Assessing Risk of E. coli Resuspension from Intertidal Estuarine Sediments: Implications for Water Quality

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling Sites
2.2. E. coli Enumeration
2.3. Erosion Potential of Sediments
3. Results
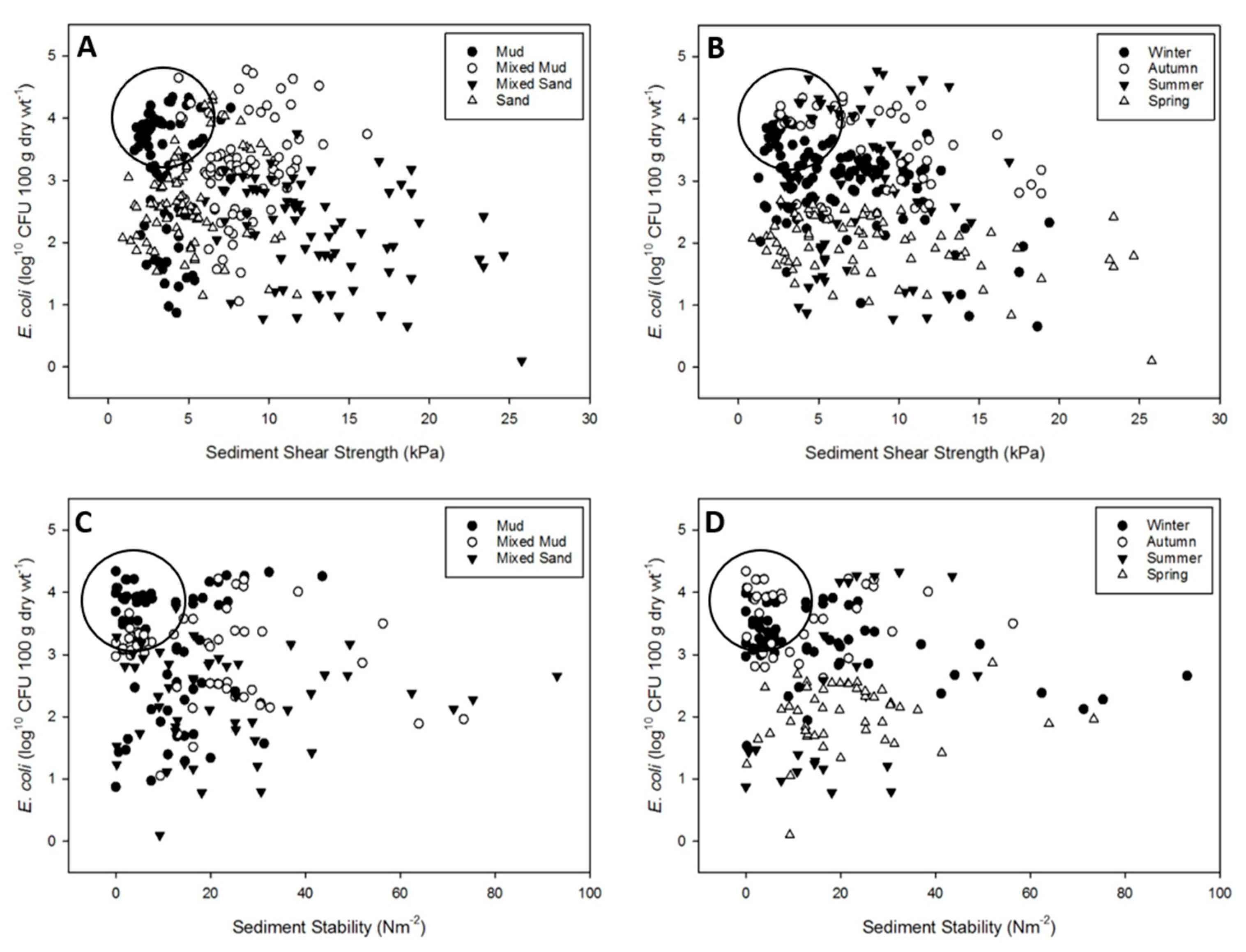
3.1. Spatio-Temporal Variability in E. coli Resuspension Potential in the Ythan Estuary
3.2. Resuspension Potential of E. coli: Seasonal Transects at Two Estuaries
3.3. Modelling of Erosion Thresholds
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Jeng, H.C.; England, A.J.; Bradford, H.B. Indicator organisms associated with stormwater suspended particles and estuarine sediment. J. Environ. Sci. Health A 2005, 40, 779–791. [Google Scholar] [CrossRef]
- Pachepsky, Y.A.; Shelton, D.R. Escherichia coli and fecal coliforms in freshwater and estuarine sediments. Crit. Rev. Environ. Sci. Techno.l 2011, 41, 1067–1110. [Google Scholar] [CrossRef]
- England, L.S.; Lee, H.; Trevors, J.T. Bacterial survival in soil: Effect of clays and protozoa. Soil Biol. Biochem. 1993, 25, 525–531. [Google Scholar] [CrossRef]
- Hassard, F.; Andrews, A.; Jones, D.L.; Parsons, L.; Jones, V.; Cox, B.A.; Daldorph, P.; Brett, H.; McDonald, J.E.; Malham, S.K. Physicochemical factors influence the abundance and culturability of human enteric pathogens and fecal indicator organisms in estuarine water and sediment. Front. Microbiol. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Davies, C.M.; Long, J.A.H.; Donald, M.; Ashbolt, N.J. Survival of fecal microorganisms in marine and freshwater sediments. Appl. Environ. Microbiol. 1995, 61, 1888–1896. [Google Scholar] [PubMed]
- Craig, D.L.; Fallowfield, H.J.; Cromar, N.J. Use of microcosms to determine persistence of Escherichia coli in recreational coastal water and sediment and validation with in situ measurements. J. Appl. Microbiol. 2004, 96, 922–930. [Google Scholar] [CrossRef] [PubMed]
- Mehta, A.J.; Partheniades, E. Resuspension potential of deposited cohesive sediment beds. In Estuarine Comparisons; Kennedy, V.S., Ed.; Academic Press: New York, NY, USA, 1982. [Google Scholar]
- Defew, E.C.; Tolhurst, T.J.; Paterson, D.M. Site-specific features influence sediment stability of intertidal flats. Hydrol. Earth Syst. Sci. 2002, 6, 971–982. [Google Scholar] [CrossRef]
