
Insights into the Antimicrobial Resistance Profile of a Next Generation Probiotic Akkermansia muciniphila DSM 22959

 ,
,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Phenotypic Antimicrobial Susceptibility Testing
2.3. In Silico Analysis of Antibiotic Resistance Genes (ARG)
3. Results and Discussion
3.1. Phenotypic Antimicrobial Susceptibility Profile
3.2. Prevalence of ARG within the Genomes of A. muciniphila Species
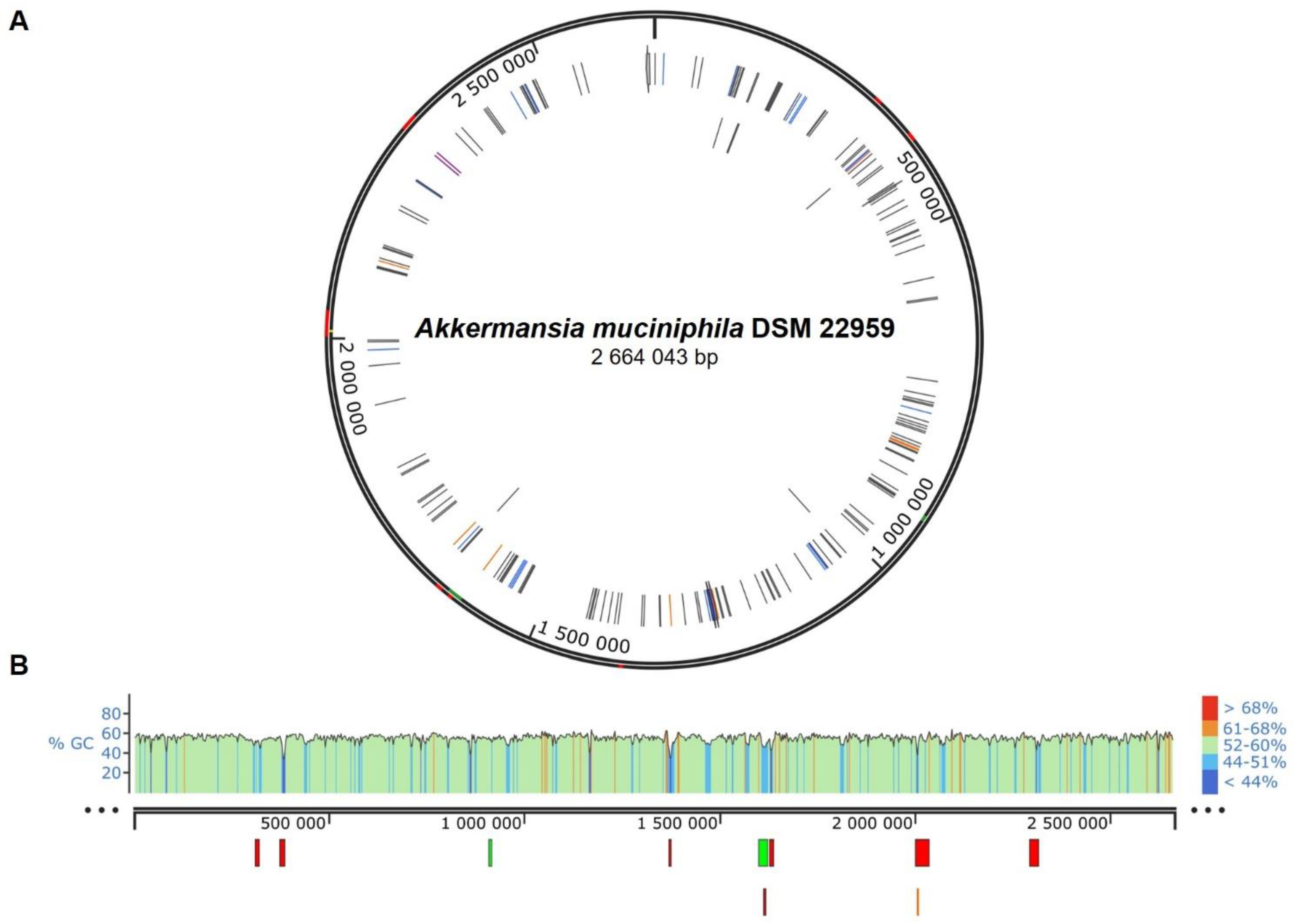
3.3. ARG and Putative Mobile Elements within A. muciniphila DSM 22959 Genome
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S.; et al. Expert Consensus Document: The International Scientific Association for Probiotics and Prebiotics Consensus Statement on the Scope and Appropriate Use of the Term Probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, D.; Machado, D.; Andrade, J.C.; Mendo, S.; Gomes, A.M.; Freitas, A.C. Evolving Trends in Next-Generation Probiotics: A 5W1H Perspective. Crit. Rev. Food Sci. Nutr. 2020, 60, 1783–1796. [Google Scholar] [CrossRef] [PubMed]
- Andrade, J.C.; Almeida, D.; Domingos, M.; Seabra, C.L.; Machado, D.; Freitas, A.C.; Gomes, A.M. Commensal Obligate Anaerobic Bacteria and Health: Production, Storage, and Delivery Strategies. Front. Bioeng. Biotechnol. 2020, 8, 550. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Bose, S.; Lim, S.; Seo, J.; Shin, J.; Lee, D.; Chung, W.-H.; Song, E.-J.; Nam, Y.-D.; Kim, H. Beneficial Effects of Newly Isolated Akkermansia Muciniphila Strains from the Human Gut on Obesity and Metabolic Dysregulation. Microorganisms 2020, 8, 1413. [Google Scholar] [CrossRef]
- Derrien, M.; Vaughan, E.E.; Plugge, C.M.; de Vos, W.M. Akkermansia Municiphila Gen. Nov., Sp. Nov., a Human Intestinal Mucin-Degrading Bacterium. Int. J. Syst. Evol. Microbiol. 2004, 54, 1469–1476. [Google Scholar] [CrossRef] [Green Version]
- Derrien, M.; Collado, M.C.; Ben-Amor, K.; Salminen, S.; De Vos, W.M. The Mucin Degrader Akkermansia Muciniphila Is an Abundant Resident of the Human Intestinal Tract. Appl. Environ. Microbiol. 2008, 74, 1646–1648. [Google Scholar] [CrossRef] [Green Version]
- Depommier, C.; Everard, A.; Druart, C.; Plovier, H.; Van Hul, M.; Vieira-Silva, S.; Falony, G.; Raes, J.; Maiter, D.; Delzenne, N.M.; et al. Supplementation with Akkermansia Muciniphila in Overweight and Obese Human Volunteers: A Proof-of-Concept Exploratory Study. Nat. Med. 2019, 25, 1096–1103. [Google Scholar] [CrossRef]
