The Effect of UV-B Radiation on Bufo arenarum Embryos Survival and Superoxide Dismutase Activity

Abstract
:Introduction
Materials and Methods
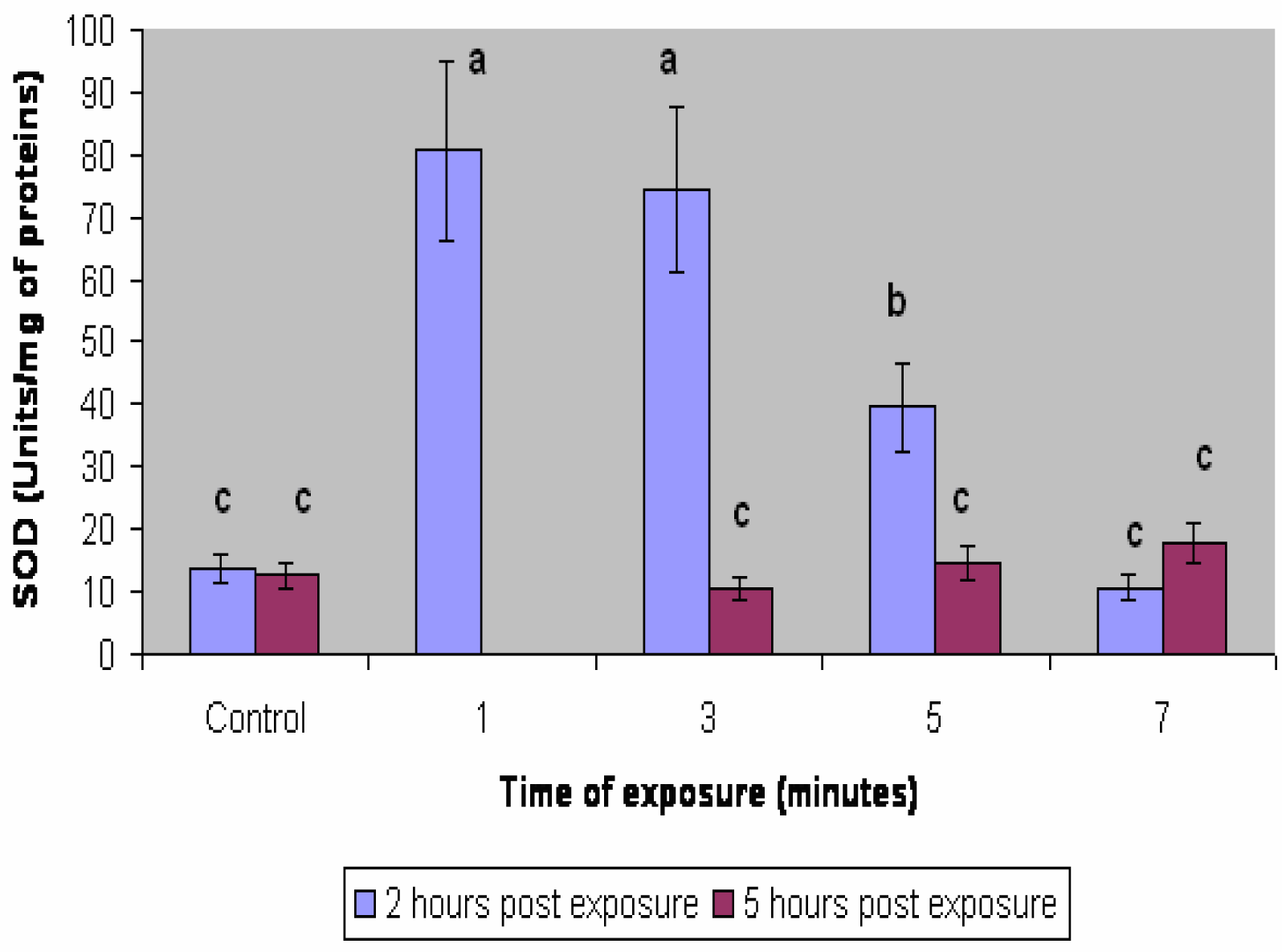
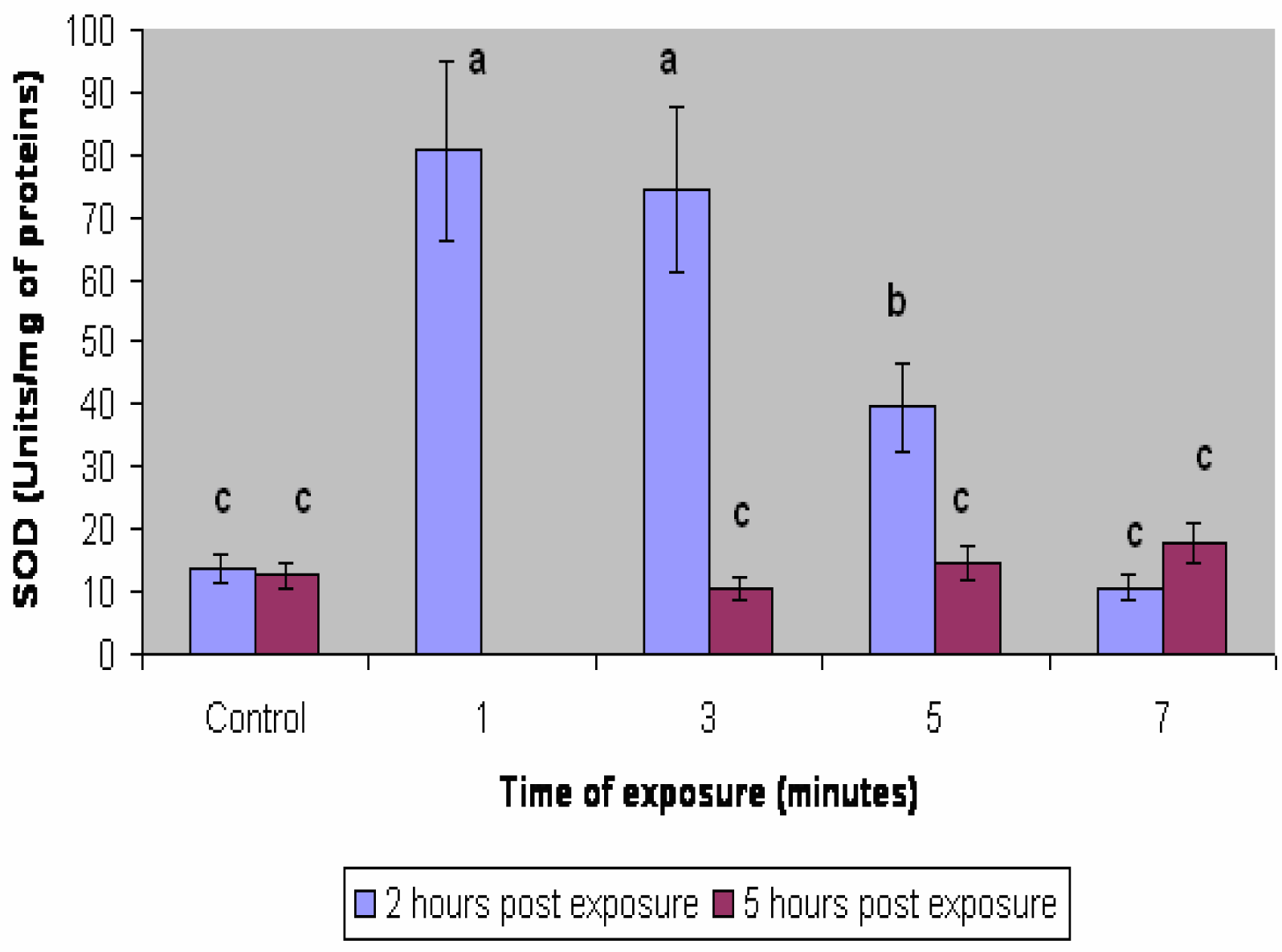
Results
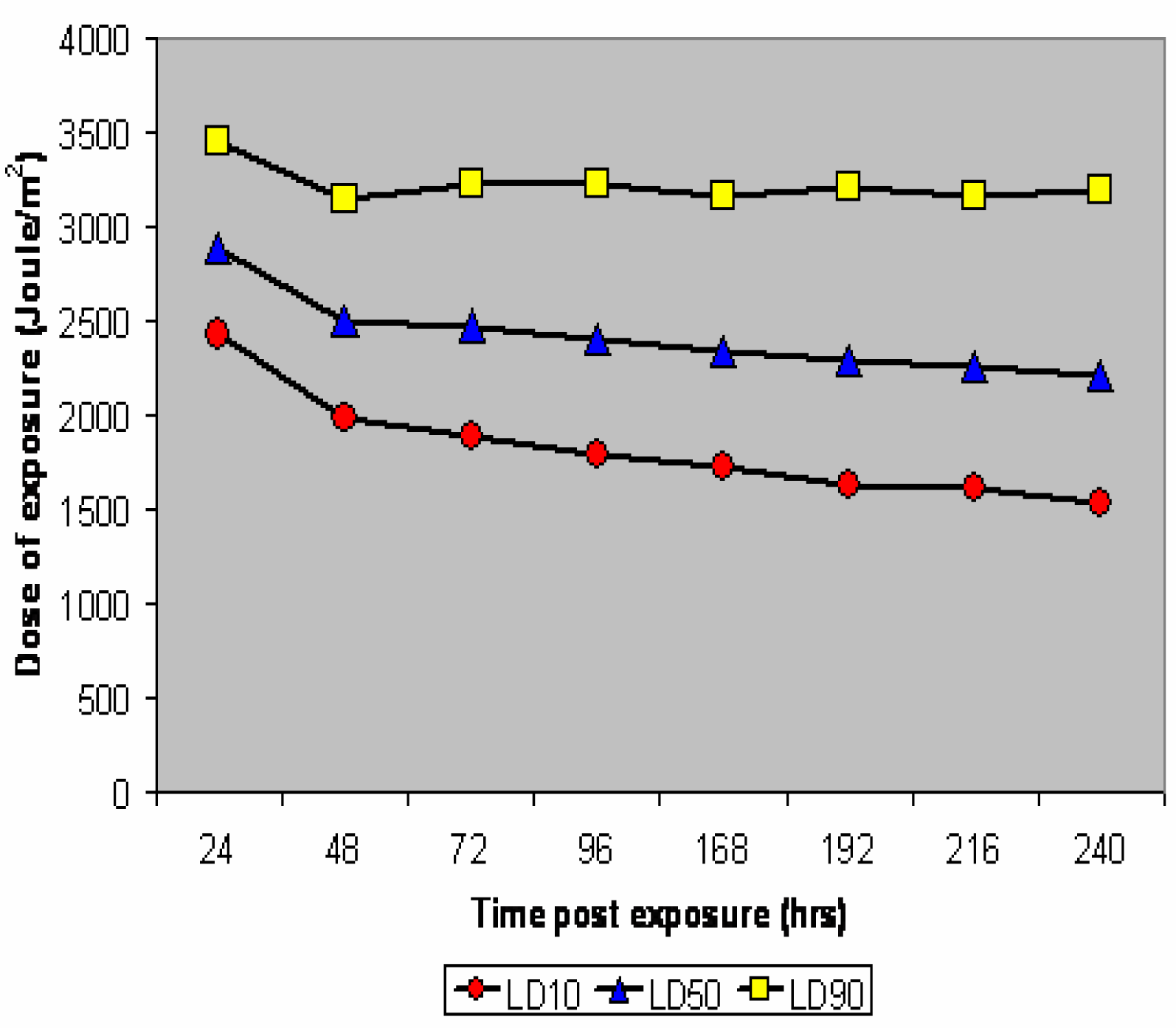
UV-B Toxicity and TOP Curves
Discussion
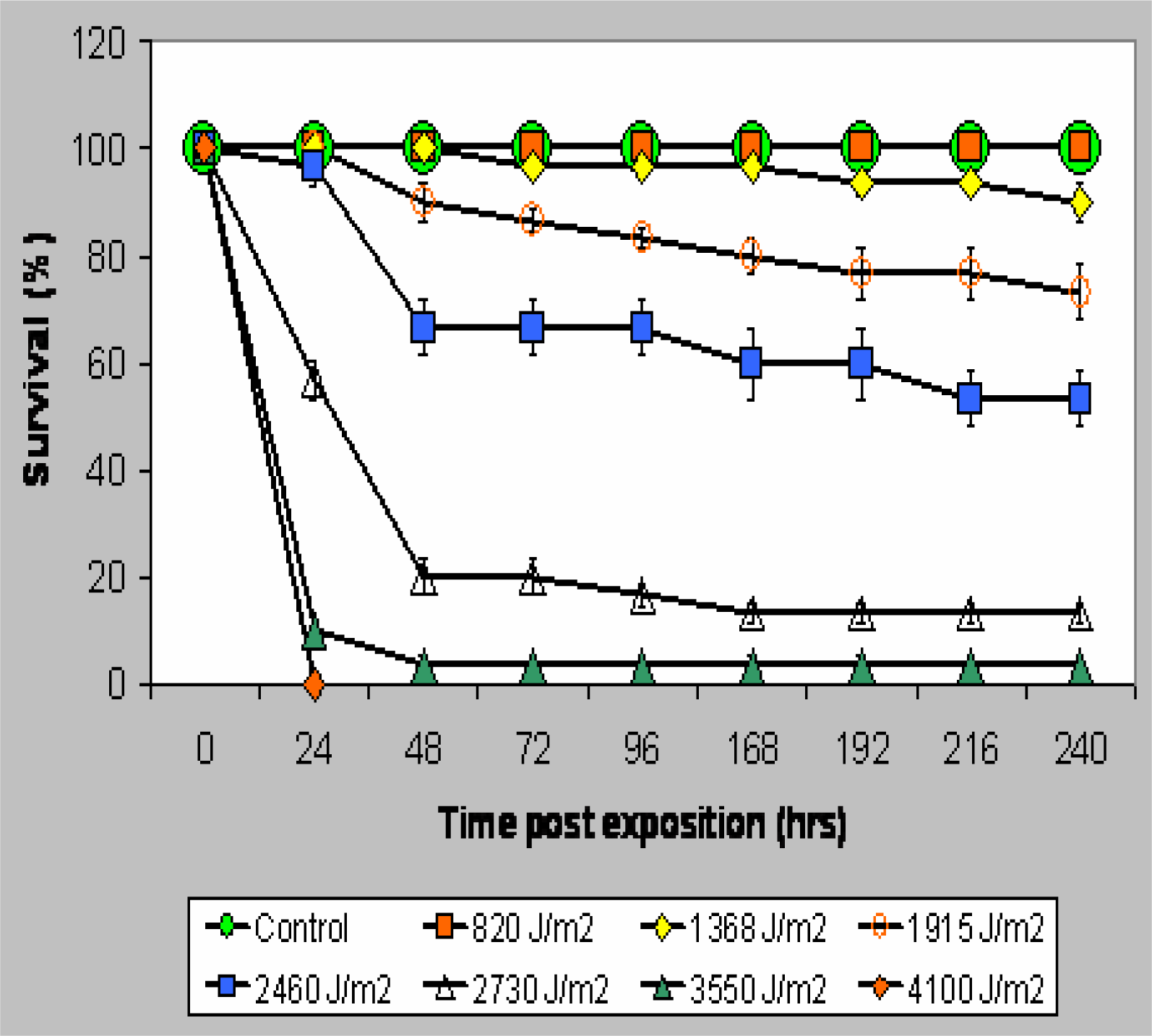

{kind=link}
{kind=link}
{kind=link}
| Time post exposition(hr) | LE10 | LE50 | LE90 |
|---|---|---|---|
| 24 | 2424 (2244 – 2553) | 2891 (2768 – 3030) | 3462 (3249 – 3778) |
| 48 | 1979 (1761 – 2124) | 2512 (2359 – 2621) | 3134 (2935 – 3478) |
| 72 | 1892 (1384 – 2151) | 2471 (2184 – 2749) | 3227 (2875 – 4210) |
| 96 | 1783 (1559 – 1946) | 2397 (2250 – 2544) | 3224 (2989 – 3609) |
| 168 | 1728 (1504 – 1889) | 2334 (2184 – 2479) | 3153 (2924 – 3230) |
| 192 | 1630 (1400 – 1796) | 2285 (2127 – 2438) | 3205 (2954 – 3615) |
| 216 | 1611 (1384 – 1777) | 2255 (2097 – 2405) | 3156 (2910 – 3571) |
| 240 | 529 (1299 – 1701) | 2206 (2045 – 2364) | 3191 (2927 – 3617) |
Acknowledgments
References
- Lesser, MP; Farrell, JH; Walker, CW. Oxidative stress, DNA damage and p53 expression in the larvae of Atlantic cod (Gadus morhua) exposed to ultraviolet (290–400 nm) radiation. J. Exp. Biol 2001, 204, 157–164. [Google Scholar]
- Barber, AA; Bernheim, F. Lipid peroxidation: its measurement, occurrence, and significance in animal tissues. Adv. Gerontol. Res 1967, 2, 355–403. [Google Scholar]
