
Implementing Precision Medicine in Human Frailty through Epigenetic Biomarkers

 ,
,  , , and
, , and
Abstract
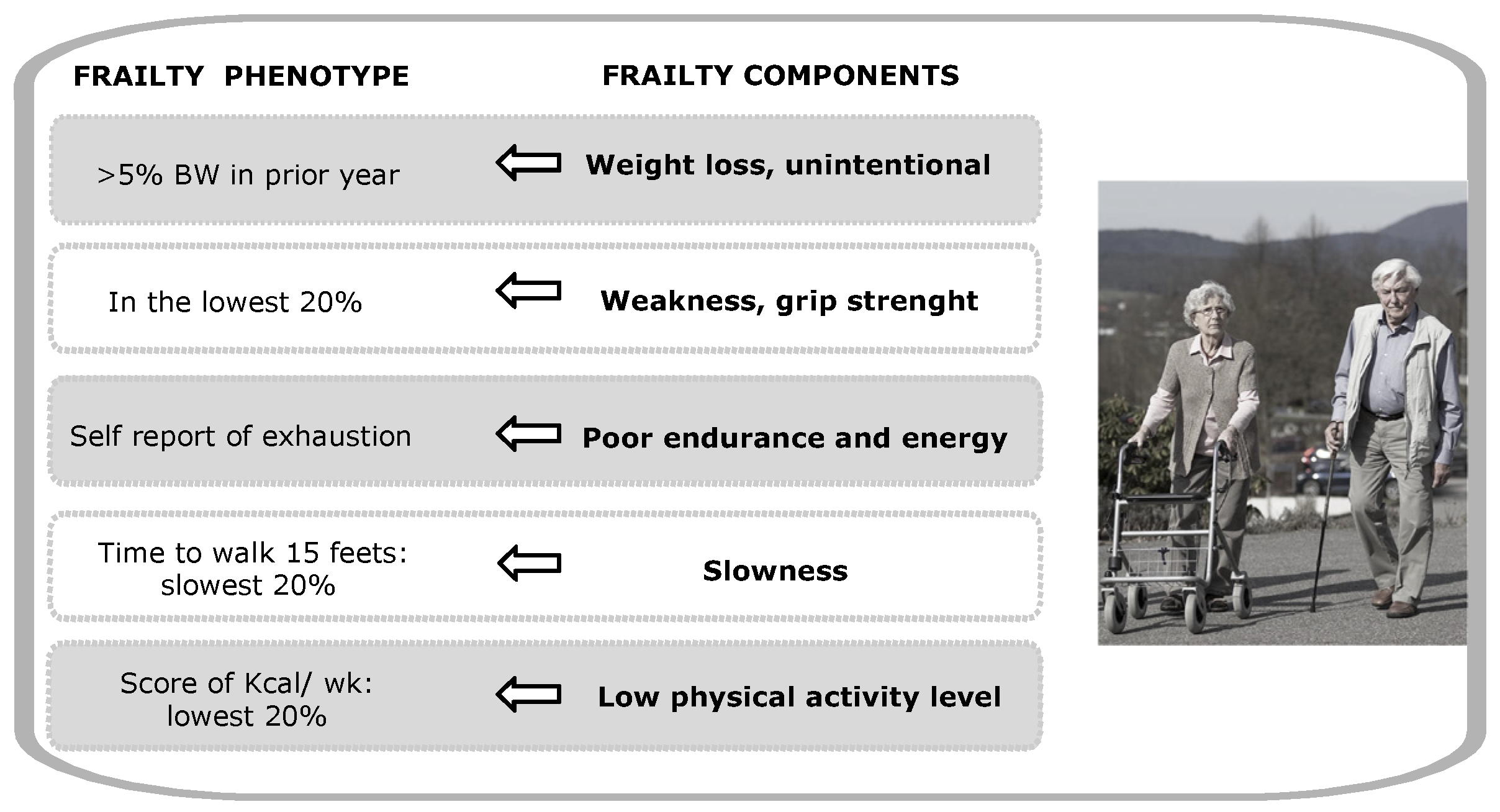
:1. Introduction
2. Materials and Methods
3. Results
3.1. Epigenetic Mechanisms Related to Aging and Frailty
3.1.1. DNA Methylation
3.1.2. Histone Post-Translational Modifications and Histone Variants
3.1.3. Non-Coding RNA
3.2. Epigenetic Biomarkers Associated with Aging and Frailty
3.2.1. DNA Methylation as a Frailty Biomarker
3.2.2. Histone PTMs and Histone Variants as Frailty Biomarkers
3.2.3. Non-Coding RNAs as Frailty Biomarkers
3.3. Epigenetic Biomarkers to Follow-up the Interventions in Frailty
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fried, L.P.; Tangen, C.M.; Walston, J.; Newman, A.B.; Hirsch, C.; Gottdiener, J.; Seeman, T.; Tracy, R.; Kop, W.J.; Burke, G.; et al. Frailty in Older Adults: Evidence for a Phenotype. J. Gerontol. Ser. A Biol. Sci. Med. Sci. 2001, 56, M146–M157. [Google Scholar] [CrossRef]
- Rodriguez-Mañas, L.; Fried, L.P. Frailty in the clinical scenario. Lancet 2015, 385, e7–e9. [Google Scholar] [CrossRef]
- Bisset, E.S.; Howlett, S.E. The biology of frailty in humans and animals: Understanding frailty and promoting translation. Aging Med. 2019, 2, 27–34. [Google Scholar] [CrossRef]
- Rockwood, K. Conceptual Models of Frailty: Accumulation of Deficits. Can. J. Cardiol. 2016, 32, 1046–1050. [Google Scholar] [CrossRef] [PubMed]
- van Deursen, J.M. The role of senescent cells in ageing. Nature 2014, 509, 439–446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viña, J.; Tarazona-Santabalbina, F.J.; Pérez-Ros, P.; Martínez-Arnau, F.M.; Borras, C.; Olaso-Gonzalez, G.; Salvador-Pascual, A.; Gomez-Cabrera, M.C. Biology of frailty: Modulation of ageing genes and its importance to prevent age-associated loss of function. Mol. Asp. Med. 2016, 50, 88–108. [Google Scholar] [CrossRef]
- Nascimento, C.M.; Ingles, M.; Salvador-Pascual, A.; Cominetti, M.R.; Gomez-Cabrera, M.C.; Viña, J. Sarcopenia, frailty and their prevention by exercise. Free. Radic. Biol. Med. 2019, 132, 42–49. [Google Scholar] [CrossRef]
- Bock, J.-O.; König, H.-H.; Brenner, H.; Haefeli, W.E.; Quinzler, R.; Matschinger, H.; Saum, K.-U.; Schöttker, B.; Heider, D. Associations of frailty with health care costs—Results of the ESTHER cohort study. BMC Health Serv. Res. 2016, 16, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Collard, R.M.; Boter, H.; Schoevers, R.A.; Oude Voshaar, R.C. Prevalence of Frailty in Community-Dwelling Older Persons: A Systematic Review. J. Am. Geriatr. Soc. 2012, 60, 1487–1492. [Google Scholar] [CrossRef]
- Eeles, E.M.P.; White, S.V.; O’Mahony, S.M.; Bayer, A.J.; Hubbard, R.E. The impact of frailty and delirium on mortality in older inpatients. Age Ageing 2012, 41, 412–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krishnan, M.; Beck, S.; Havelock, W.; Eeles, E.; Hubbard, R.E.; Johansen, A. Predicting outcome after hip fracture: Using a frailty index to integrate comprehensive geriatric assessment results. Age Ageing 2014, 43, 122–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hubbard, R.E.; Andrew, M.K.; Fallah, N.; Rockwood, K. Comparison of the prognostic importance of diagnosed diabetes, co-morbidity and frailty in older people. Diabet. Med. 2010, 27, 603–606. [Google Scholar] [CrossRef] [PubMed]
- Sipila, S.; Taaffe, D.R.; Cheng, S.; Puolakka, J.; Toivanen, J.; Suominen, H. Effects of hormone replacement therapy and high-impact physical exercise on skeletal muscle in post-menopausal women: A randomized placebo-controlled study. Clin. Sci. 2001, 101, 147–157. [Google Scholar] [CrossRef]
- Clegg, A.; Young, J.; Iliffe, S.; Rikkert, M.O.; Rockwood, K. Frailty in elderly people. Lancet 2013, 381, 752–762. [Google Scholar] [CrossRef] [Green Version]
- Marzetti, E.; Leeuwenburgh, C. Skeletal muscle apoptosis, sarcopenia and frailty at old age. Exp. Gerontol. 2006, 41, 1234–1238. [Google Scholar] [CrossRef] [PubMed]
- Vaz Fragoso, C.A.; Enright, P.L.; McAvay, G.; Van Ness, P.H.; Gill, T.M. Frailty and Respiratory Impairment in Older Persons. Am. J. Med. 2012, 125, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Lutski, M.; Haratz, S.; Weinstein, G.; Goldbourt, U.; Tanne, D. Impaired Cerebral Hemodynamics and Frailty in Patients with Cardiovascular Disease. J. Gerontol. Ser. A 2018, 73, 1714–1721. [Google Scholar] [CrossRef] [Green Version]