- de Deckere, E.M.G.T.; van de Koppel, J.; Heip, C.H.R. The influence of Corophium volutator abundance on resuspension. In Life at Interfaces and Under Extreme Conditions; Liebezeit, G., Dittmann, S., Kröncke, I., Eds.; Springer Netherlands: Dordrecht, The Netherlands, 2002; pp. 37–42. [Google Scholar]
- Perkins, T.L.; Clements, K.; Baas, J.H.; Jago, C.F.; Jones, D.L.; Malham, S.K.; McDonald, J.E. Sediment composition influences spatial variation in the abundance of human pathogen indicator bacteria within an estuarine environment. PLoS ONE 2014, 9, e112951. [Google Scholar] [CrossRef]
- Wyness, A.J.; Paterson, D.M.; Mendo, T.; Defew, E.C.; Stutter, M.I.; Avery, L.M. Factors affecting the spatial and temporal distribution of E. coli in intertidal estuarine sediments. Sci. Total Envir. 2019, 661, 155–167. [Google Scholar] [CrossRef] [PubMed]
- Schang, C.; Lintern, A.; Cook, P.L.M.; Rooney, G.; Coleman, R.; Murphy, H.M.; Deletic, A.; McCarthy, D. Escherichia coli survival and transfer in estuarine bed sediments. River Res. Appl. 2018, 34, 606–614. [Google Scholar] [CrossRef]
- Le Hir, P.; Monbet, Y.; Orvain, F. Sediment erodability in sediment transport modelling: Can we account for biota effects? Cont. Shelf. Res. 2007, 27, 1116–1142. [Google Scholar] [CrossRef]
- Underwood, G.J.; Paterson, D.M. Seasonal changes in diatom biomass, sediment stability and biogenic stabilization in the Severn estuary. J. Mar. Biol. Assn. UK 1993, 73, 871–887. [Google Scholar] [CrossRef]
- Grabowski, R.C.; Droppo, I.G.; Wharton, G. Erodibility of cohesive sediment: The importance of sediment properties. Earth-Sci. Rev. 2011, 105, 101–120. [Google Scholar] [CrossRef]
- Gerbersdorf, S.U.; Jancke, T.; Westrich, B. Physico-chemical and biological sediment properties determining erosion resistance of contaminated riverine sediments—Temporal and vertical pattern at the lauffen reservoir/river neckar, Germany. Limnologica 2005, 35, 132–144. [Google Scholar] [CrossRef]
- Black, K.S.; Tolhurst, T.J.; Paterson, D.M.; Hagerthey, S.E. Working with natural cohesive sediments. J. Hydraul. Eng-Asce. 2002, 128, 2–8. [Google Scholar] [CrossRef]
- Grabowski, R.C. Measuring the Shear Strength of Cohesive Sediment in the Field. Available online: https://pdfs.semanticscholar.org/0192/ec0a4822ada087fc428fc04f9376cfcf97b9.pdf (accessed on 4 September 2019).