- Moody, CS; Hassan, HM. Mutagenicity of oxygen free radicals. Proc Natl. Acad. Sci. U S A 1982, 79, 2855–9285. [Google Scholar]
- Del Maestro, R; Thaw, HH; Bjork, J; Planker, M; Arfors, KE. Free radicals as mediators of tissue injury. Acta. Physiol. Scand. Suppl 1980, 492, 43–57. [Google Scholar]
- McCord, JM. Free radicals and inflammation: protection of synovial fluid by superoxide dismutase. Science 1974, 185, 529–531. [Google Scholar]
- International Programme on Chemical Safety. Environmental Health Criteria 160. Ultraviolet Radiation. World Health Organization 1994. [Google Scholar]
- Desai, ID; Sawant, PL; Tappel, AL. Peroxidative and radiation damage to isolated lysosomes. Biochim. Biophys. Acta 1964, 86, 277–285. [Google Scholar]
- Putvinsky, AV; Sokolov, AI; Roshchupkin, DI; Vladimirov, YA. Electric breakdown of bilayer phospholipid membranes under ultraviolet irradiation-induced lipid peroxidation. FEBS Lett 1979, 106, 53–55. [Google Scholar]
- Azizova, AO; Islomov, AI; Roshchupkin, DI; Predvoditelev, DA; Remizov, AN; Vladimirov, YA. Free radicals formed on ultraviolet irradiation of the lipids of biological membrane. Biophysics 1980, 24, 407–414. [Google Scholar]
- Sasaki, H; Akamatsu, H; Horio, T. Effects of a single exposure to UV-B radiation on the activities and protein levels of copper-zinc and manganese superoxide dismutase in cultured human keratinocytes. Photochem. Photobiol 1997, 65, 707–713. [Google Scholar]
