Investigating the Current and Future Co-Occurrence of Ambrosia artemisiifolia and Ophraella communa in Europe through Ecological Modelling and Remote Sensing Data Analysis


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species
2.2. Dataset and Study Area
2.3. Ecological Niche Modelling
2.4. Post-Modelling Analyses
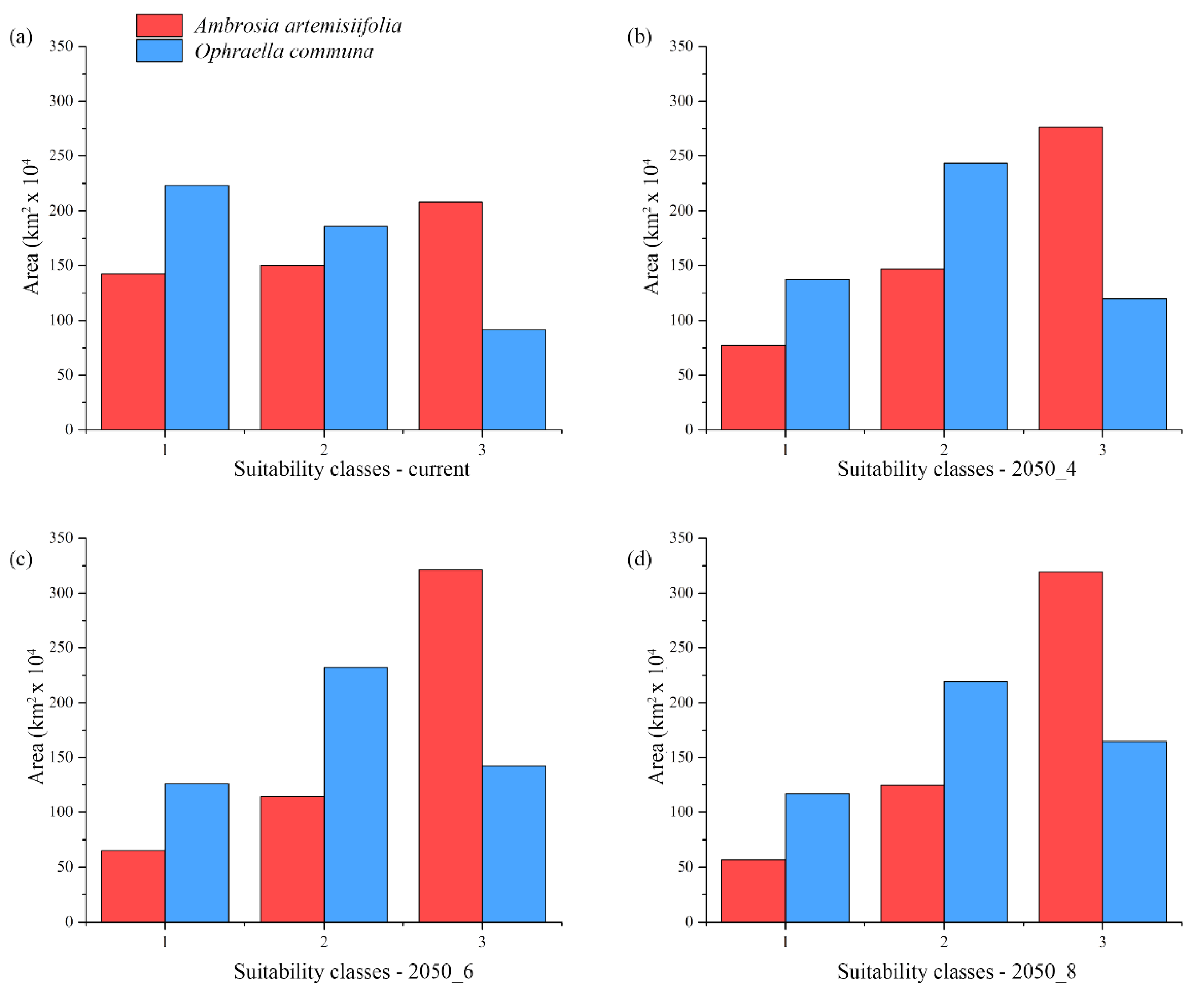
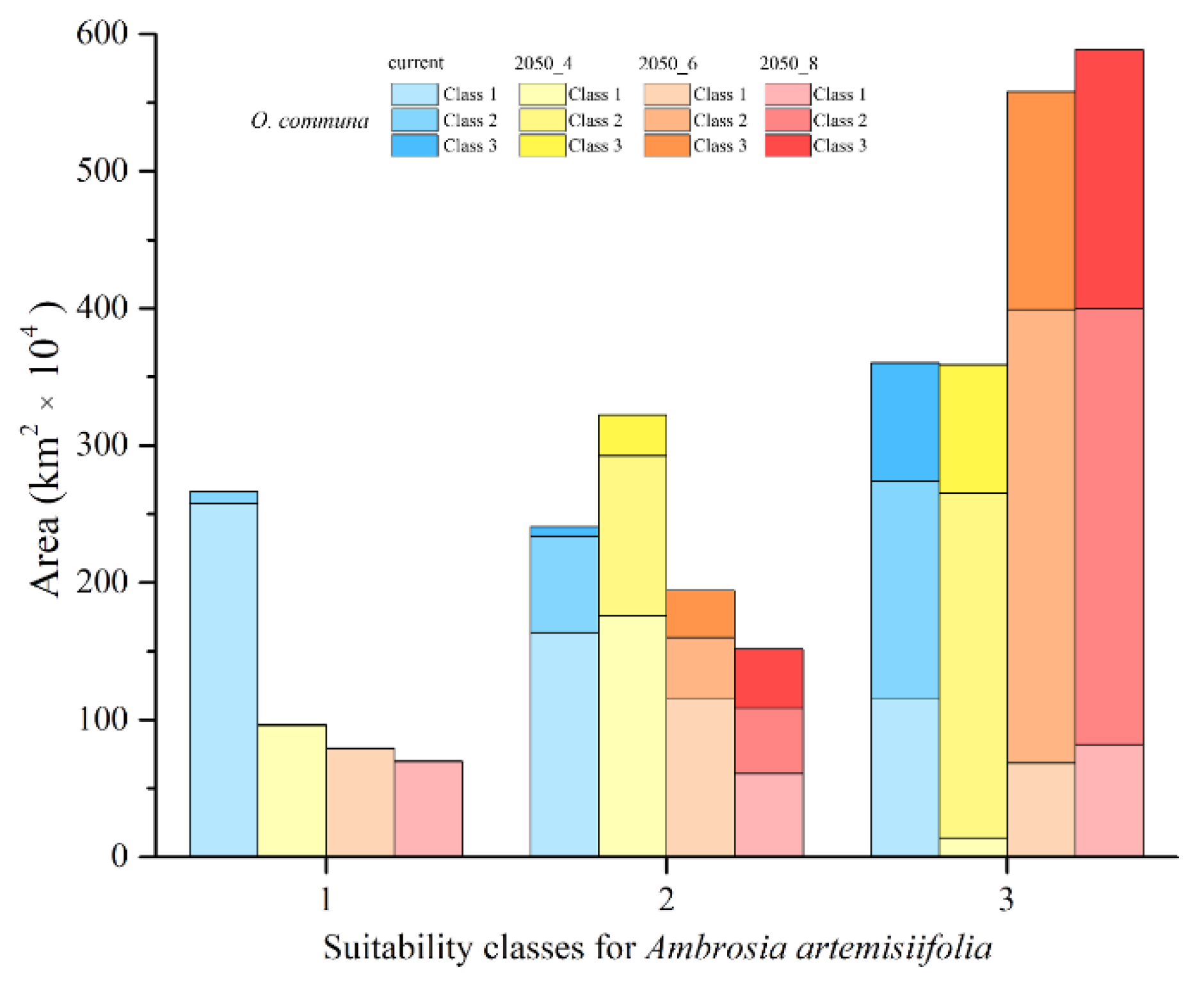
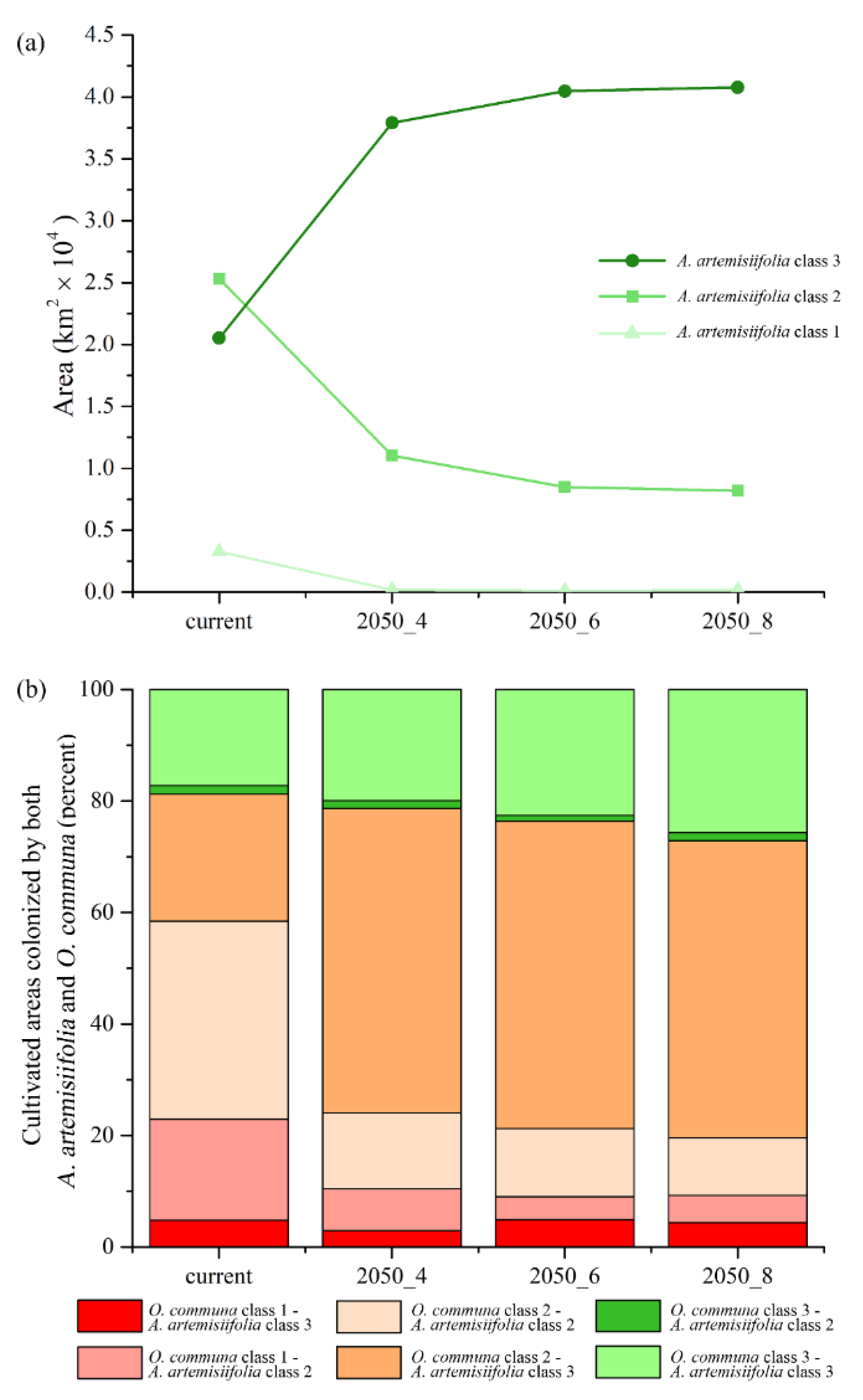
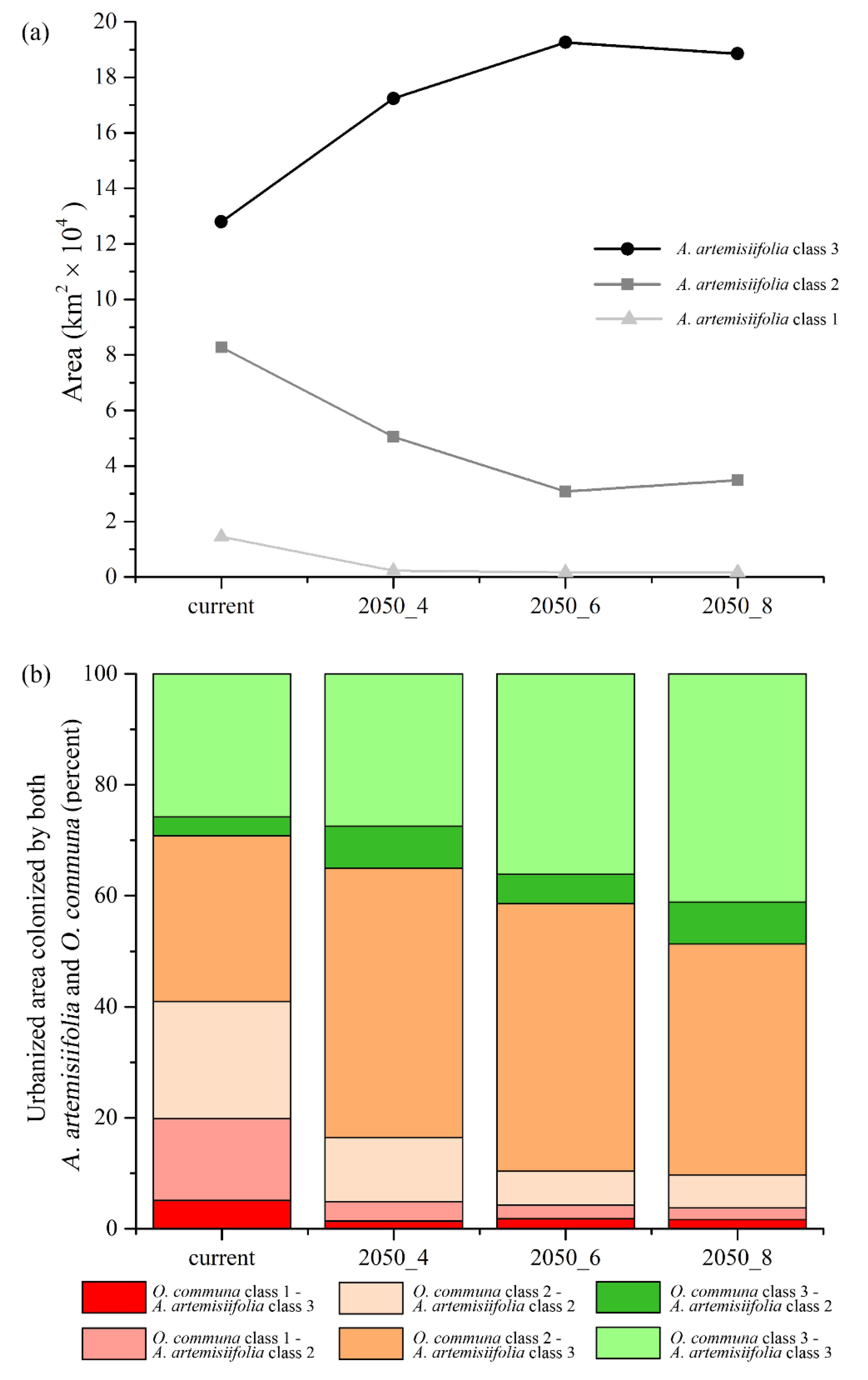
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cerasoli, F.; Iannella, M.; Biondi, M. Between the hammer and the anvil: How the combined effect of global warming and the non-native common slider could threaten the European pond turtle. Manag. Biol. Invasions 2019, 10, 428–448. [Google Scholar] [CrossRef]
- Doherty, T.S.; Dickman, C.R.; Nimmo, D.G.; Ritchie, E.G. Multiple threats, or multiplying the threats? Interactions between invasive predators and other ecological disturbances. Biol. Conserv. 2015, 190, 60–68. [Google Scholar] [CrossRef]
- Gallardo, B.; Clavero, M.; Sánchez, M.I.; Vilà, M. Global ecological impacts of invasive species in aquatic ecosystems. Glob. Chang. Biol. 2016, 22, 151–163. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.R.; Carpenter, S.R.; Vander Zanden, M.J. Invasive species triggers a massive loss of ecosystem services through a trophic cascade. Proc. Natl. Acad. Sci. USA 2016, 113, 4081–4085. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beggs, P.J. Impacts of climate change on aeroallergens: Past and future. Clin. Exp. Allergy 2004, 34, 1507–1513. [Google Scholar] [CrossRef] [PubMed]