- EFSA Panel on Biological Hazards; Koutsoumani, K.; Allende, A.; Alvarez-Ordónez, A.; Bolton, D.; Bover-Cid, S.; Chemaly, M.; Davies, R.; De Cesare, A.; Hilbert, F.; et al. Update of the List of QPS-Recommended Biological Agents Intentionally Added to Food or Feed as Notified to EFSA 12: Suitability of Taxonomic Units Notified to EFSA until March 2020. EFSA J. 2020, 18, 6174. [Google Scholar] [CrossRef]
- Li, T.; Teng, D.; Mao, R.; Hao, Y.; Wang, X.; Wang, J. A Critical Review of Antibiotic Resistance in Probiotic Bacteria. Food Res. Int. 2020, 136, 109571. [Google Scholar] [CrossRef]
- Sharma, P.; Tomar, S.K.; Goswami, P.; Sangwan, V.; Singh, R. Antibiotic Resistance among Commercially Available Probiotics. Food Res. Int. 2014, 57, 176–195. [Google Scholar] [CrossRef]
- Aminov, R. History of Antimicrobial Drug Discovery: Major Classes and Health Impact. Biochem. Pharmacol. 2017, 133, 4–19. [Google Scholar] [CrossRef] [PubMed]
- Hernando-Amado, S.; Coque, T.M.; Baquero, F.; Martínez, J.L. Defining and Combating Antibiotic Resistance from One Health and Global Health Perspectives. Nat. Microbiol. 2019, 4, 1432–1442. [Google Scholar] [CrossRef]
- Sirichoat, A.; Flórez, A.B.; Vázquez, L.; Buppasiri, P.; Panya, M.; Lulitanond, V.; Mayo, B. Antibiotic Susceptibility Profiles of Lactic Acid Bacteria from the Human Vagina and Genetic Basis of Acquired Resistances. Int. J. Mol. Sci. 2020, 21, 2594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Antimicrobial Resistance Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Campedelli, I.; Mathur, H.; Salvetti, E.; Clarke, S.; Rea, M.C.; Torriani, S.; Ross, R.P.; Hill, C.; O’Toole, P.W. Genus-Wide Assessment of Antibiotic Resistance in Lactobacillus spp. Appl. Environ. Microbiol. 2019, 85, e01738-18. [Google Scholar] [CrossRef] [Green Version]
- Selvin, J.; Maity, D.; Sajayan, A.; Kiran, G.S. Revealing Antibiotic Resistance in Therapeutic and Dietary Probiotic Supplements. J. Glob. Antimicrob. Resist. 2020, 22, 202–205. [Google Scholar] [CrossRef] [PubMed]
