Climatic Conditions: Conventional and Nanotechnology-Based Methods for the Control of Mosquito Vectors Causing Human Health Issues



Abstract

1. Introduction
2. Methodology
3. Results
3.1. Climatic Conditions Influence on Mosquito Vectors
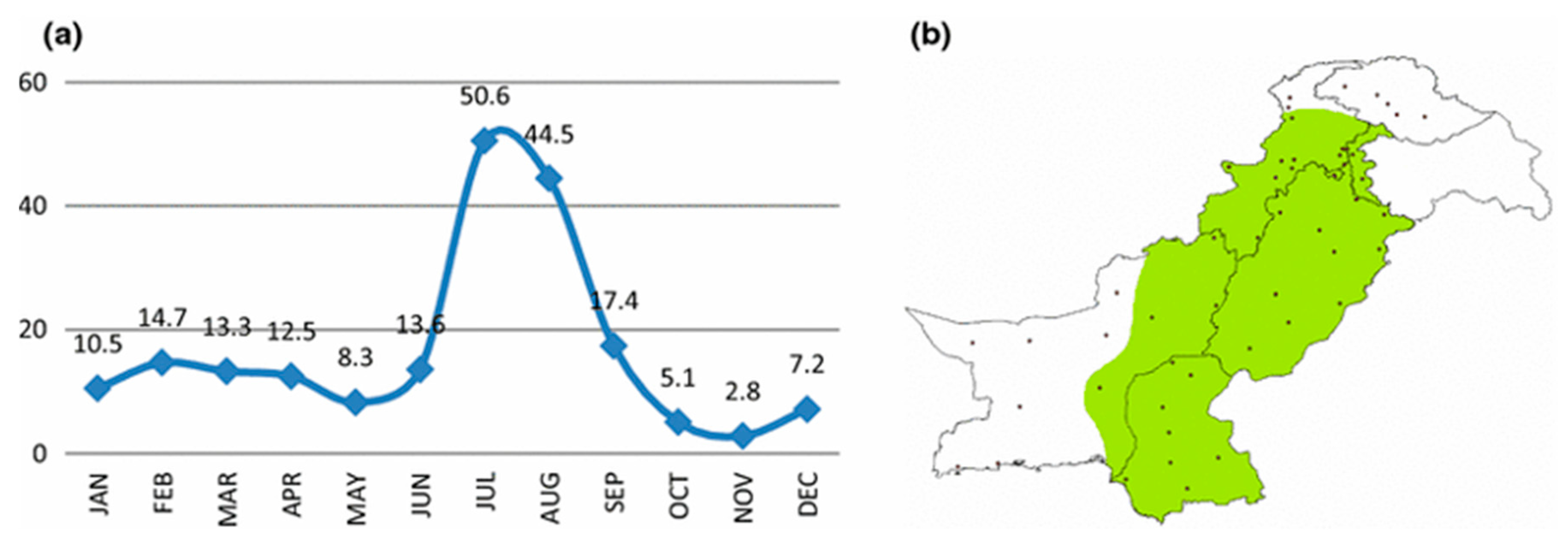
3.1.1. Seasonality of Infectious Diseases
3.1.2. Vector-Borne Diseases
- The reproduction and survival rates of vectors in a certain environment.
- The level and degree of vector-activity; typically, the rate of biting and the period of year.
- Reproduction and development rate of pathogens inside the vector.
3.1.3. Temperature Sensitivity
3.1.4. Precipitation Sensitivity
3.1.5. Humidity Sensitivity
3.1.6. Sea Level Sensitivity
3.2. Mosquito-Borne Diseases and Health Impacts
3.2.1. Vector-Host Relationships
3.2.2. Diseases Transmitted by the Mosquito Vectors
3.2.3. Treatment and Vaccines for Mosquito Vectors
3.3. Conventional Methods Used to Control Mosquito Populations
3.3.1. Overview
3.3.2. Source Reduction (Environmental Control)
3.3.3. Trapping (Mechanical Control)
3.3.4. Insecticides (Chemical Control)
3.3.5. Biological Control
3.3.6. Genetic Control
3.3.7. Integrative Vector Management Approach
4. Nanotechnology-Based Methods
4.1. Overview
4.2. Nanoliposomes
4.3. Nanosuspensions
4.4. Polymer-Based Nanoparticles
4.5. Green-Synthesized Metallic Nanoparticles
5. Drive to Transform Sick to Healthy Life under Changing Climatic Conditions
6. Pakistani Perspective with Reference to Other Countries
7. Conclusions
Outlook
- An environmental management approach is very important for the control of breeding sites of mosquitoes especially during and after monsoons. These sites include open drainage, marshes and standing water in pots.
- Control of breeding sites as a result of increased rainfall and change in rainfall pattern due to climate change, which increases the vector population like mosquitoes and provide more breeding spaces.
- Awareness campaign among the general public at community level and in schools regarding the spread and control of malaria and dengue fever (especially the time period for the diseases).
- Personal protective approaches include use of proper clothing, nets, mosquito repellent lotions and other protective measures in homes, businesses, schools and offices.
- Allocation of enough funds by the authorities for health departments to control the spread of malaria and dengue fever in the community.
- Use of biological, chemical and physical methods to diminish mosquito-related diseases.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Dengue Control. Geneva. Available online: http://www.who.int/denguecontrol/en (accessed on 28 March 2019).
- World Health Organization. Neglected Tropical Diseases. Geneva. Available online: http://www.who.int/neglected_diseases/diseases/en (accessed on 28 March 2019).
- World Health Organization. World Health Day—7 April 2014. Geneva. Available online: http://www.who.int/campaigns/world-health-day/2014/event/en (accessed on 28 March 2014).
- World Health Organization. World Malaria Report 2013. Geneva. Available online: http://www.who.int/malaria/publications/world_malaria_ report_2013/en (accessed on 28 March 2019).
