
Scrutinizing the Impact of Alternating Electromagnetic Fields on Molecular Features of the Model Plant Arabidopsis thaliana

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material and Growth Conditions
2.2. Exposure to Electromagnetic Field Conditions
2.3. Visualization of Phenotype
2.4. Determination of Photosynthetic Parameters
2.5. Isolation of Total RNA and Sodium Acetate Precipitation for Gene Expression Analysis
2.5.1. RNA Sequencing
2.5.2. Transcriptome-Based Gene Identification and Quantification by qRT-PCR
2.5.3. Gene Onotology (GO) Analysis
2.6. Metabolite Profiling
2.7. Statistical Analysis
3. Results
3.1. The Variation of the Phenotype
3.2. The Response of Photosynthetic Parameters to Electromagnetic Fields
3.3. Effect of Electromagnetic Fields on the Leaf Transcriptome
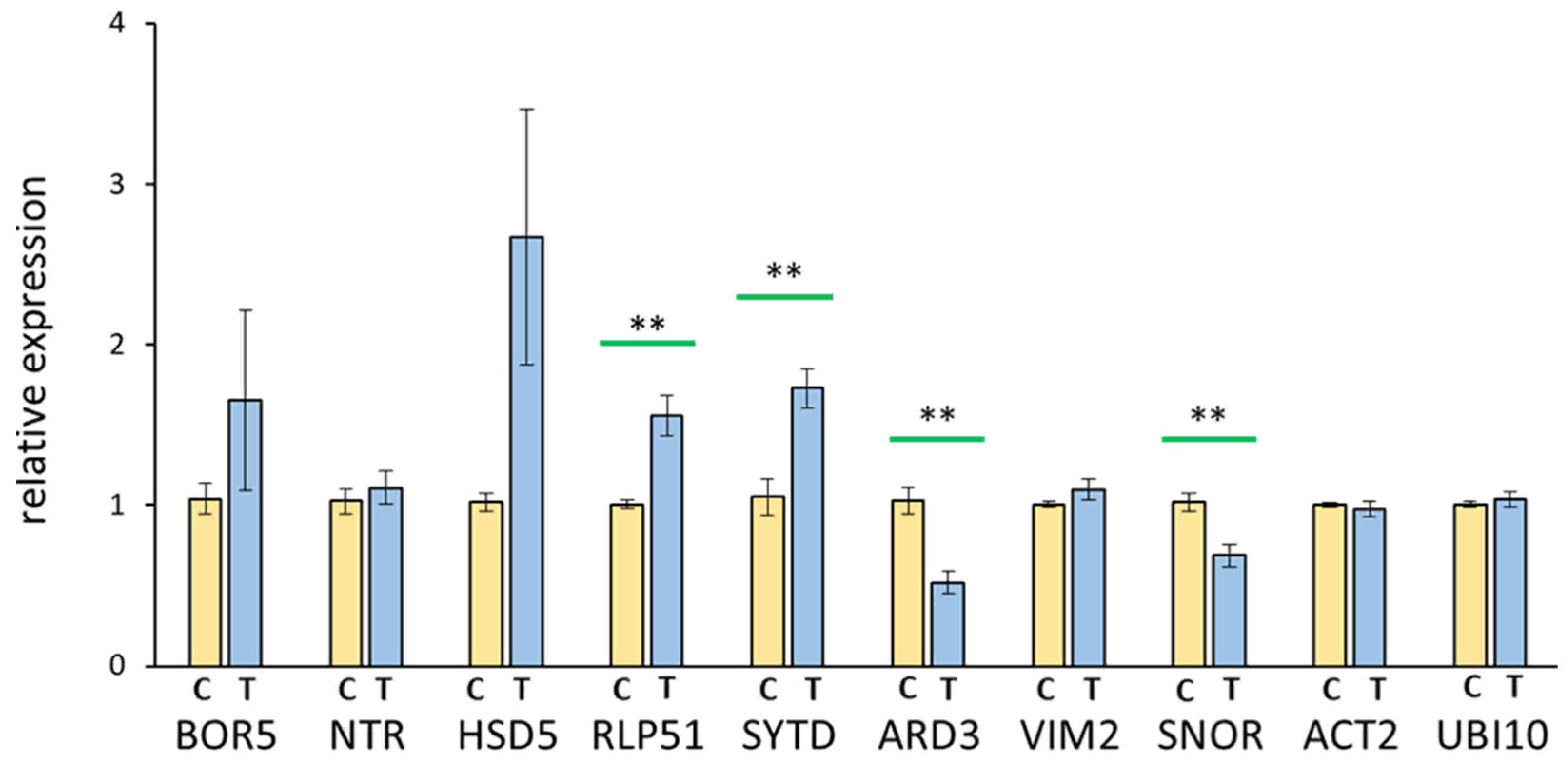
3.4. Real-Time Quantitative PCR
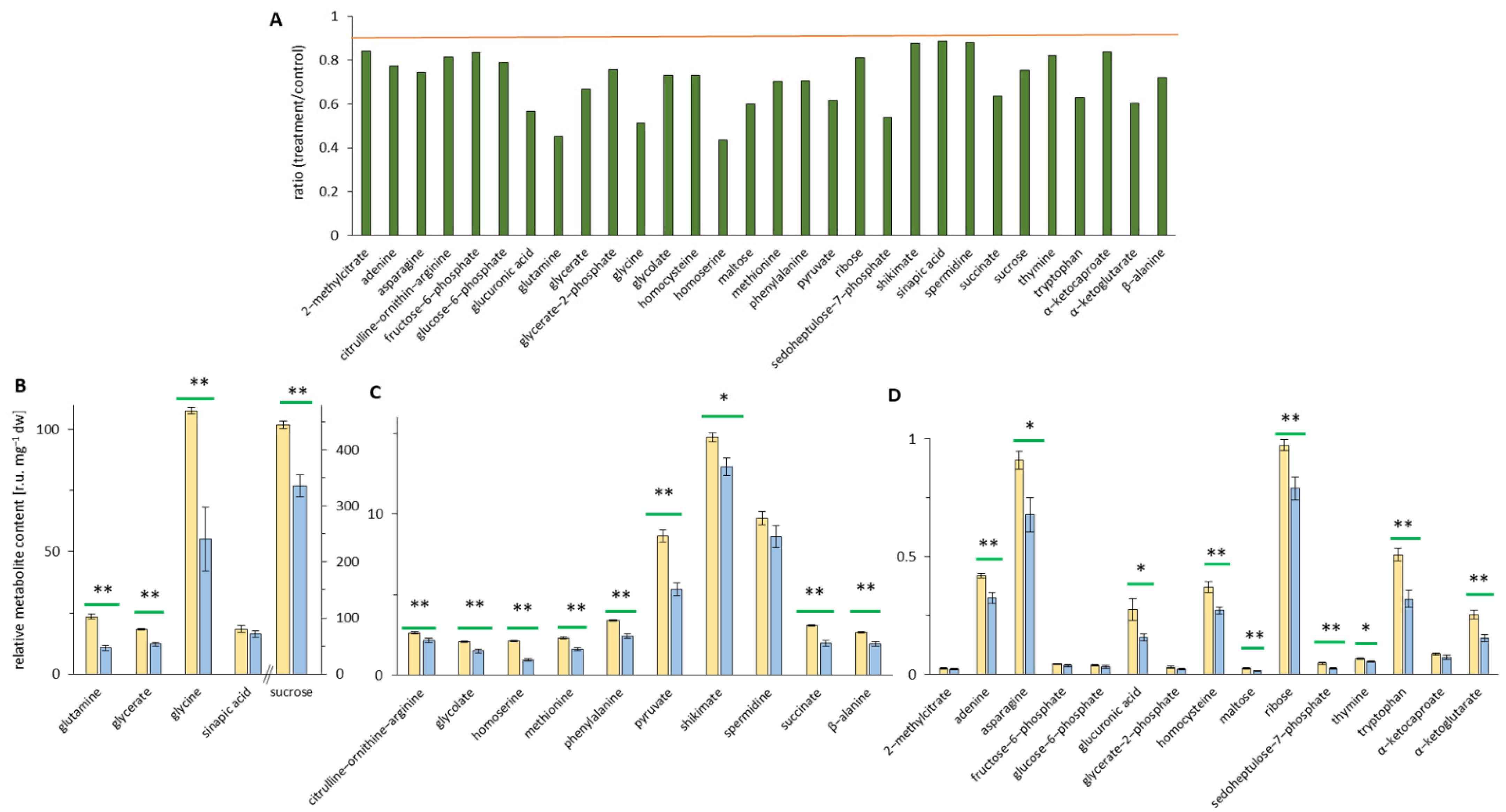
3.5. EMF Effects on Leaf Metabolome
4. Discussion
4.1. Changes in Photosynthetic Parameters
4.2. Variation in Transcript Amounts May Hide a More Severe Reorganization of the Transcriptome
4.3. EMF-Induced Metabolic Alterations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Minorsky, P.V. Do geomagnetic variations affect plant function? J. Atmos. Sol.-Terr. Phys. 2007, 69, 1770–1774. [Google Scholar] [CrossRef]
- Drzewiecka, E.M.; Kozlowska, W.; Paukszto, L.; Zmijewska, A.; Wydorski, P.J.; Jastrzebski, J.P.; Franczak, A. Effect of the Electromagnetic Field (EMF) Radiation on Transcriptomic Profile of Pig Myometrium during the Peri-Implantation Period-An In Vitro Study. Int. J. Mol. Sci. 2021, 22, 7322. [Google Scholar] [CrossRef] [PubMed]
- Guidelines for limiting exposure to time-varying electric and magnetic fields (1 Hz to 100 kHz). Health Phys. 2010, 99, 818–836. [CrossRef] [PubMed]