- D’Eramo, JL; Fridman, O; Herkovits, J. Respuesta antioxidante en embriones de Bufo arenarum expuestos a dosis letales de radiación ultravioleta (UV-b). In Toxicología y Química Ambiental. Con tribuciones Para un Desarrollo Sustentable; Society of Environmental Toxicology and Chemistry: Pensacola, FL. USA; Herkovits, J, Ed.; 2003; pp. 147–149. [Google Scholar]
- Blaustein, AR; Wake, DB. Declining amphibian populations: A global phenomenon? Trends Ecol. Evol 1990, 5, 203–204. [Google Scholar]
- Tietge, JE; Diamond, SA; Ankley, GT; DeFoe, DL; Holcombe, GW; Jensen, KM; Degitz, SJ; Elonen, GE; Hammer, E. Ambient solar UV radiation causes mortality in larvae of three species of Rana under controlled exposure conditions. Photochem. Photobiol 2001, 74, 261–268. [Google Scholar]
- Blaustein, AR; Hoffman, PD; Hokit, DG; Kiesecker, JM; Walls, SC; Hays, JB. UV repair and resistance to solar UV-B in amphibian eggs: a link to population declines? Proc. Natl. Acad. Sci. U S A 1994, 91, 1791–1795. [Google Scholar]
- Crump, D; Berril, M; Coulson, D; Lean, DRS; McGillvray, L; Smith, A. Sensitivity of amphibian embryos, tadpoles and larvae to enhanced UV-B radiation in natural pond conditions. Can. J. Zool 1999, 77, 1956–1966. [Google Scholar]
- Perez-Coll, CS; Herkovits, J. Stage dependent uptake of cadmium by Bufo arenarum embryos. Bull. Environ. Contam. Toxicol 1996, 56, 663–669. [Google Scholar]