- Juliano, S.A.; Lounibos, L.P. Ecology of invasive mosquitoes: Effects on resident species and on human health. Ecol. Lett. 2005, 8, 558–574. [Google Scholar] [CrossRef] [PubMed]
- Lake, I.R.; Jones, N.R.; Agnew, M.; Goodess, C.M.; Giorgi, F.; Hamaoui-Laguel, L.; Semenov, M.A.; Solomon, F.; Storkey, J.; Vautard, R.; et al. Climate Change and Future Pollen Allergy in Europe. Environ. Health Perspect. 2016, 125, 385–391. [Google Scholar] [CrossRef]
- Bradshaw, C.J.A.; Leroy, B.; Bellard, C.; Roiz, D.; Albert, C.; Fournier, A.; Barbet-Massin, M.; Salles, J.M.; Simard, F.; Courchamp, F. Massive yet grossly underestimated global costs of invasive insects. Nat. Commun. 2016, 7, 12986. [Google Scholar] [CrossRef]
- Crowley, S.L.; Hinchliffe, S.; McDonald, R.A. Conflict in invasive species management. Front. Ecol. Environ. 2017, 15, 133–141. [Google Scholar] [CrossRef] [Green Version]
- Gilioli, G.; Schrader, G.; Carlsson, N.; Van Donk, E.; Van Leeuwen, C.H.; Martín, P.R.; Pasquali, S.; Vilà, M.; Vos, S. Environmental risk assessment for invasive alien species: A case study of apple snails affecting ecosystem services in Europe. Environ. Impact Assess. Rev. 2017, 65, 1–11. [Google Scholar] [CrossRef]
- Oreska, M.P.; Aldridge, D.C. Estimating the financial costs of freshwater invasive species in Great Britain: A standardized approach to invasive species costing. Biol. Invasions 2011, 13, 305–319. [Google Scholar] [CrossRef]
- Padilha, B.; Vinicius, F. Current distribution of invasive feral pigs in Brazil: Economic impacts and ecological uncertainty. Nat. Conserv. 2015, 13, 84–87. [Google Scholar]