- Drew, W.; Wilson, D.; Sapey, E. Frailty and the immune system. J. Aging Res. Health 2017, 2, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Abel, G.A.; Buckstein, R. Integrating Frailty, Comorbidity, and Quality of Life in the Management of Myelodysplastic Syndromes. Am. Soc. Clin. Oncol. Educ. Book 2016, 36, e337–e344. [Google Scholar] [CrossRef]
- Walston, J.; Fedarko, N.; Yang, H.; Leng, S.; Beamer, B.; Espinoza, S.; Lipton, A.; Zheng, H.; Becker, K. The Physical and Biological Characterization of a Frail Mouse Model. J. Gerontol. Ser. A Boil. Sci. Med. Sci. 2008, 63, 391–398. [Google Scholar] [CrossRef] [Green Version]
- Boyle, P.A.; Buchman, A.S.; Wilson, R.S.; Leurgans, S.E.; Bennett, D.A. Physical frailty is associated with incident mild cognitive impairment in community-based older persons. J. Am. Geriatr. Soc. 2010, 58, 248–255. [Google Scholar] [CrossRef]
- Lipton, M.L.; Ifrah, C.; Stewart, W.F.; Fleysher, R.; Sliwinski, M.J.; Kim, M.; Lipton, R.B. Validation of HeadCount-2w for estimation of two-week heading: Comparison to daily reporting in adult amateur player. J. Sci. Med. Sport 2018, 21, 363–367. [Google Scholar] [CrossRef]
- Beltrán-García, J.; Osca-Verdegal, R.; Mena-Mollá, S.; García-Giménez, J.L. Epigenetic IVD Tests for Personalized Precision Medicine in Cancer. Front. Genet. 2019, 10, 621. [Google Scholar] [CrossRef]
- Garcia-Gimenez, J.L.; Sanchis-Gomar, F.; Lippi, G.; Mena, S.; Ivars, D.; Gomez-Cabrera, M.C.; Vina, J.; Pallardo, F.V. Epigenetic biomarkers: A new perspective in laboratory diagnostics. Clin. Chim. Acta 2012, 413, 1576–1582. [Google Scholar] [CrossRef] [PubMed]
- García-Giménez, J.L.; Seco-Cervera, M.; Tollefsbol, T.O.; Romá-Mateo, C.; Peiró-Chova, L.; Lapunzina, P.; Pallardó, F.V. Epigenetic biomarkers: Current strategies and future challenges for their use in the clinical laboratory. Crit. Rev. Clin. Lab. Sci. 2017, 54, 529–550. [Google Scholar] [CrossRef]
- Sandoval, J.; Peiró-Chova, L.; Pallardó, F.V.; García-Giménez, J.L. Epigenetic biomarkers in laboratory diagnostics: Emerging approaches and opportunities. Expert Rev. Mol. Diagn. 2013, 13, 457–471. [Google Scholar] [CrossRef]
- Pal, S.; Tyler, J.K. Epigenetics and aging. Sci. Adv. 2016, 2, e1600584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrucci, L.; Cavazzini, C.; Corsi, A.; Bartali, B.; Russo, C.R.; Lauretani, F.; Corsi, A.M.; Bandinelli, S.; Guralnik, J.M. Biomarkers of frailty in older persons. J. Endocrinol. Investig. 2002, 25, 10–15. [Google Scholar]
- van der Graaf, A.; Wardenaar, R.; Neumann, D.A.; Taudt, A.; Shaw, R.G.; Jansen, R.C.; Schmitz, R.J.; Colomé-Tatché, M.; Johannes, F. Rate, spectrum, and evolutionary dynamics of spontaneous epimutations. Proc. Natl. Acad. Sci. USA 2015, 112, 6676–6681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cruickshanks, H.A.; McBryan, T.; Nelson, D.M.; VanderKraats, N.D.; Shah, P.P.; van Tuyn, J.; Singh Rai, T.; Brock, C.; Donahue, G.; Dunican, D.S.; et al. Senescent cells harbour features of the cancer epigenome. Nat. Cell Biol. 2013, 15, 1495–1506. [Google Scholar] [CrossRef] [PubMed]
- Jung, M.; Pfeifer, G.P. Aging and DNA methylation. BMC Biol. 2015, 13, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez-Verjan, J.C.; Ramírez-Aldana, R.; Pérez-Zepeda, M.U.; Quiroz-Baez, R.; Luna-López, A.; Gutierrez Robledo, L.M. Systems biology and network pharmacology of frailty reveal novel epigenetic targets and mechanisms. Sci. Rep. 2019, 9, 10593. [Google Scholar] [CrossRef] [PubMed]
- Fraga, M.F.; Ballestar, E.; Paz, M.F.; Ropero, S.; Setien, F.; Ballestar, M.L.; Heine-Suner, D.; Cigudosa, J.C.; Urioste, M.; Benitez, J.; et al. From The Cover: Epigenetic differences arise during the lifetime of monozygotic twins. Proc. Natl. Acad. Sci. USA 2005, 102, 10604–10609. [Google Scholar] [CrossRef] [Green Version]
- Veitia, R.A.; Govindaraju, D.R.; Bottani, S.; Birchler, J.A. Aging: Somatic Mutations, Epigenetic Drift and Gene Dosage Imbalance. Trends Cell Biol. 2017, 27, 299–310. [Google Scholar] [CrossRef]