- Neut, C.; Mahieux, S.; Dubreuil, L.J. Antibiotic Susceptibility of Probiotic Strains: Is It Reasonable to Combine Probiotics with Antibiotics? Med. Mal. Infect. 2017, 47, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Anisimova, E.A.; Yarullina, D.R. Antibiotic Resistance of LACTOBACILLUS Strains. Curr. Microbiol. 2019, 76, 1407–1416. [Google Scholar] [CrossRef]
- EFSA-FEEDAP Scientific Opinion on Guidance on the Assessment of Bacterial Susceptibility to Antimicrobials of Human and Veterinary Importance. EFSA J. 2012, 10, 2740. [CrossRef]
- EFSA Panel on Additives and Products or Substances used in Animal Feed (FEEDAP); Rychen, G.; Aquilina, G.; Azimonti, G.; Bampidis, V.; de Lourdes Bastos, M.; Bories, G.; Chesson, A.; Cocconcelli, P.S.; Flachowsky, G.; et al. Guidance on the Characterisation of Microorganisms Used as Feed Additives or as Production Organisms. EFSA J. 2018, 16, e05206. [Google Scholar] [CrossRef]
- Cozzolino, A.; Vergalito, F.; Tremonte, P.; Iorizzo, M.; Lombardi, S.J.; Sorrentino, E.; Luongo, D.; Coppola, R.; Di Marco, R.; Di Marco, R.; et al. Preliminary Evaluation of the Safety and Probiotic Potential of Akkermansia Muciniphila DSM 22959 in Comparison with Lactobacillus Rhamnosus GG. Microorganisms 2020, 8, 189. [Google Scholar] [CrossRef] [Green Version]
- Dubourg, G.; Lagier, J.C.; Armougom, F.; Robert, C.; Audoly, G.; Papazian, L.; Raoult, D. High-Level Colonisation of the Human Gut by Verrucomicrobia Following Broad-Spectrum Antibiotic Treatment. Int. J. Antimicrob. Agents 2013, 41, 149–155. [Google Scholar] [CrossRef] [PubMed]
- van Passel, M.W.J.; Kant, R.; Zoetendal, E.G.; Plugge, C.M.; Derrien, M.; Malfatti, S.A.; Chain, P.S.G.; Woyke, T.; Palva, A.; de Vos, W.M.; et al. The Genome of Akkermansia Muciniphila, a Dedicated Intestinal Mucin Degrader, and Its Use in Exploring Intestinal Metagenomes. PLoS ONE 2011, 6, e16876. [Google Scholar] [CrossRef] [Green Version]
- Guo, X.; Li, S.; Zhang, J.; Wu, F.; Li, X.; Wu, D.; Zhang, M.; Ou, Z.; Jie, Z.; Yan, Q.; et al. Genome Sequencing of 39 Akkermansia Muciniphila Isolates Reveals Its Population Structure, Genomic and Functional Diverisity, and Global Distribution in Mammalian Gut Microbiotas. BMC Genom. 2017, 18, 800. [Google Scholar] [CrossRef]
- DSMZ Akkermansia Muciniphila DSM 22959. Available online: https://www.dsmz.de/collection/catalogue/details/culture/DSM-22959 (accessed on 15 January 2021).
