Creating an Internal Environment of Cognitive and Psycho-Emotional Well-Being through an External Movement-Based Environment: An Overview of Quadrato Motor Training



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
1.1. Neuro-Bio-Sociological Environments are Substrates for Cognitive and Psycho-Emotional Well-Being
1.2. The Impact of An External Movement-Based Environment on the Internal Environment
2. QMT Effects: Neuropsychological Research
2.1. Electrophysiological Effects of QMT
2.2. Neuroanatomical Effects of QMT
2.3. Molecular Effects of QMT
2.4. Behavioral Effects of QMT and Associations with Other Parameters
3. What May Differentiate QMT from Other Forms of MM?
4. Conclusions and Implication for Environmental Research and Public Health
Author Contributions
Funding
Conflicts of Interest
References
- Charles, S.T.; Carstensen, L.L. Social and Emotional Aging. Annu. Rev. Psychol. 2010, 61, 383–409. [Google Scholar] [CrossRef] [PubMed]
- Zammit, A.R.; Starr, J.M.; Johnson, W.; Deary, I.J. Profiles of physical, emotional and psychosocial wellbeing in the Lothian birth cohort 1936. BMC Geriatr 2012, 12, 64. [Google Scholar] [CrossRef] [PubMed]
- Riggs, N.R.; Shin, H.-S.; Unger, J.B.; Spruijt-Metz, D.; Pentz, M.A. Prospective Associations between Bilingualism and Executive Function in Latino Children: Sustained Effects While Controlling for Biculturalism. J. Immigr. Minor. Health 2014, 16, 914–921. [Google Scholar] [CrossRef] [PubMed]
- Diamond, A. Close Interrelation of Motor Development and Cognitive Development and of the Cerebellum and Prefrontal Cortex. Child Dev. 2000, 71, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Fuchs, E.; Flügge, G. Adult Neuroplasticity: More Than 40 Years of Research. Neural Plast. 2014, 2014, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Ganguly, K.; Poo, M. Activity-Dependent Neural Plasticity from Bench to Bedside. Neuron 2013, 80, 729–741. [Google Scholar] [CrossRef] [PubMed]
- Hübener, M.; Bonhoeffer, T. Neuronal Plasticity: Beyond the Critical Period. Cell 2014, 159, 727–737. [Google Scholar] [CrossRef]
- Zilles, K. Neuronal plasticity as an adaptive property of the central nervous system. Ann. Anat. Anat. Anz. 1992, 174, 383–391. [Google Scholar] [CrossRef]
- Glenn, A.L.; Raine, A. Neurocriminology: Implications for the punishment, prediction and prevention of criminal behaviour. Nat. Rev. Neurosci. 2014, 15, 54–63. [Google Scholar] [CrossRef]
- Lenroot, R.K.; Giedd, J.N. The changing impact of genes and environment on brain development during childhood and adolescence: Initial findings from a neuroimaging study of pediatric twins. Dev. Psychopathol. 2008, 20, 1161–1175. [Google Scholar] [CrossRef]
- Robinson, G.E.; Fernald, R.D.; Clayton, D.F. Genes and social behavior. Science 2008, 322, 896–900. [Google Scholar] [CrossRef] [PubMed]
- Dang, L.C.; O’Neil, J.P.; Jagust, W.J. Genetic effects on behavior are mediated by neurotransmitters and large-scale neural networks. Neuroimage 2013, 66, 203–214. [Google Scholar] [CrossRef] [PubMed]
- McAdams, T.A.; Gregory, A.M.; Eley, T.C. Genes of Experience: Explaining the Heritability of Putative Environmental Variables Through Their Association with Behavioural and Emotional Traits. Behav. Genet. 2013, 43, 314–328. [Google Scholar] [CrossRef] [PubMed]
- Wootton, R.E.; Davis, O.S.P.; Mottershaw, A.L.; Wang, R.A.H.; Haworth, C.M.A. Genetic and environmental correlations between subjective wellbeing and experience of life events in adolescence. Eur. Child Adolesc. Psychiatry 2017, 26, 1119–1127. [Google Scholar] [CrossRef] [PubMed]
- Kanherkar, R.R.; Bhatia-Dey, N.; Csoka, A.B. Epigenetics across the human lifespan. Front. Cell Dev. Biol. 2014, 2, 49. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, S.; Mariotti, V.; Iofrida, C.; Pellegrini, S. Genes and Aggressive Behavior: Epigenetic Mechanisms Underlying Individual Susceptibility to Aversive Environments. Front. Behav. Neurosci. 2018, 12, 117. [Google Scholar] [CrossRef] [PubMed]
- Beckman, M. Neuroscience: Crime, culpability, and the adolescent brain. Science 2004, 305, 596–599. [Google Scholar] [CrossRef]
- Brenhouse, H.C.; Andersen, S.L. Developmental trajectories during adolescence in males and females: A cross-species understanding of underlying brain changes. Neurosci. Biobehav. Rev. 2011, 35, 1687–1703. [Google Scholar] [CrossRef]
- Leshem, R. Using Dual Process Models to Examine Impulsivity Throughout Neural Maturation. Dev. Neuropsychol. 2016, 41, 125–143. [Google Scholar] [CrossRef]