- Balmori, A. Electromagnetic pollution from phone masts. Effects on wildlife. Pathophysiology 2009, 16, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Shckorbatov, Y. The Main Approaches of Studying the Mechanisms of Action of Artificial Electromagnetic Fields on Cell. J. Elec. Electron Syst. 2014, 3, 1000123. [Google Scholar] [CrossRef] [Green Version]
- Gajšek, P.; Ravazzani, P.; Grellier, J.; Samaras, T.; Bakos, J.; Thuróczy, G. Review of Studies Concerning Electromagnetic Field (EMF) Exposure Assessment in Europe: Low Frequency Fields (50 Hz–100 kHz). Int. J. Environ. Res. Public Health 2016, 13, 875. [Google Scholar] [CrossRef] [Green Version]
- Levitt, B.B.; Lai, H.C.; Manville, A.M. Effects of non-ionizing electromagnetic fields on flora and fauna, part 1. Rising ambient EMF levels in the environment. Rev. Environ. Health 2021, 37(1), 81–122. [Google Scholar] [CrossRef]
- Vian, A.; Davies, E.; Gendraud, M.; Bonnet, P. Plant Responses to High Frequency Electromagnetic Fields. Biomed Res. Int. 2016, 2016, 1830262. [Google Scholar] [CrossRef] [Green Version]
- Vian, A.; Faure, C.; Girard, S.; Davies, E.; Hallé, F.; Bonnet, P.; Ledoigt, G.; Paladian, F. Plants Respond to GSM-Like Radiation. Plant Signal. Behav. 2007, 2, 522–524. [Google Scholar] [CrossRef] [Green Version]
- Efthimiadou, A.; Katsenios, N.; Karkanis, A.; Papastylianou, P.; Triantafyllidis, V.; Travlos, I.; Bilalis, D.J. Effects of presowing pulsed electromagnetic treatment of tomato seed on growth, yield, and lycopene content. Sci. World J. 2014, 2014, 369745. [Google Scholar] [CrossRef]
- Mahajan, T.S.; Pandey, O.P. Magnetic-time model at off-season germination. Int. Agrophys. 2014, 28, 57–62. [Google Scholar] [CrossRef]
- Menegatti, R.D.; de Oliveira, L.O.; Da Costa, Á.; Braga, E.; Bianchi, V.J. Magnetic field and gibberelic acid as pre-germination treatments of passion fruit seeds. Rev. Cien. Agr. 2019, 17, 15–22. [Google Scholar] [CrossRef]