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Davis, J.J.; Wattam, A.R.; Aziz, R.K.; Brettin, T.; Butler, R.; Butler, R.M.; Chlenski, P.; Conrad, N.; Dickerman, A.; Dietrich, E.M.; et al. The PATRIC Bioinformatics Resource Center: Expanding Data and Analysis Capabilities. Nucleic Acids Res. 2020, 48, D606–D612. [Google Scholar] [CrossRef] [Green Version]
- Bortolaia, V.; Kaas, R.S.; Ruppe, E.; Roberts, M.C.; Schwarz, S.; Cattoir, V.; Philippon, A.; Allesoe, R.L.; Rebelo, A.R.; Florensa, A.F.; et al. ResFinder 4.0 for Predictions of Phenotypes from Genotypes. J. Antimicrob. Chemother. 2020, 75, 3491–3500. [Google Scholar] [CrossRef]
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.-L.V.; Cheng, A.A.; Liu, S.; et al. CARD 2020: Antibiotic Resistome Surveillance with the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2019, 48, D517–D525. [Google Scholar] [CrossRef]
- Jia, B.; Raphenya, A.R.; Alcock, B.; Waglechner, N.; Guo, P.; Tsang, K.K.; Lago, B.A.; Dave, B.M.; Pereira, S.; Sharma, A.N.; et al. CARD 2017: Expansion and Model-Centric Curation of the Comprehensive Antibiotic Resistance Database. Nucleic Acids Res. 2017, 45, D566–D573. [Google Scholar] [CrossRef] [PubMed]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L. IslandViewer 4: Expanded Prediction of Genomic Islands for Larger-Scale Datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef]
- Liu, M.; Li, X.; Xie, Y.; Bi, D.; Sun, J.; Li, J.; Tai, C.; Deng, Z.; Ou, H.Y. ICEberg 2.0: An Updated Database of Bacterial Integrative and Conjugative Elements. Nucleic Acids Res. 2019, 47, D660–D665. [Google Scholar] [CrossRef]
- Johansson, M.H.K.; Bortolaia, V.; Tansirichaiya, S.; Aarestrup, F.M.; Roberts, A.P.; Petersen, T.N. Detection of Mobile Genetic Elements Associated with Antibiotic Resistance in Salmonella Enterica Using a Newly Developed Web Tool: MobileElementFinder. J. Antimicrob. Chemother. 2021, 76, 101–109. [Google Scholar] [CrossRef]
- Song, W.; Sun, H.X.; Zhang, C.; Cheng, L.; Peng, Y.; Deng, Z.; Wang, D.; Wang, Y.; Hu, M.; Liu, W.; et al. Prophage Hunter: An Integrative Hunting Tool for Active Prophages. Nucleic Acids Res. 2019, 47, W74–W80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Liang, Y.; Lynch, K.H.; Dennis, J.J.; Wishart, D.S. PHAST: A Fast Phage Search Tool. Nucleic Acids Res. 2011, 39, W347–W352. [Google Scholar] [CrossRef] [PubMed]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A Better, Faster Version of the PHAST Phage Search Tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [Green Version]
- Carver, T.; Harris, S.R.; Berriman, M.; Parkhill, J.; McQuillan, J.A. Artemis: An Integrated Platform for Visualization and Analysis of High-Throughput Sequence-Based Experimental Data. Bioinformatics 2012, 28, 464–469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lonsway, D.R.; Elrod, M.G.; Kendrick, N.; Tiller, R.; Sullivan, M.M.; Edwards, J.R.; Blaney, D.D.; Karlsson, M. Correlation between Etest and Reference Broth Microdilution for Antimicrobial Susceptibility Testing of Burkholderia Pseudomallei. Microb. Drug Resist. 2020, 26, 311–318. [Google Scholar] [CrossRef]