- Shulman, E.P.; Smith, A.R.; Silva, K.; Icenogle, G.; Duell, N.; Chein, J.; Steinberg, L. The dual systems model: Review, reappraisal, and reaffirmation. Dev. Cognit. Neurosci. 2016, 17, 103–117. [Google Scholar] [CrossRef]
- Spear, L.P. The adolescent brain and age-related behavioral manifestations. Neurosci. Biobehav. Rev. 2000, 24, 417–463. [Google Scholar] [CrossRef]
- Hebb, D.O. The Organization of Behavior: A Neuropsycholocigal Theory; Wiley Book in Clinical Psychology: New York, NY, USA, 1949. [Google Scholar]
- Cooke, S.F.; Bliss, T.V.P. Plasticity in the human central nervous system. Brain 2006, 129, 1659–1673. [Google Scholar] [CrossRef] [PubMed]
- Shors, T.J.; Anderson, M.L.; Curlik II, D.M.; Nokia, M.S. Use it or lose it: How neurogenesis keeps the brain fit for learning. Behav. Brain Res. 2012, 227, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Leshem, R.; Glicksohn, J. A Critical Look at the Relationship Between Impulsivity and Decision-Making in Adolescents: Are They Related or Separate Factors? Dev. Neuropsychol. 2012, 37, 712–731. [Google Scholar] [CrossRef] [PubMed]
- Luna, B.; Padmanabhan, A.; O’Hearn, K. What has fMRI told us about the Development of Cognitive Control through Adolescence? Brain Cognit. 2010, 72, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Luciana, M.; Collins, P.F.; Olson, E.A.; Schissel, A.M. Tower of London performance in healthy adolescents: The development of planning skills and associations with self-reported inattention and impulsivity. Dev. Neuropsychol. 2009, 34, 461–475. [Google Scholar] [CrossRef]
- Steinberg, L.; Albert, D.; Cauffman, E.; Banich, M.; Graham, S.; Woolard, J. Age differences in sensation seeking and impulsivity as indexed by behavior and self-report: Evidence for a dual systems model. Dev. Psychol. 2008, 44, 1764–1778. [Google Scholar] [CrossRef]
- Pesce, C.; Ben-Soussan, T.D. ‘Cogito ergo sum’ or ‘ambulo ergo sum’? New perspectives in developmental exercise and cognition research. In Exercise-Cognition Interaction: Neuroscience Perspectives; McMorris, T., Ed.; Elsevier: London, UK, 2016; pp. 251–281. [Google Scholar]
- Banks, S.J.; Eddy, K.T.; Angstadt, M.; Nathan, P.J.; Phan, K.L. Amygdala-frontal connectivity during emotion regulation. Soc. Cognit. Affect. Neurosci. 2007, 2, 303–312. [Google Scholar] [CrossRef]
- Boecker, H.; Hillman, C.H.; Scheef, L.; Strüder, H.K. Functional Neuroimaging in Exercise and Sport Sciences; Springer: New York, NY, USA, 2012. [Google Scholar]
- McMorris, T. Exercise-Cognition Interaction: Neuroscience Perspectives; Elsevier: London, UK, 2016. [Google Scholar]
- Ströhle, A. Physical activity, exercise, depression and anxiety disorders. J. Neural Transm. 2009, 116, 777. [Google Scholar] [CrossRef]
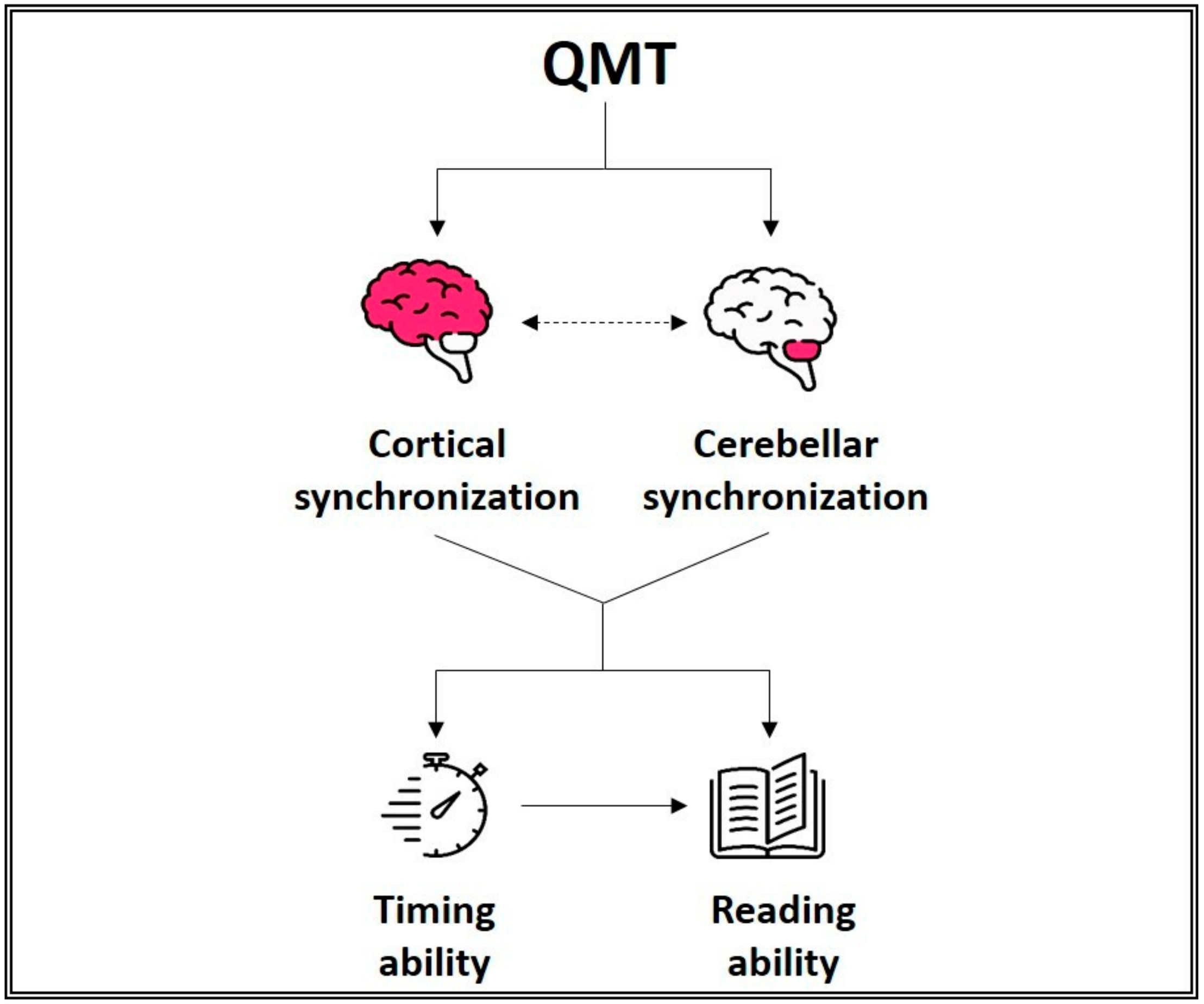
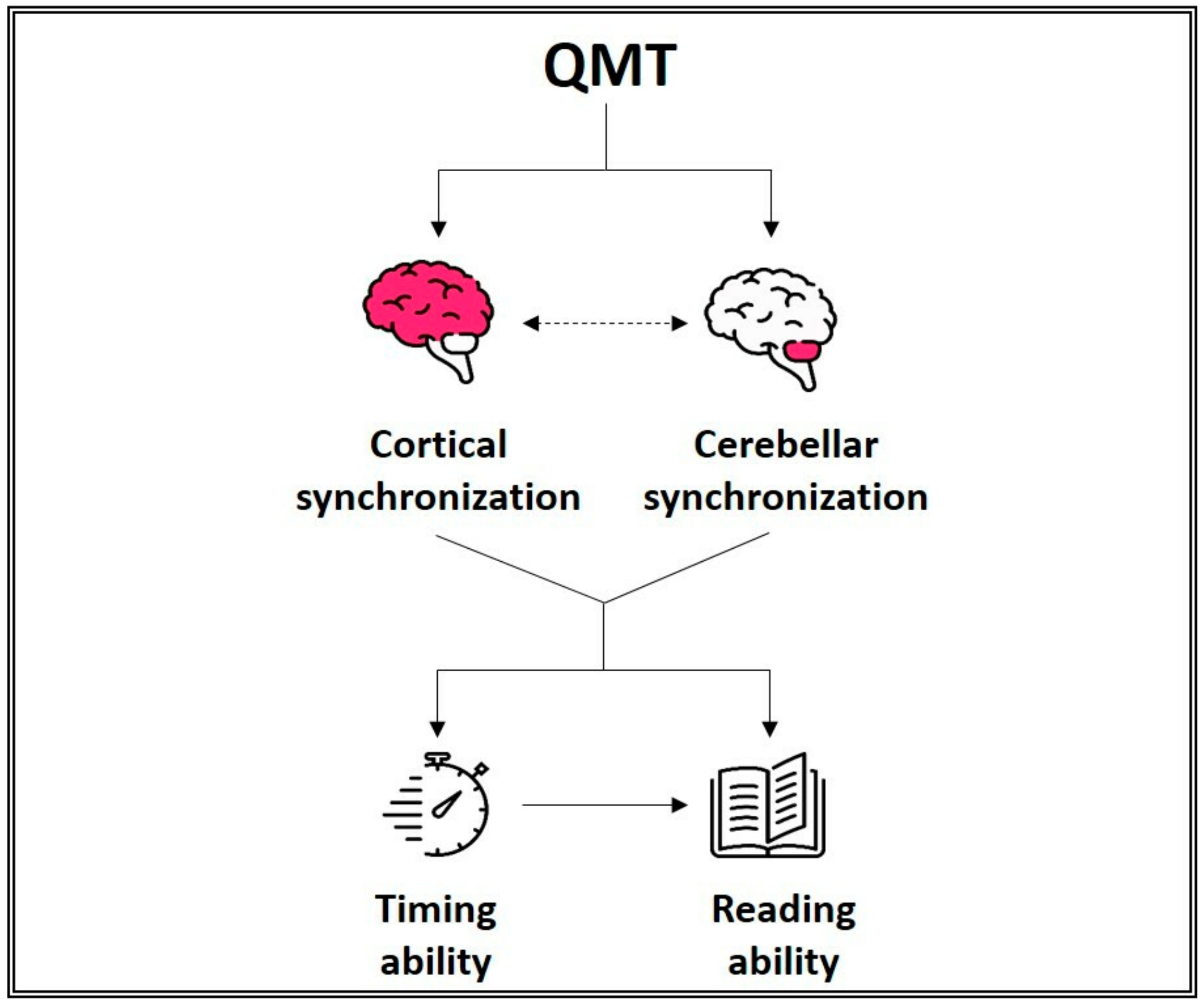
- Ben-Soussan, T.D.; Avirame, K.; Glicksohn, J.; Goldstein, A.; Harpaz, Y.; Ben-Shachar, M. Changes in cerebellar activity and inter-hemispheric coherence accompany improved reading performance following Quadrato Motor Training. Front. Syst. Neurosci. 2014, 8, 81. [Google Scholar] [CrossRef]
- Cirrik, S.; Hacioglu, G. Neurophysiological Effects of Exercise. In Fitness Medicine; Sozen, H., Ed.; IntechOpen: London, UK, 2016; pp. 1014–1150. [Google Scholar]
- Aloe, L.; Luisa Rocco, M.; Omar Balzamino, B.; Micera, A. Nerve growth factor: A focus on neuroscience and therapy. Curr. Neuropharmacol. 2015, 13, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Bathina, S.; Das, U.N. Brain-derived neurotrophic factor and its clinical implications. Arch. Med. Sci. 2015, 11, 1164. [Google Scholar] [CrossRef] [PubMed]
- Venditti, S.; Verdone, L.; Pesce, C.; Tocci, N.; Caserta, M.; Ben-Soussan, T.D. Creating well-being: Increased creativity and proNGF decrease following Quadrato Motor Training. Biomed. Res. Int. 2015, 2015, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lasaponara, S.; Glicksohn, J.; Mauro, F.; Ben-Soussan, T.D. Contingent negative variation and P3 modulations following mindful movement training. Prog. Brain Res. 2019, 244, 101–114. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A. Transient hypofrontality as a mechanism for the psychological effects of exercise. Psychiatry Res. 2006, 145, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.S.Y.; Liu, G.; Liang, D.; Deng, K.; Wu, J.; Yan, J.H. Special Issue-Therapeutic Benefits of Physical Activity for Mood: A Systematic Review on the Effects of Exercise Intensity, Duration, and Modality. J. Psychol. 2019, 153, 102–125. [Google Scholar] [CrossRef]
- Pesce, C. Shifting the Focus from Quantitative to Qualitative Exercise Characteristics in Exercise and Cognition Research. J. Sport Exerc. Psychol. 2012, 34, 766–786. [Google Scholar] [CrossRef] [PubMed]
