Education and Lifestyle Factors Are Associated with DNA Methylation Clocks in Older African Americans

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Sample
2.2. Methylation Measures
2.3. DNAm Age Calculation and Blood Cell Counts
2.4. Assessment of Education, Lifestyle Factors and Other Covariates
2.5. Statistical Analyses
3. Results
3.1. Descriptive Statistics
3.2. Correlation among DNAm Age Estimators
3.3. Associations between Education or Lifestyle Risk Factors and DNAm Age Acceleration
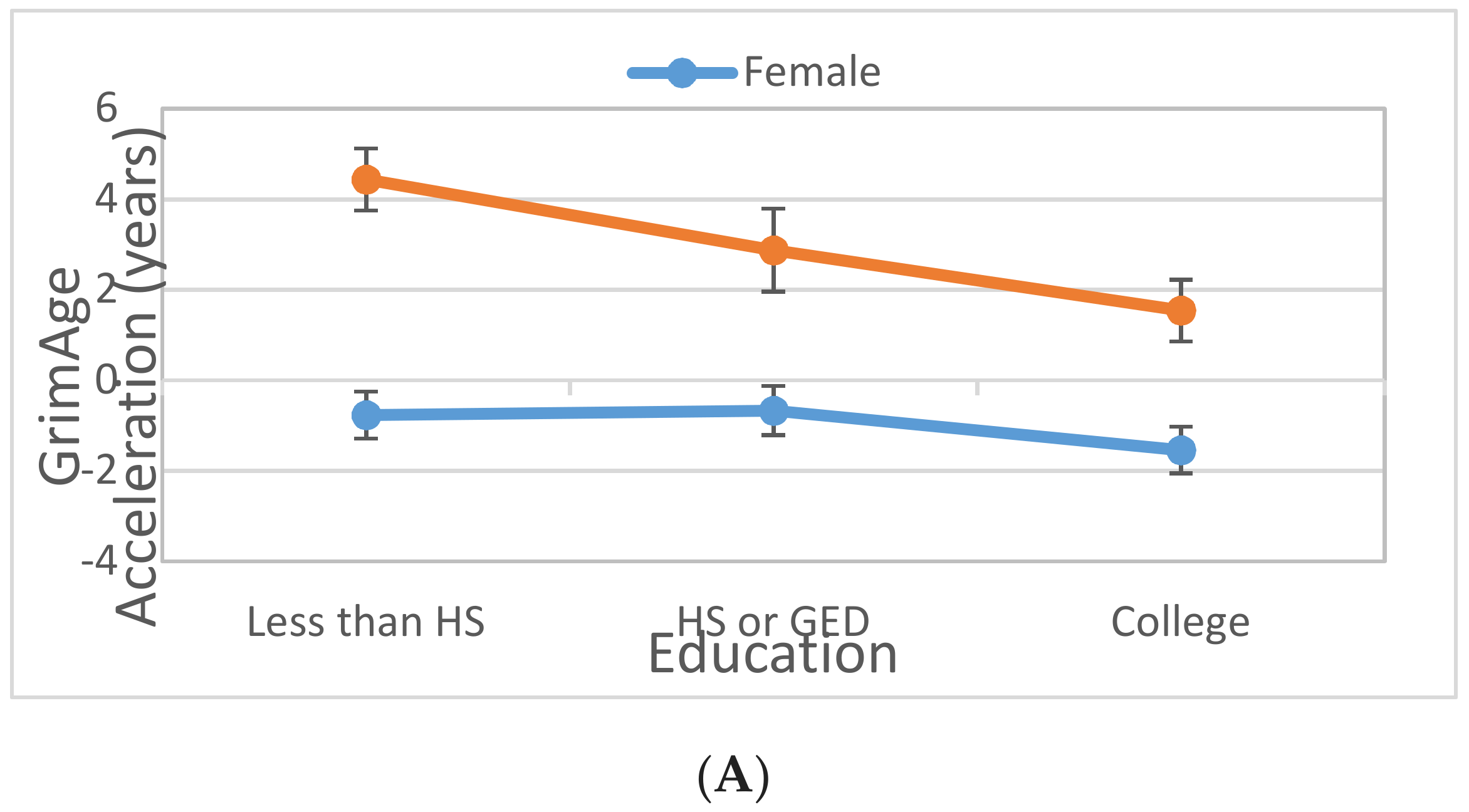
3.4. Interaction between Gender and Lifestyle Factors on DNAm Age Acceleration
3.5. Association between GrimAge Components and Education or Lifestyle Risk Factors
3.6. Association between Education or Lifestyle Factors and Longitudinal Change in DNAm Age Acceleration
4. Discussion
4.1. Associations with GrimAge Acceleration and Its Components
4.2. Gender and DNAm Clocks
4.3. Education and DNAm Clocks
4.4. Smoking and DNAm Clocks
4.5. BMI and DNAm Clocks
4.6. Alcohol Consumption and DNAm Clocks
4.7. Physical Activity and DNAm Clocks
4.8. Lifestyle Factors and Longitudinal Change in DNAm Age Acceleration
4.9. Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Marioni, R.E.; Suderman, M.; Chen, B.H.; Horvath, S.; Bandinelli, S.; Morris, T.; Beck, S.; Ferrucci, L.; Pedersen, N.L.; Relton, C.L.; et al. Tracking the Epigenetic Clock Across the Human Life Course: A Meta-analysis of Longitudinal Cohort Data. J. Gerontol. Ser. A 2018, 74, 57–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Field, A.E.; Robertson, N.A.; Wang, T.; Havas, A.; Ideker, T.; Adams, P.D. DNA Methylation Clocks in Aging: Categories, Causes, and Consequences. Mol. Cell 2018, 71, 882–895. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hannum, G.; Guinney, J.; Zhao, L.; Zhang, L.; Hughes, G.; Sadda, S.; Klotzle, B.; Bibikova, M.; Fan, J.B.; Gao, Y.; et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol. Cell 2013, 49, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S. DNA methylation age of human tissues and cell types. Genome Biol. 2013, 14, R115. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.E.; Lu, A.T.; Quach, A.; Chen, B.H.; Assimes, T.L.; Bandinelli, S.; Hou, L.; Baccarelli, A.A.; Stewart, J.D.; Li, Y.; et al. An epigenetic biomarker of aging for lifespan and healthspan. Aging 2018, 10, 573–591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, A.T.; Quach, A.; Wilson, J.G.; Reiner, A.P.; Aviv, A.; Raj, K.; Hou, L.; Baccarelli, A.A.; Li, Y.; Stewart, J.D.; et al. DNA methylation GrimAge strongly predicts lifespan and healthspan. Aging 2019, 11, 303–327. [Google Scholar] [CrossRef] [PubMed]
- Marioni, R.E.; Shah, S.; McRae, A.F.; Ritchie, S.J.; Muniz-Terrera, G.; Harris, S.E.; Gibson, J.; Redmond, P.; Cox, S.R.; Pattie, A.; et al. The epigenetic clock is correlated with physical and cognitive fitness in the Lothian Birth Cohort 1936. Int. J. Epidemiol. 2015, 44, 1388–1396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perna, L.; Zhang, Y.; Mons, U.; Holleczek, B.; Saum, K.U.; Brenner, H. Epigenetic age acceleration predicts cancer, cardiovascular, and all-cause mortality in a German case cohort. Clin. Epigenetics 2016, 8, 64. [Google Scholar] [CrossRef]