- Azrad, M.; Tkhawkho, L.; Isakovich, N.; Nitzan, O.; Peretz, A. Antimicrobial Susceptibility of Campylobacter Jejuni and Campylobacter Coli: Comparison between Etest and a Broth Dilution Method. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 23. [Google Scholar] [CrossRef]
- Hakvoort, H.; Bovenkamp, E.; Greenwood-Quaintance, K.E.; Schmidt-Malan, S.M.; Mandrekar, J.N.; Schuetz, A.N.; Patel, R. Imipenem-Relebactam Susceptibility Testing of Gram-Negative Bacilli by Agar Dilution, Disk Diffusion, and Gradient Strip Methods Compared with Broth Microdilution. J. Clin. Microbiol. 2020, 58, e00695-20. [Google Scholar] [CrossRef] [PubMed]
- Courvalin, P. Genotypic Approach to the Study of Bacterial Resistance to Antibiotics. Antimicrob. Agents Chemother. 1991, 35, 1019–1023. [Google Scholar] [CrossRef] [Green Version]
- Brinkac, L.; Voorhies, A.; Gomez, A.; Nelson, K.E. The Threat of Antimicrobial Resistance on the Human Microbiome. Microb. Ecol. 2017, 74, 1001–1008. [Google Scholar] [CrossRef]
- Anjum, M.F.; Zankari, E.; Hasman, H. Molecular Methods for Detection of Antimicrobial Resistance. In Antimicrobial Resistance in Bacteria from Livestock and Companion Animals; American Society of Microbiology: Washington, DC, USA, 2017; Volume 5, pp. 33–50. [Google Scholar]
- Wright, G.D. The Antibiotic Resistome. Expert Opin. Drug Discov. 2010, 5, 779–788. [Google Scholar] [CrossRef]
- Benson, D.A.; Cavanaugh, M.; Clark, K.; Karsch-Mizrachi, I.; Ostell, J.; Pruitt, K.D.; Sayers, E.W. GenBank. Nucleic Acids Res. 2018, 46, D41–D47. [Google Scholar] [CrossRef] [Green Version]
- Pruitt, K.D.; Tatusova, T.; Brown, G.R.; Maglott, D.R. NCBI Reference Sequences (RefSeq): Current Status, New Features and Genome Annotation Policy. Nucleic Acids Res. 2012, 40, D130–D135. [Google Scholar] [CrossRef] [Green Version]
- Aziz, R.K.; Bartels, D.; Best, A.; DeJongh, M.; Disz, T.; Edwards, R.A.; Formsma, K.; Gerdes, S.; Glass, E.M.; Kubal, M.; et al. The RAST Server: Rapid Annotations Using Subsystems Technology. BMC Genom. 2008, 9, 75. [Google Scholar] [CrossRef] [Green Version]
- Wattam, A.R.; Abraham, D.; Dalay, O.; Disz, T.L.; Driscoll, T.; Gabbard, J.L.; Gillespie, J.J.; Gough, R.; Hix, D.; Kenyon, R.; et al. PATRIC, the Bacterial Bioinformatics Database and Analysis Resource. Nucleic Acids Res. 2014, 42, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Antonopoulos, D.A.; Assaf, R.; Aziz, R.K.; Brettin, T.; Bun, C.; Conrad, N.; Davis, J.J.; Dietrich, E.M.; Disz, T.; Gerdes, S.; et al. PATRIC as a Unique Resource for Studying Antimicrobial Resistance. Brief. Bioinform. 2019, 20, 1094–1102. [Google Scholar] [CrossRef]