- WHO. Malaria vaccine: WHO position paper-January 2016. Vaccine 2018, 36, 3576–3577. [Google Scholar] [CrossRef] [PubMed]
- Deichstetter, P. The Effect of Climate Change on Mosquito-Borne Diseases. Am. Biol. Teach. 2017, 79, 169–173. [Google Scholar] [CrossRef]
- Githeko, A.K.; Lindsay, S.W.; Confalonieri, U.E.; Patz, J.A. Climate change and vector-borne diseases: A regional analysis. Bull. World Health OrgAnophels 2000, 78, 1136–1147. [Google Scholar]
- Haider, Z.; Ahmad, F.Z.; Mahmood, A.; Waseem, T.; Shafiq, I.; Raza, T.; Qazi, J.; Siddique, N.; Humayun, M.A. Dengue fever in Pakistan: A paradigm shift; changing epidemiology and clinical patterns. Perspect. Public Health 2015, 135, 294–298. [Google Scholar] [CrossRef] [PubMed]
- Yi, H.; Devkota, B.R.; Yu, J.; Oh, K.; Kim, J.; Kim, H. Effects of global warming on mosquitoes & mosquito-borne diseases and the new strategies for mosquito control. Entomol. Res. 2014, 44, 215–235. [Google Scholar] [CrossRef]
- Mohammed, A.; Chadee, D.D. Effects of different temperature regimens on the development of Aedes aegypti (L.) (Diptera: Culicidae) mosquitoes. Acta Trop. 2011, 119, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred Reporting Items for Systematic Reviews and Meta-Analyses: The PRISMA Statement. J. Clin. Epidemiol. 2009, 62, 1006–1012. [Google Scholar] [CrossRef]
- Eisen, L.; Monaghan, A.J.; Lozano-Fuentes, S.; Steinhoff, D.F.; Hayden, M.H.; Bieringer, P.E. The Impact of Temperature on the Bionomics ofAedes (Stegomyia) aegypti, With Special Reference to the Cool Geographic Range Margins. J. Med Èntomol. 2014, 51, 496–516. [Google Scholar] [CrossRef] [PubMed]
- Hamlet, A.; Jean, K.; Perea, W.; Yactayo, S.; Biey, J.; Van Kerkhove, M.; Ferguson, N.; Garske, T. The seasonal influence of climate and environment on yellow fever transmission across Africa. PLoS Negl. Trop. Dis. 2018, 12, e0006284. [Google Scholar] [CrossRef]
- Madico, G.; McDonald, J.; Gilman, R.H.; Cabrera, L.; Sterling, C.R. Epidemiology and Treatment of Cyclospora cayetanensis Infection in Peruvian Children. Clin. Infect. Dis. 1997, 24, 977–981. [Google Scholar] [CrossRef]
- Moore, P.S. Meningococcal Meningitis in Sub-Saharan Africa: A Model for the Epidemic Process. Clin. Infect. Dis. 1992, 14, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Magori, K.; Drake, J.M. The Population Dynamics of Vector-borne Diseases. Nat. Educ. Knowl. 2013, 4, 14. [Google Scholar]
- Kovats, R.S.; Campbell-Lendrum, D.H.; McMichael, A.J.; Woodward, A.; Cox, J.S. Early effects of climate change: Do they include changes in vector-borne disease? Philos. Trans. R. Soc. Lond. Ser. B. Biol. Sci. 2001, 356, 1057–1068. [Google Scholar] [CrossRef] [PubMed]
- Bradley, D.J. Human tropical diseases in a changing environment. Ciba Found. Symp. 1993, 175, 146–162. [Google Scholar] [PubMed]
- Lindsay, S.W.; Birley, M.H. Climate change and malaria transmission. Ann. Trop. Med. Parasitol. 1996, 90, 573–588. [Google Scholar] [CrossRef] [PubMed]
- Beserra, E.B.; Fernandes, C.R.M.; Silva, S.A.D.O.; Da Silva, L.A.; Santos, J.W. Efeitos da temperatura no ciclo de vida, exigências térmicas e estimativas do número de gerações anuais de Aedes aegypti (Diptera, Culicidae). Iheringia. Série Zool. 2009, 99, 142–148. [Google Scholar] [CrossRef]
- Marinho, R.A.; Beserra, E.B.; Bezerra-Gusmão, M.A.; Porto, V.D.; Olinda, R.A.; dos Santos, C.A. Effects of temperature on the life cycle, expansion, and dispersion of Aedes aegypti (Diptera: Culicidae) in three cities in Paraiba, Brazil. J. Vector Ecol. 2016, 41, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gubler, D.J.; Reiter, P.; Ebi, K.L.; Yap, W.; Nasci, R.; Patz, J.A. Climate variability and change in the United States: Potential impacts on vector- and rodent-borne diseases. Environ. Health Perspect. 2001, 109, 223–233. [Google Scholar] [PubMed]
- Cox, J.; Craig, M.; Sueur, D.L.; Sharp, B. Mapping Malaria Risk in the Highlands of Africa. In Mapping Climate Vulnerability and Poverty in Africa: Report to the Department for International Development; ILRI: Nairobi, Kenya, 1999; p. 96. [Google Scholar]
- Ko, A.I.; Galvão Reis, M.; Ribeiro Dourado, C.M.; Johnson, W.D., Jr.; Riley, L.W. Urban epidemic of severe leptospirosis in Brazil. Salvador Leptospirosis Study Group. Lancet 1999, 354, 820–825. [Google Scholar] [CrossRef]
- Bai, L.; Morton, L.C.; Liu, Q. Climate change and mosquito-borne diseases in China: A review. Glob. Health 2013, 9, 10. [Google Scholar] [CrossRef]
- Yang, G.J.; Gao, Q.; Zhou, S.S.; Malone, J.B.; McCarroll, J.C.; Tanner, M.; Vounatsou, P.; Bergquist, R.; Utzinger, J.; Zhou, X.N. Mapping and predicting malaria transmission in the People’s Republic of China, using integrated biology-driven and statistical models. Geospat. Health 2010, 5, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Focks, D.A.; Daniels, E.; Haile, D.G.; Keesling, J.E. A Simulation Model of the Epidemiology of Urban Dengue Fever: Literature Analysis, Model Development, Preliminary Validation, and Samples of Simulation Results. Am. J. Trop. Med. Hyg. 1995, 53, 489–506. [Google Scholar] [CrossRef] [PubMed]
- Reeves, W.C.; Hardy, J.L.; Reisen, W.K.; Milby, M.M. Potential Effect of Global Warming on Mosquito-Borne Arboviruses. J. Med Èntomol. 1994, 31, 323–332. [Google Scholar] [CrossRef] [PubMed]