- Sousa-Victor, P.; Gutarra, S.; García-Prat, L.; Rodriguez-Ubreva, J.; Ortet, L.; Ruiz-Bonilla, V.; Jardí, M.; Ballestar, E.; González, S.; Serrano, A.L.; et al. Geriatric muscle stem cells switch reversible quiescence into senescence. Nature 2014, 506, 316–321. [Google Scholar] [CrossRef]
- Bellizzi, D.; D’Aquila, P.; Montesanto, A.; Corsonello, A.; Mari, V.; Mazzei, B.; Lattanzio, F.; Passarino, G. Global DNA methylation in old subjects is correlated with frailty. AGE 2012, 34, 169–179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montesanto, A.; Lagani, V.; Martino, C.; Dato, S.; De Rango, F.; Berardelli, M.; Corsonello, A.; Mazzei, B.; Mari, V.; Lattanzio, F.; et al. A novel, population-specific approach to define frailty. AGE 2010, 32, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Horvath, S.; Raj, K. DNA methylation-based biomarkers and the epigenetic clock theory of ageing. Nat. Rev. Genet. 2018, 19, 371–384. [Google Scholar] [CrossRef] [PubMed]
- Roetker, N.S.; Pankow, J.S.; Bressler, J.; Morrison, A.C.; Boerwinkle, E. Prospective Study of Epigenetic Age Acceleration and Incidence of Cardiovascular Disease Outcomes in the ARIC Study (Atherosclerosis Risk in Communities). Circ. Genom. Precis. Med. 2018, 11, e001937. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 2013, 14, R115. [Google Scholar] [CrossRef] [Green Version]
- Ambatipudi, S.; Horvath, S.; Perrier, F.; Cuenin, C.; Hernandez-Vargas, H.; Le Calvez-Kelm, F.; Durand, G.; Byrnes, G.; Ferrari, P.; Bouaoun, L.; et al. DNA methylome analysis identifies accelerated epigenetic ageing associated with postmenopausal breast cancer susceptibility. Eur. J. Cancer 2017, 75, 299–307. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.E.; Hosgood, H.D.; Chen, B.; Absher, D.; Assimes, T.; Horvath, S. DNA methylation age of blood predicts future onset of lung cancer in the women’s health initiative. Aging 2015, 7, 690–700. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.E.; Lu, A.T.; Bennett, D.A.; Horvath, S. Epigenetic age of the pre-frontal cortex is associated with neuritic plaques, amyloid load, and Alzheimer’s disease related cognitive functioning. Aging 2015, 7, 1198–1211. [Google Scholar] [CrossRef]
- Chen, B.H.; Marioni, R.E.; Colicino, E.; Peters, M.J.; Ward-Caviness, C.K.; Tsai, P.-C.; Roetker, N.S.; Just, A.C.; Demerath, E.W.; Guan, W.; et al. DNA methylation-based measures of biological age: Meta-analysis predicting time to death. Aging 2016, 8, 1844–1865. [Google Scholar] [CrossRef] [Green Version]
- Levine, M.E.; Lu, A.T.; Quach, A.; Chen, B.H.; Assimes, T.L.; Bandinelli, S.; Hou, L.; Baccarelli, A.A.; Stewart, J.D.; Li, Y.; et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging 2018, 10, 573–591. [Google Scholar] [CrossRef] [Green Version]
- Marioni, R.E.; Shah, S.; McRae, A.F.; Ritchie, S.J.; Muniz-Terrera, G.; Harris, S.E.; Gibson, J.; Redmond, P.; Cox, S.R.; Pattie, A.; et al. The epigenetic clock is correlated with physical and cognitive fitness in the Lothian Birth Cohort 1936. Int. J. Epidemiol. 2015, 44, 1388–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wilson, R.; Heiss, J.; Breitling, L.P.; Saum, K.-U.; Schöttker, B.; Holleczek, B.; Waldenberger, M.; Peters, A.; Brenner, H. DNA methylation signatures in peripheral blood strongly predict all-cause mortality. Nat. Commun. 2017, 8, 14617. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Leung, D.; Thrush, K.; Zhao, W.; Ratliff, S.; Tanaka, T.; Schmitz, L.L.; Smith, J.A.; Ferrucci, L.; Levine, M.E. Underlying features of epigenetic aging clocks in vivo and in vitro. Aging Cell 2020, 19, e13229. [Google Scholar] [CrossRef] [PubMed]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.-B.; Gao, Y.; et al. Genome-wide Methylation Profiles Reveal Quantitative Views of Human Aging Rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weidner, C.; Lin, Q.; Koch, C.; Eisele, L.; Beier, F.; Ziegler, P.; Bauerschlag, D.; Jöckel, K.-H.; Erbel, R.; Mühleisen, T.; et al. Aging of blood can be tracked by DNA methylation changes at just three CpG sites. Genome Biol. 2014, 15, R24. [Google Scholar] [CrossRef] [Green Version]