- Pesce, C.; Faigenbaum, A.D.; Goudas, M.; Tomporowski, P.D. Coupling our plough of thoughtful moving to the star of children’s right to play: From neuroscience to multisectoral promotion. In Physical Activity and Educational Achievement: Insights from Exercise Neuroscience; Meeusen, R., Schaefer, S., Tomporowski, P., Bailey, R., Eds.; Routledge: London, UK, 2017; pp. 1–28. [Google Scholar]
- Lobelo, F.; Stoutenberg, M.; Hutber, A. The exercise is medicine global health initiative: A 2014 update. Br. J. Sports Med. 2014, 48, 1627–1633. [Google Scholar] [CrossRef]
- Diamond, A.; Ling, D.S. Conclusions about interventions, programs, and approaches for improving executive functions that appear justified and those that, despite much hype, do not. Dev. Cognit. Neurosci. 2016, 18, 34–48. [Google Scholar] [CrossRef]
- Diamond, A.; Ling, D.S. Aerobic-Exercise and resistance-training interventions have been among the least effective ways to improve executive functions of any method tried thus far. Dev. Cogn. Neurosci. 2018, 10, S1878929318300094. [Google Scholar] [CrossRef]
- Ben-Soussan, T.D.; Glicksohn, J.; Berkovich-Ohana, A. From Cerebellar Activation and Connectivity to Cognition: A Review of the Quadrato Motor Training. Biomed. Res. Int. 2015, 2015, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Paoletti, P. Crescere Nell’eccellenza; Armando Publishing: Rome, Italy, 2008. [Google Scholar]
- Gutmann, B.; Hülsdünker, T.; Mierau, J.; Strüder, H.K.; Mierau, A. Exercise-induced changes in EEG alpha power depend on frequency band definition mode. Neurosci. Lett. 2018, 662, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Henz, D.; Schöllhorn, W.I. EEG Brain Activity in Dynamic Health Qigong Training: Same Effects for Mental Practice and Physical Training? Front. Psychol. 2017, 8, 154. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kamei, T.; Toriumi, Y.; Kimura, H.; Kumano, H.; Ohno, S.; Kimura, K. Decrease in serum cortisol during Yoga exercise is correlated with alpha wave activation. Percept. Mot. Skills 2000, 90, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Lardon, M.T.; Polich, J. EEG changes from long-term physical exercise. Biol. Psychol. 1996, 44, 19–30. [Google Scholar] [CrossRef]
- Liu, Y.; Mimura, K.; Wang, L.; Ikuda, K. Physiological benefits of 24-style Taijiquan exercise in middle-aged Women. J. Physiol. Anthropol. Appl. Hum. Sci. 2003, 22, 219–225. [Google Scholar] [CrossRef]
- Pan, Z.; Su, X.; Fang, Q.; Hou, L.; Lee, Y.; Chen, C.C.; Lamberth, J.; Kim, M.-L. The effects of Tai Chi intervention on healthy elderly by means of neuroimaging and EEG: A systematic review. Front. Aging Neurosci. 2018, 10, 110. [Google Scholar] [CrossRef] [PubMed]
- Petruzzello, S.J.; Landers, D.M. State anxiety reduction and exercise: Does hemispheric activation reflect such changes? Med. Sci. Sports Exerc. 1994, 26, 1028–1035. [Google Scholar] [CrossRef]
- Cahn, B.R.; Polich, J. Meditation states and traits: EEG, ERP, and neuroimaging studies. Psychol. Bull. 2006, 132, 180–211. [Google Scholar] [CrossRef]
- Lomas, T.; Ivtzan, I.; Fu, C.H.Y. A systematic review of the neurophysiology of mindfulness on EEG oscillations. Neurosci. Biobehav. Rev. 2015, 57, 401–410. [Google Scholar] [CrossRef]
- Van Lutterveld, R.; van Dellen, E.; Pal, P.; Yang, H.; Stam, C.J.; Brewer, J. Meditation is associated with increased brain network integration. Neuroimage 2017, 158, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Ben-Soussan, T.D.; Berkovich-Ohana, A.; Glicksohn, J.; Goldstein, A. A suspended act: Increased reflectivity and gender-dependent electrophysiological change following Quadrato Motor Training. Front. Psychol. 2014, 5, 55. [Google Scholar] [CrossRef] [PubMed]
- Lasaponara, S.; Mauro, F.; Ben-Soussan, T.D.; Carducci, F.; Tombini, M.; Quattrocchi, C.C.; Errante, Y.; Mallio, C.A.; Paoletti, P. Electrophysiological indexes of eyes open and closed resting states conditions following the Quadrato Motor Training. Int J. Bioelectromagn. 2016, 18, 99–108. [Google Scholar]
- Lasaponara, S.; Mauro, F.; Carducci, F.; Paoletti, P.; Tombini, M.; Quattrocchi, C.C.; Mallio, C.A.; Errante, Y.; Scarciolla, L.; Ben-Soussan, T.D. Increased alpha band functional connectivity following the Quadrato Motor Training: A longitudinal study. Front. Hum. Neurosci. 2017, 11, 282. [Google Scholar] [CrossRef] [PubMed]
- Dotan Ben-Soussan, T.; Glicksohn, J.; Goldstein, A.; Berkovich-Ohana, A.; Donchin, O. Into the square and out of the box: The effects of Quadrato Motor Training on creativity and alpha coherence. PLoS ONE 2013, 8, e55023. [Google Scholar] [CrossRef]