- Horvath, S.; Ritz, B.R. Increased epigenetic age and granulocyte counts in the blood of Parkinson’s disease patients. Aging 2015, 7, 1130–1142. [Google Scholar] [CrossRef]
- Levine, M.E.; Lu, A.T.; Bennett, D.A.; Horvath, S. Epigenetic age of the pre-frontal cortex is associated with neuritic plaques, amyloid load, and Alzheimer’s disease related cognitive functioning. Aging 2015, 7, 1198–1211. [Google Scholar] [CrossRef]
- Horvath, S.; Langfelder, P.; Kwak, S.; Aaronson, J.; Rosinski, J.; Vogt, T.F.; Eszes, M.; Faull, R.L.; Curtis, M.A.; Waldvogel, H.J.; et al. Huntington’s disease accelerates epigenetic aging of human brain and disrupts DNA methylation levels. Aging 2016, 8, 1485–1512. [Google Scholar] [CrossRef] [PubMed]
- Levine, M.E.; Hosgood, H.D.; Chen, B.; Absher, D.; Assimes, T.; Horvath, S. DNA methylation age of blood predicts future onset of lung cancer in the women’s health initiative. Aging 2015, 7, 690–700. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.H.; Marioni, R.E.; Colicino, E.; Peters, M.J.; Ward-Caviness, C.K.; Tsai, P.C.; Roetker, N.S.; Just, A.C.; Demerath, E.W.; Guan, W.; et al. DNA methylation-based measures of biological age: Meta-analysis predicting time to death. Aging 2016, 8, 1844–1865. [Google Scholar] [CrossRef] [PubMed]
- Marioni, R.E.; Shah, S.; McRae, A.F.; Chen, B.H.; Colicino, E.; Harris, S.E.; Gibson, J.; Henders, A.K.; Redmond, P.; Cox, S.R.; et al. DNA methylation age of blood predicts all-cause mortality in later life. Genome Biol. 2015, 16, 25. [Google Scholar] [CrossRef] [PubMed]
- Christiansen, L.; Lenart, A.; Tan, Q.; Vaupel, J.W.; Aviv, A.; McGue, M.; Christensen, K. DNA methylation age is associated with mortality in a longitudinal Danish twin study. Aging Cell 2016, 15, 149–154. [Google Scholar] [CrossRef]
- Fraga, M.F.; Ballestar, E.; Paz, M.F.; Ropero, S.; Setien, F.; Ballestar, M.L.; Heine-Suner, D.; Cigudosa, J.C.; Urioste, M.; Benitez, J.; et al. Epigenetic differences arise during the lifetime of monozygotic twins. Proc. Natl. Acad. Sci. USA 2005, 102, 10604–10609. [Google Scholar] [CrossRef] [Green Version]
- Madsen, M.; Andersen, A.M.; Christensen, K.; Andersen, P.K.; Osler, M. Does educational status impact adult mortality in Denmark? A twin approach. Am. J. Epidemiol. 2010, 172, 225–234. [Google Scholar] [CrossRef]
- Cutler, D.M.; Lleras-Muney, A. Understanding differences in health behaviors by education. J. Health Econ. 2010, 29, 1–28. [Google Scholar] [CrossRef] [Green Version]
- Conti, G.; Heckman, J.; Urzua, S. The Education-Health Gradient. Am. Econ. Rev. 2010, 100, 234–238. [Google Scholar] [CrossRef] [Green Version]