- Jepsen, R.; Roberts, J.; Lick, W. Effects of bulk density on sediment erosion rates. Water Air Soil Pollut. 1997, 99, 21–31. [Google Scholar] [CrossRef]
- Paterson, D.M. Short-term changes in the erodibility of intertidal cohesive sediments related to the migratory behavior of epipelic diatoms. Limnol. Oceanogr. 1989, 34, 223–234. [Google Scholar] [CrossRef]
- Passarelli, C.; Hubas, C.; Paterson, D.M. Mudflat Ecosystem Engineers and Services. In Mudflat Ecology; Beninger, P.G., Ed.; Springer: New York, NY, USA, 2018; pp. 243–269. [Google Scholar]
- de Deckere, E.M.G.T.; Tolhurst, T.J.; de Brouwer, J.F.C. Destabilization of cohesive intertidal sediments by infauna. Estuar. Coast. Shelf Sci. 2001, 53, 665–669. [Google Scholar] [CrossRef]
- Whitehouse, R. Dynamics of Estuarine Muds: A manual for Practical Applications; Thomas Telford: London, UK, 2000. [Google Scholar]
- Dade, W.B.; Self, R.L.; Pellerin, N.B.; Moffet, A.; Jumars, P.A.; Nowell, A.R.M. The effects of bacteria on the flow behaviour of clay-seawater suspensions. J. Sediment. Res. 1996, 66, 39–42. [Google Scholar]
- Dade, W.B.; Davis, J.D.; Nichols, P.D.; Nowell, A.R.M.; Thistle, D.; Trexler, M.B.; White, D.C. Effects of bacterial exopolymer adhesion on the entrainment of sand. Geomicrobiol. J. 1990, 8, 1–16. [Google Scholar] [CrossRef]
- Gerbersdorf, S.U.; Bittner, R.; Lubarsky, H.; Manz, W.; Paterson, D.M. Microbial assemblages as ecosystem engineers of sediment stability. J. Soils Sediments 2009, 9, 640–652. [Google Scholar] [CrossRef]
- De Brouwer, J.F.; Wolfstein, K.; Ruddy, G.K.; Jones, T.E.; Stal, L.J. Biogenic stabilization of intertidal sediments: The importance of extracellular polymeric substances produced by benthic diatoms. Microbial Ecol. 2005, 49, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Austen, I.; Andersen, T.J.; Edelvang, K. The influence of benthic diatoms and invertebrates on the erodibility of an intertidal a mudflat, the danish wadden sea. Estuar. Coast. Shelf Sci. 1999, 49, 99–111. [Google Scholar] [CrossRef]
- Underwood, G.J.; Paterson, D.M. The importance of extracellular carbohydrate production by marine epipelic diatoms. Advan. Botan. Res. 2003, 40, 183–240. [Google Scholar]
- Lubarsky, H.V. The Impact of Microbial Extracellular Polymeric Substances on Sediment Stability. Available online: http://agris.fao.org/agris-search/search.do?recordID=AV2012073631 (accessed on 4 September 2019).
