Campylobacteriosis, Salmonellosis, Yersiniosis, and Listeriosis as Zoonotic Foodborne Diseases: A Review


Abstract
:1. Introduction
2. Campylobacteriosis
3. Salmonellosis
4. Yersiniosis
5. Listeriosis
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2015. EFSA J. 2016, 14, e04634. [Google Scholar] [CrossRef]
- Karesh, W.B.; Dobson, A.; Lloyd-Smith, J.O.; Lubroth, J.; Dixon, M.A.; Bennett, M.; Aldrich, S.; Harrington, T.; Formenty, P.; Loh, E.H.; et al. Ecology of zoonoses: Natural and unnatural histories. Lancet 2012, 380, 1936–1945. [Google Scholar] [CrossRef]
- Zhang, H.L.; Mnzava, K.W.; Mitchell, S.T.; Melubo, M.L.; Kibona, T.J.; Cleaveland, S.; Kazwala, R.R.; Crump, J.A.; Sharp, J.P.; Halliday, J.E.B. Mixed Methods Survey of Zoonotic Disease Awareness and Practice among Animal and Human Healthcare Providers in Moshi, Tanzania. PLoS Negl. Trop. Dis. 2016, 10, e0004476. [Google Scholar] [CrossRef] [PubMed]
- Bonansea, S.J. Veterinary Hygiene: Applied to the Protection of Man against Zoonosis. Public Health Pap. Rep. 1906, 32, 320–324. [Google Scholar] [PubMed]
- The Zoonoses. Am. J. Public Health Nat. Health 1953, 43, 476–477.
- Levin, S. Zoonoses. In Goldman’s Cecil Medicine, 24th ed.; Goldman, L., Schafer, A.I., Eds.; W.B. Sounders: Philadelphia, PA, USA, 2012; Chapter 336; pp. 1964–1967. ISBN 9781455753369, 9780323295253, 9781437736083. [Google Scholar]
- Chikeka, I.; Dumler, J.S. Neglected Bacterial Zoonoses. Clin. Microbiol. Infect. 2015, 21, 404–415. [Google Scholar] [CrossRef] [PubMed]
- Cantas, L.; Suer, K. Review: The Important Bacterial Zoonoses in “One Health” Concept. Front. Public Health 2014, 2, 144. [Google Scholar] [CrossRef] [PubMed]
- Scanes, C.G. Animal and Human Disease: Zoonosis, Vectors, Food-Borne Diseases and Allergies. In Animals and Human Society; Scanes, C.G., Toukhsati, S.R., Eds.; Academic Press: London, UK; San Diego, CA, USA; Cambridge, UK; Oxford, UK, 2018; Chapter 14; pp. 331–348. ISBN 9780128054383. [Google Scholar]
- World Health Organization (WHO). WHO estimates of the global burden of foodborne diseases. In Foodborne Disease Burden Epidemiology Reference Group 2007–2015; World Health Organization: Geneva, Switzerland, 2015. [Google Scholar]
- Keerthirathne, T.P.; Ross, K.; Fallowfield, H.; Whiley, H. A Review of Temperature, pH, and Other Factors that Influence the Survival of Salmonella in Mayonnaise and Other Raw Egg Products. Pathogens 2016, 5, 63. [Google Scholar] [CrossRef] [PubMed]
- Rohde, A.; Hammerl, J.A.; Al Dahouk, S. Detection of foodborne bacterial zoonoses by fluorescence in situ hybridization. Food Control 2016, 69, 297–305. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2016. EFSA J. 2017, 15, e5077. [Google Scholar] [CrossRef]
- Zeng, D.; Chen, Z.; Jiang, Y.; Xue, F.; Li, B. Advances and Challenges in Viability Detection of Foodborne Pathogens. Front. Microbiol. 2016, 7, 1833. [Google Scholar] [CrossRef] [PubMed]
- McLinden, T.; Sargeant, J.M.; Thomas, M.K.; Papadopoulos, A.; Fazil, A. Component costs of foodborne illness: A scoping review. BMC Public Health 2014, 14, 509. [Google Scholar] [CrossRef] [PubMed]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The Community Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents, Antimicrobial Resistance and Foodborne Outbreaks in the European Union in 2005. EFSA J. 2006, 4, 94. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The Community Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents, Antimicrobial Resistance and Foodborne Outbreaks in the European Union in 2006. EFSA J. 2007, 5, 130. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The Community Summary Report on Trends and Sources of Zoonoses and Zoonotic Agents in the European Union in 2007. EFSA J. 2009, 7, 223. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The Community Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in the European Union in 2008. EFSA J. 2010, 8, 1496. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2009. EFSA J. 2011, 9, 2090. [Google Scholar] [CrossRef] [Green Version]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2010. EFSA J. 2012, 10, 2597. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2011. EFSA J. 2013, 11, 3129. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2012. EFSA J. 2014, 12, 3547. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union Summary Report on Trends and Sources of Zoonoses, Zoonotic Agents and Food-borne Outbreaks in 2013. EFSA J. 2015, 13, 3991. [Google Scholar] [CrossRef]