- Quach, A.; Levine, M.E.; Tanaka, T.; Lu, A.T.; Chen, B.H.; Ferrucci, L.; Ritz, B.; Bandinelli, S.; Neuhouser, M.L.; Beasley, J.M.; et al. Epigenetic clock analysis of diet, exercise, education, and lifestyle factors. Aging 2017, 9, 419–446. [Google Scholar] [CrossRef] [Green Version]
- Van Dongen, J.; Nivard, M.G.; Willemsen, G.; Hottenga, J.J.; Helmer, Q.; Dolan, C.V.; Ehli, E.A.; Davies, G.E.; van Iterson, M.; Breeze, C.E.; et al. Genetic and environmental influences interact with age and sex in shaping the human methylome. Nat. Commun. 2016, 7, 11115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fiorito, G.; McCrory, C.; Robinson, O.; Carmeli, C.; Rosales, C.O.; Zhang, Y.; Colicino, E.; Dugue, P.A.; Artaud, F.; McKay, G.J.; et al. Socioeconomic position, lifestyle habits and biomarkers of epigenetic aging: A multi-cohort analysis. Aging 2019, 11, 2045–2070. [Google Scholar] [CrossRef] [PubMed]
- Horvath, S.; Gurven, M.; Levine, M.E.; Trumble, B.C.; Kaplan, H.; Allayee, H.; Ritz, B.R.; Chen, B.; Lu, A.T.; Rickabaugh, T.M.; et al. An epigenetic clock analysis of race/ethnicity, sex, and coronary heart disease. Genome Biol. 2016, 17, 171. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Chen, B.H.; Assimes, T.L.; Ferrucci, L.; Horvath, S.; Levine, M.E. The role of epigenetic aging in education and racial/ethnic mortality disparities among older U.S. Women. Psychoneuroendocrinology 2019, 104, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Daniels, P.R.; Kardia, S.L.; Hanis, C.L.; Brown, C.A.; Hutchinson, R.; Boerwinkle, E.; Turner, S.T.; Genetic Epidemiology Network of Arteriopathy, s. Familial aggregation of hypertension treatment and control in the Genetic Epidemiology Network of Arteriopathy (GENOA) study. Am. J. Med. 2004, 116, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Fortin, J.-P.; Fertig, E.; Hansen, K. shinyMethyl: Interactive quality control of Illumina 450k DNA methylation arrays in R. F1000Research 2014, 3, 175. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Niu, L.; Li, L.; Taylor, J.A. ENmix: A novel background correction method for Illumina HumanMethylation450 BeadChip. Nucleic Acids Res. 2015, 44, e20. [Google Scholar] [CrossRef] [PubMed]
- Fortin, J.P.; Triche, T.J., Jr.; Hansen, K.D. Preprocessing, normalization and integration of the Illumina HumanMethylationEPIC array with minfi. Bioinformatics 2017, 33, 558–560. [Google Scholar] [CrossRef]
- Niu, L.; Xu, Z.; Taylor, J.A. RCP: A novel probe design bias correction method for Illumina Methylation BeadChip. Bioinformatics 2016, 32, 2659–2663. [Google Scholar] [CrossRef]
- Lehne, B.; Drong, A.W.; Loh, M.; Zhang, W.; Scott, W.R.; Tan, S.T.; Afzal, U.; Scott, J.; Jarvelin, M.R.; Elliott, P.; et al. A coherent approach for analysis of the Illumina HumanMethylation450 BeadChip improves data quality and performance in epigenome-wide association studies. Genome Biol. 2015, 16, 37. [Google Scholar] [CrossRef]
- DNA Methylation Age Calculator: New Methylation Age Calculator. Available online: https://dnamage.genetics.ucla.edu/new (accessed on 28 August 2019).