- Baron, S.A.; Diene, S.M.; Rolain, J.M. Human Microbiomes and Antibiotic Resistance. Hum. Microbiome J. 2018, 10, 43–52. [Google Scholar] [CrossRef]
- Rolain, J.M. Food and Human Gut as Reservoirs of Transferable Antibiotic Resistance Encoding Genes. Front. Microbiol. 2013, 4, 173. [Google Scholar] [CrossRef] [Green Version]
- Rolain, J.M.; Abat, C.; Jimeno, M.T.; Fournier, P.E.; Raoult, D. Do We Need New Antibiotics? Clin. Microbiol. Infect. 2016, 22, 408–415. [Google Scholar] [CrossRef] [Green Version]
- Peterson, E.; Kaur, P. Antibiotic Resistance Mechanisms in Bacteria: Relationships between Resistance Determinants of Antibiotic Producers, Environmental Bacteria, and Clinical Pathogens. Front. Microbiol. 2018, 9, 2928. [Google Scholar] [CrossRef]
- Everard, A.; Belzer, C.; Geurts, L.; Ouwerkerk, J.P.; Druart, C.; Bindels, L.B.; Guiot, Y.; Derrien, M.; Muccioli, G.G.; Delzenne, N.M.; et al. Cross-Talk between Akkermansia Muciniphila and Intestinal Epithelium Controls Diet-Induced Obesity. Proc. Natl. Acad. Sci. USA 2013, 110, 9066–9071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrade, F.F.; Silva, D.; Rodrigues, A.; Pina-Vaz, C. Colistin Update on Its Mechanism of Action and Resistance, Present and Future Challenges. Microorganisms 2020, 8, 1716. [Google Scholar] [CrossRef]
- McInnes, R.S.; McCallum, G.E.; Lamberte, L.E.; van Schaik, W. Horizontal Transfer of Antibiotic Resistance Genes in the Human Gut Microbiome. Curr. Opin. Microbiol. 2020, 53, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Jeannot, K.; Xiao, Y.; Knapp, C.W. Editorial: Horizontal Gene Transfer Mediated Bacterial Antibiotic Resistance. Front. Microbiol. 2019, 10, 1933. [Google Scholar] [CrossRef] [Green Version]
- Zankari, E.; Hasman, H.; Kaas, R.S.; Seyfarth, A.M.; Agersø, Y.; Lund, O.; Larsen, M.V.; Aarestrup, F.M. Genotyping Using Whole-Genome Sequencing Is a Realistic Alternative to Surveillance Based on Phenotypic Antimicrobial Susceptibility Testing. J. Antimicrob. Chemother. 2013, 68, 771–777. [Google Scholar] [CrossRef]
- Juhas, M.; van der Meer, J.R.; Gaillard, M.; Harding, R.M.; Hood, D.W.; Crook, D.W. Genomic Islands: Tools of Bacterial Horizontal Gene Transfer and Evolution. FEMS Microbiol. Rev. 2009, 33, 376–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertelli, C.; Tilley, K.E.; Brinkman, F.S.L. Microbial Genomic Island Discovery, Visualization and Analysis. Brief. Bioinform. 2019, 20, 1685–1698. [Google Scholar] [CrossRef] [Green Version]
- da Silva Filho, A.C.; Raittz, R.T.; Guizelini, D.; De Pierri, C.R.; Augusto, D.W.; dos Santos-Weiss, I.C.R.; Marchaukoski, J.N. Comparative Analysis of Genomic Island Prediction Tools. Front. Genet. 2018, 9, 619. [Google Scholar] [CrossRef] [Green Version]