- Da Silva, J.A.T.; Dobránszki, J. Magnetic fields: How is plant growth and development impacted? Protoplasma 2016, 253, 231–248. [Google Scholar] [CrossRef]
- Belyavskaya, N.A. Biological effects due to weak magnetic field on plants. Adv. Space Res. 2004, 34, 1566–1574. [Google Scholar] [CrossRef]
- Shine, M.B.; Guruprasad, K.N.; Anand, A. Enhancement of germination, growth, and photosynthesis in soybean by pre-treatment of seeds with magnetic field. Bioelectromagnetics 2011, 32, 474–484. [Google Scholar] [CrossRef]
- Dhawi, F. Why Magnetic Fields are Used to Enhance a Plant’s Growth and Productivity? ARRB 2014, 4, 886–896. [Google Scholar] [CrossRef]
- Maffei, M.E. Magnetic field effects on plant growth, development, and evolution. Front. Plant Sci. 2014, 5, 445. [Google Scholar] [CrossRef] [Green Version]
- Martinez, E.; Florez, M.; Carbonell, M.V. Stimulatory Effect of the Magnetic Treatment on the Germination of Cereal Seeds. IJEAB 2017, 2, 375–381. [Google Scholar] [CrossRef]
- Danho, S.; Schoellhorn, W.; Aclan, M. Innovative technical implementation of the Schumann resonances and its influence on organisms and biological cells. IOP Conf. Ser.: Mater. Sci. Eng. 2019, 564, 12081. [Google Scholar] [CrossRef]
- Reina, F.G.; Pascual, L.A. Influence of a stationary magnetic field on water relations in lettuce seeds. Part I: Theoretical considerations. Bioelectromagnetics 2001, 22, 589–595. [Google Scholar] [CrossRef]
- Sukhov, V.; Sukhova, E.; Sinitsyna, Y.; Gromova, E.; Mshenskaya, N.; Ryabkova, A.; Ilin, N.; Vodeneev, V.; Mareev, E.; Price, C. Influence of Magnetic Field with Schumann Resonance Frequencies on Photosynthetic Light Reactions in Wheat and Pea. Cells 2021, 10, 149. [Google Scholar] [CrossRef] [PubMed]