- McEwen, L.M.; Jones, M.J.; Lin, D.T.S.; Edgar, R.D.; Husquin, L.T.; MacIsaac, J.L.; Ramadori, K.E.; Morin, A.M.; Rider, C.F.; Carlsten, C.; et al. Systematic evaluation of DNA methylation age estimation with common preprocessing methods and the Infinium MethylationEPIC BeadChip array. Clin. Epigenetics 2018, 10, 123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhingra, R.; Kwee, L.C.; Diaz-Sanchez, D.; Devlin, R.B.; Cascio, W.; Hauser, E.R.; Gregory, S.; Shah, S.; Kraus, W.E.; Olden, K. Evaluating DNA methylation age on the Illumina MethylationEPIC Bead Chip. PloS ONE 2019, 14, e0207834. [Google Scholar] [CrossRef] [PubMed]
- Houseman, E.A.; Accomando, W.P.; Koestler, D.C.; Christensen, B.C.; Marsit, C.J.; Nelson, H.H.; Wiencke, J.K.; Kelsey, K.T. DNA methylation arrays as surrogate measures of cell mixture distribution. BMC Bioinformatics 2012, 13, 86. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Leung, D.; Levine, M. Comparative analysis of epigenetic aging clocks from CpG characteristics to functional associations. bioRxiv 2019, 512483. [Google Scholar] [CrossRef] [Green Version]
- Jougasaki, M.; Burnett, J.C., Jr. Adrenomedullin: Potential in physiology and pathophysiology. Life Sci. 2000, 66, 855–872. [Google Scholar] [CrossRef]
- Murty, M.; Sharma, U.; Pandey, V.; Kankare, S. Serum cystatin C as a marker of renal function in detection of early acute kidney injury. Ind. J. Nephrol. 2013, 23, 180. [Google Scholar] [CrossRef] [PubMed]
- Taglieri, N.; Koenig, W.; Kaski, J.C. Cystatin C and cardiovascular risk. Clin. Chem. 2009, 55, 1932–1943. [Google Scholar] [CrossRef]
- Shlipak, M.G.; Sarnak, M.J.; Katz, R.; Fried, L.F.; Seliger, S.L.; Newman, A.B.; Siscovick, D.S.; Stehman-Breen, C. Cystatin C and the risk of death and cardiovascular events among elderly persons. N. Eng. J. Med. 2005, 352, 2049–2060. [Google Scholar] [CrossRef]
- Knight, E.L.; Verhave, J.C.; Spiegelman, D.; Hillege, H.L.; De Zeeuw, D.; Curhan, G.C.; De Jong, P.E. Factors influencing serum cystatin C levels other than renal function and the impact on renal function measurement. Kidney Int. 2004, 65, 1416–1421. [Google Scholar] [CrossRef] [Green Version]
- Juhan-Vague, I.; Alessi, M. PAI-1, obesity, insulin resistance and risk of cardiovascular events. Thrombos. Haemost. 1997, 78, 656–660. [Google Scholar] [CrossRef]
- Alessi, M.-C.; Juhan-Vague, I. PAI-1 and the metabolic syndrome: Links, causes, and consequences. Arterioscler. Thrombos. Vasc. Biol. 2006, 26, 2200–2207. [Google Scholar] [CrossRef] [PubMed]
- Barton, P.J.; Birks, E.J.; Felkin, L.E.; Cullen, M.E.; Koban, M.U.; Yacoub, M.H. Increased expression of extracellular matrix regulators TIMP1 and MMP1 in deteriorating heart failure. J. Heart Lung Transplant. 2003, 22, 738–744. [Google Scholar] [CrossRef]
- Fong, K.M.; Kida, Y.; Zimmerman, P.V.; Smith, P.J. TIMP1 and adverse prognosis in non-small cell lung cancer. Clin. Cancer Res. 1996, 2, 1369–1372. [Google Scholar] [PubMed]
- Lee, S.W.; Song, K.E.; Shin, D.S.; Ahn, S.M.; Ha, E.S.; Kim, D.J.; Nam, M.S.; Lee, K.-W. Alterations in peripheral blood levels of TIMP-1, MMP-2, and MMP-9 in patients with type-2 diabetes. Diabetes Res. Clin. Pract. 2005, 69, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Bataille, R.; Boccadoro, M.; Klein, B.; Durie, B.; Pileri, A. C-reactive protein and beta-2 microglobulin produce a simple and powerful myeloma staging system. Blood 1992, 80, 733–737. [Google Scholar] [PubMed]