- European Food Safety Authority (EFSA); European Centre for Disease Prevention and Control (ECDC). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2014. EFSA J. 2015, 13, 4329. [Google Scholar] [CrossRef]
- Modi, S.; Brahmbhatt, M.N.; Chatur, Y.A.; Nayak, J.B. Prevalence of Campylobacter species in milk and milk products, their virulence gene profile and anti-bio gram. Vet. World 2015, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Kaakoush, N.O.; Castaño-Rodríguez, N.; Mitchell, H.M.; Man, S.M. Global Epidemiology of Campylobacter Infection. Clin. Microbiol. Rev. 2015, 28, 687–720. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.; Leite, D.; Fernandes, M.; Mena, C.; Gibbs, P.A.; Teixeira, P. Campylobacter spp. as a Foodborne Pathogen: A Review. Front. Microbiol. 2011, 2, 200. [Google Scholar] [CrossRef] [PubMed]
- Krutkiewicz, A. Kampylobakterioza u ludzi i zwierząt. Życie Weter. 2008, 83, 285–288. (In Polish) [Google Scholar]
- Bronnec, V.; Turoňová, H.; Bouju, A.; Cruveiller, S.; Rodrigues, R.; Demnerova, K.; Tresse, O.; Haddad, N.; Zagorec, M. Adhesion, Biofilm Formation, and Genomic Features of Campylobacter jejuni Bf, an Atypical Strain Able to Grow under Aerobic Conditions. Front. Microbiol. 2016, 7, 1002. [Google Scholar] [CrossRef] [PubMed]
- Wieczorek, K.; Osek, J. Antimicrobial Resistance Mechanisms among Campylobacter. BioMed Res. Int. 2013, 2013, 340605. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.J.; Shank, J.M.; Johnson, J.G. Current and Potential Treatments for Reducing Campylobacter Colonization in Animal Hosts and Disease in Humans. Front. Microbiol. 2017, 8, 487. [Google Scholar] [CrossRef] [PubMed]
- Galate, L.; Bangde, S. Campylobacter—A Foodborne Pathogen. Int. J. Sci. Res. 2015, 4, 1250–1259. [Google Scholar]
- Kashoma, I.P.; Kassem, I.I.; Kumar, A.; Kessy, B.M.; Gebreyes, W.; Kazwala, R.R.; Rajashekara, G. Antimicrobial Resistance and Genotypic Diversity of Campylobacter Isolated from Pigs, Dairy, and Beef Cattle in Tanzania. Front. Microbiol. 2015, 6, 1240. [Google Scholar] [CrossRef] [PubMed]
- Ravel, A.; Pintar, K.; Nesbitt, A.; Pollari, F. Non food-related risk factors of campylobacteriosis in Canada: A matched case-control study. BMC Public Health 2016, 16, 1016. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg Goldstein, R.E.; Cruz-Cano, R.; Jiang, C.; Palmer, A.; Blythe, D.; Ryan, P.; Hogan, B.; White, B.; Dunn, J.R.; Libby, T.; et al. Association between community socioeconomic factors, animal feeding operations, and campylobacteriosis incidence rates: Foodborne Diseases Active Surveillance Network (FoodNet), 2004–2010. BMC Infect. Dis. 2016, 16, 354. [Google Scholar] [CrossRef] [PubMed]
- Laprade, N.; Cloutier, M.; Lapen, D.R.; Topp, E.; Wilkes, G.; Villemur, R.; Khan, I.U.H. Detection of virulence, antibiotic resistance and toxin (VAT) genes in Campylobacter species using newly developed multiplex PCR assays. J. Microbiol. Methods 2016, 124, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Klančnik, A.; Vučković, D.; Jamnik, P.; Abram, M.; Možina, S.S. Stress Response and Virulence of Heat-Stressed Campylobacter jejuni. Microbes Environ. 2014, 29, 338–345. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.L.; Williams, D.W. Campylobacter. In Microbiology of Waterborne Diseases, 2nd ed.; Percival, S., Yates, M., Williams, D., Chalmers, R., Gray, N., Eds.; Academic Press: London, UK, 2014; Chapter 4; pp. 65–78. ISBN 978-0-12-415846-7. [Google Scholar]