- Vidal-Bralo, L.; Lopez-Golan, Y.; Gonzalez, A. Simplified Assay for Epigenetic Age Estimation in Whole Blood of Adults. Front. Genet. 2016, 7, 126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, Q.; Wagner, W. Epigenetic Aging Signatures Are Coherently Modified in Cancer. PLoS Genet. 2015, 11, e1005334. [Google Scholar] [CrossRef]
- Bergsma, T.; Rogaeva, E. DNA Methylation Clocks and Their Predictive Capacity for Aging Phenotypes and Healthspan. Neurosci. Insights 2020, 15, 263310552094222. [Google Scholar] [CrossRef]
- Breitling, L.P.; Saum, K.-U.; Perna, L.; Schöttker, B.; Holleczek, B.; Brenner, H. Frailty is associated with the epigenetic clock but not with telomere length in a German cohort. Clin. Epigenet. 2016, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gale, C.R.; Marioni, R.E.; Harris, S.E.; Starr, J.M.; Deary, I.J. DNA methylation and the epigenetic clock in relation to physical frailty in older people: The Lothian Birth Cohort 1936. Clin. Epigenet. 2018, 10, 101. [Google Scholar] [CrossRef]
- Kim, D.H.; Schneeweiss, S.; Glynn, R.J.; Lipsitz, L.A.; Rockwood, K.; Avorn, J. Measuring Frailty in Medicare Data: Development and Validation of a Claims-Based Frailty Index. J. Gerontol. Ser. A 2018, 73, 980–987. [Google Scholar] [CrossRef] [PubMed]
- García-Giménez, J.L.; Romá-Mateo, C.; Pallardó, F.V. Oxidative post-translational modifications in histones. BioFactors 2019, 45, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Benayoun, B.A.; Pollina, E.A.; Brunet, A. Epigenetic regulation of ageing: Linking environmental inputs to genomic stability. Nat. Rev. Mol. Cell Biol. 2015, 16, 593–610. [Google Scholar] [CrossRef] [Green Version]
- Siddiqui, M.S.; François, M.; Fenech, M.F.; Leifert, W.R. Persistent γH2AX: A promising molecular marker of DNA damage and aging. Mutat. Res. Mutat. Res. 2015, 766, 1–19. [Google Scholar] [CrossRef]
- Sedelnikova, O.A.; Horikawa, I.; Redon, C.; Nakamura, A.; Zimonjic, D.B.; Popescu, N.C.; Bonner, W.M. Delayed kinetics of DNA double-strand break processing in normal and pathological aging. Aging Cell 2008, 7, 89–100. [Google Scholar] [CrossRef]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Vasudevan, S.; Tong, Y.; Steitz, J.A. Switching from repression to activation: MicroRNAs can up-regulate translation. Science 2007, 318, 1931–1934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Mercer, T.R.; Mattick, J.S. Structure and function of long noncoding RNAs in epigenetic regulation. Nat. Struct. Mol. Biol. 2013, 20, 300–307. [Google Scholar] [CrossRef]
- Kobashigawa, J.; Dadhania, D.; Bhorade, S.; Adey, D.; Berger, J.; Bhat, G.; Budev, M.; Duarte-Rojo, A.; Dunn, M.; Hall, S.; et al. Report from the American Society of Transplantation on frailty in solid organ transplantation. Am. J. Transplant. 2019, 19, 984–994. [Google Scholar] [CrossRef] [PubMed]
- Sannicandro, A.J.; Soriano-Arroquia, A.; Goljanek-Whysall, K. Micro(RNA)-managing muscle wasting. J. Appl. Physiol. 2019, 127, 619–632. [Google Scholar] [CrossRef] [PubMed]
- Smith-Vikos, T.; Liu, Z.; Parsons, C.; Gorospe, M.; Ferrucci, L.; Gill, T.M.; Slack, F.J. A serum miRNA profile of human longevity: Findings from the Baltimore Longitudinal Study of Aging (BLSA). Aging 2016, 8, 2971–2987. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valadi, H.; Ekström, K.; Bossios, A.; Sjöstrand, M.; Lee, J.J.; Lötvall, J.O. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol. 2007, 9, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Rusanova, I.; Fernández-Martínez, J.; Fernández-Ortiz, M.; Aranda-Martínez, P.; Escames, G.; García-García, F.J.; Mañas, L.; Acuña-Castroviejo, D. Involvement of plasma miRNAs, muscle miRNAs and mitochondrial miRNAs in the pathophysiology of frailty. Exp. Gerontol. 2019, 124, 110637. [Google Scholar] [CrossRef]
- García-Giménez, J.L. Epigenetic Biomarkers and Diagnostics; Academic Press: Amsterdam, The Netherlands, 2015. [Google Scholar]
- Theou, O.; Sluggett, J.K.; Bell, J.S.; Lalic, S.; Cooper, T.; Robson, L.; Morley, J.E.; Rockwood, K.; Visvanathan, R. Frailty, Hospitalization, and Mortality in Residential Aged Care. J. Gerontol. Ser. A 2018, 73, 1090–1096. [Google Scholar] [CrossRef]