- Yilmaz, H.; Çelik, H.; Gurel, O.; Bilgic, M.; Namuslu, M.; Bozkurt, H.; Ayyildiz, A.; Inan, O.; Bavbek, N.; Akcay, A. Increased serum levels of GDF-15 associated with mortality and subclinical atherosclerosis in patients on maintenance hemodialysis. Herz 2015, 40, 305–312. [Google Scholar] [CrossRef]
- Rohatgi, A.; Patel, P.; Das, S.R.; Ayers, C.R.; Khera, A.; Martinez-Rumayor, A.; Berry, J.D.; McGuire, D.K.; De Lemos, J.A. Association of growth differentiation factor-15 with coronary atherosclerosis and mortality in a young, multiethnic population: Observations from the Dallas Heart Study. Clin. Chem. 2012, 58, 172–182. [Google Scholar] [CrossRef]
- Eggers, K.M.; Kempf, T.; Wallentin, L.; Wollert, K.C.; Lind, L. Change in growth differentiation factor 15 concentrations over time independently predicts mortality in community-dwelling elderly individuals. Clin. Chem. 2013, 59, 1091–1098. [Google Scholar] [CrossRef]
- Skau, E.; Henriksen, E.; Wagner, P.; Hedberg, P.; Siegbahn, A.; Leppert, J. GDF-15 and TRAIL-R2 are powerful predictors of long-term mortality in patients with acute myocardial infarction. Eur. J. Prev. Cardiol. 2017, 24, 1576–1583. [Google Scholar] [CrossRef]
- Ahima, R.S.; Flier, J.S. Leptin. Ann. Rev. Physiol. 2000, 62, 413–437. [Google Scholar] [CrossRef] [Green Version]
- Simpkin, A.J.; Hemani, G.; Suderman, M.; Gaunt, T.R.; Lyttleton, O.; McArdle, W.L.; Ring, S.M.; Sharp, G.C.; Tilling, K.; Horvath, S.; et al. Prenatal and early life influences on epigenetic age in children: A study of mother-offspring pairs from two cohort studies. Hum. Mol. Genet. 2016, 25, 191–201. [Google Scholar] [CrossRef] [PubMed]
- Tan, Q.; Heijmans, B.T.; Hjelmborg, J.V.; Soerensen, M.; Christensen, K.; Christiansen, L. Epigenetic drift in the aging genome: A ten-year follow-up in an elderly twin cohort. Int. J. Epidemiol. 2016, 45, 1146–1158. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, P.; Huen, K.; Dave, V.; Barcellos, L.; Eskenazi, B.; Holland, N. Sex differences in DNA methylation assessed by 450 K BeadChip in newborns. BMC Genom. 2015, 16, 911. [Google Scholar] [CrossRef] [PubMed]
- Ryan, J.; Wrigglesworth, J.; Loong, J.; Fransquet, P.D.; Woods, R.L. A systematic review and meta-analysis of environmental, lifestyle and health factors associated with DNA methylation age. J. Gerontol. A Biol. Sci. Med. Sci. 2019, glz099. [Google Scholar] [CrossRef] [PubMed]
- Chen, E.; Miller, G.E.; Yu, T.; Brody, G.H. The Great Recession and health risks in African American youth. Brain Behav. Immun. 2016, 53, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Simons, R.L.; Lei, M.K.; Beach, S.R.; Philibert, R.A.; Cutrona, C.E.; Gibbons, F.X.; Barr, A. Economic hardship and biological weathering: The epigenetics of aging in a U.S. sample of black women. Soc. Sci. Med. 2016, 150, 192–200. [Google Scholar] [CrossRef] [PubMed]
- Joehanes, R.; Just, A.C.; Marioni, R.E.; Pilling, L.C.; Reynolds, L.M.; Mandaviya, P.R.; Guan, W.; Xu, T.; Elks, C.E.; Aslibekyan, S.; et al. Epigenetic Signatures of Cigarette Smoking. Circ. Cardiovasc. Genet. 2016, 9, 436–447. [Google Scholar] [CrossRef] [Green Version]
- Barcelona, V.; Huang, Y.; Brown, K.; Liu, J.; Zhao, W.; Yu, M.; Kardia, S.L.R.; Smith, J.A.; Taylor, J.Y.; Sun, Y.V. Novel DNA methylation sites associated with cigarette smoking among African Americans. Epigenetics 2019, 14, 383–391. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Zhao, W.; Ammous, F.; Turner, S.T.; Mosley, T.H.; Zhou, X.; Smith, J.A. Longitudinal analysis of epigenome-wide DNA methylation reveals novel smoking-related loci in African Americans. Epigenetics 2019, 14, 171–184. [Google Scholar] [CrossRef] [Green Version]