- Herkovits, J; Herkovits, FD; Pérez-Coll, CS. Lead Toxicity TOP and Time Dependent Lead-Zinc Antagonism in Bufo arenarum Embryos. Abstract Book of the 17th Annual Meeting of the Society of Environmental Toxicology and Chemistry, Washington, DC; 17–21 November 1996; p. 141. [Google Scholar]
- Perez-Coll, C; Herkovits, J. Lethal and teratogenic effects of naringenin evaluated by mean of an amphibian embryo toxicity test (AMPHITOX). Food Chem. Toxicol 2004, 42, 299–306. [Google Scholar]
- Herkovits, J; Pérez-Coll, CS. AMPHITOX: A Standardized Set of Toxicity Tests employing Amphibian Embryos. It’s Potential for Customized Hazard Assessment. In Multiple Stressor Effects in Relation to Declining Amphibian Populations; ASTM; Linder, G, Krest, S, Sparling, D, Little, EE, Eds.; ASTM International, 2003; pp. 46–60. [Google Scholar]
- Herkovits, J; Herkovits, FD; Pérez-Coll, CS. Identification of aluminium toxicity and aluminium-zinc interaction in amphibian Bufo arenarum embryos. Environ. Sci 1997, 5, 57–64. [Google Scholar]
- Herkovits, J; Helguero, LA. Copper toxicity and copper-zinc interactions in amphibian embryos. Sci. Total Environ 1998, 221, 1–10. [Google Scholar]
- Herkovits, J; Perez-Coll, C. Bioensayos para test de toxicidad con embriones de anfibio “ANFITOX”. basado en Bufo arenarum. Test Agudo (ANFIAGU), Crónico corto (ANFICOR), Crónico (ANFICRO) y de Estadios Tempranos del Desarrollo (ANFIEMB). Ingeniería Sanitaria y Ambiental 1999, 42, 24–30, 43, 50–55.. [Google Scholar]