- Soberón, J.; Peterson, A.T. Interpretation of Models of Fundamental Ecological Niches and Species’ Distributional Areas. Biodivers. Inform. 2005, 2, 1–10. [Google Scholar] [CrossRef]
- Lei, J.; Chen, L.; Li, H. Using ensemble forecasting to examine how climate change promotes worldwide invasion of the golden apple snail (Pomacea canaliculata). Environ. Monit. Assess. 2017, 189, 404. [Google Scholar] [CrossRef]
- Ricciardi, A.; Blackburn, T.M.; Carlton, J.T.; Dick, J.T.; Hulme, P.E.; Iacarella, J.C.; Jeschke, J.M.; Liebhold, A.M.; Lockwood, J.L.; MacIsaac, H.J.; et al. Invasion Science: A Horizon Scan of Emerging Challenges and Opportunities. Trends Ecol. Evol. 2017, 32, 464–474. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.H.; Caulfield, F.A. Rising CO2 and pollen production of common ragweed (Ambrosia artemisiifolia L.), a known allergy-inducing species: Implications for public health. Funct. Plant Biol. 2000, 27, 893–898. [Google Scholar] [CrossRef]
- D’Alessandro, P.; Iannella, M.; Frasca, R.; Biondi, M. Distribution patterns and habitat preference for the genera-group Blepharida s.l. in Sub-Saharan Africa (Coleoptera: Chrysomelidae: Galerucinae: Alticini). ZooL. Anz. A J. Comp. ZooL. 2018, 277, 23–32. [Google Scholar] [CrossRef]
- Iannella, M.; Cerasoli, F.; D’Alessandro, P.; Console, G.; Biondi, M. Coupling GIS spatial analysis and Ensemble Niche Modelling to investigate climate change-related threats to the Sicilian pond turtle Emys trinacris, an endangered species from the Mediterranean. PeerJ 2018, 6, e4969. [Google Scholar] [CrossRef]
- Iannella, M.; D’Alessandro, P.; Biondi, M. Evidences for a shared history for spectacled salamanders, haplotypes and climate. Sci. Rep. 2018, 8, 16507. [Google Scholar] [CrossRef]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef] [Green Version]
