Effect of β-Glucan and Black Tea in a Functional Bread on Short Chain Fatty Acid Production by the Gut Microbiota in a Gut Digestion/Fermentation Model †



Abstract
:1. Introduction
2. Materials and Methods
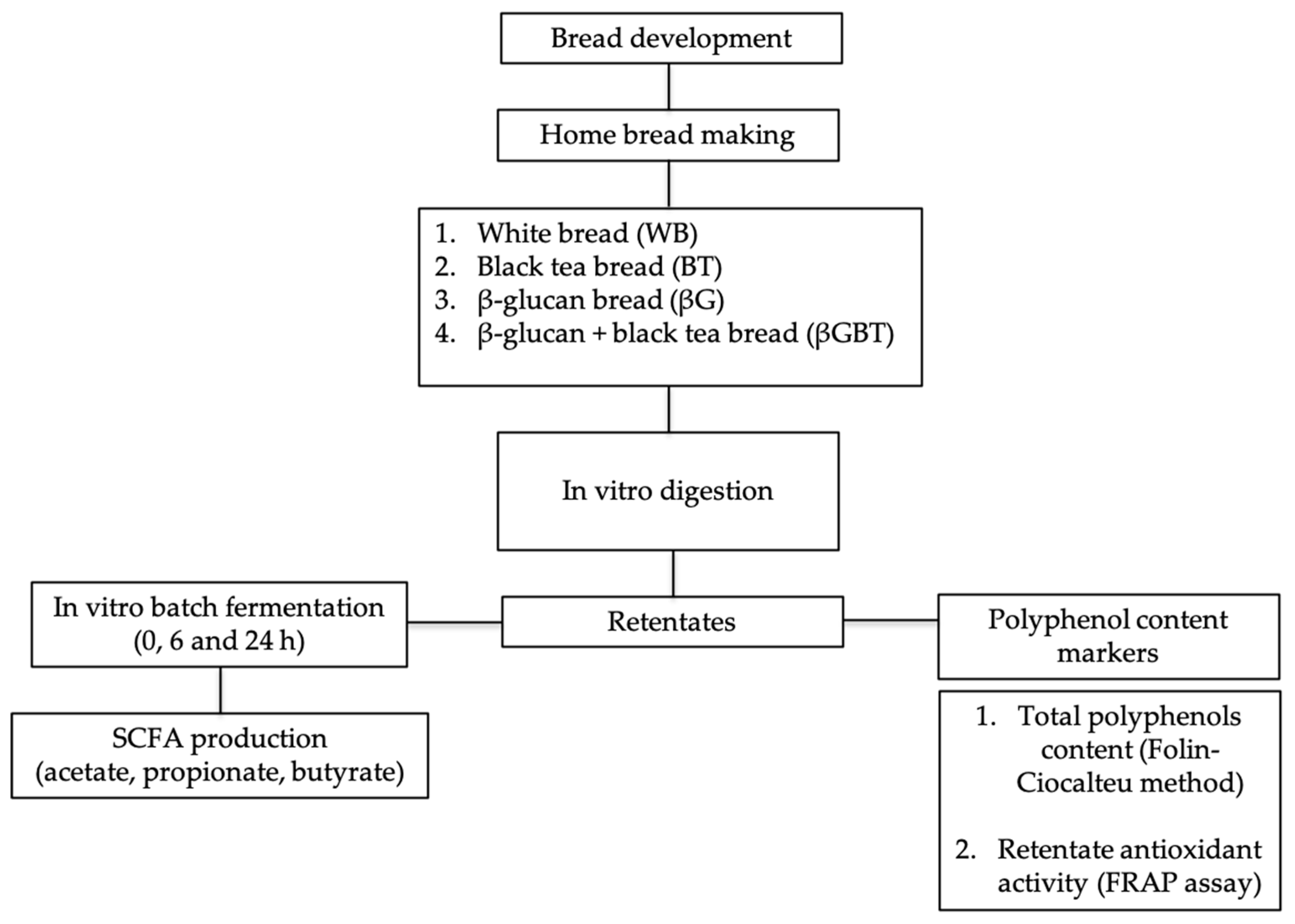
2.1. Study Design
2.2. Bread Development
2.2.1. Materials
2.2.2. Bread Preparation
2.3. In Vitro Digestion Model
- (i)
- Oral phase: Bread samples containing 50 mg available carbohydrate were incubated with α-amylase (50 U/sample) (Sigma-Aldrich, Dorset, UK) at 37 °C for 5 min (mimicking oral chewing).