- Perkins, R.G.; Underwood, G.J.; Brotas, V.; Snow, G.C.; Jesus, B.; Ribiero, L. Responses of microphytobenthos to light: Primary production and carbohydrate allocation over an emersion period. Mar. Ecol-Progr. Ser. 2001, 223, 101–112. [Google Scholar] [CrossRef]
- Bhaskar, P.V.; Bhosle, N.B. Microbial extracellular polymeric substances in marine biogeochemical processes. Curr. Sci. 2005, 88, 45–53. [Google Scholar]
- Lubarsky, H.V.; Hubas, C.; Chocholek, M.; Larson, F.; Manz, W.; Paterson, D.M.; Gerbersdorf, S.U. The stabilisation potential of individual and mixed assemblages of natural bacteria and microalgae. PLoS ONE 2010, 5, e13794. [Google Scholar] [CrossRef]
- Tolhurst, T.J.; Black, K.S.; Shayler, S.A.; Mather, S.; Black, I.; Baker, K.; Paterson, D.M. Measuring the in situ erosion shear stress of intertidal sediments with the cohesive strength meter (csm). Estuar. Coast. Shelf Sci. 1999, 49, 281–294. [Google Scholar] [CrossRef]
- Cavanaugh, J.E. Unifying the derivations for the akaike and corrected akaike information criteria. Stat. Probab. Lett. 1997, 33, 201–208. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013. [Google Scholar]
- Amos, C.L.; Van Wagoner, N.A.; Daborn, G.R. The influence of subaerial exposure on the bulk properties of fine-grained intertidal sediment from Minas Basin, Bay of Fundy. Estuar. Coast. Shelf Sci. 1988, 27, 1–13. [Google Scholar] [CrossRef]
- McPhail, C.D.; Stidson, R.T. Bathing water signage and predictive water quality models in Scotland. Aquat. Ecosyst. Health 2009, 12, 183–186. [Google Scholar] [CrossRef]
- Stidson, R.T.; Gray, C.A.; McPhail, C.D. Development and use of modelling techniques for real-time bathing water quality predictions. Water Environ. J. 2012, 26, 7–18. [Google Scholar] [CrossRef]
- SEPA. Scottish Bathing Water: 2014–2015. Available online: http://www.sepa.org.uk/media/143136/sepa_bathing_waters_2014-15_web.pdf (accessed on 4 September 2019).
- Martinez-Manzanares, E.; Moriñigo, M.A.; Castro, D.; Balebona, M.C.; Sanchez, J.M.; Borrego, J.J. Influence of the faecal pollution of marine sediments on the microbial content of shellfish. Mar. Pollut. Bull. 1992, 24, 342–349. [Google Scholar] [CrossRef]
- Clements, K.; Quilliam, R.S.; Jones, D.L.; Wilson, J.; Malham, S.K. Spatial and temporal heterogeneity of bacteria across an intertidal shellfish bed: Implications for regulatory monitoring of faecal indicator organisms. Sci. Total Envir. 2015, 506–507, 1–9. [Google Scholar] [CrossRef]
- Cho, K.H.; Pachepsky, Y.A.; Kim, J.H.; Kim, J.W.; Park, M.H. The modified SWAT model for predicting fecal coliforms in the Wachusett reservoir watershed, USA. Water Res. 2012, 46, 4750–4760. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.H.; Pachepsky, Y.A.; Oliver, D.M.; Muirhead, R.W.; Park, Y.; Quilliam, R.S.; Shelton, D.R. Modeling fate and transport of fecally-derived microorganisms at the watershed scale: State of the science and future opportunities. Water Res. 2016, 100, 38–56. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Bedford, K.; Yen, C. Storm and entrainment effects on tributary sediment loads. J. Hydraul. Eng. 1994, 120, 81–103. [Google Scholar] [CrossRef]
- Jamieson, R.C.; Joy, D.M.; Kostaschuk, R.; Gordon, R.J. Resuspension of sediment-associated Escherichia coli in a natural stream. J.Environ. Qual. 2005, 34, 581–589. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Lin, B.; Falconer, R.A. Modelling enteric bacteria level in coastal and estuarine waters. Eng. Comput. Mech. 2008, 161, 179–186. [Google Scholar] [CrossRef]