- Shabani, F.; Kumar, L.; Ahmadi, M. A comparison of absolute performance of different correlative and mechanistic species distribution models in an independent area. Ecol. Evol. 2016, 6, 5973–5986. [Google Scholar] [CrossRef] [Green Version]
- Iannella, M.; D’Alessandro, P.; Longo, S.; Biondi, M. New records and potential distribution by Ecological Niche Modelling of the adventive leaf beetle Monoxia obesula Blake in the Mediterranean area (Coleoptera, Chrysomelidae, Galerucinae). Bull. Insectology 2019, 72, 135–142. [Google Scholar]
- Bosio, G.; Massobrio, V.; Chersi, C.; Scavarda, G.; Clark, S. Spread of the ragweed leaf beetle, Ophraella communa LeSage, 1986 (Coleoptera Chrysomelidae), in Piedmont Region (northwestern Italy). Boll. Della Soc. Entomol. Ital. 2014, 146, 17–30. [Google Scholar] [CrossRef]
- Komíves, T.; Béres, I.; Reisinger, P.; Lehoczky, E.; Berke, J.; Tamás, J.; Páldy, A.; Csornai, G.; Nádor, G.; Kardeván, P. New strategy of the integrated protection against common ragweed (Ambrosia artemisiifolia L.). Hung. Weed Res. Technol. 2006, 7, 5–49. [Google Scholar]
- Lommen, S.T.; Ciappetta, S.; Ghiani, A.; Asero, R.; Gentili, R.; Müller-Schärer, H.; Citterio, S. Defoliation of common ragweed by Ophraella communa beetle does not affect pollen allergenicity in controlled conditions. Plant Biosyst. 2017, 151, 1094–1100. [Google Scholar] [CrossRef]
- Bonini, M.; Šikoparija, B.; Prentović, M.; Cislaghi, G.; Colombo, P.; Testoni, C.; Lommen, S.; Müller-Schärer, H.; Smith, M. A follow-up study examining airborne Ambrosia pollen in the Milan area in 2014 in relation to the accidental introduction of the ragweed leaf beetle Ophraella communa. Aerobiologia 2016, 32, 371–374. [Google Scholar] [CrossRef]