- (ii)
- Gastric phase: The pH of the mixture was adjusted to 2.5 using HCL (0.15 M, pH 2.5) and incubated with pepsin (0.7 mL, 2 mg/mL in 0.02 M HCl) at 37 °C for 2 h in a shaking water bath.
- (iii)
- Small intestine (duodenum and ileum) phase: The pH was adjusted to 7.0 using sodium hydroxide (6 M). A porcine extract bile acid (Sigma B8631) (2.7 mL, 150 ng/mL in 0.15 M sodium bicarbonate) and pancreatin (mixture of amylase, proteases and lipase, 2.7 mL, 75 mg/mL in 0.15 M sodium bicarbonate) (Sigma-Aldrich, Dorset, UK) were added and incubated at 37 °C for 4 h on a shaking water bath. Absorption in the small intestine was simulated after digestion using dialysis tubing (molecular weight 500–1000 Dalton cut-off, 35 cm long with flat width of 31 mm and diameter of 20 mm) (Spectrum Laboratories, Rancho Dominguez, CA, USA) for 6 h in 2 L distilled water.
- (iv)
- The non-digested/dialyzed products (retentate) were carefully removed from the dialysis tube and freeze-dried. The freeze-dried sample was used as substrate in the in vitro fermentation model.
2.4. Determination of Total Polyphenols
2.5. Antioxidant Potential in Retentates—Ferric Reducing Ability of Plasma (FRAP) Assay
2.6. In Vitro Batch Fermentation
2.7. Statistical Analysis
3. Results
3.1. Total Polyphenols and Antioxidant Potential
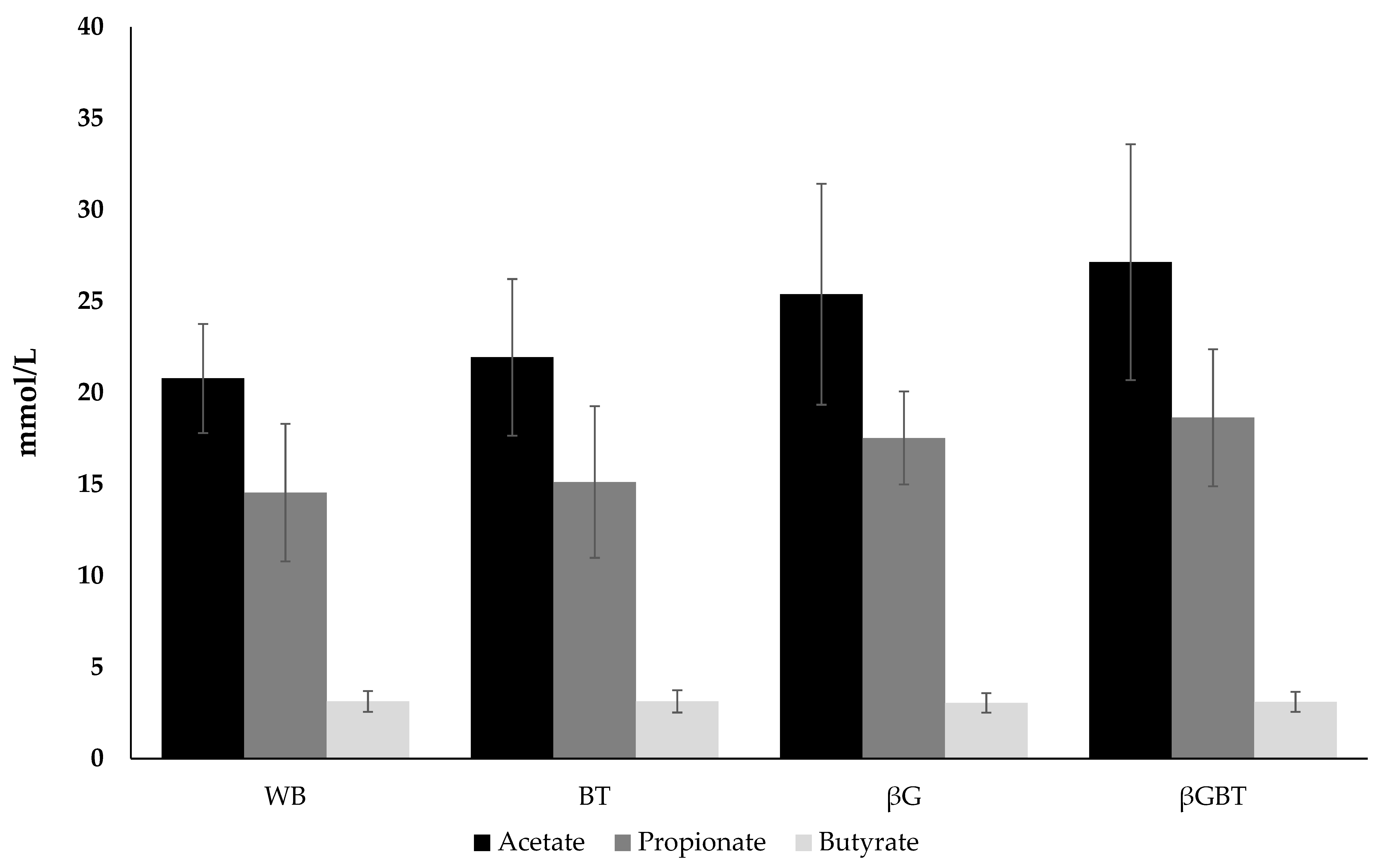
3.2. Short Chain Fatty Acid Production
3.2.1. Fermentation pH
3.2.2. Effects of Adding β-Glucan in Bread on SCFA Production
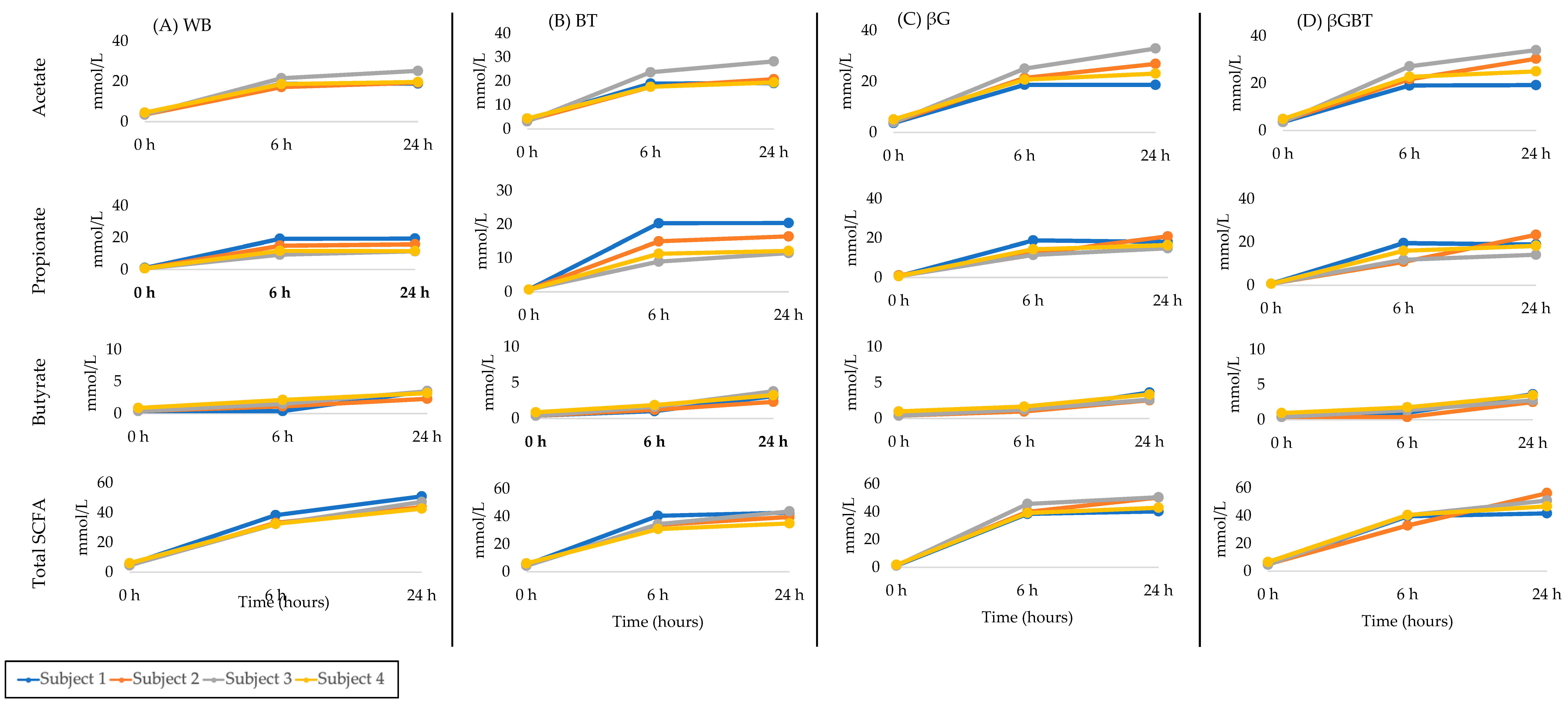
3.2.3. Donor SCFA Variability
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rosell, C.M. Chapter 1—The science of doughs and bread quality. In Flour and Breads and Their Fortification in Health and Disease Prevention, 1st ed.; Preedy, V.R., Watson, R.R., Patel, V.B., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 3–14. [Google Scholar]
- Hayta, M.; Gamze, Ö. Phytochemical fortification of flour and bread. In Flour and Breads and Their Fortification in Health and Disease Prevention, 1st ed.; Preedy, V.R., Watson, R.R., Patel, V.B., Eds.; Academic Press: San Diego, CA, USA, 2011; pp. 293–300. [Google Scholar]