- Chiang, Y.N.; Penadés, J.R.; Chen, J. Genetic Transduction by Phages and Chromosomal Islands: The New and Noncanonical. PLoS Pathog. 2019, 15, e1007878. [Google Scholar] [CrossRef] [Green Version]
- Casjens, S. Prophages and Bacterial Genomics: What Have We Learned so Far? Mol. Microbiol. 2003, 49, 277–300. [Google Scholar] [CrossRef]
- Fortier, L.C.; Sekulovic, O. Importance of Prophages to Evolution and Virulence of Bacterial Pathogens. Virulence 2013, 4, 354–365. [Google Scholar] [CrossRef]
- Ramisetty, B.C.M.; Sudhakari, P.A. Bacterial “grounded” Prophages: Hotspots for Genetic Renovation and Innovation. Front. Genet. 2019, 10, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loh, B.; Chen, J.; Manohar, P.; Yu, Y.; Hua, X.; Leptihn, S. A Biological Inventory of Prophages in A. Baumannii Genomes Reveal Distinct Distributions in Classes, Length and Genomic Positions. Front. Microbiol. 2020, 11, 579802. [Google Scholar] [CrossRef] [PubMed]
- Martínez, J.L. Bottlenecks in the Transferability of Antibiotic Resistance from Natural Ecosystems to Human Bacterial Pathogens. Front. Microbiol. 2012, 2, 265. [Google Scholar] [CrossRef] [PubMed] [Green Version]

{kind=link}
| Antimicrobial | MIC (µg/mL) in Broth Microdilution | MIC (µg/mL) in E-Test® | EFSA-FEEDAP Cut-Off Values (µg/mL) |
|---|---|---|---|
| Ampicillin | ≤2 | 0.25–0.75 | 8 |
| Gentamicin | >4 | 16–32 | 2 |
| Kanamycin | >16 | ≥256 | 8 |
| Streptomycin | >32 | 192–256 | 16 |
| Tetracycline | ≤2 | 0.125–0.25 | 8 |
| Ciprofloxacin | >0.12 | ≥32 | 0.06 |
| Colistin | ≤0.5 | <0.016 | 2 |
| Fosfomycin | 4–8 | 0.5–1 | 8 |
| Mode of Action | CARD Definition | # Hits | Antibiotics | # Hits |
|---|---|---|---|---|
| Antibiotic target in susceptible species | Antibiotic-sensitive wild-type bacterial components; might suffer mutations that render them not susceptible | 4073 | Bicyclomycin | 387 |
| D-cycloserine, Cycloserine | 569 | |||
| Brodimoprim, Iclaprim, Tetroxoprim, Trimethoprim | 189 | |||
| Enacyloxin IIa, Kirromycin, Pulvomycin | 193 | |||
| Ciprofloxacin, Clofazimine, Gatifloxacin, Levofloxacin, Moxifloxacin, Nalidixic acid, Ofloxacin, Sparfloxacin, Trovafloxacin | 193 | |||
| Ciprofloxacin, Clofazimine, Clorobiocin, Coumermycin, Coumermycin A1, Gatifloxacin, Levofloxacin, Moxifloxacin, Nalidixic acid, Novobiocin, Ofloxacin, Sparfloxacin, Trovafloxacin | 193 | |||
| Fosfomycin | 197 | |||
| Fosmidomycin | 190 | |||
| Fusidic acid | 250 | |||
| Ethionamide, Isoniazid, Triclosan | 189 | |||
| Isoniazid, Triclosan | 379 | |||
| Mupirocin (Pseudomonic acid) | 194 | |||
| Daptomycin | 189 | |||
| Daptomycin, Rifabutin, Rifampin, Rifamycin | 195 | |||
| Dapsone, Mafenide, Sulfacetamide, Sulfadiazine, Sulfadimidine, Sulfadoxine, Sulfamethizole, Sulfamethoxazole, Sulfasalazine, Sulfasoxazole | 1190 | |||
| Tetracycline, Tigecycline | 188 | |||
| Aminoglycosides (Streptomycin) | 188 | |||
| Protein altering cell wall charge conferring antibiotic resistance | Cell wall alteration | 188 | Peptide antibiotics (Daptomycin) | 188 |