- Ben-Soussan, T.D.; Glicksohn, J.; Ohana, A.B.; Donchin, O.; Goldstein, A. Step in time: Changes in EEG coherence during a time estimation task following Quadrato Motor Training. In Proceedings of the 27th Annual Meeting of the International Society for Psychophysics, Herzliya, Israel, 24–27 October 2011; Volume 27, pp. 239–244. [Google Scholar]
- Shaw, J.C. Intention as a component of the alpha-rhythm response to mental activity. Int. J. Psychophysiol. 1996, 24, 7–23. [Google Scholar] [CrossRef]
- Ben-Soussan, T.D.; Glicksohn, J.; Berkovich-Ohana, A. Attentional effort, mindfulness, and altered states of consciousness experiences following Quadrato Motor Training. Mindfulness 2017, 8, 59–67. [Google Scholar] [CrossRef]
- Engel, A.K.; Fries, P. Neuronal oscillations, coherence, and consciousness. In The Neurology of Conciousness, 2nd ed.; Laureys, S., Gosseries, O., Tononi, G., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 49–60. [Google Scholar]
- Sauseng, P.; Klimesch, W.; Schabus, M.; Doppelmayr, M. Fronto-parietal EEG coherence in theta and upper alpha reflect central executive functions of working memory. Int. J. Psychophysiol. 2005, 57, 97–103. [Google Scholar] [CrossRef]
- Travis, F.; Tecce, J.; Arenander, A.; Wallace, R.K. Patterns of EEG coherence, power, and contingent negative variation characterize the integration of transcendental and waking states. Biol. Psychol. 2002, 61, 293–319. [Google Scholar] [CrossRef]
- Paoletti, P.; Ben-Soussan, T.D.; Giubilei, F.; Marzano, N.; Lopez, S.; Brunno, E.; Orzi, F.; Bianchi, V.; Babiloni, C.; Buttinelli, C. Subjects with amnesic mild cognitive impairment can do Quadrato Motor Training with possible beneficial effects on resting state electroencephalographic rhythms. In Proceedings of the Federation of European Neuroscience Societies (FENS), Copenhagen, Denmark, 2–6 July 2016. [Google Scholar]
- Nicolson, R.I.; Fawcett, A.J.; Dean, P. Developmental dyslexia: The cerebellar deficit hypothesis. Trends Neurosci. 2001, 24, 508–511. [Google Scholar] [CrossRef]
- Pernet, C.R.; Poline, J.B.; Demonet, J.F.; Rousselet, G.A. Brain classification reveals the right cerebellum as the best biomarker of dyslexia. BMC Neurosci. 2009, 10, 67. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D. Disorders of the cerebellum: Ataxia, dysmetria of thought, and the cerebellar cognitive affective syndrome. J. Neuropsychiatry Clin. Neurosci. 2004, 16, 367–378. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, C.J.; Stein, J.F. The cerebellum and dyslexia. Cortex 2011, 47, 101–116. [Google Scholar] [CrossRef] [PubMed]
- Buckner, R.L. The cerebellum and cognitive function: 25 years of insight from Anatomy and Neuroimaging. Neuron 2013, 80, 807–815. [Google Scholar] [CrossRef] [PubMed]
- Ivry, R.B.; Spencer, R.M.; Zelaznik, H.N.; Diedrichsen, J. The cerebellum and event timing. Ann. N. Y. Acad. Sci. 2002, 978, 302–317. [Google Scholar] [CrossRef] [PubMed]
- Stoodley, C.J.; Schmahmann, J.D. The cerebellum and language: Evidence from patients with cerebellar degeneration. Brain Lang. 2009, 110, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Nicolson, R.I.; Fawcett, A.J.; Dean, P. Time estimation deficits in developmental dyslexia: Evidence of cerebellar involvement. Proc. Biol. Sci. 1995, 259, 43–47. [Google Scholar] [PubMed]
- Babiloni, C.; Del Percio, C.; Lizio, R.; Marzano, N.; Infarinato, F.; Soricelli, A.; Salvatore, E.; Ferri, R.; Bonforte, C.; Tedeschi, G.; et al. Cortical sources of resting state electroencephalographic alpha rhythms deteriorate across time in subjects with amnesic mild cognitive impairment. Neurobiol. Aging 2014, 35, 130–142. [Google Scholar] [CrossRef] [PubMed]
- Babiloni, C.; Benussi, L.; Binetti, G.; Cassetta, E.; Dal Forno, G.; Del Percio, C.; Ferreri, F.; Ferri, R.; Frisoni, G.; Ghidoni, R.; et al. Apolipoprotein E and alpha brain rhythms in mild cognitive impairment: A multicentric Electroencephalogram study. Ann. Neurol. 2006, 59, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Dierks, T.; Jelic, V.; Pascual-Marqui, R.D.; Wahlund, L.-O.; Julin, P.; Linden, D.E.J.; Maurer, K.; Winblad, B.; Nordberg, A. Spatial pattern of cerebral glucose metabolism (PET) correlates with localization of intracerebral EEG-generators in Alzheimer’s disease. Clin. Neurophysiol. 2000, 111, 1817–1824. [Google Scholar] [CrossRef]
- Huang, C.; Wahlund, L.-O.; Dierks, T.; Julin, P.; Winblad, B.; Jelic, V. Discrimination of Alzheimer’s disease and mild cognitive impairment by equivalent EEG sources: A cross-sectional and longitudinal study. Clin. Neurophysiol. 2000, 111, 1961–1967. [Google Scholar] [CrossRef]