- Plank, L.; Zak, D.; Getzner, M.; Follak, S.; Essl, F.; Dullinger, S.; Kleinbauer, I.; Moser, D.; Gattringer, A. Benefits and costs of controlling three allergenic alien species under climate change and dispersal scenarios in Central Europe. Environ. Sci. Policy 2016, 56, 9–21. [Google Scholar] [CrossRef]
- Buters, J.; Alberternst, B.; Nawrath, S.; Wimmer, M.; Traidl-Hoffmann, C.; Starfinger, U.; Behrendt, H.; Schmidt-Weber, C.; Bergmann, K.-C. Ambrosia artemisiifolia (ragweed) in Germany—Current presence, allergological relevance and containment procedures. Allergo J. Int. 2015, 24, 108–120. [Google Scholar] [CrossRef]
- Gerber, E.; Schaffner, U.; Gassmann, A.; Hinz, H.L.; Seier, M.; Müller-Schärer, H. Prospects for biological control of Ambrosia artemisiifolia in Europe: Learning from the past. Weed Res. 2011, 51, 559–573. [Google Scholar] [CrossRef]
- Karrer, G.; Skjøth, C.A.; Šikoparija, B.; Smith, M.; Berger, U.; Essl, F. Ragweed (Ambrosia) pollen source inventory for Austria. Sci. Total Environ. 2015, 523, 120–128. [Google Scholar] [CrossRef]
- Rasmussen, K.; Thyrring, J.; Muscarella, R.; Borchsenius, F. Climate-change-induced range shifts of three allergenic ragweeds (Ambrosia L.) in Europe and their potential impact on human health. PeerJ 2017, 5, e3104. [Google Scholar] [CrossRef]
- SMARTER project Sustainable Management of Ambrosia artemisiifolia in Europe. In Proceedings of the Sustainable Management of Ambrosia artemisiifolia in Europe Final Conference, Vianden, Luxembourg, 11–13 September 2016.