- Collerton, J.; Gautrey, H.E.; van Otterdijk, S.D.; Davies, K.; Martin-Ruiz, C.; von Zglinicki, T.; Kirkwood, T.B.L.; Jagger, C.; Mathers, J.C.; Strathdee, G. Acquisition of aberrant DNA methylation is associated with frailty in the very old: Findings from the Newcastle 85+ Study. Biogerontology 2014, 15, 317–328. [Google Scholar] [CrossRef] [PubMed]
- Mitnitski, A.; Rockwood, K. The rate of aging: The rate of deficit accumulation does not change over the adult life span. Biogerontology 2016, 17, 199–204. [Google Scholar] [CrossRef]
- Vetter, V.M.; Meyer, A.; Karbasiyan, M.; Steinhagen-Thiessen, E.; Hopfenmüller, W.; Demuth, I. Epigenetic Clock and Relative Telomere Length Represent Largely Different Aspects of Aging in the Berlin Aging Study II (BASE-II). J. Gerontol. Ser. A 2019, 74, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Vetter, V.M.; Spira, D.; Banszerus, V.L.; Demuth, I. Epigenetic Clock and Leukocyte Telomere Length Are Associated with Vitamin D Status but not with Functional Assessments and Frailty in the Berlin Aging Study II. J. Gerontol. Ser. A 2020, 75, 2056–2063. [Google Scholar] [CrossRef] [PubMed]
- Shulman, K.I. Clock-drawing: Is it the ideal cognitive screening test? Int. J. Geriatr. Psychiatry 2000, 15, 548–561. [Google Scholar] [CrossRef]
- Gräff, J.; Tsai, L.H. Histone acetylation: Molecular mnemonics on the chromatin. Nat. Rev. Neurosci. 2013, 14, 97–111. [Google Scholar] [CrossRef]
- Burns, A.M.; Gräff, J. Cognitive epigenetic priming: Leveraging histone acetylation for memory amelioration. Curr. Opin. Neurobiol. 2020, 67, 75–84. [Google Scholar] [CrossRef]
- Yoshihara, T.; Machida, S.; Tsuzuki, T.; Kakigi, R.; Chang, S.W.; Sugiura, T.; Naito, H. Age-related changes in histone modification in rat gastrocnemius muscle. Exp. Gerontol. 2019, 125, 110658. [Google Scholar] [CrossRef]
- Walsh, M.E.; Bhattacharya, A.; Sataranatarajan, K.; Qaisar, R.; Sloane, L.; Rahman, M.M.; Kinter, M.; Van Remmen, H. The histone deacetylase inhibitor butyrate improves metabolism and reduces muscle atrophy during aging. Aging Cell 2015, 14, 957–970. [Google Scholar] [CrossRef] [PubMed]
- Valdiglesias, V.; Sánchez-Flores, M.; Marcos-Pérez, D.; Lorenzo-López, L.; Maseda, A.; Millán-Calenti, J.C.; Pásaro, E.; Laffon, B. Exploring Genetic Outcomes as Frailty Biomarkers. J. Gerontol. Ser. A 2019, 74, 168–175. [Google Scholar] [CrossRef]
- Ipson, B.R.; Fletcher, M.B.; Espinoza, S.E.; Fisher, A.L. Identifying Exosome-Derived MicroRNAs as Candidate Biomarkers of Frailty. J. Frailty Aging 2018, 7, 100–103. [Google Scholar] [CrossRef]
- Rusanova, I.; Diaz-Casado, M.E.; Fernández-Ortiz, M.; Aranda-Martínez, P.; Guerra-Librero, A.; García-García, F.J.; Escames, G.; Mañas, L.; Acuña-Castroviejo, D. Analysis of Plasma MicroRNAs as Predictors and Biomarkers of Aging and Frailty in Humans. Oxidative Med. Cell. Longev. 2018, 2018, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Giménez, J.L.; Rubio-Belmar, P.A.; Peiró-Chova, L.; Hervás, D.; González-Rodríguez, D.; Ibañez-Cabellos, J.S.; Bas-Hermida, P.; Mena-Mollá, S.; García-López, E.M.; Pallardó, F.V.; et al. Circulating miRNAs as diagnostic biomarkers for adolescent idiopathic scoliosis. Sci. Rep. 2018, 8, 2646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greco, E.A.; Pietschmann, P.; Migliaccio, S. Osteoporosis and Sarcopenia Increase Frailty Syndrome in the Elderly. Front. Endocrinol. 2019, 10, 255. [Google Scholar] [CrossRef]
- Wu, I.C.; Lin, C.C.; Hsiung, C.A. Emerging roles of frailty and inflammaging in risk assessment of age-related chronic diseases in older adults: The intersection between aging biology and personalized medicine. Biomedicine 2015, 5, 1. [Google Scholar] [CrossRef]
- Zheng, Y.; Kong, J.; Li, Q.; Wang, Y.; Li, J. Role of miRNAs in skeletal muscle aging. Clin. Interv. Aging 2018, 13, 2407–2419. [Google Scholar] [CrossRef] [Green Version]
- Sathyan, S.; Barzilai, N.; Atzmon, G.; Milman, S.; Ayers, E.; Verghese, J. Genetic Insights Into Frailty: Association of 9p21-23 Locus With Frailty. Front. Med. 2018, 5, 105. [Google Scholar] [CrossRef] [Green Version]
- Pasmant, E.; Sabbagh, A.; Vidaud, M.; Bièche, I. ANRIL, a long, noncoding RNA, is an unexpected major hotspot in GWAS. FASEB J. 2011, 25, 444–448. [Google Scholar] [CrossRef]