- Anand, A.; Kumari, A.; Thakur, M.; Koul, A. Hydrogen peroxide signaling integrates with phytohormones during the germination of magnetoprimed tomato seeds. Sci. Rep. 2019, 9, 8814. [Google Scholar] [CrossRef] [PubMed]
- Nyakane, N.E.; Markus, E.D.; Sedibe, M.M. The Effects of Magnetic Fields on Plants Growth: A Comprehensive Review. Int. J. Food Eng. 2019, 5, 79–87. [Google Scholar] [CrossRef]
- Da, A.; Teixeira Silva, J.; Dobránszki, J. How do magnetic fields affect plants in vitro? In Vitro Cell. Dev. Biol.-Plant 2015, 51, 233–240. [Google Scholar] [CrossRef]
- Grellier, J.; Ravazzani, P.; Cardis, E. Potential health impacts of residential exposures to extremely low frequency magnetic fields in Europe. Environ. Int. 2014, 62, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rawlins, J.C. Introduction to Alternating Current. In Basic AC Circuits; Elsevier: Amsterdam, The Netherlands, 2000; pp. 1–28. ISBN 9780750671736. [Google Scholar]
- Klughammer, C.; Schreiber, U. Deconvolution of ferredoxin, plastocyanin, and P700 transmittance changes in intact leaves with a new type of kinetic LED array spectrophotometer. Photosynth. Res. 2016, 128, 195–214. [Google Scholar] [CrossRef] [Green Version]
- Balti, H.; Abassi, M.; Dietz, K.-J.; Kumar, V. Differences in Ionic, Enzymatic, and Photosynthetic Features Characterize Distinct Salt Tolerance in Eucalyptus Species. Plants 2021, 10, 1401. [Google Scholar] [CrossRef]
- Jiang, H.; Lei, R.; Ding, S.-W.; Zhu, S. Skewer: A fast and accurate adapter trimmer for next-generation sequencing paired-end reads. BMC Bioinform. 2014, 15, 182. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [Green Version]
- R: A Language and Environment for Statistical Computing. Available online: https://www.gbif.org/tool/81287/r-a-language-and-environment-for-statistical-computing (accessed on 10 February 2015).