- Vincent, G.; Deslauriers, S.; Cloutier, D. Problems and eradication of Ambrosia artemisiifolia L. in Quebec in the urban and suburban environments. Allerg. Immunol. 1992, 24, 84–89. [Google Scholar]
- Nishide, Y.; Fukano, Y.; Doi, H.; Satoh, T.; Inoue, H.; Boriani, M. Origins and genetic diversity of the ragweed beetles, Ophraella communa (Coleoptera: Chrysomelidae), that were introduced into Italy and Japan based on an analysis of mitochondrial DNA sequence data. Eur. J. Èntomol. 2015, 112, 613–618. [Google Scholar] [CrossRef]
- Buttenschøn, R.M.; Waldispühl, S.; Bohren, C. Guidelines for Management of Common Ragweed, Ambrosia artemisiifolia; Euphresco Report; University of Copenhagen: Copenhagen, Denmark, 2010. [Google Scholar]
- Sun, Y.; Zhou, Z.; Wang, R.; Müller-Schärer, H. Biological control opportunities of ragweed are predicted to decrease with climate change in East Asia. Biodivers. Sci. 2017, 25, 1285–1294. [Google Scholar] [Green Version]
- Lommen, S.T.; Jolidon, E.F.; Sun, Y.; Eduardo, J.B. An early suitability assessment of two exotic Ophraella species (Coleoptera: Chrysomelidae) for biological control of invasive ragweed in Europe. Eur. J. Èntomol. 2017, 114, 160–169. [Google Scholar] [CrossRef]
- Meng, L.; Li, B. Advances on biology and host specificity of the newly introduced beetle, Ophraella communa Lesage (Coleoptera: Chrysomelidae), attacking Ambrosia artemisiifolia (Compositae) in continent of China. Chin. J. Biol. Control 2005, 21, 65–69. [Google Scholar]
- Zhou, Z.S.; Chen, H.S.; Zheng, X.W.; Guo, J.Y.; Guo, W.; Li, M.; Luo, M.; Wan, F.H. Control of the invasive weed Ambrosia artemisiifolia with Ophraella communa and Epiblema strenuana. Biocontrol Sci. Technol. 2014, 24, 950–964. [Google Scholar] [CrossRef]
- Dernovici, S.A.; Teshler, M.P.; Watson, A.K. Is sunflower (Helianthus annuus) at risk to damage from Ophraella communa, a natural enemy of common ragweed (Ambrosia artemisiifolia)? Biocontrol Sci. Technol. 2006, 16, 669–686. [Google Scholar] [CrossRef]
- Cao, Z.; Wang, H.; Meng, L.; Li, B. Risk to nontarget plants from Ophraella communa (Coleoptera: Chrysomelidae), a potential biological control agent of alien invasive weed Ambrosia artemisiifolia (Asteraceae) in China. Appl. Èntomol. Zool. 2011, 46, 375–381. [Google Scholar] [CrossRef]
- Case, M.J.; Stinson, K.A. Climate change impacts on the distribution of the allergenic plant, common ragweed (Ambrosia artemisiifolia) in the eastern United States. PLoS ONE 2018, 13, e0205677. [Google Scholar] [CrossRef]
- Leiblein, M.C.; Cunze, S.; Tackenberg, O. Range Expansion of Ambrosia artemisiifolia in Europe Is Promoted by Climate Change. ISRN Ecol. 2013, 2013, 610126. [Google Scholar]
- Liu, X.; Li, J.; Zhao, C.; Quan, Z.; Zhao, X.; Gong, L. Prediction of potential suitable area of Ambrosia artemisiifolia L. in China based on MAXENT and ArcGIS. Acta Phytophylacica Sin. 2016, 43, 1041–1048. [Google Scholar]
- Qin, Z.; DiTommaso, A.; Wu, R.S.; Huang, H.Y. Potential distribution of two Ambrosia species in China under projected climate change. Weed Res. 2014, 54, 520–531. [Google Scholar] [CrossRef]
- Cardarelli, E.; Musacchio, A.; Montagnani, C.; Bogliani, G.; Citterio, S.; Gentili, R. Ambrosia artemisiifolia control in agricultural areas: Effect of grassland seeding and herbivory by the exotic leaf beetle Ophraella communa. NeoBiota 2018, 37, 55–76. [Google Scholar] [CrossRef]
- Bonini, M.; Šikoparija, B.; Skjøth, C.; Cislaghi, G.; Colombo, P.; Testoni, C.; Smith, M. Ambrosia pollen source inventory for Italy: A multi-purpose tool to assess the impact of the ragweed leaf beetle (Ophraella communa LeSage) on populations of its host plant. Int. J. Biometeorol. 2018, 62, 597–608. [Google Scholar] [CrossRef]
- Mouttet, R.; Augustinus, B.; Bonini, M.; Chauvel, B.; Desneux, N.; Gachet, E.; Le Bourgeois, T.; Müller-Schärer, H.; Thibaudon, M.; Schaffner, U. Estimating economic benefits of biological control of Ambrosia artemisiifolia by Ophraella communa in southeastern France. Basic Appl. Ecol. 2018, 33, 14–24. [Google Scholar] [CrossRef]
- Müller-Schärer, H.E.; Lommen, S.T.; Rossinelli, M.; Bonini, M.; Boriani, M.; Bosio, G.; Schaffner, U.; Müller-Schärer, H. Ophraella communa, the ragweed leaf beetle, has successfully landed in Europe: Fortunate coincidence or threat? Weed Res. 2014, 54, 109–119. [Google Scholar]