- Brennan, C.S.; Blake, D.E.; Ellis, P.R.; Schofield, J.D. Effects of guar galactomannan on wheat bread microstructure and on the in vitro and in vivo digestibility of starch in bread. J. Cereal Sci. 1996, 24, 151–160. Available online: https://www.sciencedirect.com/science/article/pii/S073352109690048X (accessed on 6 January 2019). [CrossRef]
- Jalil, A.M.; Edwards, C.A.; Combet, E.; Ibrahim, M.; Garcia, A.L. Combined effects of added beta glucan and black tea in breads on starch functionality. Int. J. Food Sci. Nutr. 2015, 66, 159–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agostoni, C.; Bresson, J.-L.; Fairweather-Tait, S.; Flynn, A.; Golly, I.; Korhonen, H.; Lagiou, P.; Løvik, M.; Marchelli, R.; Martin, A.; et al. Scientific opinion on the substantiation of health claims related to beta-glucans from oats and barley and maintenance of normal blood LDL-cholesterol concentrations (ID 1236, 1299), increase in satiety leading to a reduction in energy intake (ID 851, 852), reduction of post-prandial glycaemic responses (ID 821, 824), and “digestive function” (ID 850) pursuant to Article 13(1) of Regulation (EC) No 1924/20061. ESFA J. 2011, 9, 2207–2228. Available online: https://www.efsa.europa.eu/en/efsajournal/pub/2207 (accessed on 6 January 2019).
- Panahi, S.; Ezatagha, A.; Jovanovski, E.; Jenkins, A.; Temelli, F.; Vasanthan, T.; Vuksan, V. Glycemic effect of oat and barley beta-glucan when incorporated into a snack bar: A dose escalation study. J. Am. Coll. Nutr. 2014, 33, 442–449. [Google Scholar] [CrossRef] [PubMed]
- Juvonen, K.R.; Salmenkallio-Marttila, M.; Lyly, M.; Liukkonen, K.H.; Lähteenmäki, L.; Laaksonen, D.E.; Uuusitupa, M.I.; Herzig, K.H.; Poutanen, K.S.; Karhunen, L.J. Semisolid meal enriched in oat bran decreases plasma glucose and insulin levels, but does not change gastrointestinal peptide responses or short-term appetite in healthy subjects. Nutr. Metab. Cardiovasc. Dis. 2011, 21, 748–756. [Google Scholar] [CrossRef] [PubMed]
- Hughes, S.A.; Shewry, P.R.; Gibson, G.R.; McCleary, B.V.; Rastall, R.A. In vitro fermentation of oat and barley derived beta-glucans by human faecal microbiota. FEMS Microbiol. Ecol. 2008, 64, 482–493. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.S.; Morrison, D.J.; Frost, G. Control of appetite and energy intake by SCFA: What are the potential underlying mechanisms? Proc. Nutr. Soc. 2014, 74, 328–336. [Google Scholar] [CrossRef]