- Melzer, D.; Frayling, T.M.; Murray, A.; Hurst, A.J.; Harries, L.W.; Song, H.; Khaw, K.; Luben, R.; Surtees, P.G.; Bandinelli, S.S.; et al. A common variant of the p16(INK4a) genetic region is associated with physical function in older people. Mech. Ageing Dev. 2007, 128, 370–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, K.M.; Noh, J.H.; Yoon, J.H.; Abdelmohsen, K.; Gorospe, M. Long noncoding RNAs in diseases of aging. Biochim. Biophys. Acta 2016, 1859, 209–221. [Google Scholar] [CrossRef] [Green Version]
- Dey, B.K.; Pfeifer, K.; Dutta, A. The H19 long noncoding RNA gives rise to microRNAs miR-675-3p and miR-675-5p to promote skeletal muscle differentiation and regeneration. Genes Dev. 2014, 28, 491–501. [Google Scholar] [CrossRef] [Green Version]
- McKay, B.R.; Ogborn, D.I.; Bellamy, L.M.; Tarnopolsky, M.A.; Parise, G. Myostatin is associated with age-related human muscle stem cell dysfunction. FASEB J. 2012, 26, 2509–2521. [Google Scholar] [CrossRef]
- Watts, R.; Johnsen, V.L.; Shearer, J.; Hittel, D.S. Myostatin-induced inhibition of the long noncoding RNA Malat1 is associated with decreased myogenesis. Am. J. Physiol. Cell Physiol. 2013, 304, C995–C1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legnini, I.; Morlando, M.; Mangiavacchi, A.; Fatica, A.; Bozzoni, I. A feedforward regulatory loop between HuR and the long noncoding RNA linc-MD1 controls early phases of myogenesis. Mol. Cell 2014, 53, 506–514. [Google Scholar] [CrossRef] [Green Version]
- Pardo, P.S.; Boriek, A.M. The physiological roles of Sirt1 in skeletal muscle. Aging (Albany NY) 2011, 3, 430–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viña, J.; Rodriguez-Mañas, L.; Salvador-Pascual, A.; Tarazona-Santabalbina, F.J.; Gomez-Cabrera, M.C. Exercise: The lifelong supplement for healthy ageing and slowing down the onset of frailty. J. Physiol. 2016, 594, 1989–1999. [Google Scholar] [CrossRef] [Green Version]
- Arc-Chagnaud, C.; Millan, F.; Salvador-Pascual, A.; Correas, A.G.; Olaso-Gonzalez, G.; De la Rosa, A.; Carretero, A.; Gomez-Cabrera, M.C.; Viña, J. Reversal of age-associated frailty by controlled physical exercise: The pre-clinical and clinical evidences. Sports Med. Health Sci. 2019, 1, 33–39. [Google Scholar] [CrossRef]
- Ramírez-Vélez, R.; Izquierdo, M. Editorial: Precision Physical Activity and Exercise Prescriptions for Disease Prevention: The Effect of Interindividual Variability Under Different Training Approaches. Front. Physiol. 2019, 10, 646. [Google Scholar] [CrossRef] [PubMed]
- Espinoza, S.E.; Jiwani, R.; Wang, J.; Wang, C.P. Review of Interventions for the Frailty Syndrome and the Role of Metformin as a Potential Pharmacologic Agent for Frailty Prevention. Clin. Ther. 2019, 41, 376–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaeberlein, M. How healthy is the healthspan concept? Geroscience 2018, 40, 361–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palliyaguru, D.L.; Moats, J.M.; Di Germanio, C.; Bernier, M.; de Cabo, R. Frailty index as a biomarker of lifespan and healthspan: Focus on pharmacological interventions. Mech. Ageing Dev. 2019, 180, 42–48. [Google Scholar] [CrossRef]
- Garcia-Valles, R.; Gomez-Cabrera, M.C.; Rodriguez-Manas, L.; Garcia-Garcia, F.J.; Diaz, A.; Noguera, I.; Olaso-Gonzalez, G.; Vina, J. Life-long spontaneous exercise does not prolong lifespan but improves health span in mice. Longev. Health 2013, 2, 14. [Google Scholar] [CrossRef] [Green Version]
- Gomez-Cabrera, M.C.; Garcia-Valles, R.; Rodriguez-Mañas, L.; Garcia-Garcia, F.J.; Olaso-Gonzalez, G.; Salvador-Pascual, A.; Tarazona-Santabalbina, F.J.; Viña, J. A New Frailty Score for Experimental Animals Based on the Clinical Phenotype: Inactivity as a Model of Frailty. J. Gerontol. A Biol. Sci. Med. Sci. 2017, 72, 885–891. [Google Scholar] [CrossRef] [Green Version]