- Sun, Y.; Brönnimann, O.; Roderick, G.K.; Poltavsky, A.; Lommen, S.T.E.; Müller-Schärer, H. Climatic suitability ranking of biological control candidates: A biogeographic approach for ragweed management in Europe. Ecosphere 2017, 8, e01731. [Google Scholar] [CrossRef]
- Sun, Y.; Brönnimann, O.; Müller-Schärer, H. Climatic suitability of the accidentally introduced leaf beetle Ophraella communa in Europe: A potential biological control candidate for ragweed. Not. della Soc. Bot. Ital. 2016, 1, 108110. [Google Scholar]
- Cerasoli, F.; Iannella, M.; D’Alessandro, P.; Biondi, M. Comparing pseudo-absences generation techniques in Boosted Regression Trees models for conservation purposes: A case study on amphibians in a protected area. PLoS ONE 2017, 12, e0187589. [Google Scholar] [CrossRef]
- Brunetti, M.; Magoga, G.; Iannella, M.; Biondi, M.; Montagna, M. Phylogeography and species distribution modelling of Cryptocephalus barii (Coleoptera: Chrysomelidae): Is this alpine endemic species close to extinction? ZooKeys 2019, 856, 3. [Google Scholar] [CrossRef]
- Iannella, M.; D’Alessandro, P.; Biondi, M. Entomological knowledge in Madagascar by GBIF datasets: Estimates on the coverage and possible biases (Insecta). Fragm. Entomol. 2019, 51, 1–10. [Google Scholar] [CrossRef]
- Iannella, M.; Liberatore, L.; Biondi, M. The effects of a sudden urbanization on micromammal communities: A case study of post-earthquake L’Aquila (Abruzzi Region, Italy). Ital. J. Zool. 2016, 83, 1–8. [Google Scholar] [CrossRef]
- Barnes, E.R.; Jhala, A.J.; Knezevic, S.Z.; Sikkema, P.H.; Lindquist, J.L. Common ragweed (Ambrosia artemisiifolia L.) interference with soybean in Nebraska. Agron. J. 2018, 110, 646–653. [Google Scholar] [CrossRef]
- Yamazaki, K.; Imai, C.; Natuhara, Y. Rapid population growth and food-plant exploitation pattern in an exotic leaf beetle, Ophraella communa LeSage (Coleoptera: Chrysomelidae), in western Japan. Appl. Entomol. Zool. 2000, 35, 215–223. [Google Scholar] [CrossRef]
- ESRI, ArcMap 10.0. ESRI, Redlands, California. 2010. [Google Scholar]
- Aiello-Lammens, M.E.; Boria, R.A.; Radosavljevic, A.; Vilela, B.; Anderson, R.P. spThin: An R package for spatial thinning of species occurrence records for use in ecological niche models. Ecography 2015, 38, 541–545. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 15 June 2019).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high-resolution interpolated climate surfaces for global land areas. Int. J. Clim. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J. Collinearity: A review of methods to deal with it and a simulation study evaluating their performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Elith, J.; Graham, C.H.; Anderson, R.P.; Dudík, M.; Ferrier, S.; Guisan, A.; Hijmans, R.J.; Huettmann, F.; Leathwick, J.R.; Lehmann, A.; et al. Novel methods improve prediction of species’ distributions from occurrence data. Ecography 2006, 29, 129–151. [Google Scholar] [CrossRef]
- Gent, P.R.; Danabasoglu, G.; Donner, L.J.; Holland, M.M.; Hunke, E.C.; Jayne, S.R.; Lawrence, D.M.; Neale, R.B.; Rasch, P.J.; Vertenstein, M.; et al. The Community Climate System Model Version 4. J. Clim. 2011, 24, 4973–4991. [Google Scholar] [CrossRef]
- Marti, O.; Braconnot, P.; Dufresne, J.L.; Bellier, J.; Benshila, R.; Bony, S.; Brockmann, P.; Cadule, P.; Caubel, A.; Codron, F. Key features of the IPSL ocean atmosphere model and its sensitivity to atmospheric resolution. Clim. Dyn. 2010, 34, 1–26. [Google Scholar] [CrossRef]
- Watanabe, S.; Hajima, T.; Sudo, K.; Nagashima, T.; Takemura, T.; Okajima, H.; Nozawa, T.; Kawase, H.; Abe, M.; Yokohata, T. MIROC-ESM 2010: Model description and basic results of CMIP5-20c3m experiments. Geosci. Model Dev. 2011, 4, 845. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef]
- Brown, J.L. SDMtoolbox: A python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. Methods Ecol. Evol. 2014, 5, 694–700. [Google Scholar] [CrossRef]
- Iannella, M.; Cerasoli, F.; Biondi, M. Unraveling climate influences on the distribution of the parapatric newts Lissotriton vulgaris meridionalis and L. italicus. Front. Zool. 2017, 14, 55. [Google Scholar] [CrossRef]
- Cianfrani, C.; Le Lay, G.; Maiorano, L.; Satizábal, H.F.; Loy, A.; Guisan, A. Adapting global conservation strategies to climate change at the European scale: The otter as a flagship species. Biol. Conserv. 2011, 144, 2068–2080. [Google Scholar] [CrossRef]