- Del Conte, E; Sirlin, L. The first stages of Bufo arenarum development (in spanish). Acta Zool. Lilloana 1951, 12, 495–499. [Google Scholar]
- Lowry, OH; Rosebrough, HJ; Farr, AI; Randall, RJ. Protein measurement with the Folin phenol reagent. J. Biol. Chem 1951, 193, 265–275. [Google Scholar]
- Herkovits, J; Cardellini, P; Pavanati, C; Perez’Coll, CS. Susceptibility of early life stages of Xenopus laevis to cadmium. Environ. Toxicol. Chem 1977, 16, 312–316. [Google Scholar]
- Pahkala, M; Laurila, A; Merila, J. Effects of ultraviolet-B radiation on behaviour and growth of three species of amphibian larvae. Chemosphere 2003, 51, 197–204. [Google Scholar]
- Blaustein, AR; Belden, LK. Amphibian defenses against ultraviolet-B radiation. Evol. Dev 2003, 5, 89–97. [Google Scholar]
- Alvarez, S; Boveris, A. Antioxidant adaptive response in human blood mononuclear cells exposed to UV-B. J. Photochem. Photobiol. B 1997, 38, 152–157. [Google Scholar]
- Iizawa, O; Kato, T; Tagami, H; Akamatsu, H; Niwa, Y. Long-term follow-up study of changes in lipid peroxide levels and the activity of superoxide dismutase, catalase and glutathione peroxidase in mouse skin after acute and chronic UV irradiation. Arch. Dermatol. Res 1994, 286, 47–52. [Google Scholar]