- Reiter, P. Weather, Vector Biology, and Arboviral Recrudescence. In The Arboviruses: Epidemiology and Ecology; Monath, T.P., Ed.; CRC Press: Boca Raton, FL, USA, 1988; pp. 245–255. [Google Scholar]
- Reiter, P. Global-warming and vector-borne disease in temperate regions and at high altitude. Lancet 1998, 351, 839–840. [Google Scholar] [CrossRef]
- Reisen, W.K. Effect of Temperature on Culex tarsalis (Diptera: Culicidae) from the Coachella and San Joaquin Valleys of California. J. Med Èntomol. 1995, 32, 636–645. [Google Scholar] [CrossRef] [PubMed]
- Kramer, L.D.; Hardy, J.L.; Presser, S.B. Effect of Temperature of Extrinsic Incubation on the Vector Competence of Culex tarsalis for Western Equine Encephalomyelitis Virus. Am. J. Trop. Med. Hyg. 1983, 32, 1130–1139. [Google Scholar] [CrossRef] [PubMed]
- Day, J.F.; Curtis, G.A. Influence of Rainfall on Culex nigripalpus (Diptera: Culicidae) Blood-Feeding Behavior in Indian River County, Florida. Ann. Èntomol. Soc. Am. 1989, 82, 32–37. [Google Scholar] [CrossRef]
- Hardy, J.L.; Meyer, R.P.; Presser, S.B.; Milby, M.M. Temporal Variations in the Susceptibility of a Semi-Isolated Population of Culex tarsalis to Peroral Infection with Western Equine Encephalomyelitis and St. Louis Encephalitis Viruses. Am. J. Trop. Med. Hyg. 1990, 42, 500–511. [Google Scholar] [CrossRef]
- Lansdowne, C.; Hacker, C.S. The Effect of Fluctuating Temperature and Humidity on the Adult Life Table Characteristics of Five Strains of Aedes Aegypti. J. Med Èntomol. 1975, 11, 723–733. [Google Scholar] [CrossRef]
- United Nations (UN); Intergovernmental Panel on Climate Change (IPCC). Fourth Assessment Report Climate Change 2007; Intergovernmental Panel on Climate Change: Geneva, Switzerland, 2007. [Google Scholar]
- Gratz, N.G. Emerging and resurging vector-borne diseases. Annu. Rev. Èntomol. 1999, 44, 51–75. [Google Scholar] [CrossRef]
- Halstead, S.B. Dengue. Lancet 2007, 370, 1644–1652. [Google Scholar] [CrossRef]
- Scholte, E.J.; Schaffner, F. Waiting for the Tiger: Establishment and Spread of the Ae. albopictus Mosquito in Europe. In Emerging Pests and Vector-borne Disease in Europe; Takken, W., Knols, B.G.J., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2007; pp. 241–460. [Google Scholar]
- Guerra, C.A.; Gikandi, P.W.; Tatem, A.J.; Noor, A.M.; Smith, D.L.; I Hay, S.; Snow, R.W. The Limits and Intensity of Plasmodium falciparum Transmission: Implications for Malaria Control and Elimination Worldwide. PLoS Med. 2008, 5, e38. [Google Scholar] [CrossRef] [PubMed]
- WHO. Dengue and Severe Dengue. 2019 Report. Available online: https://www.who.int/news-room/fact-sheets/detail/dengue-and-severe-dengue (accessed on 2 July 2019).
- Kantor, I.N. Dengue, Zika, Chikungunya y el Desarrollo de Vacunas; MEDICINA: Buenos Aires, Argentina, 2018; Volume 78, pp. 23–28. ISBN 1669-9106. [Google Scholar]
- Paixão, E.S.; Teixeira, M.G.; Rodrigues, L.C. Zika, chikungunya and dengue: The causes and threats of new and re-emerging arboviral diseases. BMJ Glob. Health 2018, 3, 000530. [Google Scholar] [CrossRef] [PubMed]
- NIH, NIH Begins Study of Vaccine to Protect Against Mosquito-Borne Diseases. 2017. Available online: https://www.nih.gov/news-events/news-releases/nih-begins-study-vaccine-protect-against-mosquito-borne-diseases (accessed on 18 August 2019).
- WHO. Yellow Fever. Available online: https://www.who.int/news-room/fact-sheets/detail/yellow-fever (accessed on 18 August 2019).
- Chebabi Abramides, G.; Roiz, D.; Guitart, R.; Quintana, S.; Guerrero, I.; Gimenez, N. Effectiveness of a multiple intervention strategy for the control of the tiger mosquito (Ae. albopictus) in Spain. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, D.M.; Unlu, I.; Crepeau, T.; Farajollahi, A.; Healy, S.P.; Bartlett-Healy, K.; Strickman, D.; Gaugler, R.; Hamilton, G.; Kline, D.; et al. Area-wide management of Ae. albopictus. Part 2, Gauging the efficacy of traditional integrated pest control measures against urban container mosquitoes. Pest Manag Sci. 2013, 69, 1351–1361. [Google Scholar] [CrossRef] [PubMed]
- Dowling, Z.; Armbruster, P.; LaDeau, S.L.; Decotiis, M.; Mottley, J.; Leisnham, P.T. Linking Mosquito Infestation to Resident Socioeconomic Status, Knowledge, and Source Reduction Practices in Suburban Washington, DC. EcoHealth 2013, 10, 36–47. [Google Scholar] [CrossRef]
- Petrić, D.; Bellini, R.; Scholte, E.-J.; Rakotoarivony, L.M.; Schaffner, F. Monitoring population and environmental parameters of invasive mosquito species in Europe. Parasites Vectors 2014, 7, 187. [Google Scholar] [CrossRef]
- Jacups, S.P.; Rapley, L.P.; Johnson, P.H.; Benjamin, S.; Ritchie, S.A. Bacillus thuringiensis var. israelensis misting for control of Aedes in cryptic ground containers in North Queensland, Australia. Am. J. Trop. Med. Hyg. 2013, 88, 490–496. [Google Scholar] [CrossRef]
- Kline, D.L. Semiochemicals, traps/targets and mass trapping technology for mosquito management. J. Am. Mosq. Control. Assoc. 2007, 23, 241–251. [Google Scholar] [CrossRef]
- Mackay, A.J.; Amador, M.; Barrera, R. An improved autocidal gravid ovitrap for the control and surveillance of Aedes aegypti. Parasites Vectors 2013, 6, 225. [Google Scholar] [CrossRef]
- Cook, S.M.; Khan, Z.R.; Pickett, J.A. The Use of Push-Pull Strategies in Integrated Pest Management. Annu. Rev. Èntomol. 2007, 52, 375–400. [Google Scholar] [CrossRef] [PubMed]