- Jeong, J. EEG dynamics in patients with Alzheimer’s disease. Clin. Neurophysiol. 2004, 115, 1490–1505. [Google Scholar] [CrossRef] [PubMed]
- Moretti, D. Computerized processing of EEG–EOG–EMG artifacts for multi-centric studies in EEG oscillations and event-related potentials. Int. J. Psychophysiol. 2003, 47, 199–216. [Google Scholar] [CrossRef]
- Adler, G.; Brassen, S.; Jajcevic, A. EEG coherence in Alzheimer’s dementia. J. Neural Transm. 2003, 110, 1051–1058. [Google Scholar] [CrossRef] [PubMed]
- Babiloni, C.; Triggiani, A.I.; Lizio, R.; Cordone, S.; Tattoli, G.; Bevilacqua, V.; Soricelli, A.; Ferri, R.; Nobili, F.; Gesualdo, L.; et al. Classification of single normal and Alzheimer’s disease individuals from cortical sources of resting state EEG rhythms. Front. Neurosci. 2016, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Jelic, V.; Johansson, S.-E.; Almkvist, O.; Shigeta, M.; Julin, P.; Nordberg, A.; Winblad, B.; Wahlund, L.-O. Quantitative electroencephalography in mild cognitive impairment: Longitudinal changes and possible prediction of Alzheimer’s disease. Neurobiol. Aging 2000, 21, 533–540. [Google Scholar] [CrossRef]
- Knott, V.; Mohr, E.; Mahoney, C.; Ilivitsky, V. Electroencephalographic coherence in Alzheimer’s disease: Comparisons with a control group and population norms. J. Geriatr. Psychiatry Neurol. 2000, 13, 1–8. [Google Scholar] [CrossRef]
- Kumar, R.; Nguyen, H.D.; Macey, P.M.; Woo, M.A.; Harper, R.M. Regional brain axial and radial diffusivity changes during development. J. Neurosci. Res. 2012, 90, 346–355. [Google Scholar] [CrossRef]
- Roberts, R.E.; Anderson, E.J.; Husain, M. White matter microstructure and cognitive function. Neuroscientist 2013, 19, 8–15. [Google Scholar] [CrossRef]
- Song, S.-K.; Yoshino, J.; Le, T.Q.; Lin, S.-J.; Sun, S.-W.; Cross, A.H.; Armstrong, R.C. Demyelination increases radial diffusivity in corpus callosum of mouse brain. Neuroimage 2005, 26, 132–140. [Google Scholar] [CrossRef]
- Song, S.-K.; Sun, S.-W.; Ramsbottom, M.J.; Chang, C.; Russell, J.; Cross, A.H. Dysmyelination revealed through MRI as increased radial (but unchanged axial) diffusion of water. Neuroimage 2002, 17, 1429–1436. [Google Scholar] [CrossRef] [PubMed]
- Caminiti, R.; Carducci, F.; Piervincenzi, C.; Battaglia-Mayer, A.; Confalone, G.; Visco-Comandini, F.; Pantano, P.; Innocenti, G.M. Diameter, Length, Speed, and Conduction Delay of Callosal Axons in Macaque Monkeys and Humans: Comparing Data from Histology and Magnetic Resonance Imaging Diffusion Tractography. J. Neurosci. 2013, 33, 14501–14511. [Google Scholar] [CrossRef] [PubMed]
- Mac Donald, C.; Dikranian, K.; Song, S.; Bayly, P.; Holtzman, D.; Brody, D. Detection of traumatic axonal injury with diffusion tensor imaging in a mouse model of traumatic brain injury. Exp. Neurol. 2007, 205, 116–131. [Google Scholar] [CrossRef] [PubMed]
- Bennett, I.J.; Madden, D.J.; Vaidya, C.J.; Howard, D.V.; Howard, J.H. Age-related differences in multiple measures of white matter integrity: A diffusion tensor imaging study of healthy aging. Hum. Brain Mapp 2009, 31, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Keller, T.A.; Just, M.A. Altering cortical connectivity: Remediation-induced changes in the white matter of poor readers. Neuron 2009, 64, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Batouli, S.A.H.; Saba, V. At least eighty percent of brain grey matter is modifiable by physical activity: A review study. Behav. Brain Res. 2017, 332, 204–217. [Google Scholar] [CrossRef] [PubMed]
- Erickson, K.I.; Leckie, R.L.; Weinstein, A.M. Physical activity, fitness, and gray matter volume. Neurobiol. Aging 2014, 35, S20–S28. [Google Scholar] [CrossRef] [PubMed]
- Froeliger, B.; Garland, E.L.; McClernon, F.J. Yoga meditation practitioners exhibit greater gray matter volume and fewer reported cognitive failures: Results of a preliminary voxel-based morphometric analysis. Evid. Based Complement. Altern. Med. 2012, 2012, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Gothe, N.P.; Hayes, J.M.; Temali, C.; Damoiseaux, J.S. Differences in brain structure and function among Yoga practitioners and controls. Front. Integr. Neurosci. 2018, 12, 26. [Google Scholar] [CrossRef] [PubMed]
- Hölzel, B.K.; Brunsch, V.; Gard, T.; Greve, D.N.; Koch, K.; Sorg, C.; Lazar, S.W.; Milad, M.R. Mindfulness-based stress reduction, fear conditioning, and the uncinate fasciculus: A pilot study. Front. Behav. Neurosci. 2016, 10, 124. [Google Scholar] [CrossRef] [PubMed]