- Shindo, Y; Witt, E; Han, D; Packer, L. Dose-response effects of acute ultraviolet irradiation on antioxidants and molecular markers of oxidation in murine epidermis and dermis. J. Invest. Dermatol 1994, 102, 470–475. [Google Scholar]
© 2006 by the authors; licensee MDPI, Basel, Switzerland This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Herkovits, J.; D´Eramo, J.L.; Fridman, O. The Effect of UV-B Radiation on Bufo arenarum Embryos Survival and Superoxide Dismutase Activity. Int. J. Environ. Res. Public Health 2006, 3, 43-47. https://doi.org/10.3390/ijerph2006030006
Herkovits J, D´Eramo JL, Fridman O. The Effect of UV-B Radiation on Bufo arenarum Embryos Survival and Superoxide Dismutase Activity. International Journal of Environmental Research and Public Health. 2006; 3(1):43-47. https://doi.org/10.3390/ijerph2006030006
Chicago/Turabian StyleHerkovits, J., J. L. D´Eramo, and O. Fridman. 2006. "The Effect of UV-B Radiation on Bufo arenarum Embryos Survival and Superoxide Dismutase Activity" International Journal of Environmental Research and Public Health 3, no. 1: 43-47. https://doi.org/10.3390/ijerph2006030006
APA StyleHerkovits, J., D´Eramo, J. L., & Fridman, O. (2006). The Effect of UV-B Radiation on Bufo arenarum Embryos Survival and Superoxide Dismutase Activity. International Journal of Environmental Research and Public Health, 3(1), 43-47. https://doi.org/10.3390/ijerph2006030006