- Alhabeeb, H.; Chambers, E.S.; Frost, G.; Morrison, D.J.; Preston, T. Inulin propionate ester increases satiety and decreases appetite but does not affect gastric emptying in healthy humans. Proc. Nutr. Soc. 2014, 73, E21. [Google Scholar] [CrossRef]
- Li, S.H.; Zhao, P.; Tian, H.B.; Chen, L.H.; Cui, L.Q. Effect of grape polyphenols on blood pressure: A Meta-analysis of randomized controlled trials. PLoS ONE 2015, 10, e0137665. [Google Scholar] [CrossRef]
- Chiva-Blanch, G.; Arranz, S.; Lamuela-Raventos, R.M.; Estruch, R. Effects of wine, alcohol and polyphenols on cardiovascular disease risk factors: Evidences from human studies. Alcohol 2013, 48, 270–277. [Google Scholar] [CrossRef]
- Coe, S.; Ryan, L. Impact of polyphenol-rich sources on acute postprandial glycaemia: A systematic review. J. Nutr. Sci. 2016, 5, e24. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Jiménez, J.; Díaz-Rubio, M.E.; Saura-Calixto, F. Non-extractable polyphenols, a major dietary antioxidant: Occurrence, metabolic fate and health effects. Nutr. Res. Rev. 2013, 26, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Bamia, C.; Lagiou, P.; Jenab, M.; Trichopoulou, A.; Fedirko, V.; Aleksandrova, K.; Pischon, T.; Overvad, K.; Olsen, A.; Tjønneland, A.; et al. Coffee, tea and decaffeinated coffee in relation to hepatocellular carcinoma in a European population: Multicentre, prospective cohort study. Int. J. Cancer 2016, 136, 1899–1908. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.T.; Lin-Shiau, S.Y.; Shyur, L.F.; Lin, J.K. Pu-erh tea polysaccharides decrease blood sugar by inhibition of alpha-glucosidase activity in vitro and in mice. Food Funct. 2015, 6, 1539–1546. [Google Scholar] [CrossRef] [PubMed]
- Satoh, T.; Igarashi, M.; Yamada, S.; Takahashi, N.; Watanabe, K. Inhibitory effect of black tea and its combination with acarbose on small intestinal α-glucosidase activity. J. Ethnopharmacol. 2015, 161, 147–155. [Google Scholar] [CrossRef] [PubMed]
- Hertog, M.G.L.; Hollman, P.C.H.; Putte, B.V.D. Content of potentially anticarcinogenic flavonoids of tea infusions, wines, and fruit juices. J. Agric. Food Chem. 1993, 41, 1242–1246. [Google Scholar] [CrossRef]
- Unno, T.; Osakabe, N. Green tea extract and black tea extract differentially influence cecal levels of short-chain fatty acids in rats. Food Sci. Nutr. 2018, 6, 728–735. [Google Scholar] [CrossRef] [Green Version]