- Paz-Soldan, V.A.; Plasai, V.; Morrison, A.C.; Rios-Lopez, E.J.; Guedez-Gonzales, S.; Grieco, J.P.; Mundal, K.; Chareonviriyaphap, T.; Achee, N.L. Initial assessment of the acceptability of a push-pull Ae. aegypti control strategy in Iquitos, Peru and Kanchanaburi, Thailand. Am. J. Trop. Med. Hyg. 2011, 84, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Lima, J.B.P.; Rosa-Freitas, M.G.; Rodovalho, C.M.; Santos, F.; Lourenço-De-Oliveira, R. Is there an efficient trap or collection method for sampling Anopheles darlingi and other malaria vectors that can describe the essential parameters affecting transmission dynamics as effectively as human landing catches? - A Review. Memórias Do Inst. Oswaldo Cruz 2014, 109, 685–705. [Google Scholar] [CrossRef] [PubMed]
- Salazar, F.V.; Achee, N.L.; Grieco, J.P.; Prabaripai, A.; Ojo, T.A.; Eisen, L.; Dureza, C.; Polsomboon, S.; Chareonviriyaphap, T. Effect of Ae. aegypti exposure to spatial repellent chemicals on BG-Sentinel (TM) trap catches. Parasit. Vect. 2013, 6, 145. [Google Scholar] [CrossRef] [PubMed]
- Suman, D.S.; Wang, Y.; Bilgrami, A.L.; Gaugler, R. Ovicidal activity of three insect growth regulators against Aedes and Culex mosquitoes. Acta Trop. 2013, 128, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Mulla, M.S. The future of insect growth regulators in vector control. J. Am. Mosq. Control. Assoc. 1995, 11, 269–273. [Google Scholar] [PubMed]
- Ocampo, C.B.; Julieth, M.N.; Carabali, M.; Alexander, N.; Osorio, L. Reduction in dengue cases observed during mass control of Aedes (Stegomyia) in street catch basins in an endemic urban area in Colombia. Acta Trop. 2014, 132, 15–22. [Google Scholar] [CrossRef]
- PubChem. U.S National library of medicine (NIH), National Center for Biotechnology Information. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Fendona#section=Depositor-Supplied-Synonyms (accessed on 29 August 2019).
- PubChem. U.S National library of medicine (NIH), National Center for Biotechnology Information. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/lambda-Cyhalothrin (accessed on 29 August 2019).
- PubChem. U.S National library of medicine (NIH), National Center for Biotechnology Information. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/deltamethrin (accessed on 29 August 2019).
- PubChem. U.S National library of medicine (NIH), National Center for Biotechnology Information. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Fenpropathrin (accessed on 29 August 2019).
- Regnault-Roger, C. The potential of botanical essential oils for insect pest control. Integr. Pest Manag. Rev. 1997, 2, 25–34. [Google Scholar] [CrossRef]
- Roman, P. Essential oils for the development of eco-friendly mosquito larvicides: A review. Ind. Crop. Prod. 2015, 76, 174–187. [Google Scholar]
- Dias, C.N.; Moraes, D.F. Essential oils and their compounds as Ae. aegypti L. (Diptera: Culicidae) larvicides: Review. Parasitol. Res. 2014, 113, 565–592. [Google Scholar] [CrossRef]
- Maheswaran, R.; Ignacimuthu, S. A novel herbal formulation against dengue vector mosquitoes Ae. aegypti and Ae. albopictus. Parasitol. Res. 2012, 110, 1801–1813. [Google Scholar] [CrossRef] [PubMed]
- Mikami, A.; Yamashita, N. The inhibitory effects of a neem formulation on emergence of Oc. japonicus and Culex pipiens pallens. Med. Entomol. Zool. 2004, 55, 239–242. [Google Scholar] [CrossRef]
- Felipe, C.F.B.; Fonsêca, K.S.; Barbosa, A.L.R.; Bezerra, J.N.S.; Neto, M.A.; França Fonteles, M.M.; Barros Viana, G.S. Alterations in behavior and memory induced by the essential oil of Zingiber officinale Roscoe (ginger) in mice are cholinergic-dependent. J. Med. Plants Res. 2008, 2, 163–170. [Google Scholar]
- Rameshwar, S.R. Mechanism of action of insecticidal secondary metabolites of plant origin. Crop Prot. 2010, 29, 913–920. [Google Scholar]
- Federici, B.A.; Park, H.W.; Bideshi, D.K.; Wirth, M.C.; Johnson, J.J.; Sakano, Y.; Tang, M. Developing recombinant bacteria for control of mosquito larvae. J. Am. Mosq. Control. Assoc. 2007, 23, 164–175. [Google Scholar] [CrossRef]
- Bloomquist, J.R.; Boina, D.R.; Chow, E.; Carlier, P.R.; Reina, M.; González-Coloma, A. Mode of action of the plant-derived silphinenes on insect and mammalian GABAA receptor/chloride channel complex. Pestic. Biochem. Physiol. 2008, 91, 17–23. [Google Scholar] [CrossRef]
- Casida, J.E. Pyrethrum, the Natural Insecticide; Academic Press: New York, NY, USA, 1973. [Google Scholar]
- Khambay, B.P.; Batty, D.; Jewess, P.J.; Bateman, G.L.; Hollomon, D.W. Mode of action and pesticidal activity of the natural product dunnione and of some analogues. Pest Manag. Sci. 2003, 59, 174–182. [Google Scholar] [CrossRef]
- Copping, L.G.; Menn, J.J. Biopesticides: A review of their action, applications and efficacy. Pest Manag. Sci. 2000, 56, 651–676. [Google Scholar] [CrossRef]
- Bloomquist, J.R. Ion Channels as Targets for Insecticides. Annu. Rev. Èntomol. 1996, 41, 163–190. [Google Scholar] [CrossRef]
- Enan, E.E. Molecular and pharmacological analysis of an octopamine receptor from american cockroach and fruit fly in response to plant essential oils. Arch. Insect Biochem. Physiol. 2005, 59, 161–171. [Google Scholar] [CrossRef]