- Hölzel, B.K.; Carmody, J.; Vangel, M.; Congleton, C.; Yerramsetti, S.M.; Gard, T.; Lazar, S.W. Mindfulness practice leads to increases in regional brain gray matter density. Psychiatry Res. 2011, 191, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Last, N.; Tufts, E.; Auger, L.E. The effects of meditation on grey matter atrophy and neurodegeneration: A systematic review. J. Alzheimers Dis. 2017, 56, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Sexton, C.E.; Betts, J.F.; Demnitz, N.; Dawes, H.; Ebmeier, K.P.; Johansen-Berg, H. A systematic review of MRI studies examining the relationship between physical fitness and activity and the white matter of the ageing brain. Neuroimage 2016, 131, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-Y.; Lu, Q.; Fan, M.; Yang, Y.; Posner, M.I. Mechanisms of white matter changes induced by meditation. Proc. Natl. Acad. Sci. USA 2012, 109, 10570–10574. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.-Y.; Lu, Q.; Geng, X.; Stein, E.A.; Yang, Y.; Posner, M.I. Short-term meditation induces white matter changes in the anterior cingulate. Proc. Natl. Acad. Sci. USA 2010, 107, 15649–15652. [Google Scholar] [CrossRef] [PubMed]
- Barysheva, M.; Jahanshad, N.; Foland-Ross, L.; Altshuler, L.L.; Thompson, P.M. White matter microstructural abnormalities in bipolar disorder: A whole brain diffusion tensor imaging study. Neuroimage Clin. 2013, 2, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Mayo, C.D.; Mazerolle, E.L.; Ritchie, L.; Fisk, J.D.; Gawryluk, J.R. Longitudinal changes in microstructural white matter metrics in Alzheimer’s disease. Neuroimage Clin. 2017, 13, 330–338. [Google Scholar] [CrossRef]
- Taubert, M.; Villringer, A.; Ragert, P. Learning-related gray and white matter changes in humans: An update. Neuroscientist 2012, 18, 320–325. [Google Scholar] [CrossRef]
- Ben-Soussan, T.D.; Berkovich-Ohana, A.; Piervincenzi, C.; Glicksohn, J.; Carducci, F. Embodied cognitive flexibility and neuroplasticity following Quadrato Motor Training. Front. Psychol. 2015, 6, 1021. [Google Scholar] [CrossRef]
- Ben-Soussan, T.D.; Piervincenzi, C.; Venditti, S.; Verdone, L.; Caserta, M.; Carducci, F. Increased cerebellar volume and BDNF level following quadrato motor training: Correlation between Cerebellar Volume and BDNF Level. Synapse 2015, 69, 1–6. [Google Scholar] [CrossRef]
- Piervincenzi, C.; Ben-Soussan, T.D.; Mauro, F.; Mallio, C.A.; Errante, Y.; Quattrocchi, C.C.; Carducci, F. White matter microstructural changes following Quadrato Motor Training: A longitudinal study. Front. Hum. Neurosci. 2017, 11, 590. [Google Scholar] [CrossRef] [PubMed]
- Melillo, R.; Leisman, G. Neurobehavioral Disorders of Childhood: An Evolutionary Perspective; Springer: New York, NY, USA, 2010. [Google Scholar]
- Watson, T.C.; Becker, N.; Apps, R.; Jones, M.W. Back to front: Cerebellar connections and interactions with the prefrontal cortex. Front. Syst. Neurosci. 2014, 8, 4. [Google Scholar] [CrossRef]
- Mamah, D.; Conturo, T.E.; Harms, M.P.; Akbudak, E.; Wang, L.; McMichael, A.R.; Gado, M.H.; Barch, D.M.; Csernansky, J.G. Anterior thalamic radiation integrity in schizophrenia: A diffusion-tensor imaging study. Psychiatry Res. 2010, 183, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Van Der Werf, Y.; Jolles, J.; Witter, M.; Uylings, H. Contributions of thalamic nuclei to declarative memory functioning. Cortex 2003, 39, 1047–1062. [Google Scholar] [CrossRef]
- Bruijn, S.M.; Van Impe, A.; Duysens, J.; Swinnen, S.P. White matter microstructural organization and gait stability in older adults. Front. Aging Neurosci. 2014, 6, 104. [Google Scholar] [CrossRef] [PubMed]
- Vercruysse, S.; Leunissen, I.; Vervoort, G.; Vandenberghe, W.; Swinnen, S.; Nieuwboer, A. Microstructural changes in white matter associated with freezing of gait in Parkinson’s disease: FOg-related white matter changes. Mov. Disord. 2015, 30, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Papagno, C.; Miracapillo, C.; Casarotti, A.; Romero Lauro, L.J.; Castellano, A.; Falini, A.; Casaceli, G.; Fava, E.; Bello, L. What is the role of the uncinate fasciculus? Surgical removal and proper name retrieval. Brain 2011, 134, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Von Der Heide, R.J.; Skipper, L.M.; Klobusicky, E.; Olson, I.R. Dissecting the uncinate fasciculus: Disorders, controversies and a hypothesis. Brain 2013, 136, 1692–1707. [Google Scholar] [CrossRef] [PubMed]
- Hofer, S.; Frahm, J. Topography of the human corpus callosum revisited—Comprehensive fiber tractography using diffusion tensor magnetic resonance imaging. NeuroImage 2006, 32, 989–994. [Google Scholar] [CrossRef] [PubMed]
- Lewin, G.R.; Carter, B.D. Neurotrophic Factors; Springer: Berlin, Germany, 2014. [Google Scholar]