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Andrew, S. FastQC A Quality Control Tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 24 November 2010).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009; ISBN 9780387981413. [Google Scholar]
- Galili, T. dendextend: An R package for visualizing, adjusting and comparing trees of hierarchical clustering. Bioinformatics 2015, 31, 3718–3720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Vandesompele, J.; de Preter, K.; Pattyn, F.; Poppe, B.; van Roy, N.; de Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mi, H.; Muruganujan, A.; Huang, X.; Ebert, D.; Mills, C.; Guo, X.; Thomas, P.D. Protocol Update for large-scale genome and gene function analysis with the PANTHER classification system (v.14.0). Nat. Protoc. 2019, 14, 703–721. [Google Scholar] [CrossRef]
- Mi, H.; Ebert, D.; Muruganujan, A.; Mills, C.; Albou, L.-P.; Mushayamaha, T.; Thomas, P.D. PANTHER version 16: A revised family classification, tree-based classification tool, enhancer regions and extensive API. Nucleic Acids Res. 2021, 49, D394–D403. [Google Scholar] [CrossRef]
- Kind, T.; Wohlgemuth, G.; Lee, D.Y.; Lu, Y.; Palazoglu, M.; Shahbaz, S.; Fiehn, O. FiehnLib: Mass spectral and retention index libraries for metabolomics based on quadrupole and time-of-flight gas chromatography/mass spectrometry. Anal. Chem. 2009, 81, 10038–10048. [Google Scholar] [CrossRef] [Green Version]
- Plassmeier, J.; Barsch, A.; Persicke, M.; Niehaus, K.; Kalinowski, J. Investigation of central carbon metabolism and the 2-methylcitrate cycle in Corynebacterium glutamicum by metabolic profiling using gas chromatography-mass spectrometry. J. Biotechnol. 2007, 130, 354–363. [Google Scholar] [CrossRef]
- Larkindale, J.; Hall, J.D.; Knight, M.R.; Vierling, E. Heat stress phenotypes of Arabidopsis mutants implicate multiple signaling pathways in the acquisition of thermotolerance. Plant Physiol. 2005, 138, 882–897. [Google Scholar] [CrossRef] [Green Version]
- Liu, Q.; Kasuga, M.; Sakuma, Y.; Abe, H.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Two transcription factors, DREB1 and DREB2, with an EREBP/AP2 DNA binding domain separate two cellular signal transduction pathways in drought- and low-temperature-responsive gene expression, respectively, in Arabidopsis. Plant Cell 1998, 10, 1391–1406. [Google Scholar] [CrossRef] [Green Version]
- Parker, J.E.; Holub, E.B.; Frost, L.N.; Falk, A.; Gunn, N.D.; Daniels, M.J. Characterization of eds1, a mutation in Arabidopsis suppressing resistance to Peronospora parasitica specified by several different RPP genes. Plant Cell 1996, 8, 2033–2046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klevanik, A. Magnetic-field effects on primary reactions in Photosystem I. Biochim. Biophys. Acta (BBA)—Bioenerg. 1996, 1275, 237–243. [Google Scholar] [CrossRef] [Green Version]
- Niklas, J.; Brahmachari, U.; Utschig, L.M.; Poluektov, O.G. D-Band EPR and ENDOR Spectroscopy of 15N-Labeled Photosystem I. Appl. Magn. Reson. 2021, 53, 1–19. [Google Scholar] [CrossRef]