- Kostyukovsky, M.; Rafaeli, A.; Gileadi, C.; Demchenko, N.; Shaaya, E. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manag. Sci. 2002, 58, 1101–1106. [Google Scholar] [CrossRef] [PubMed]
- Enan, E.E. Molecular response of Drosophila melanogaster tyramine receptor cascade to plant essential oils. Insect Biochem. Mol. Biol. 2005, 35, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Subbiah, P. Current trends in the control of mosquito vectors by means of biological larvicides. J. Biofertil. Biopestici. 2012, 3. [Google Scholar] [CrossRef]
- Mahmood, F. Laboratory bioassay to compare susceptibilities of Ae. aegypti and Anophels albimanus to Bacillus thuringiensis var. israelensis as affected by their feeding rates. J. Am. Mosq. Control Assoc. 1998, 14, 69–71. [Google Scholar] [PubMed]
- Singh, G.J.; Gill, S.S. An electron microscope study of the toxic action of Bacillus sphaericus in Culex quinquefasciatus larvae. J. Invertebr. Pathol. 1998, 52, 237–247. [Google Scholar] [CrossRef]
- Poopathi, S.; Mani, T.R.; Rao, D.R.; Baskaran, G.; Kabilan, L. Susceptibilty levels of resistance of Culex quinquefasciatus to the insecticidal toxin of Bacillus sphaericus (strain 2362). Environ. Ecol. 2000, 18, 703–710. [Google Scholar]
- Poopathi, S.; Mani, T.R.; Raghunatha, R.D.; Baskaran, G.; Lalitha, K. Effect of Bacillus sphaericus and Bacillus thuringiensis yare israelensis on the ultrastructural changes in the midgut of Culex quinquefasciatus Say (Diptera: Culicidae). J. Ent. Res. 1999, 23, 347–357. [Google Scholar]
- Charles, J.F.; Silva-Filha, M.H.; Nielsen-LeRoux, C.; Humphreys, M.J.; Berry, C. Binding of the 51- and 42-kDa individual components from the Bacillus sphaericus crystal toxin to mosquito larval midgut membranes from Culex and Anophels sp. (Diptera: Culicidae). FEMS Microbiol. Lett. 1997, 156, 153–159. [Google Scholar] [CrossRef]
- Silva-Filha, M.H.; Nielsen-LeRoux, C.; Charles, J.F. Identification of the receptor for Bacillus sphaericus crystal toxin in the brush border membrane of the mosquito Culex pipiens (Diptera: Culicidae). Insect Biochem. Mol. Biol. 1999, 29, 711–721. [Google Scholar] [CrossRef]
- Baumann, P.; A Clark, M.; Baumann, L.; Broadwell, A.H. Bacillus sphaericus as a mosquito pathogen: Properties of the organism and its toxins. Microbiol. Rev. 1991, 55, 425–436. [Google Scholar]
- Ignoffo, C.M.; Couch, T.L.; Garcia, C.; Kroha, M.J. Relative activity of Bacillus thuringiensis var. kurstaki and B. thuringiensis var. israelensis against larvae of Ae. aegypti, Culex quinquefasciatus, Trichoplusia ni, Heliothis zea, and Heliothis virescens. J. Econ. Entomol. 1981, 74, 218–222. [Google Scholar] [CrossRef] [PubMed]
- Höfte, H.; Whiteley, H.R. Insecticidal crystal proteins of Bacillus thuringiensis. Microbiol. Rev. 1989, 53, 242–255. [Google Scholar] [PubMed]
- Delecluse, A.; Rosso, M.L.; Ragni, A. Cloning and expression of a novel toxin gene from Bacillus thuringiensis sub sp. jegathesan encoding a highly mosquitocidal protein. Appl. Environ. Microbiol. 1995, 61, 4230–4235. [Google Scholar] [PubMed]
- Frutos, R.; Rang, C.; Royer, M. Managing Insect Resistance to Plants ProducingBacillus thuringiensisToxins. Crit. Rev. Biotechnol. 1999, 19, 227–276. [Google Scholar] [CrossRef]
- Pimentel, D.; Burgess, M. Effects of Single Versus Combinations of Insecticides on the Development of Resistance. Environ. Èntomol. 1985, 14, 582–589. [Google Scholar] [CrossRef]
- McGaughey, W.H.; Gould, F.; Gelernter, W. Bt resistance management. Nat. Biotechnol. 1998, 16, 144–146. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.B.; Tabashnik, B.E. Experimental evidence that refuges delay insect adaptation to Bacillus thuringiensis. Proc. R. Soc. B Biol. Sci. 1997, 264, 605–610. [Google Scholar] [CrossRef]
- Wirth, M.C.; Zaritsky, A.; Ben-Dov, E.; Manasherob, R.; Khasdan, V.; Boussiba, S.; Walton, W.E. Cross-resistance spectra of Culex quinquefasciatus resistant to mosquitocidal toxins of Bacillus thuringiensis towards recombinant Escherichia coli expressing genes from B. thuringiensis subsp. israelensis. Environ. Microbiol. 2007, 9, 1393–1401. [Google Scholar] [CrossRef]
- Alphey, L. Genetic control of mosquitoes. Annu. Rev. Entomol. 2014, 59, 205–224. [Google Scholar] [CrossRef]
- Alphey, L.; Benedict, M.; Bellini, R.; Clark, G.G.; Dame, D.A.; Service, M.W.; Dobson, S.L. Sterile-Insect Methods for Control of Mosquito-Borne Diseases: An Analysis. Vector Borne Zoonotic Dis. 2010, 10, 295–311. [Google Scholar] [CrossRef]
- Lambrechts, L.; Koella, J.C.; Boëte, C. Can transgenic mosquitoes afford the fitness cost? Trends Parasitol. 2008, 24, 4–7. [Google Scholar] [CrossRef] [PubMed]
- FAO/IAEA. Status and Risk Assessment of the Use of Transgenic Arthropods in Plant Protection. In Proceedings of the a Technical Meeting Organized by the Joint FAO/IAEA Programme of Nuclear Techniques in Food and Agriculture and the Secretariat of the International Plant Protection Convention, Rome, Italy, 8–12 April 2002. [Google Scholar]
- Benedict, M. The first releases of transgenic mosquitoes: An argument for the sterile insect technique. Trends Parasitol. 2003, 19, 349–355. [Google Scholar] [CrossRef]
- Dame, D.; Curtis, C.; Benedict, M.; Robinson, A.; Knols, B. Historical applications of induced sterilization in field populations of mosquitoes. Malar. J. 2009, 8, S2. [Google Scholar] [CrossRef] [PubMed]