- Levi-Montalcini, R.; Hamburger, V. A diffusible agent of mouse sarcoma, producing hyperplasia of sympathetic ganglia and hyperneurotization of viscera in the chick embryo. J. Exp. Zool. 1953, 123, 233–287. [Google Scholar] [CrossRef]
- Barde, Y.A.; Edgar, D.; Thoenen, H. Purification of a new neurotrophic factor from mammalian brain. EMBO J. 1982, 1, 549–553. [Google Scholar] [CrossRef] [PubMed]
- Zoladz, J.A.; Pilc, A. The effect of physical activity on the brain derived neurotrophic factor: From animal to human studies. J. Physiol. Pharmacol. 2010, 61, 533–541. [Google Scholar] [PubMed]
- Caserta, M.; Ben-Soussan, T.D.; Vetriani, V.; Venditti, S.; Verdone, L. Influence of Quadrato Motor Training on salivary proNGF and proBDNF. Front. Neurosci. 2019, 13, 58. [Google Scholar] [CrossRef] [PubMed]
- Maisonpierre, P.C.; Belluscio, L.; Friedman, B.; Alderson, R.F.; Wiegand, S.J.; Furth, M.E.; Lindsay, R.M.; Yancopoulos, G.D. NT-3, BDNF, and NGF in the developing rat nervous system: Parallel as well as reciprocal patterns of expression. Neuron 1990, 5, 501–509. [Google Scholar] [CrossRef]
- Huang, T.; Larsen, K.T.; Ried-Larsen, M.; Møller, N.C.; Andersen, L.B. The effects of physical activity and exercise on brain-derived neurotrophic factor in healthy humans: A review. Scand. J. Med. Sci. Sports 2014, 24, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.M.C.; Wong, M.L.; Lau, B.W.-M.; Lee, J.C.-D.; Yau, S.-Y.; So, K.-F. Aerobic exercise interacts with neurotrophic factors to predict cognitive functioning in adolescents. Psychoneuroendocrinology 2014, 39, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Pareja-Galeano, H.; Brioche, T.; Sanchis-Gomar, F.; Montal, A.; Jovaní, C.; Martínez-Costa, C.; Gomez-Cabrera, M.C.; Viña, J. Impact of exercise training on neuroplasticity-related growth factors in adolescents. J. Musculoskelet. Neuronal Interact. 2013, 13, 368–371. [Google Scholar]
- Babaei, P.; Damirchi, A.; Mehdipoor, M.; Tehrani, B.S. Long term habitual exercise is associated with lower resting level of serum BDNF. Neurosci. Lett. 2014, 566, 304–308. [Google Scholar] [CrossRef]
- Ben-Soussan, T.D.; Glicksohn, J.; De Fano, A.; Mauro, F.; Marson, F.; Modica, M.; Pesce, C. Embodied time: Time production in advanced Quadrato and Aikido practitioners. Psych. J. 2019, 8, 8–16. [Google Scholar] [CrossRef]
- Ben-Soussan, T.D.; Glicksohn, J. Gender-Dependent Changes in Time Production Following Quadrato Motor Training in Dyslexic and Normal Readers. Front. Comput. Neurosci. 2018, 12, 71. [Google Scholar] [CrossRef]
- Paoletti, P.; Glicksohn, J.; Ben-Soussan, T.D. Inner Design Technology: Improved Affect by Quadrato Motor Training. In The Amygdala-Where Emotions Shape Perception, Learning and Memories; Ferry, B., Ed.; InTech: Hicksville, NY, USA, 2017. [Google Scholar]
- Dietrich, A.; Kanso, R. A review of EEG, ERP, and neuroimaging studies of creativity and insight. Psychol. Bull. 2010, 136, 822–848. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, A. Who’s afraid of a cognitive neuroscience of creativity? Methods 2007, 42, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Runco, M.A.; Acar, S. Divergent Thinking as an Indicator of Creative Potential. Creat. Res. J. 2012, 24, 66–75. [Google Scholar] [CrossRef]
- Dietrich, A. The cognitive neuroscience of creativity. Psychon. Bull. Rev. 2004, 11, 1011–1026. [Google Scholar] [CrossRef] [PubMed]
- Glicksohn, J.; Kinberg, Z. Performance on embedded figures tests: Profiling individual differences. J. Individ. Differ. 2009, 30, 152–162. [Google Scholar] [CrossRef]
- Glicksohn, J. Entering trait and context into a cognitive-timer model for time estimation. J. Environ. Psychol. 1996, 16, 361–370. [Google Scholar] [CrossRef]
- Glicksohn, J. Temporal cognition and the phenomenology of time: A multiplicative function for apparent duration. Conscious. Cognit. 2001, 10, 1–25. [Google Scholar] [CrossRef]
- Kujala, J.; Pammer, K.; Cornelissen, P.; Roebroeck, A.; Formisano, E.; Salmelin, R. Phase coupling in a cerebro-cerebellar network at 8–13 Hz during reading. Cereb. Cortex 2007, 17, 1476–1485. [Google Scholar] [CrossRef]
- Tallal, P.; Miller, S.; Fitch, R.H. Neurobiological basis of speech: A case for the preeminence of temporal processing. Ann. N. Y. Acad. Sci. 1993, 682, 27–47. [Google Scholar] [CrossRef]
- Wolff, P.H. Timing precision and rhythm in developmental dyslexia. Read. Writ. 2002, 15, 179–206. [Google Scholar] [CrossRef]
- Wolff, P.H.; Michel, G.F.; Ovrut, M.; Drake, C. Rate and timing precision of motor coordination in developmental dyslexia. Dev. Psychol. 1990, 26, 349–359. [Google Scholar] [CrossRef]