- Stalmach, A.; Mullen, W.; Steiling, H.; Williamson, G.; Lean, M.E.; Crozier, A. Absorption, metabolism, and excretion of green tea flavan-3-ols in humans with an ileostomy. Mol. Nutr. Food Res. 2010, 54, 323–334. [Google Scholar] [CrossRef]
- Roowi, S.; Stalmach, A.; Mullen, W.; Lean, M.E.; Edwards, C.A.; Crozier, A. Green tea flavan-3-ols: Colonic degradation and urinary excretion of catabolites by humans. J. Agric. Food Chem. 2010, 58, 1296–1304. [Google Scholar] [CrossRef]
- Auger, C.; Mullen, W.; Hara, Y.; Crozier, A. Bioavailability of polyphenon E flavan-3-ols in humans with an ileostomy. J. Nutr. 2008, 138, 1535s–1542s. [Google Scholar] [CrossRef]
- Chai, Y.; Wang, M.; Zhang, G. Interaction between amylose and tea polyphenols modulates the postprandial glycemic response to high-amylose maize starch. J. Agric. Food Chem. 2013, 61, 8608–8615. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Urpi-Sarda, M.; Boto-Ordoñez, M.; Knox, C.; Llorach, R.; Eisner, R.; Cruz, J.; Neveu, V.; Wishart, D.; Manach, C. Phenol-Explorer 2.0: A major update of the Phenol-Explorer database integrating data on polyphenol metabolism and pharmacokinetics in humans and experimental animals. Database 2012. [Google Scholar] [CrossRef]
- Stewart, A.J.; Mullen, W.; Burns, J.; Lean, M.E.; Brighenti, F.; Crozier, A. HPLC-MS analysis of phenolic compounds and purine alkaloids in green and black tea. J. Agric. Food Chem. 2004, 52, 2807–2815. [Google Scholar] [CrossRef]
- Jacobs, S.M.; Izydorczyk, M.S.; Preston, K.R.; Dexter, J.E. Evaluation of baking procedures for incorporation of barley roller milling fractions containing high levels of dietary fibre into bread. J. Sci. Food Agric. 2008, 88, 558–568. [Google Scholar] [CrossRef]
- Aura, A.-M.; Härkönen, H.; Fabritius, M.; Poutanena, K. Development of an in vitro enzymic digestion method for removal of starch and protein and assessment of its performance using rye and wheat breads. J. Cereal Sci. 1999, 29, 139–152. [Google Scholar] [CrossRef]
- Slinkard, K.; Singleton, V.L. Total phenol analyses: Automation and comparison with manual methods. Am. J. Enol. Vitic. 1977, 28, 49–55. Available online: http://www.ajevonline.org/content/28/1/49 (accessed on 7 January 2019).