- Lindbohm, J.V.; Kaprio, J.; Jousilahti, P.; Salomaa, V.; Korja, M. Sex, Smoking, and Risk for Subarachnoid Hemorrhage. Stroke 2016, 47, 1975–1981. [Google Scholar] [CrossRef] [Green Version]
- Sørheim, I.-C.; Johannessen, A.; Gulsvik, A.; Bakke, P.S.; Silverman, E.K.; DeMeo, D.L. Gender differences in COPD: Are women more susceptible to smoking effects than men? Thorax 2010, 65, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Krishnan, E.; Sokka, T.; Hannonen, P. Smoking-gender interaction and risk for rheumatoid arthritis. Arthritis Res. Ther. 2003, 5, R158–R162. [Google Scholar] [CrossRef] [PubMed]
- Janisch, F.; Shariat, S.F.; Schernhammer, E.; Rink, M.; Fajkovic, H. The interaction of gender and smoking on bladder cancer risks. Curr. Opin. Urol. 2019, 29, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Dick, K.J.; Nelson, C.P.; Tsaprouni, L.; Sandling, J.K.; Aïssi, D.; Wahl, S.; Meduri, E.; Morange, P.-E.; Gagnon, F.; Grallert, H. DNA methylation and body-mass index: A genome-wide analysis. Lancet 2014, 383, 1990–1998. [Google Scholar] [CrossRef]
- Mendelson, M.M.; Marioni, R.E.; Joehanes, R.; Liu, C.; Hedman, Å.K.; Aslibekyan, S.; Demerath, E.W.; Guan, W.; Zhi, D.; Yao, C. Association of body mass index with DNA methylation and gene expression in blood cells and relations to cardiometabolic disease: A Mendelian randomization approach. PLoS Med. 2017, 14, e1002215. [Google Scholar] [CrossRef] [PubMed]
- Wahl, S.; Drong, A.; Lehne, B.; Loh, M.; Scott, W.R.; Kunze, S.; Tsai, P.-C.; Ried, J.S.; Zhang, W.; Yang, Y. Epigenome-wide association study of body mass index, and the adverse outcomes of adiposity. Nature 2017, 541, 81. [Google Scholar] [CrossRef] [PubMed]
- Richmond, R.C.; Sharp, G.C.; Ward, M.E.; Fraser, A.; Lyttleton, O.; McArdle, W.L.; Ring, S.M.; Gaunt, T.R.; Lawlor, D.A.; Smith, G.D. DNA methylation and BMI: Investigating identified methylation sites at HIF3A in a causal framework. Diabetes 2016, 65, 1231–1244. [Google Scholar] [CrossRef]
- Morales, E.; Groom, A.; Lawlor, D.A.; Relton, C.L. DNA methylation signatures in cord blood associated with maternal gestational weight gain: Results from the ALSPAC cohort. BMC Res. Notes 2014, 7, 278. [Google Scholar] [CrossRef]
- Liu, X.; Chen, Q.; Tsai, H.J.; Wang, G.; Hong, X.; Zhou, Y.; Zhang, C.; Liu, C.; Liu, R.; Wang, H. Maternal preconception body mass index and offspring cord blood DNA methylation: Exploration of early life origins of disease. Environ. Mol. Mutagen. 2014, 55, 223–230. [Google Scholar] [CrossRef]
- Beach, S.R.; Dogan, M.V.; Lei, M.K.; Cutrona, C.E.; Gerrard, M.; Gibbons, F.X.; Simons, R.L.; Brody, G.H.; Philibert, R.A. Methylomic aging as a window onto the influence of lifestyle: Tobacco and alcohol use alter the rate of biological aging. J. Am. Geriatr. Soc. 2015, 63, 2519–2525. [Google Scholar] [CrossRef]
- Chan, A.W. Racial differences in alcohol sensitivity. Alcohol Alcohol. 1986, 21, 93–104. [Google Scholar] [PubMed]
- Liu, C.; Marioni, R.E.; Hedman, Å.K.; Pfeiffer, L.; Tsai, P.-C.; Reynolds, L.M.; Just, A.C.; Duan, Q.; Boer, C.G.; Tanaka, T. A DNA methylation biomarker of alcohol consumption. Mol. Psychiatry 2018, 23, 422. [Google Scholar] [CrossRef] [PubMed]