- Pfannschmidt, T.; Yang, C. The hidden function of photosynthesis: A sensing system for environmental conditions that regulates plant acclimation responses. Protoplasma 2012, 249 (Suppl. 2), S125–S136. [Google Scholar] [CrossRef]
- Unal, D.; García-Caparrós, P.; Kumar, V.; Dietz, K.-J. Chloroplast-associated molecular patterns as concept for fine-tuned operational retrograde signalling. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2020, 375, 20190443. [Google Scholar] [CrossRef]
- Kilian, J.; Whitehead, D.; Horak, J.; Wanke, D.; Weinl, S.; Batistic, O.; D’Angelo, C.; Bornberg-Bauer, E.; Kudla, J.; Harter, K. The AtGenExpress global stress expression data set: Protocols, evaluation and model data analysis of UV-B light, drought and cold stress responses. Plant J. 2007, 50, 347–363. [Google Scholar] [CrossRef]
- Coolen, S.; Proietti, S.; Hickman, R.; Davila Olivas, N.H.; Huang, P.-P.; van Verk, M.C.; van Pelt, J.A.; Wittenberg, A.H.J.; de Vos, M.; Prins, M.; et al. Transcriptome dynamics of Arabidopsis during sequential biotic and abiotic stresses. Plant J. 2016, 86, 249–267. [Google Scholar] [CrossRef] [Green Version]
- Rogers, A.; Dietz, K.-J.; Gifford, M.L.; Lunn, J.E. The importance of independent replication of treatments in plant science. J. Exp. Bot. 2021, 72, 5270–5274. [Google Scholar] [CrossRef]
- Salon, C.; Avice, J.-C.; Colombié, S.; Dieuaide-Noubhani, M.; Gallardo, K.; Jeudy, C.; Ourry, A.; Prudent, M.; Voisin, A.-S.; Rolin, D. Fluxomics links cellular functional analyses to whole-plant phenotyping. J. Exp. Bot. 2017, 68, 2083–2098. [Google Scholar] [CrossRef]
- Dietz, K.-J.; Heber, U. Light and CO2 limitation of photosynthesis and states of the reactions regenerating ribulose 1,5-bisphosphate or reducing 3-phosphoglycerate. Biochim. Biophys. Acta (BBA)—Bioenerg. 1986, 848, 392–401. [Google Scholar] [CrossRef]
- Heber, U.; Neimanis, S.; Dietz, K.J.; Viil, J. Assimilatory power as a driving force in photosynthesis. Biochim. Biophys. Acta (BBA)—Bioenerg. 1986, 852, 144–155. [Google Scholar] [CrossRef]
- Engqvist, M.K.M.; Schmitz, J.; Gertzmann, A.; Florian, A.; Jaspert, N.; Arif, M.; Balazadeh, S.; Mueller-Roeber, B.; Fernie, A.R.; Maurino, V.G. GLYCOLATE OXIDASE3, a Glycolate Oxidase Homolog of Yeast l-Lactate Cytochrome c Oxidoreductase, Supports l-Lactate Oxidation in Roots of Arabidopsis. Plant Physiol. 2015, 169, 1042–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pazur, A.; Rassadina, V. Transient effect of weak electromagnetic fields on calcium ion concentration in Arabidopsis thaliana. BMC Plant Biol. 2009, 9, 47. [Google Scholar] [CrossRef] [Green Version]
- Dietz, K.-J. Synergism and antagonism in plant acclimation to abiotic stress combinations. Turk J. Bot. 2021, 45, 587–600. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| A GO Cellular Component With Increased Abundance | A.t. (Ref.) | EMF Effect | Expected | EMF- Enriched | p-Value (FDR) |
|---|---|---|---|---|---|
| Microtubule (0005874) | 182 | 18 | 7.39 | 2.44 | 2.78 × 10-2 |
| Unclassified (UNCLASSIFIED) | 1929 | 111 | 78.34 | 1.42 | 1.01 × 10-2 |