| Gene conferring resistance via absence | Deletion of gene or gene product results in resistance. For example, deletion of a porin gene blocks drug from entering the cell. | 2 | Aminoglycosides (Streptomycin) | 2 |
| Antibiotic inactivation enzyme | Enzyme that catalyses the inactivation of an antibiotic resulting in resistance. Inactivation includes chemical modification, destruction, etc. | 211 | Beta-lactam antibiotics | 187 |
| Lincosamides (Clindamycin, Lincomycin) | 20 | |||
| Aminoglycosides (Streptomycin) | 4 | |||
| Efflux pump conferring antibiotic resistance | Subunits of efflux proteins that pump antibiotic out of a cell to confer resistance. | 568 | Acriflavin, Amikacin, Arbekacin, Cefepime, Chloramphenicol, Ciprofloxacin, Erythromycin, Gentamicin C, Meropenem, Norfloxacin, Ofloxacin, Tetracycline, Tobramycin | 373 |
| Macrolides (Erythromycin) | 194 | |||
| Tetracyclines | 1 | |||
| Antibiotic target protection protein | These proteins confer antibiotic resistance by bind the antibiotic target to prevent antibiotic binding. | 5 | Tetracyclines | 5 |
| Antibiotic target modifying enzyme | Enzymes that confer resistance by modifying (mutational alteration or enzymatic modification) antibiotic targets. | 59 | Azithromycin, Clarithromycin, Clindamycin, Dalfopristin, Dirithromycin, Erythromycin, Griseoviridin, Lincomycin, Madumycin II, Oleandomycin, Ostreogrycin B3, Patricin A, Patricin B, Pristinamycin IA, Pristinamycin IB, Pristinamycin IIA, Quinupristin, Roxithromycin, Spiramycin, Telithromycin, Tylosin, Vernamycin B-gamma, Vernamycin C, Virginiamycin S2 | 59 |
| Regulator modulating expression of antibiotic resistance genes * | Mechanism activated by the presence of a specific antibiotic. | 1 | Multiple antibiotic resistance * | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machado, D.; Barbosa, J.C.; Almeida, D.; Andrade, J.C.; Freitas, A.C.; Gomes, A.M. Insights into the Antimicrobial Resistance Profile of a Next Generation Probiotic Akkermansia muciniphila DSM 22959. Int. J. Environ. Res. Public Health 2022, 19, 9152. https://doi.org/10.3390/ijerph19159152
Machado D, Barbosa JC, Almeida D, Andrade JC, Freitas AC, Gomes AM. Insights into the Antimicrobial Resistance Profile of a Next Generation Probiotic Akkermansia muciniphila DSM 22959. International Journal of Environmental Research and Public Health. 2022; 19(15):9152. https://doi.org/10.3390/ijerph19159152
Chicago/Turabian StyleMachado, Daniela, Joana Cristina Barbosa, Diana Almeida, José Carlos Andrade, Ana Cristina Freitas, and Ana Maria Gomes. 2022. "Insights into the Antimicrobial Resistance Profile of a Next Generation Probiotic Akkermansia muciniphila DSM 22959" International Journal of Environmental Research and Public Health 19, no. 15: 9152. https://doi.org/10.3390/ijerph19159152