- Tarazona-Santabalbina, F.J.; Gomez-Cabrera, M.C.; Perez-Ros, P.; Martinez-Arnau, F.M.; Cabo, H.; Tsaparas, K.; Salvador-Pascual, A.; Rodriguez-Manas, L.; Vina, J. A Multicomponent Exercise Intervention that Reverses Frailty and Improves Cognition, Emotion, and Social Networking in the Community-Dwelling Frail Elderly: A Randomized Clinical Trial. J. Am. Med. Dir. Assoc. 2016, 17, 426–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Viña, J.; Salvador-Pascual, A.; Tarazona-Santabalbina, F.J.; Rodriguez-Mañas, L.; Gomez-Cabrera, M.C. Exercise training as a drug to treat age associated frailty. Free. Radic. Biol. Med. 2016, 98, 159–164. [Google Scholar] [CrossRef]
- Marques, E.; Carvalho, J.; Soares, J.M.; Marques, F.; Mota, J. Effects of resistance and multicomponent exercise on lipid profiles of older women. Maturitas 2009, 63, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Cadore, E.L.; Casas-Herrero, A.; Zambom-Ferraresi, F.; Idoate, F.; Millor, N.; Gomez, M.; Rodriguez-Manas, L.; Izquierdo, M. Multicomponent exercises including muscle power training enhance muscle mass, power output, and functional outcomes in institutionalized frail nonagenarians. Age (Dordr) 2014, 36, 773–785. [Google Scholar] [CrossRef] [Green Version]
- Gine-Garriga, M.; Roque-Figuls, M.; Coll-Planas, L.; Sitja-Rabert, M.; Salva, A. Physical exercise interventions for improving performance-based measures of physical function in community-dwelling, frail older adults: A systematic review and meta-analysis. Arch. Phys. Med. Rehabil. 2014, 95, 753–769.e753. [Google Scholar] [CrossRef]
- Gudlaugsson, J.; Gudnason, V.; Aspelund, T.; Siggeirsdottir, K.; Olafsdottir, A.S.; Jonsson, P.V.; Arngrimsson, S.A.; Harris, T.B.; Johannsson, E. Effects of a 6-month multimodal training intervention on retention of functional fitness in older adults: A randomized-controlled cross-over design. Int. J. Behav. Nutr. Phys. Act. 2012, 9, 107. [Google Scholar] [CrossRef] [Green Version]
- Jeon, M.Y.; Jeong, H.; Petrofsky, J.; Lee, H.; Yim, J. Effects of a randomized controlled recurrent fall prevention program on risk factors for falls in frail elderly living at home in rural communities. Med. Sci. Monit. 2014, 20, 2283–2291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.C.; Chang, K.C.; Tsauo, J.Y.; Hung, J.W.; Huang, Y.C.; Lin, S.I.; Fall Prevention Initiatives in Taiwan (FPIT) Investigators. Effects of a multifactorial fall prevention program on fall incidence and physical function in community-dwelling older adults with risk of falls. Arch. Phys. Med. Rehabil. 2013, 94, 606–615.e601. [Google Scholar] [CrossRef]
- Rosendahl, E.; Lindelöf, N.; Littbrand, H.; Yifter-Lindgren, E.; Lundin-Olsson, L.; Håglin, L.; Gustafson, Y.; Nyberg, L. High-intensity functional exercise program and protein-enriched energy supplement for older persons dependent in activities of daily living: A randomised controlled trial. Aust. J. Physiother. 2006, 52, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Serra-Rexach, J.A.; Bustamante-Ara, N.; Villarán, M.H.; Gil, P.G.; Ibáñez, M.J.S.; Bsc, N.B.S.; Bsc, V.O.S.; Bsc, N.G.S.; Bsc, A.B.M.P.; Gallardo, C.; et al. Short-term, light- to moderate-intensity exercise training improves leg muscle strength in the oldest old: A randomized controlled trial. J. Am. Geriatr. Soc. 2011, 59, 594–602. [Google Scholar] [CrossRef]
- Zech, A.; Drey, M.; Freiberger, E.; Hentschke, C.; Bauer, J.M.; Sieber, C.C.; Pfeifer, K. Residual effects of muscle strength and muscle power training and detraining on physical function in community-dwelling prefrail older adults: A randomized controlled trial. BMC Geriatr. 2012, 12, 68. [Google Scholar] [CrossRef] [Green Version]
- Cesari, M.; Vellas, B.; Hsu, F.C.; Newman, A.B.; Doss, H.; King, A.C.; Manini, T.M.; Church, T.; Gill, T.M.; Miller, M.E.; et al. A physical activity intervention to treat the frailty syndrome in older persons-results from the LIFE-P study. J. Gerontol. A Biol. Sci. Med. Sci. 2015, 70, 216–222. [Google Scholar] [CrossRef] [PubMed]
- Fielding, R.A.; Guralnik, J.M.; King, A.C.; Pahor, M.; McDermott, M.M.; Tudor-Locke, C.; Manini, T.M.; Glynn, N.W.; Marsh, A.P.; Axtell, R.S.; et al. Dose of physical activity, physical functioning and disability risk in mobility-limited older adults: Results from the LIFE study randomized trial. PLoS ONE 2017, 12, e0182155. [Google Scholar] [CrossRef] [PubMed]