- Windbichler, N.; Papathanos, P.A.; Crisanti, A. Targeting the X chromosome during spermatogenesis induces Y chromosome transmission ratio distortion and early dominant embryo lethality in Anophels gambiae. PLoS Genet. 2008, 4, e1000291. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Genet. 2008, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Burt, A.; Trivers, R. Genes in Conflict: The Biology of Selfish Genetic Elements; Belknap Press/Harvard University Press: Cambridge, MA, USA, 2006; p. 610. [Google Scholar]
- Burt, A. Site-specific selfish genes as tools for the control and genetic engineering of natural populations. Proc. R. Soc. B Biol. Sci. 2003, 270, 921–928. [Google Scholar] [CrossRef]
- Chen, C.H.; Huang, H.; Ward, C.M.; Su, J.T.; Schaeffer, L.V.; Guo, M.; Hay, B.A. A synthetic maternal-effect selfish genetic element drives population replacement in Drosophila. Science 2007, 316, 597–600. [Google Scholar] [CrossRef]
- Lee, S.W.; Chang, S.H.; Lai, Y.S.; Lin, C.C.; Tsai, C.M.; Lee, Y.C.; Chen, J.C.; Huang, C.L. Effect of Temperature on the Growth of Silver Nanoparticles Using Plasmon-Mediated Method under the Irradiation of Green LEDs. Materials 2014, 7, 7781–7798. [Google Scholar] [CrossRef]
- Santos-Magalhães, N.S.; Mosqueira, V.C.F. Nanotechnology applied to the treatment of malaria. Adv. Drug Deliv. Rev. 2010, 62, 560–575. [Google Scholar] [CrossRef]
- Lasic, D.D. Novel applications of liposomes. Trends Biotechnol. 1998, 16, 307–321. [Google Scholar] [CrossRef]
- Peeters, P.A.M.; Huiskamp, C.W.E.M.; Eling, W.M.C.; Crommelin, D.J.A. Chloroquine containing liposomes in the chemotherapy of murine malaria. Parasitology 1989, 98, 381–386. [Google Scholar] [CrossRef] [PubMed]
- Agrawal, A.K.; Singhal, A.; Gupta, C. Functional drug targeting to erythrocytes in vivo using antibody bearing liposomes as drug vehicles. Biochem. Biophys. Res. Commun. 1987, 148, 357–361. [Google Scholar] [CrossRef]
- Wu, Y.; Yang, W.; Wang, C.; Hu, J.; Fu, S. Chitosan nanoparticles as a novel delivery system for ammonium glycyrrhizinate. Int. J. Pharm. 2005, 295, 235–245. [Google Scholar] [CrossRef] [PubMed]
- Nakache, E.; Poulain, N.; Candau, F.; Orecchioni, A.; Irache, J. Biopolymer and Polymer Nanoparticles and Their Biomedical Applications. In Handbook of Nanostructured Materials and Nanotechnology; Nalwa, H.S., Ed.; Academic Press: New York, NY, USA, 2000. [Google Scholar]
- Baird, J.K.; Hoffman, S.L. Primaquine therapy for malaria. Clin. Infect. Dis. 2000, 39, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Omwoyo, W.N.; Ogutu, B.; Oloo, F.; Swai, H.; Kalombo, L.; Melariri, P.; Mahanga, G.M.; Gathirwa, J.W. Preparation, characterization, and optimization of primaquine-loaded solid lipid nanoparticles. Int. J. Nanomed. 2014, 9, 3865–3874. [Google Scholar]
- Baird, J.; Rieckmann, K.H. Can primaquine therapy for vivax malaria be improved? Trends Parasitol. 2003, 19, 115–120. [Google Scholar] [CrossRef]
- Dennis, E.; Peoples, V.A.; Johnson, F.; Bibbs, R.K.; Topps, D.; Bopda-Waffo, A.; Coats, M.T. Utilizing Nanotechnology to Combat Malaria. J. Infect. Dis. Ther. 2015, 3, 4. [Google Scholar] [CrossRef]
- Ahmed, T.A.; Aljaeid, B.M. Preparation, characterization, and potential application of chitosan, chitosan derivatives, and chitosan metal nanoparticles in pharmaceutical drug delivery. Drug Des. Dev. Ther. 2016, 10, 483–507. [Google Scholar] [CrossRef]
- Kilama, W.; Ntoumi, F. Malaria: A research agenda for the eradication era. Lancet 2009, 374, 1480–1482. [Google Scholar] [CrossRef]
- Schwendener, R.A. Liposomes as vaccine delivery systems: A review of the recent advances. Ther. Adv. Vaccines 2014, 2, 159–182. [Google Scholar] [CrossRef]
- Sen, G.L. A brief history of RNAi: The silence of the genes. FASEB J. 2006, 20, 1293–1299. [Google Scholar] [CrossRef] [PubMed]
- Airs, P.M.; Bartholomay, L.C. RNA Interference for Mosquito and Mosquito-Borne Disease Control. Insects 2017, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, J.; Zhu, K.Y. Chitosan/double-stranded RNA nanoparticle-mediated RNA interference to silence chitin synthase genes through larval feeding in the African malaria mosquito (Anophels gambiae). Insect Mol. Biol. 2010, 19, 683–693. [Google Scholar] [CrossRef] [PubMed]
- Bennet, D.; Kim, S. Polymer Nanoparticles for Smart Drug Delivery. In Application of Nanotechnology in Drug Delivery; IntechOpen: London, UK, 2014. [Google Scholar]
- Moon, J.J.; Suh, H.; Polhemus, M.E.; Ockenhouse, C.F.; Yadava, A.; Irvine, D.J. Antigen-Displaying Lipid-Enveloped PLGA Nanoparticles as Delivery Agents for a Plasmodium vivax Malaria Vaccine. PLoS ONE 2012, 7, e31472. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, S.; Das, S.; Chakraborty, S.P.; Sahu, S.K.; Pramanik, P.; Roy, S. Synthesis, characterization of chitosan--tripolyphosphate conjugated chloroquine nanoparticle and it’s in vivo anti-malarial efficacy against rodent parasite: A dose and duration dependent approach. Int. J. Pharm. 2012, 434, 292–305. [Google Scholar] [CrossRef]
- Tripathy, S.; Das, S.; Dash, S.K.; Chattopadhyay, S.; Roy, S. The impact of nanochloroquine on restoration of hepatic and splenic. J. Nanopart. 2013, 9, 106152. [Google Scholar] [CrossRef]