- Glicksohn, J.; Ohana, A.B.; Dotan, T.B.; Goldstein, A.; Donchin, O. Time Production and EEG alpha revisited. Neuroquantology 2009, 7. [Google Scholar] [CrossRef]
- Bekius, A.; Cope, T.E.; Grube, M. The beat to read: A cross-lingual link between rhythmic regularity perception and reading skill. Front. Hum. Neurosci. 2016, 10, 425. [Google Scholar] [CrossRef] [PubMed]
- Wallace, B.A. The Buddhist tradition of Samatha: Methods for refining and examining consciousness. J. Conscious. Stud. 1999, 6, 175–187. [Google Scholar]
- Bradburn, N.M. The Structure of Psychological Well-Being; Aldine: Chicago, IL, USA, 1969. [Google Scholar]
- Meeks, S.; Van Haitsma, K.; Kostiwa, I.; Murrell, S.A. Positivity and Well-being Among Community-Residing Elders and Nursing Home Residents: What Is the Optimal Affect Balance? J. Gerontol. Ser. B Psychol. Sci. Soc. Sci. 2012, 67, 460–467. [Google Scholar] [CrossRef] [PubMed]
- Verkley, H.; Stolk, J. Does happiness lead into idleness. In How Harmful is Happiness? Consequences of Enjoying Life or Not; Veenhoven, R.E., Ed.; Universitaire Pers Rotterdam: Rotterdam, The Netherlands, 1989; pp. 79–93. [Google Scholar]
- Jerusalem, M.; Schwarzer, R. Self-efficacy as a resource factor in stress appraisal processes. In Self-Efficacy: Thought Control of Action; Schwarzer, R., Ed.; Hemisphere Publishing Corp: Washington, DC, USA, 1992; pp. 195–213. [Google Scholar]
- Albaugh, M.D.; Ducharme, S.; Karama, S.; Watts, R.; Lewis, J.D.; Orr, C.; Nguyen, T.-V.; Mckinstry, R.C.; Botteron, K.N.; Evans, A.C.; et al. Anxious/depressed symptoms are related to microstructural maturation of white matter in typically developing youths. Dev. Psychopathol. 2017, 29, 751–758. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.-H.; Wu, Y.-T. Alterations in white matter micro-integrity of the superior longitudinal fasciculus and anterior thalamic radiation of young adult patients with depression. Psychol. Med. 2014, 44, 2825–2832. [Google Scholar] [CrossRef]
- Miyake, A.; Friedman, N.P.; Emerson, M.J.; Witzki, A.H.; Howerter, A.; Wager, T.D. The Unity and Diversity of Executive Functions and Their Contributions to Complex “Frontal Lobe” Tasks: A Latent Variable Analysis. Cognit. Psychol. 2000, 41, 49–100. [Google Scholar] [CrossRef]
- Larkey, L.; Jahnke, R.; Etnier, J.; Gonzalez, J. Meditative Movement as a category of exercise: Implications for research. J. Phys. Act. Health 2009, 6, 230–238. [Google Scholar] [CrossRef]
- Bishop, S.R.; Lau, M.; Shapiro, S.; Carlson, L.; Anderson, N.D.; Carmody, J.; Segal, Z.V.; Abbey, S.; Speca, M.; Velting, D.; et al. Mindfulness: A proposed operational definition. Clin. Psychol. 2004, 11, 230–241. [Google Scholar] [CrossRef]
- Kabat-Zinn, J. Full Catastrophe Living: Using the Wisdom of Your Body and Mind to Face Stress, Pain, and Illness; Dell Publishing: New York, NY, USA, 1991. [Google Scholar]
- Biddle, S.J.; Mutrie, N. Psychology of Physical Activity: Determinants, Well-Being and Interventions; Routledge: Abingdon, UK, 2008. [Google Scholar]
- Tomporowski, P.D.; Pesce, C. Exercise, Sports, and Performance Arts Benefit Cognition via a Common Process. Psychol. Bull. 2019, in press. [Google Scholar] [CrossRef] [PubMed]



© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Fano, A.; Leshem, R.; Ben-Soussan, T.D. Creating an Internal Environment of Cognitive and Psycho-Emotional Well-Being through an External Movement-Based Environment: An Overview of Quadrato Motor Training. Int. J. Environ. Res. Public Health 2019, 16, 2160. https://doi.org/10.3390/ijerph16122160
De Fano A, Leshem R, Ben-Soussan TD. Creating an Internal Environment of Cognitive and Psycho-Emotional Well-Being through an External Movement-Based Environment: An Overview of Quadrato Motor Training. International Journal of Environmental Research and Public Health. 2019; 16(12):2160. https://doi.org/10.3390/ijerph16122160
Chicago/Turabian StyleDe Fano, Antonio, Rotem Leshem, and Tal Dotan Ben-Soussan. 2019. "Creating an Internal Environment of Cognitive and Psycho-Emotional Well-Being through an External Movement-Based Environment: An Overview of Quadrato Motor Training" International Journal of Environmental Research and Public Health 16, no. 12: 2160. https://doi.org/10.3390/ijerph16122160
APA StyleDe Fano, A., Leshem, R., & Ben-Soussan, T. D. (2019). Creating an Internal Environment of Cognitive and Psycho-Emotional Well-Being through an External Movement-Based Environment: An Overview of Quadrato Motor Training. International Journal of Environmental Research and Public Health, 16(12), 2160. https://doi.org/10.3390/ijerph16122160