections were observed under a transmission electron microscope (JEM-1200).
2.4. Statistical Analysis
Data and statistical analyses were performed using SPSS 19.0 software. One-way analysis of variance (ANOVA), two-way ANOVA, the least-significant-difference (LSD) fitting method, and correlation analysis were used to determine the effects of DSE inoculation on seedling roots under different levels of drought stress. The significance level was set at α = 0.05. Before variance analyses, all measured data were tested to ensure that the data obeyed the assumptions of a normal distribution and homogeneity of variance. Related charts were prepared using Origin Pro8.
4. Discussion
Maintaining organelle structure is essential for maintaining the normal physiological function of cells. Under adverse conditions, the destruction of organelle structures eventually leads to a decrease or even loss of cell physiological function [
27,
28]. With the aggravation of drought stress, the ultrastructure of root cells of Non-AV seedlings became more damaged, the electron density of the endoplasm in root cells decreased significantly, the chromatin became highly condensed, the number of mitochondria decreased, the inner mitochondrial membrane ridges disappeared, the cell membrane was damaged, the cytoplasm leaked from the cell, and the cells appeared vacuolated. Under water stress conditions, seedlings inoculated with
A. vagum suffered significantly less damage in terms of root cell ultrastructure than the Non-AV group. Under the three water stress conditions, the root cell structure of AV seedlings remained intact; the cell membrane was clearly visible; the intracellular electron density was uniform; the nucleolus was obvious; and the mitochondria were clear, complete, and numerous. Under drought stress, ultrastructural changes are mainly caused by dehydration of the cytoplasm, which results in further damage, such as membrane damage, cytoplasmic leakage, and metabolic arrest. The colonization of the host by the DSE strain
A. vagum appeared to alleviate water loss in plants to a certain extent and, hence, reduced the degree of drought stress, thus protecting the integrity of plant cells and ensuring the normal physiological activities of seedlings. Our results confirmed that the symbiosis between the DSE and the host could improve the drought tolerance of the host at the cell level. Previous studies have suggested that the network of DSE hyphae that connect the root cells of the plant with the rhizosphere could help plants to maintain water content and transport nutrients to the plant under drought stress conditions [
29]. In addition, DSEs could also increase proline content and antioxidant enzyme activity in host plants [
30]. Proline and antioxidant enzymes play active roles in maintaining the osmotic pressure of cells and enhancing the ability of scavenging reactive oxygen species. An increase in proline and antioxidant enzyme levels might explain why the structural integrity of the root cells was maintained in seedlings inoculated with
A. vagum; however, the specific mechanism still needs to be further investigated.
Changes in root morphology and configuration are important effects caused by some microorganisms. Under the same degree of drought stress, the topological index of AV seedlings was smaller than that of Non-AV seedlings, and the trend of root development towards a forked branch structure was obvious. In addition, the root volume, root branch number, root tip number, and root fresh weight values of the AV group were significantly higher than those of Non-AV seedlings. These findings are similar to the results of a citrus drought resistance study reported by Zhang et al. [
26], which indicated that DSEs could promote root development and growth. Both the AV and Non-AV groups showed significantly lower levels of root growth at higher levels of drought stress. However, under low water conditions, the total root length of AV seedlings was significantly greater than that of Non-AV seedlings. A possible explanation for this finding is that the symbiotic relationship between
O. hosiei seedlings and
A. vagum enabled the plant to produce more fine roots under drought stress conditions, which promoted the uptake of water and nutrients by roots owing to the increase in the length of active fine roots and the number of root tips, thereby reducing the potential harm caused by drought conditions. This study supports previous findings that endophytic fungi could help the host to improve root morphology and configuration under drought stress conditions, thereby expanding the absorption range of roots and increasing the contact between roots and soil, which promotes the ability of plants to adapt to adverse stress.
Plant hormones are a group of natural organic compounds that play important roles in regulating plant root development under water stress conditions. Under drought stress conditions, plants rapidly accumulate and synthesize a large amount of ABA in roots and then transport ABA to the aboveground parts through the xylem in the transpiration stream to participate in leaf stomatal regulation, reduce stomatal conductance, reduce transpiration loss, and improve plant water retention capacity [
31,
32]. In addition, accumulation of auxin (IAA) in the root system reduces water use and modulates plant hydraulic properties to enable the expression of water-saving traits under drought [
33]. In the present study,
A. vagum inoculation significantly promoted the accumulation of ABA and IAA in
O. hosiei seedling roots. This suggests that
A. vagum could promote stomatal closure and reduce transpiration by inducing ABA accumulation in host roots under water stress conditions, and rapidly adjust root water status to enhance plant drought resistance. In addition to ABA,
A. vagum also affected the GA content levels in plants. The content of GA changed little with the aggravation of water stress; however, the GA levels of seedlings inoculated with
A. vagum were significantly lower than those of the Non-AV group. These results indicate that DSEs could coordinate the balance of growth-related hormones to promote plant growth and, hence, improve plant drought tolerance.
Previous research has shown that the ratio of growth-promoting hormones to stress-related hormones is often more important than the role of a single hormone in plants [
34]. Changes in hormone interactions under drought stress conditions have been demonstrated in many studies. In this study, we showed that water stress reduced the GA/ABA and ZR/IAA ratios of seedlings. Apart from the IAA/ABA ratio of DSE-inoculated seedlings, which was higher than that of noninoculated seedlings, the ratios of GA/ABA, ZR/ABA, and ZR/IAA of
A.-
vagum-inoculated seedlings were lower than those of noninoculated seedlings. This suggests that
A. vagum may coordinate the balance of host hormones through adjusting hormone ratios after symbiosis with the host, slowing down plant growth and development to reduce the excessive consumption of water, maintaining normal cell swelling pressure to maintain normal water balance, promoting stomatal closure to reduce water transpiration, and increasing cell solute concentration to enhance plant water absorption capacity, thereby enhancing plant drought resistance. However, how DSEs induce the production of hormones and regulate the balance of hormones in host plants needs to be further elucidated from a molecular perspective.
Root morphological development is closely related to plant hormone levels [
11]. López-Coria et al. [
35] showed that root changes may be attributable to auxin signaling, which promotes division and elongation of epidermal cells [
36]. Xiong et al. [
5] found that ABA is also involved in crop root development under drought stress and is an essential hormone for maintaining root growth under drought conditions. Our results showed that the hormone content levels and ratios of hormones in roots as well as the root morphological structure of seedlings inoculated with
A. vagum were different to those of the noninoculated control seedlings. The present results suggest that DSEs might influence root morphology and structure by regulating hormone content levels and the ratio of hormones in roots in order to better adapt the host to a drought stress environment. DSEs not only help the host root system improve drought resistance but also help the root system adapt to the drought stress environment by modulating the morphological structure of the root, showing a positive response to the drought stress.








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}