- Dugue, P.A.; Bassett, J.K.; Joo, J.E.; Baglietto, L.; Jung, C.H.; Wong, E.M.; Fiorito, G.; Schmidt, D.; Makalic, E.; Li, S.; et al. Association of DNA Methylation-Based Biological Age With Health Risk Factors and Overall and Cause-Specific Mortality. Am. J. Epidemiol. 2018, 187, 529–538. [Google Scholar] [CrossRef] [PubMed]
- Sillanpää, E.; Ollikainen, M.; Kaprio, J.; Wang, X.; Leskinen, T.; Kujala, U.M.; Törmäkangas, T. Leisure-time physical activity and DNA methylation age—A twin study. Clin. Epigen. 2019, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Gale, C.R.; Marioni, R.E.; Cukic, I.; Chastin, S.F.; Dall, P.M.; Dontje, M.L.; Skelton, D.A.; Deary, I.J.; Seniors USP Team. The epigenetic clock and objectively measured sedentary and walking behavior in older adults: The Lothian Birth Cohort 1936. Clin. Epigen. 2018, 10, 4. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Karlsson, R.; Lampa, E.; Zhang, Q.; Hedman, Å.K.; Almgren, M.; Almqvist, C.; McRae, A.F.; Marioni, R.E.; Ingelsson, E. Epigenetic influences on aging: A longitudinal genome-wide methylation study in old Swedish twins. Epigenetics 2018, 13, 975–987. [Google Scholar] [CrossRef]
- Jylhävä, J.; Hjelmborg, J.; Soerensen, M.; Munoz, E.; Tan, Q.; Kuja-Halkola, R.; Mengel-From, J.; Christensen, K.; Christiansen, L.; Hägg, S. Longitudinal changes in the genetic and environmental influences on the epigenetic clocks across old age: Evidence from two twin cohorts. EBioMedicine 2019, 40, 710–716. [Google Scholar] [CrossRef] [Green Version]
- Wilson, R.; Wahl, S.; Pfeiffer, L.; Ward-Caviness, C.K.; Kunze, S.; Kretschmer, A.; Reischl, E.; Peters, A.; Gieger, C.; Waldenberger, M. The dynamics of smoking-related disturbed methylation: A two time-point study of methylation change in smokers, non-smokers and former smokers. BMC Genom. 2017, 18, 805. [Google Scholar] [CrossRef]
- Nevalainen, T.; Kananen, L.; Marttila, S.; Jylhävä, J.; Mononen, N.; Kähönen, M.; Raitakari, O.T.; Hervonen, A.; Jylhä, M.; Lehtimäki, T. Obesity accelerates epigenetic aging in middle-aged but not in elderly individuals. Clin. Epigenet. 2017, 9, 20. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Cross-Sectional (N = 1100) | Longitudinal (N = 266) | |||
|---|---|---|---|---|
| Variable Name | Mean (SD) or N (%) | Phase 1 Mean (SD) or N (%) | Phase 2 Mean (SD) or N (%) 1 | Pearson r or Kappa 2 |
| HorvathAge (years) | 53.89 (9.98) | 51.19 (9.18) | 55.12 (9.19) *** | 0.919 |
| HannumAge (years) | 47.71 (10.75) | 44.27 (9.75) | 49.14 (10.02) *** | 0.960 |
| PhenoAge (years) | 44.22 (12.66) | 40.21 (11.62) | 44.68 (11.69) *** | 0.922 |
| GrimAge (years) | 54.31 (9.49) | 52.00 (9.09) | 55.44 (9.15) *** | 0.972 |
| IEAA (years) | 0.15 (4.79) | −0.01 (4.77) | −0.46 (5.10) * | 0.748 |
| EEAA (years) | 0.27 (5.85) | −0.36 (5.54) | −1.02 (6.52) ** | 0.852 |
| PhenoAA (years) | 0.38 (7.17) | −0.54 (6.81) | −1.46 (6.27) ** | 0.764 |
| GrimAA (years) | 0.11 (4.95) | 0.23 (4.56) | −0.58 (4.63) *** | 0.911 |
| Chronological Age (years) | 57.05 (10.48) | 53.97 (9.77) | 59.42 (9.35) *** | 0.994 |
| Gender (male) | 319 (29.00%) | 77 (28.95%) | ||
| Education | ||||
| Less than HS | 374 (34.00%) | 80 (30.08%) | ||
| HS/GED | 292 (26.55%) | 71 (26.69%) | ||
| At least some college | 434 (39.45%) | 115 (43.23%) | ||
| Smoking | ||||
| Never | 666 (60.55%) | 160 (60.15%) | ||
| Former smoker | 255 (23.18%) | 63 (23.68%) | ||
| Current smoker | 179 (16.27%) | 43 (16.17%) | ||