- Ferrucci, L.; Levine, M.E.; Kuo, P.L.; Simonsick, E.M. Time and the Metrics of Aging. Circ. Res. 2018, 123, 740–744. [Google Scholar] [CrossRef] [PubMed]
- Derbre, F.; Gratas-Delamarche, A.; Gomez-Cabrera, M.C.; Viña, J. Inactivity-induced oxidative stress: A central role in sarcopenia? Eur. J. Sport Sci. 2012, 14, S98–S108. [Google Scholar] [CrossRef] [PubMed]
- Barrès, R.; Yan, J.; Egan, B.; Treebak, J.T.; Rasmussen, M.; Fritz, T.; Caidahl, K.; Krook, A.; O’Gorman, D.J.; Zierath, J.R. Acute exercise remodels promoter methylation in human skeletal muscle. Cell Metab. 2012, 15, 405–411. [Google Scholar] [CrossRef] [Green Version]
- Lindholm, M.E.; Marabita, F.; Gomez-Cabrero, D.; Rundqvist, H.; Ekström, T.J.; Tegnér, J.; Sundberg, C.J. An integrative analysis reveals coordinated reprogramming of the epigenome and the transcriptome in human skeletal muscle after training. Epigenetics 2014, 9, 1557–1569. [Google Scholar] [CrossRef]
- Sailani, M.R.; Halling, J.F.; Møller, H.D.; Lee, H.; Plomgaard, P.; Pilegaard, H.; Snyder, M.P.; Regenberg, B. Lifelong physical activity is associated with promoter hypomethylation of genes involved in metabolism, myogenesis, contractile properties and oxidative stress resistance in aged human skeletal muscle. Sci. Rep. 2019, 9, 3272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gensous, N.; Bacalini, M.G.; Franceschi, C.; Meskers, C.G.M.; Maier, A.B.; Garagnani, P. Age-Related DNA Methylation Changes: Potential Impact on Skeletal Muscle Aging in Humans. Front. Physiol. 2019, 10, 996. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Epigenetic Mechanism | Epigenetic Biomarker Associated to Frailty | Biospecimen | Association between the Epigenetic Change and Frailty | Reference |
|---|---|---|---|---|
| DNA methylation | Horvath’s clock | Leukocytes | Epigenetic age acceleration was correlated with clinically relevant aging-related phenotypes | [54] |
| Histone PTMs | H3K9me3 and H3K9ac and H3K27ac decreases with age | Muscle samples in rat models | Muscle loss and sarcopenia | [79] |
| γH2AX | Leukocytes | Significantly higher γH2AX values observed in individuals positive for low physical activity (p < 0.001), slow waking (p < 0.01), and low grip strength (p < 0.01) criteria | [81] | |
| Non-coding RNAs | miR-10a-3p, miR-92a-3p, miR-185–3p, miR-194–5p, miR-326, miR-532–5p, miR-576–5p, miR-760 | Plasma exosome-derived miRNAs | Associated to frailty | [82] |
| miR-21, miR-223, miR-483 | Plasma | Increased expression in frail subjects compared to control subjects | [83] | |
| miR-146a | Plasma | Low levels in frail subjects compared to robust old adults | [83] | |
| miR-1, miR-21, miR-34a, miR-146a, miR-185, miR-206, miR-223 | Plasma | Increased levels in frail subjects compared to robust old adults | [69] | |
| miR-34a-5p miR-449b-5p | Muscle biopsy | Elevated in sarcopenic muscle compared with muscle tissue from controls | [87] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Giménez, J.L.; Mena-Molla, S.; Tarazona-Santabalbina, F.J.; Viña, J.; Gomez-Cabrera, M.C.; Pallardó, F.V. Implementing Precision Medicine in Human Frailty through Epigenetic Biomarkers. Int. J. Environ. Res. Public Health 2021, 18, 1883. https://doi.org/10.3390/ijerph18041883
García-Giménez JL, Mena-Molla S, Tarazona-Santabalbina FJ, Viña J, Gomez-Cabrera MC, Pallardó FV. Implementing Precision Medicine in Human Frailty through Epigenetic Biomarkers. International Journal of Environmental Research and Public Health. 2021; 18(4):1883. https://doi.org/10.3390/ijerph18041883
Chicago/Turabian StyleGarcía-Giménez, José Luis, Salvador Mena-Molla, Francisco José Tarazona-Santabalbina, Jose Viña, Mari Carmen Gomez-Cabrera, and Federico V. Pallardó. 2021. "Implementing Precision Medicine in Human Frailty through Epigenetic Biomarkers" International Journal of Environmental Research and Public Health 18, no. 4: 1883. https://doi.org/10.3390/ijerph18041883
APA StyleGarcía-Giménez, J. L., Mena-Molla, S., Tarazona-Santabalbina, F. J., Viña, J., Gomez-Cabrera, M. C., & Pallardó, F. V. (2021). Implementing Precision Medicine in Human Frailty through Epigenetic Biomarkers. International Journal of Environmental Research and Public Health, 18(4), 1883. https://doi.org/10.3390/ijerph18041883