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Jaganath, I.; Mullen, W.; Lean, M.; Edwards, C.; Crozier, A. In vitro catabolism of rutin by human fecal bacteria and the antioxidant capacity of its catabolites. Free Radic. Biol. Med. 2009, 47, 1180–1189. [Google Scholar] [CrossRef]
- Metzler-Zebeli, B.U.; Canibe, N.; Montagne, L.; Freire, J. Resistant starch reduces large intestinal pH and promotes fecal lactobacilli and bifidobacterial in pigs. Animal 2019, 13, 64–73. Available online: https://www.ncbi.nlm.nih.gov/pubmed/29745350 (accessed on 6 January 2019). [CrossRef]
- Bown, R.L.; Gibson, J.A.; Sladen, G.E.; Hicks, B.; Dawson, A.M. Effects of lactulose and other laxatives on ileal and colonic pH as measured by a radiotelemetry device. Gut 1974, 15, 999–1004. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1413067/ (accessed on 6 January 2019). [CrossRef] [Green Version]
- Laurentin, A.; Edwards, C.A. Differential fermentation of glucose-based carbohydrates in vitro by human faecal bacteria--a study of pyrodextrinised starches from different sources. Eur. J. Nutr. 2004, 43, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Lin, Q.; Chen, Z.; Xiao, H. The interaction between tea polyphenols and rice starch during gelatinization. Food Sci. Technol. Int. 2011, 17, 569–577. [Google Scholar] [CrossRef] [Green Version]
- Sivam, A.S.; Sun-Waterhouse, D.; Waterhouse, G.I.; Quek, S.; Perera, C.O. Physicochemical properties of bread dough and finished bread with added pectin fiber and phenolic antioxidants. J. Food Sci. 2011, 76, H97–H107. [Google Scholar] [CrossRef] [PubMed]
- Sun-Waterhouse, D.; Chen, J.; Chuah, C.; Wibisono, R.; Melton, L.D.; Laing, W.; Ferguson, L.R.; Skinner, M.A. Kiwi fruit-based polyphenols and related antioxidants for functional foods: Kiwi fruit extract-enhanced gluten-free bread. Int. J. Food Sci. Nutr. 2009, 60 (Suppl. 7), 251–264. [Google Scholar] [CrossRef] [PubMed]
- Simonsen, H.T.; Nielsen, M.S.; Christensen, N.J.; Christensen, U.; La Cour, T.V.; Motawia, M.S.; Jespersen, B.P.; Engelsen, S.B.; Møller, B.L. Molecular interactions between barley and oat beta-glucans and phenolic derivatives. J. Agric. Food Chem. 2009, 57, 2056–2064. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, J.; Chen, F.; Zhao, G. Effects of molecular structure of polyphenols on their noncovalent interactions with oat beta-glucan. J. Agric. Food Chem. 2013, 61, 4533–4538. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Ming, J.; Gao, R.; Wang, Y.; Liang, Q.; Yu, H.; Zhao, G. Characterization and antioxidant activity of the complex of tea polyphenols and oat beta-glucan. J. Agric. Food Chem. 2011, 59, 10737–10746. [Google Scholar] [CrossRef]
- Stalmach, A.; Troufflard, S.; Serafini, M.; Crozier, A. Absorption, metabolism and excretion of Choladi green tea flavan-3-ols by humans. Mol. Nutr. Food Res. 2009, 53 (Suppl. 1), S44–S53. [Google Scholar] [CrossRef]
- Granatoa, D.; Shahidi, F.; Wrolstad, R.; Kilmartin, P.; Melton, L.D.T.; Hidalgo, F.J.; Miyashita, K.; van Camp, J.; Alasalvar, C.; Ismail, A.; et al. Antioxidant activity, total phenolics and flavonoids contents: Should we ban in vitro screening methods? Food Chem. 2018, 264, 471–475. [Google Scholar] [CrossRef]
- Parkar, S.G.; Trower, T.M.; Stevenson, D.E. Fecal microbial metabolism of polyphenols and its effects on human gut microbiota. Anaerobe 2013, 23, 12–19. [Google Scholar] [CrossRef]