| Continuous drinks/week | 0.67 (2.68) | 0.71 (2.82) | 0.52 (1.69) | 0.491 |
| Physical activity (hrs/day) | 1.07 (1.57) | 1.14 (1.42) | ||
| Body Mass Index (kg/m2) | 31.20 (6.48) | 31.53 (6.69) | 32.04 (6.83) *** | 0.946 |
| GrimAA | PhenoAA | IEAA | EEAA | |||||
|---|---|---|---|---|---|---|---|---|
| Beta | p-Value | Beta | p-Value | Beta | p-Value | Beta | p-Value | |
| Gender (male) | 2.410 | 1.94 × 10−17 | −0.834 | 0.106 | 1.038 | 0.003 | 2.405 | 1.94 × 10−8 |
| Education | ||||||||
| HS/GED | −0.098 | 0.745 | −0.812 | 0.153 | −0.669 | 0.081 | −1.016 | 0.028 |
| At least some college | −0.605 | 0.041 | −1.107 | 0.051 | −0.743 | 0.050 | −1.784 | 1.02 × 10−4 |
| Smoking | ||||||||
| Former smoker | 2.337 | 2.67 × 10−15 | 1.317 | 0.015 | 0.948 | 0.009 | 0.039 | 0.928 |
| Current smoker | 7.618 | 4.98 × 10−81 | 2.135 | 0.001 | 0.045 | 0.915 | 0.672 | 0.191 |
| Continuous ln(drinks/week) | 0.455 | 0.033 | 0.561 | 0.159 | 0.376 | 0.164 | −0.044 | 0.891 |
| BMI | 0.040 | 0.029 | 0.080 | 0.022 | 0.038 | 0.112 | 0.035 | 0.216 |
| GrimAA Interaction Model 1 | GrimAA Interaction Model 2 | GrimAA Interaction Model 3 | ||||
|---|---|---|---|---|---|---|
| Beta | p-Value | Beta | p-Value | Beta | p-Value | |
| Gender (male) | 3.450 | 8.79 × 10−16 | 1.684 | 2.43 × 10−5 | 2.698 | 4.75 × 10−7 |
| Education | ||||||
| HS/GED | 0.439 | 0.205 | −0.108 | 0.720 | 0.414 | 0.231 |
| At least some college | −0.114 | 0.739 | −0.592 | 0.045 | −0.175 | 0.610 |
| Smoking | ||||||
| Former smoker | 2.365 | 1.06 × 10−15 | 2.213 | 1.42 × 10−9 | 2.287 | 4.08 × 10−10 |
| Current smoker | 7.622 | 2.34 × 10−81 | 6.877 | 1.46 × 10−51 | 6.928 | 2.57 × 10−52 |
| Continuous ln (drinks/week) | 0.462 | 0.031 | 0.372 | 0.083 | 0.380 | 0.077 |
| BMI | 0.045 | 0.016 | 0.041 | 0.028 | 0.044 | 0.017 |
| Gender (male)*Education | ||||||
| HS/GED | −1.929 | 0.003 | −1.899 | 0.003 | ||
| At least some college | −1.513 | 0.006 | −1.272 | 0.022 | ||
| Gender (male)*Smoking | ||||||
| Former smoker | 0.786 | 0.189 | 0.636 | 0.290 | ||
| Current smoker | 2.170 | 0.001 | 2.026 | 0.003 | ||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, W.; Ammous, F.; Ratliff, S.; Liu, J.; Yu, M.; Mosley, T.H.; Kardia, S.L.R.; Smith, J.A. Education and Lifestyle Factors Are Associated with DNA Methylation Clocks in Older African Americans. Int. J. Environ. Res. Public Health 2019, 16, 3141. https://doi.org/10.3390/ijerph16173141
Zhao W, Ammous F, Ratliff S, Liu J, Yu M, Mosley TH, Kardia SLR, Smith JA. Education and Lifestyle Factors Are Associated with DNA Methylation Clocks in Older African Americans. International Journal of Environmental Research and Public Health. 2019; 16(17):3141. https://doi.org/10.3390/ijerph16173141
Chicago/Turabian StyleZhao, Wei, Farah Ammous, Scott Ratliff, Jiaxuan Liu, Miao Yu, Thomas H. Mosley, Sharon L. R. Kardia, and Jennifer A. Smith. 2019. "Education and Lifestyle Factors Are Associated with DNA Methylation Clocks in Older African Americans" International Journal of Environmental Research and Public Health 16, no. 17: 3141. https://doi.org/10.3390/ijerph16173141
APA StyleZhao, W., Ammous, F., Ratliff, S., Liu, J., Yu, M., Mosley, T. H., Kardia, S. L. R., & Smith, J. A. (2019). Education and Lifestyle Factors Are Associated with DNA Methylation Clocks in Older African Americans. International Journal of Environmental Research and Public Health, 16(17), 3141. https://doi.org/10.3390/ijerph16173141