- Benelli, G.; Maggi, F.; Pavela, R.; Murugan, K.; Govindarajan, M.; Vaseeharan, B.; Petrelli, R.; Cappellacci, L.; Kumar, S.; Hofer, A.; et al. Mosquito control with green nanopesticides: Towards the One Health approach? A review of non-target effects. Environ. Sci. Pollut. Res. 2017, 25, 10184–10206. [Google Scholar] [CrossRef]
- Benelli, G. Plant-mediated biosynthesis of nanoparticles as an emerging tool against mosquitoes of medical and veterinary importance: A review. Parasitol. Res. 2016, 115, 23–34. [Google Scholar] [CrossRef]
- Azarudeen, R.M.S.T.; Govindarajan, M.; Amsath, A.; Muthukumaran, U.; Benelli, G. Single-step biofabrication of silver nanocristals using Naregamia alata: A cost effective and eco-friendly control tool in the fight against Malaria, Zika virus and St Louis encephalitis mosquito vectors. J. Clust. Sci. 2017, 28, 179–203. [Google Scholar] [CrossRef]
- Benelli, G. Green synthesized nanoparticles in the fight against mosquito-borne diseases and cancer—a brief review. Enzym. Microb. Technol. 2016, 95, 58–68. [Google Scholar] [CrossRef]
- Panneerselvam, C.; Murugan, K.; Roni, M.; Aziz, A.T.; Suresh, U.; Rajaganesh, R.; Madhiyazhagan, P.; Subramaniam, J.; Dinesh, D.; Nicoletti, M.; et al. Fern-synthesize nanoparticles in the fight against malaria: LC/MS analysis of Pteridium aquilinum leaf extract and biosynthesis of silver nanoparticles with high mosquitocidal and antiplasmodial activity. Parasitol. Res. 2016, 115, 997–1013. [Google Scholar] [CrossRef] [PubMed]
- Alto, B.W.; Juliano, S.A. Precipitation and temperature effects of populations of Ae. albopictus (Diptera: Culicidae): Implications for range expansion. J. Med. Entomol. 2001, 38, 646–656. [Google Scholar] [CrossRef] [PubMed]
- Roni, M.; Murugan, K.; Panneerselvam, C.; Subramaniam, J.; Nicoletti, M.; Madhiyazhagan, P.; Dinesh, D.; Suresh, U.; Khater, H.F.; Wei, H.; et al. Characterization and biotoxicity of Hypnea musciformis-synthesized silver nanoparticles as potential eco-friendly control tool against Ae. aegypti and Plutella xylostella. Ecotoxicol. Environ. Saf. 2015, 121, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Costa, E.A.P.A.; Santos, E.M.; Correia, J.C.; Albuquerque, C.M.R. Impact of small variations in temperature and humidity on the reproductive activity and survival of Aedes aegypti (Diptera, Culicidae). Rev. Bras. Entomol. 2010, 54, 488–493. [Google Scholar] [CrossRef]
- Becker, N. Influence of climate change on mosquito development and mosquito-borne diseases in Europe. Parasitol. Res. 2008, 103, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Canyon, D.V.; Hii, J.L.K.; Muller, R. Adaptation of Aedes aegypti (Diptera: Culicidae) oviposition behaviour in response to humidity and diet. J. Insect Physiol. 1999, 45, 959–964. [Google Scholar] [CrossRef]
- Reiter, P.; Amador, M.A.; Anderson, R.A.; Clark, G.G. Short report: Dispersal of Ae. aegypti in an urban area after blood feeding as demonstrated by rubidium-marked eggs. Am. J. Trop. Med. Hyg. 1995, 52, 177–179. [Google Scholar] [CrossRef]
- Mogi, M.; Miyagi, I.; Abadi, K.; Syafruddin. Inter- and intraspecific variation in resistance to desiccation by adult Aedes (Stegomyia) spp. (Diptera: Culicidae) from Indonesia. Med. Vet. Entomol. 1996, 33, 53–57. [Google Scholar]
- Ahmed, T.; Scholz, M.; Al-Faraj, F.; Niaz, W. Water-Related Impacts of Climate Change on Agriculture and Subsequently on Public Health: A Review for Generalists with Particular Reference to Pakistan. Int. J. Environ. Res. Public Health 2016, 13, 1051. [Google Scholar] [CrossRef]
- Kreft, S.; Eckstein, D.; Melchior, I. Global Climate Risk Index 2017 Who Suffers Most from Extreme Weather Events? Weather-related Loss Events in 2015 and 1996 to 2015, 2017; Germanwatch e.V.: Bonn, Germany, 2014. [Google Scholar]
- Jahan, F. Short report: Dengue Fever (DF) in Pakistan. Asia Pac. Fam. Med. 2011, 10, 1. [Google Scholar] [CrossRef]
- Kazmi, D.H.; Li, J.; Ruan, C.; Zhao, S.; Li, Y.; Sen, Z. A statistical downscaling model for summer rainfall over Pakistan. Clim. Dyn. 2016, 47, 2653–2666. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Impacts of Temperature on Certain Vectors and Vector-Borne Pathogens | Impacts of Changes in Precipitation on Vector-Borne Pathogens | Impacts of Higher Sea Level on Vector-Borne Pathogens |
|---|---|---|
Vector
| Vector
| Vector Reducing or eradicating breeding sites of mosquitoes (e.g., Cs. melanura). The relationship between vector biology and climatic conditions is complex due to the natural capability of vectors to seek out appropriate microclimates for their survival such as hiding places under vegetation, pit toilets during hot or dry weather and tunnels during cold weather [23]. |
| Pathogen | Pathogen
| |
Vertebrate host
|
| Name | International Union of Pure and Applied Chemistry Name | Molar Mass | Pack Size | Applications | Reference |
|---|---|---|---|---|---|
| Fendona 10% [C22H19Cl2NO3] | [cyano-(3-phenoxyphenyl) methyl] 3-(2,2-dichloroethenyl)-2,2-dimethylcyclopropane-1-carboxylate | 416.300 g/mol | 100, 200 and 1000 mL | It has a broad action spectrum against a large variety of insect pests in households and public facilities. | [61] |
| Lambda Cyhalothrin [C23H19ClF3NO3] | cyano-(3-phenoxyphenyl) methyl] (1R,3R)-3-[(Z)-2-chloro-3,3,3-trifluoroprop-1-enyl]-2,2 dimethylcyclopropane-1-carboxylate | 449.854 g/mol | 250 and 500 mL | Targets aphids, butterfly larvae, cockroaches, mosquitoes and flies. | [62] |