- Goodwin, B.L.; Ruthven, C.R.; Sandler, M. Gut flora and the origin of some urinary aromatic phenolic compounds. Biochem. Pharmacol. 1994, 47, 2294–2297. Available online: https://www.sciencedirect.com/science/article/abs/pii/0006295294902682?via%3Dihub (accessed on 7 January 2019). [CrossRef]
- Liu, A.B.; Tao, S.; Lee, M.-J.; Hu, Q.; Meng, X.; Lin, Y.; Yang, C.S. Effects of gut microbiota and time of treatment on tissue levels of green tea polyphenols in mice. Biofactors 2018, 4, 348–360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, Y.; Zhu, J.; Zhang, M.; Ho, C.-T.; Huang, Q.; Cao, J. Metagenomics analysis of gut microbiota in a high fat diet-induced obesity mouse model fed with (-)-epigallocatechin 3-O-(3-O-methyl) gallate (EGCG3’’Me). Mol. Nutr. Food Res. 2018, 62, e1800274. [Google Scholar] [CrossRef] [PubMed]
- Nordlund, E.; Aura, A.M.; Mattila, I.; Kössö, T.; Rouau, X.; Poutanen, K. Formation of phenolic microbial metabolites and short-chain fatty acids from rye, wheat, and oat bran and their fractions in the metabolical in vitro colon model. J. Agric. Food Chem. 2012, 60, 8134–8145. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; White, P.J. Optimizing the molecular weight of oat beta-glucan for in vitro bile acid binding and fermentation. J. Agric. Food Chem. 2011, 59, 10322–10328. [Google Scholar] [CrossRef] [PubMed]
- Carlson, J.; Esparza, J.; Swan, J.; Taussig, D.; Combs, J.; Slavin, J. In vitro analysis of partially hydrolyzed guar gum fermentation differences between six individuals. Food Funct. 2016, 7, 2833–2838. [Google Scholar] [CrossRef]
- Harris, H.C.; Edwards, C.A.; Morrison, D.J. Impact of glycosidic bond configuration on short chain fatty acid production from model fermentable carbohydrates by the human gut microbiota. Nutrients 2017, 9, 26. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T.; Macfarlane, S.; Gibson, G.R. Validation of a three-stage compound continuous culture system for investigating the effect of retention time on the ecology and metabolism of bacteria in the human colon. Microb. Ecol. 1998, 35, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.S.; Li, L.; Bang, J.; Han, N.S. Application of in vitro gut fermentation models to food components: A review. Food Sci. Biotechnol. 2016, 25, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Cummings, J.H.; Pomare, E.W.; Branch, W.J.; Naylor, C.P.E.; Macfarlane, G.T. Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 1987, 28, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.S.; Viardot, A.; Psichas, A.; Morrison, D.J.; Murphy, K.G.; Zac-Varghese, S.E.; MacDougall, K.; Preston, T.; Tedford, C.; Finlayson, G.S. Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut 2015, 64, 1744–1754. [Google Scholar] [CrossRef] [PubMed]
- Psichas, A.; Sleeth, M.L.; Murphy, K.G.; Brooks, L.; Bewick, G.A.; Hanyaloglu, A.C.; Ghatei, M.A.; Bloom, S.R.; Frost, G. The short chain fatty acid propionate stimulates GLP-1 and PYY secretion via free fatty acid receptor 2 in rodents. Int. J. Obes. (Lond.) 2015, 39, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Karaki, S.; Tazoe, H.; Hayashi, H.; Kashiwabara, H.; Tooyama, K.; Suzuki, Y.; Kuwahara, A. Expression of the short-chain fatty acid receptor, GPR43, in the human colon. J. Mol. Histol. 2008, 39, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.G.; Dhillo, W.S.; Bloom, S.R. Gut peptides in the regulation of food intake and energy homeostasis. Endocr. Rev. 2006, 27, 719–727. [Google Scholar] [CrossRef] [PubMed]