- Van Gils, H.; Westinga, E.; Carafa, M.; Antonucci, A.; Ciaschetti, G. Where the bears roam in Majella National Park, Italy. J. Nat. Conserv. 2014, 22, 23–34. [Google Scholar] [CrossRef]
- Li, R.; Xu, M.; Wong, M.H.G.; Qiu, S.; Li, X.; Ehrenfeld, D.; Li, D. Climate change threatens giant panda protection in the 21st century. Biol. Conserv. 2015, 182, 93–101. [Google Scholar] [CrossRef]
- Dey, N.; Bhatt, C.; Ashour, A.S. Big Data for Remote Sensing: Visualization, Analysis and Interpretation; Springer: Cham, Switzerland, 2018. [Google Scholar]
- Lavender, S.; Lavender, A. Practical Handbook of Remote Sensing; Informa: London, UK, 2015. [Google Scholar]
- Thenkabail, P.S. Remotely Sensed Data Characterization, Classification, and Accuracies; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Kumar, L.; Mutanga, O. Google Earth Engine Applications since Inception: Usage, Trends, and Potential. Remote Sens. 2018, 10, 1509. [Google Scholar] [CrossRef]
- Landolt, E.; Bäumler, B.; Erhardt, A.; Hegg, O.; Klötzli, F.; Lämmler, W.; Nobis, M.; Rudmann-Maurer, K.; Schweingruber, F.; Theurillat, J.P. Flora Indicativa Ecological Inicator Values and Biological Attributes of the Flora of Switzerland and the Alps: Ökologische Zeigerwerte und Biologische Kennzeichen zur Flora der Schweiz und der Alpen; Haupt Verlag: Bern, Switzerland, 2010. [Google Scholar]
- Thenkabail, P.S.; Knox, J.W.; Ozdogan, M.; Gumma, M.K.; Congalton, R.G.; Wu, Z.; Milesi, C.; Finkral, A.; Marshall, M.; Mariotto, I. Assessing future risks to agricultural productivity, water resources and food security: How can remote sensing help? Photogramm. Eng. Remote Sens. 2012, 78, 773–782. [Google Scholar]
- Csontos, P.; Vitalos, M.; Barina, Z.; Kiss, L. Early distribution and spread of Ambrosia artemisiifolia in Central and Eastern Europe. Bot. Helv. 2010, 120, 75–78. [Google Scholar] [CrossRef]
- Vitalos, M.; Karrer, G. Dispersal of Ambrosia artemisiifolia seeds along roads: The contribution of traffic and mowing machines. Neobiota 2009, 8, 53–60. [Google Scholar]
- Bašić, F.; Ðikić, M.; Gadžo, D.; Gavrić, T. Investigation of allelopathic influence of invasive weed species common ragweed (Ambrosia artemisiifolia L.) on the initial growth parameters of selected plants. Rad. Poljopr. Fak. Univ. U Sarajev. Fac. Agric. Univ. Sarajevo 2018, 63, 25–34. [Google Scholar]
- Schindler, S.; Rabitsch, W.; Essl, F.; Wallner, P.; Lemmerer, K.; Follak, S.; Hutter, H.-P. Alien Species and Human Health: Austrian Stakeholder Perspective on Challenges and Solutions. Int. J. Environ. Res. Public Health 2018, 15, 2527. [Google Scholar] [CrossRef]
- Urbanowicz, C.; Hutyra, L.R.; Stinson, K.A. The effects of urbanization and land use on ragweed distribution. Ecosphere 2018, 9, e02512. [Google Scholar] [CrossRef]
- Tanaka, K.; Murata, K.; Matsuura, A. Rapid evolution of an introduced insect Ophraella communa Le Sage in new environments: Temporal changes and geographical differences in photoperiodic response. Entomol. Sci. 2015, 18, 104–112. [Google Scholar] [CrossRef]
- Zhou, Z.S.; Rasmann, S.; Li, M.; Guo, J.Y.; Chen, H.S.; Wan, F.H. Cold Temperatures Increase Cold Hardiness in the Next Generation Ophraella communa Beetles. PLoS ONE 2013, 8, e74760. [Google Scholar] [CrossRef]







© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Iannella, M.; De Simone, W.; D’Alessandro, P.; Console, G.; Biondi, M. Investigating the Current and Future Co-Occurrence of Ambrosia artemisiifolia and Ophraella communa in Europe through Ecological Modelling and Remote Sensing Data Analysis. Int. J. Environ. Res. Public Health 2019, 16, 3416. https://doi.org/10.3390/ijerph16183416
Iannella M, De Simone W, D’Alessandro P, Console G, Biondi M. Investigating the Current and Future Co-Occurrence of Ambrosia artemisiifolia and Ophraella communa in Europe through Ecological Modelling and Remote Sensing Data Analysis. International Journal of Environmental Research and Public Health. 2019; 16(18):3416. https://doi.org/10.3390/ijerph16183416
Chicago/Turabian StyleIannella, Mattia, Walter De Simone, Paola D’Alessandro, Giulia Console, and Maurizio Biondi. 2019. "Investigating the Current and Future Co-Occurrence of Ambrosia artemisiifolia and Ophraella communa in Europe through Ecological Modelling and Remote Sensing Data Analysis" International Journal of Environmental Research and Public Health 16, no. 18: 3416. https://doi.org/10.3390/ijerph16183416