| Deltamethrin [C22H19Br2NO3] | Cyano (3-phenoxy-phenyl) methyl; 3-(2,2dibromoethenyl)-2,2-dimethylcyclopropanecarboxylate (CA); [partial diff]-cyano-m-phenoxy benzyl, (1R,3R)-3-(2,2-dibromovinyl)-2,2-dimethyl -cyclopropanol-carboxylate, (S)-[partial diff]-cyano-3-phenoxybenzyl (1R)-cis-3-(2,2-dibromovinyl)-2,2-dimethylcyclopropane-carboxylate | 505.240 g/mol | 25 g/L | Control of the malaria vector as a stomach poison for insects. | [63] |
| Fenpropathrin [C22H23NO3] | 2,2,3,3-Tetramethylcyclopropane carboxylic acid cyano(3-phenoxyphenyl) methyl ester | 349.42 g/mol | 1000 L (mini-mum order) | Controls a variety of pests; specifically, mites present in fruits and vegetables. | [64] |
| System | Active Compound | Mechanism | Plant | References |
|---|---|---|---|---|
| Cholinergic system | Essential oils | Acetylecholinestrase (AChE) inhibition | Azadirachtina indica, Lavendula spp. and Mentha spp. | [70] |
| Veratrin | Nerves sodium channels | Schoenocaulon officinale | [71] | |
| Nicotine | Cholinergic acetylcholine nicotinic receptor agonist/antagonist | Nicotiana spp., Haloxylon salicornicum, Delphinium spp. and Stemona japonicum | [72] | |
| GABA system | Thymol and Silphinenes | GABA-gated chloride channel | Thymus vulgaris | [73] |
| Mitochondrial system | Pyrethrin | Potassium ion and sodium exchange disturbance | Crysanthemum cinerariaefolium | [74] |
| Rotenone | Cellular respiration inhibitor (mitochondrial complex I electron transportinhibitor or METI) | Lonchocarpus spp. | [75] | |
| Ryanodine | Calcium channel disturbance | Ryania spp. | [76] | |
| Sabadilla | Alter nerve cell membrane action | Schoenocaulon officinale | [77] | |
| Octopaminergic system | Essential oils | Octopaminergic receptors | Cedrus spp., Pinus spp., Citronella spp. and Eucalyptus spp. | [78,79] |
| Thymol | Block octopamine receptors by functioning through tyramine receptors cascade | Thymus vulgaris | [78,80] | |
| Miscellaneous | Azadirachtin | Anti-mitotic (G2/M Phase) | Azadiractina indica | [71] |
| Temperature | Relative Humidity of 60% | Relative Humidity of 80% | ||
|---|---|---|---|---|
| Eggs (Sample Number) | Oviposition Variation | Eggs (Sample Number) | Oviposition Variation | |
| 25 °C | 85.99 ± 3.16 (102) | 4–160 (37.25%) | 99.08 ± 3.56 (92) | 4–155 (55.43%) |
| 30 °C | 82.89 ± 3.33 (111) | 2–143 (37.84%) | 75.75 ± 5.03 (75) | 1–144 (45.33%) |
| 35 °C | 54.53 ± 4.81 (55) | 1–126 (14.55%) | 59.62 ± 3.41 (79) | 2–132 (7.59%) |
| Total | 78.25 ± 2.2 (268) | 79.29 ± 2.53 (246) | ||
| Reports | Saudi Arabia | Sri Lanka | Pakistan | Nepal | Malaysia | India | China | Bangladesh |
|---|---|---|---|---|---|---|---|---|
| Reported cases | 1600 | 35,008 | 1085 | 30 | 41,486 | 17,000 | 322 | 510 |
| Mortality | N/A | 346 | 13 | 0 | 90 | 105 | 0 | 0 |
| Reported cases | 2200 | 34,188 | 11,024 | 917 | 46,171 | 28,000 | 260 | 405 |
| Mortality | N/A | 229 | 40 | 5 | 135 | 112 | 0 | 0 |
| Reported cases | 2400 | 28,473 | 17,057 | 79 | 19,884 | 19,000 | 160 | 1310 |
| Mortality | N/A | 246 | 219 | 0 | 38 | 171 | 0 | 8 |
| Reported cases | 800 | 44,461 | 639 (Karachi only) | 183 | 21,900 | 50,000 | 610 | 750 |
| Mortality | N/A | 220 | 4 | 0 | 37 | 240 | 0 | 1 |
| Reported cases | 4200 | 32,063 | 4388 | 642 | 43,346 | 70,000 | 4779 | 1790 |
| Mortality | N/A | N/A | 32 | 0 | 95 | 140 | 0 | 3 |
| Reported cases | 1950 | 47,502 | 1400 | 355 | 108,698 | 40,571 | 47,056 | 355 |
| Mortality | N/A | N/A | 16 | 0 | 215 | 137 | 6 | 0 |
| Reported cases | 4200 | 29,777 | 3900 | 1500 | 120,836 | 99,913 | 4230 | 3162 |
| Mortality | N/A | N/A | 11 | 0 | 336 | 220 | 0 | N/A |
| Reported cases | N/A | 55,150 | 2500 | 594 till October | 100,028 | 1,29,166 | 2098 | 6010 |
| Mortality | N/A | N/A | 3 | N/A | 231 | 245 | N/A | N/A |
| Reported cases | N/A | 185,688 | N/A | 854 till October | 82,840 | 1,53,635 | 3195 | N/A |
| Mortality | N/A | N/A | N/A | N/A | 171 | 226 | N/A | N/A |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmed, T.; Hyder, M.Z.; Liaqat, I.; Scholz, M. Climatic Conditions: Conventional and Nanotechnology-Based Methods for the Control of Mosquito Vectors Causing Human Health Issues. Int. J. Environ. Res. Public Health 2019, 16, 3165. https://doi.org/10.3390/ijerph16173165
Ahmed T, Hyder MZ, Liaqat I, Scholz M. Climatic Conditions: Conventional and Nanotechnology-Based Methods for the Control of Mosquito Vectors Causing Human Health Issues. International Journal of Environmental Research and Public Health. 2019; 16(17):3165. https://doi.org/10.3390/ijerph16173165
Chicago/Turabian StyleAhmed, Toqeer, Muhammad Zeeshan Hyder, Irfan Liaqat, and Miklas Scholz. 2019. "Climatic Conditions: Conventional and Nanotechnology-Based Methods for the Control of Mosquito Vectors Causing Human Health Issues" International Journal of Environmental Research and Public Health 16, no. 17: 3165. https://doi.org/10.3390/ijerph16173165
APA StyleAhmed, T., Hyder, M. Z., Liaqat, I., & Scholz, M. (2019). Climatic Conditions: Conventional and Nanotechnology-Based Methods for the Control of Mosquito Vectors Causing Human Health Issues. International Journal of Environmental Research and Public Health, 16(17), 3165. https://doi.org/10.3390/ijerph16173165