- Batterham, R.L.; Cowley, M.A.; Small, C.J.; Herzog, H.; Cohen, M.A.; Dakin, C.L.; Wren, A.M.; Brynes, A.E.; Low, M.J.; Ghatei, M.A.; et al. Gut hormone PYY (3-36) physiologically inhibits food intake. Nature 2002, 418, 650–654. [Google Scholar] [CrossRef] [PubMed]
- Åman, P.; Lena, R.; Roger, A. Molecular weight distribution of beta-glucan in oat-based foods. Cereal Chem. 2004, 81, 356–360. [Google Scholar] [CrossRef]
- Verbeke, K.; Ferchaud-Roucher, V.; Preston, T.; Small, A.C.; Henckaerts, L.; Krempf, M.; Wang, H.; Vonk, R.J.; Priebe, M.G. Influence of the type of indigestible carbohydrate on plasma and urine short-chain fatty acid profiles in healthy human volunteers. Eur. J. Clin. Nutr. 2010, 64, 678–684. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Ingredient (g) | White Bread (WB) | Black Tea Bread (BT) | β-Glucan Bread (βG) | β-Glucan + Black Tea Bread (βGBT) |
|---|---|---|---|---|
| Strong white wheat flour | 500.0 | 500.0 | 500.0 | 500.0 |
| NaCl | 8.0 | 8.0 | 8.0 | 8.0 |
| Sugar | 6.0 | 6.0 | 6.0 | 6.0 |
| Dehydrated yeast | 8.0 | 8.0 | 10.0 | 10.0 |
| Butter (unsalted) | 6.0 | 6.0 | 6.0 | 6.0 |
| Skimmed milk powder | 7.0 | 7.0 | 7.0 | 7.0 |
| β-glucan | 0.0 | 0.0 | 35.0 | 35.0 |
| Black tea | 0.0 | 2.5 | 0.0 | 2.5 |
| Water (Ml) | 300 | 300 | 540 | 540 |
| Total | 835.0 | 837.5 | 1112.0 | 1114.5 |
| Bread | Total Polyphenols (μg GAE/g Retentate) | FRAP Activity of Retentate (μg Fe2+ Equivalents/g Retentate) | ||
|---|---|---|---|---|
| * Pre-Digested | ** Digested | * Pre-Digested | ** Digested | |
| WB | 222.6 ± 63.0 ade | 1077.7 ± 250.2 ab | 338.9 ± 131.2 ab | 3633.3 ± 645.0 a |
| BT | 361.1 ± 40.0 bc | 1228.9 ± 106.0 ac | 1056.2 ± 131.4 c | 4024.5 ± 198.8 a |
| βG | 229.1 ± 0.0 d | 1298.3 ± 276.0 ac | 236.2 ± 80.4 b | 3351.9 ± 552.2 a |
| βGBT | 291.5 ± 0.0 e | 1379.9 ± 146.8 c | 608.0 ± 119.0 d | 3998.8 ± 1030.4 a |
| Bread | 0 h | 6 h | 24 h |
|---|---|---|---|
| WB | 6.6 ± 0.2 | 5.5 ± 0.2 | 5.8 ± 0.4 |
| BT | 6.6 ± 0.4 | 5.4 ± 0.4 | 5.7 ± 0.8 |
| βG | 6.6 ± 0.4 | 5.1 ± 0.8 | 5.3 ± 1.0 |
| βGBT | 6.6 ± 0.4 | 5.0 ± 0.8 | 5.2 ± 1.0 |
| Bread | Individual SCFA by donor | |||
|---|---|---|---|---|
| Acetate | Propionate | Butyrate | Total | |
| Donor 1 | BT = βGBT > WB > βG | BT > WB > βGBT > βG | βGBT = βG > WB > BT | BT > WB > βGBT > βG |
| Donor 2 | βGBT > βG > BT > WB | βGBT > βG > BT > WB | βG > βGBT > BT > WB | βGBT > βG > BT > WB |
| Donor 3 | βGBT > βG > BT > WB | βG > βGBT > BT > WB | BT > WB > βG > βGBT | βGBT > βG > BT > WB |
| Donor 4 | βGBT > βG > WB > BT | βGBT > βG > BT > WB | βGBT > βG > BT > WB | βGBT > βG > BT > WB |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mhd Jalil, A.M.; Combet, E.; Edwards, C.A.; Garcia, A.L. Effect of β-Glucan and Black Tea in a Functional Bread on Short Chain Fatty Acid Production by the Gut Microbiota in a Gut Digestion/Fermentation Model. Int. J. Environ. Res. Public Health 2019, 16, 227. https://doi.org/10.3390/ijerph16020227
Mhd Jalil AM, Combet E, Edwards CA, Garcia AL. Effect of β-Glucan and Black Tea in a Functional Bread on Short Chain Fatty Acid Production by the Gut Microbiota in a Gut Digestion/Fermentation Model. International Journal of Environmental Research and Public Health. 2019; 16(2):227. https://doi.org/10.3390/ijerph16020227
Chicago/Turabian StyleMhd Jalil, Abbe M., Emilie Combet, Christine A. Edwards, and Ada L. Garcia. 2019. "Effect of β-Glucan and Black Tea in a Functional Bread on Short Chain Fatty Acid Production by the Gut Microbiota in a Gut Digestion/Fermentation Model" International Journal of Environmental Research and Public Health 16, no. 2: 227. https://doi.org/10.3390/ijerph16020227