- Fernandes, S.M.R.; Sobral, P.J.; Costa, M.H.A. Nereis diversicolor effect on the stability of cohesive intertidal sediments. Aquat. Ecol. 2006, 40, 567–579. [Google Scholar] [CrossRef]
- Paterson, D.M.; Hope, J.A.; Kenworthy, J.; Biles, C.L.; Gerbersdorf, S.U. Form, function and physics: The ecology of biogenic stabilisation. J. Soils Sediments 2018, 18, 3044–3054. [Google Scholar] [CrossRef]
- Raffaelli, D. Interactions between macro-algal mats and invertebrates in the Ythan estuary, Aberdeenshire, Scotland. Helgoland Mar. Res. 2000, 54, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Meadows, P.S.; Tait, J. Modification of sediment permeability and shear strength by two burrowing invertebrates. Mar. Biol. 1989, 101, 75–82. [Google Scholar] [CrossRef]
- McLusky, D.S. Some effects of salinity on the distribution and abundance of corophium volutator in the Ythan estuary. J. Mar. Biol. Assn. UK 1968, 48, 443–454. [Google Scholar] [CrossRef]
- Aberle, J.; Nikora, V.; Walters, R. Effects of bed material properties on cohesive sediment erosion. Mar. Geol. 2004, 207, 83–93. [Google Scholar] [CrossRef]
- Tolhurst, T.J.; Friend, P.L.; Watts, C.; Wakefield, R.; Black, K.S.; Paterson, D.M. The effects of rain on the erosion threshold of intertidal cohesive sediments. Aquat. Ecol. 2006, 40, 533–541. [Google Scholar] [CrossRef]
- Balls, P.W.; MacDonald, A.; Pugh, K.B.; Edwards, A.C. Rainfall events and their influence on nutrient distributions in the Ythan estuary (Scotland). Estuar. Coast. Shelf Sci. 1997, 44, 73–81. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predictor | Coefficient | Standardised Coefficient | p-Value |
|---|---|---|---|
| Intercept | 8.625 | 2.978 | <0.001 |
| Organic matter content | −0.549 | 0.230 | <0.050 |
| Season: spring | −0.534 | −0.534 | <0.001 |
| Season: summer | −0.629 | −0.630 | <0.001 |
| Season: winter | −0.398 | −0.398 | <0.001 |
| Median particle diameter | −0.006 | 0.548 | <0.001 |
| pH | −0.740 | −0.196 | <0.001 |
| Bulk density | −0.245 | 0.065 | <0.050 |
| Organic matter content × Median particle diameter | 0.004 | 0.929 | <0.001 |
| Predictor | Coefficient | Standardised Coefficient | p-Value |
|---|---|---|---|
| Intercept | 2.859 | 1.923 | <0.001 |
| Organic matter content | −0.202 | −0.312 | <0.001 |
| Season: spring | 0.653 | 0.653 | <0.001 |
| Season: summer | 0.683 | 0.683 | <0.001 |
| Season: winter | 0.181 | 0.181 | 0.341 |
| Colloidal carbohydrates | 0.002 | 0.253 | <0.001 |
| 5-day cumulative precipitation | −0.098 | 0.124 | 0.117 |
| Tidal amplitude | −0.338 | 0.161 | 0.063 |
| 5-day cumulative precipitation × Tidal amplitude | 0.042 | 0.248 | <0.001 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wyness, A.J.; Paterson, D.M.; Rimmer, J.E.V.; Defew, E.C.; Stutter, M.I.; Avery, L.M. Assessing Risk of E. coli Resuspension from Intertidal Estuarine Sediments: Implications for Water Quality. Int. J. Environ. Res. Public Health 2019, 16, 3255. https://doi.org/10.3390/ijerph16183255
Wyness AJ, Paterson DM, Rimmer JEV, Defew EC, Stutter MI, Avery LM. Assessing Risk of E. coli Resuspension from Intertidal Estuarine Sediments: Implications for Water Quality. International Journal of Environmental Research and Public Health. 2019; 16(18):3255. https://doi.org/10.3390/ijerph16183255
Chicago/Turabian StyleWyness, Adam J., David M. Paterson, James E. V. Rimmer, Emma C. Defew, Marc I. Stutter, and Lisa M. Avery. 2019. "Assessing Risk of E. coli Resuspension from Intertidal Estuarine Sediments: Implications for Water Quality" International Journal of Environmental Research and Public Health 16, no. 18: 3255. https://doi.org/10.3390/ijerph16183255