| Cellular component (0005575) | 25,501 | 1003 | 1035.66 | 0.97 | 1.03 × 10-2 |
| Mitochondrion (0005739) | 4385 | 134 | 178.09 | 0.75 | 9.17 × 10-3 |
| Intracellular protein-containing complex (0140535) | 710 | 13 | 28.83 | 0.45 | 3.50 × 10-2 |
| Golgi apparatus (0005794) | 1161 | 20 | 47.15 | 0.42 | 5.86 × 10-4 |
| Catalytic complex (1902494) | 1224 | 19 | 49.71 | 0.38 | 6.98 × 10-5 |
| Plastid membrane (0042170) | 476 | 7 | 19.33 | 0.36 | 4.86 × 10-2 |
| Nucleolus (0005730) | 488 | 6 | 19.82 | 0.3 | 1.59 × 10-2 |
| Endosome (0005768) | 407 | 5 | 16.53 | 0.3 | 3.74 × 10-2 |
| Membrane protein complex (0098796) | 606 | 7 | 24.61 | 0.28 | 2.49 × 10-3 |
| Thylakoid membrane (0042651) | 375 | 4 | 15.23 | 0.26 | 3.63 × 10-2 |
| Chloroplast envelope (0009941) | 601 | 6 | 24.41 | 0.25 | 7.05 × 10-4 |
| Peroxisome (0005777) | 321 | 3 | 13.04 | 0.23 | 4.05 × 10-2 |
| Chloroplast thylakoid (0009534) | 438 | 4 | 17.79 | 0.22 | 7.97 × 10-3 |
| Cytosolic ribosome (0022626) | 294 | 2 | 11.94 | 0.17 | 2.77 × 10-2 |
| Endoplasmic reticulum membrane (0005789) | 305 | 2 | 12.39 | 0.16 | 2.10 × 10-2 |
| Ribosomal subunit (0044391) | 314 | 2 | 12.75 | 0.16 | 1.53 × 10-2 |
| Chloroplast stroma (0009570) | 703 | 2 | 28.55 | 0.07 | 4.17 × 10-8 |
| B GO cellular component With decreased abundance | A.t. (Ref.) | EMF effect | Expected | EMF- enriched | p-value (FDR) |
| Plasmodesma (0009506) | 880 | 15 | 31.28 | 0.48 | 4.8 × 10-2 |
| Chloroplast stroma (0009570) | 703 | 10 | 24.99 | 0.4 | 3.0 × 10-2 |
| Ribonucleoprotein complex (1990904) | 678 | 9 | 24.10 | 0.37 | 2.4 × 10-2 |
| Nucleolus (0005730) | 488 | 5 | 17.35 | 0.29 | 2.8 × 10-2 |
| Plant-type vacuole (0000325) | 787 | 8 | 27.97 | 0.29 | 6.7 × 10-4 |
| Chloroplast envelope (0009941) | 601 | 5 | 21.36 | 0.23 | 2.2 × 10-3 |
| Endosome (0005768) | 407 | 3 | 14.47 | 0.21 | 2.4 × 10-2 |
| Trans-Golgi network (0005802) | 281 | 1 | 9.99 | 0.1 | 3.4 × 10-2 |
| Cytosolic ribosome (0022626) | 294 | 1 | 10.45 | 0.1 | 1.9 × 10-2 |
| Thylakoid (0009579) | 533 | 1 | 18.50 | 0.05 | 2.4 × 10-5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schmidtpott, S.M.; Danho, S.; Kumar, V.; Seidel, T.; Schöllhorn, W.; Dietz, K.-J. Scrutinizing the Impact of Alternating Electromagnetic Fields on Molecular Features of the Model Plant Arabidopsis thaliana. Int. J. Environ. Res. Public Health 2022, 19, 5144. https://doi.org/10.3390/ijerph19095144
Schmidtpott SM, Danho S, Kumar V, Seidel T, Schöllhorn W, Dietz K-J. Scrutinizing the Impact of Alternating Electromagnetic Fields on Molecular Features of the Model Plant Arabidopsis thaliana. International Journal of Environmental Research and Public Health. 2022; 19(9):5144. https://doi.org/10.3390/ijerph19095144
Chicago/Turabian StyleSchmidtpott, Sonja Michèle, Saliba Danho, Vijay Kumar, Thorsten Seidel, Wolfgang Schöllhorn, and Karl-Josef Dietz. 2022. "Scrutinizing the Impact of Alternating Electromagnetic Fields on Molecular Features of the Model Plant Arabidopsis thaliana" International Journal of Environmental Research and Public Health 19, no. 9: 5144. https://doi.org/10.3390/ijerph19095144