- Bolton, D.J. Campylobacter virulence and survival factors. Food Microbiol. 2016, 48, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Epps, S.V.R.; Harvey, R.B.; Hume, M.E.; Phillips, T.D.; Anderson, R.C.; Nisbet, D.J. Foodborne Campylobacter: Infections, Metabolism, Pathogenesis and Reservoirs. Int. J. Environ. Res. Public Health 2013, 10, 6292–6304. [Google Scholar] [CrossRef] [PubMed]
- Kanji, A.; Jones, M.A.; Maskell, D.J.; Grant, A.J. Campylobacter jejuni PflB is required for motility and colonisation of the chicken gastrointestinal tract. Microb. Pathog. 2015, 89, 93–99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koolman, L.; Whyte, P.; Burgess, C.; Bolton, D. Virulence gene expression, adhesion and invasion of Campylobacter jejuni exposed to oxidative stress (H2O2). Int. J. Food. Microbiol. 2016, 220, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Andrzejewska, M.; Szczepańska, B.; Śpica, D.; Klawe, J.K. Trends in the occurrence and characteristics of Campylobacter jejuni and Campylobacter coli isolates from poultry meat in Northern Poland. Food Control 2015, 51, 190–194. [Google Scholar] [CrossRef]
- Iovine, N.M. Resistance mechanisms in Campylobacter jejuni. Virulence 2013, 4, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Skarp, C.P.A.; Hännien, M.L.; Rautelin, H.I.K. Campylobacteriosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Szczepańska, B.; Andrzejewska, M.; Śpica, D.; Klawe, J. Campylobacter spp.—Niedoceniany w Polsce czynnik etiologiczny zakażeń przewodu pokarmowego. Probl. Hig. Epidemio. 2014, 95, 574–579. (In Polish) [Google Scholar]
- Kawai, F.; Paek, S.; Choi, K.-J.; Prouty, M.; Kanipes, M.I.; Guerry, P.; Yeo, H.-J. Crystal structure of JlpA, a surface-exposed lipoprotein adhesin of Campylobacter jejuni. J. Struct. Biol. 2012, 177, 583–588. [Google Scholar] [CrossRef] [PubMed]
- Rokosz-Chudziak, N.; Rastawicki, W. Wybrane mechanizmy chorobotwórczości pałeczek Campylobacter jejuni. Med. Dosw. Mikrobiol. 2014, 66, 47–58. (In Polish) [Google Scholar] [PubMed]
- Black, R.E.; Levine, M.M.; Clements, M.L.; Hughes, T.P.; Blaser, M.J. Experimental Campylobacter jejuni Infection In Humans. J. Infect. Dis. 1988, 157, 472–479. [Google Scholar] [CrossRef] [PubMed]
- Schielke, A.; Rosner, B.M.; Stark, K. Epidemiology of campylobacteriosis in Germany—Insights from 10 years of surveillance. BMC Infect. Dis. 2014, 14, 30. [Google Scholar] [CrossRef] [PubMed]
- Di Giannatale, E.; Di Serafino, G.; Zilli, K.; Alessiani, A.; Sacchini, L.; Garofolo, L.; Aprea, G.; Marotta, F. Characterization of Antimicrobial Resisance Patterns and Detection of Virulence Genes in Campylobacter Isolates in Italy. Sensors 2014, 14, 3308–3322. [Google Scholar] [CrossRef] [PubMed]
- Nyati, K.K.; Nyati, R. Role of Campylobacter jejuni Infection in the Pathogenesis of Guillain-Barré Syndrome: An Update. BioMed Res. Int. 2013, 2013, 852195. [Google Scholar] [CrossRef] [PubMed]
- Hald, B.; Skov, M.N.; Nielsen, E.M.; Rahbek, C.; Madsen, J.J.; Wainø, M.; Chriél, M.; Nordentoft, S.; Baggesen, D.L.; Madsen, M. Campylobacter jejuni and Campylobacter coli in wild birds on Danish livestock farms. Acta Vet. Scand. 2016, 58, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Josefsen, M.H.; Bhunia, A.K.; Olsson Engvall, E.; Fachmann, M.S.R.; Hoorfar, J. Monitoring in the poultry production chain—From culture to genes and beyond. J. Microbiol. Methods 2015, 112, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Newell, D.G.; Mughini-Gras, L.; Kalupahana, R.; Wagenaar, J.A. Campylobacter Epidemiology—Sources and Routes of Transmission for Human Infection. In Campylobacter: Features, Detection, and Prevention of Foodborne Disease; Klein, G., Ed.; Academic Press: London, UK; San Diego, CA, USA, 2016; pp. 85–110. ISBN 9780128036495. [Google Scholar]
- Lin, J. Novel Approaches for Campylobacter Control in Poultry. Foodborne Pathog. Dis. 2009, 6, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Saint-Cyr, M.J.; Guyard-Nicodème, M.; Messaoudi, S.; Chemaly, M.; Cappelier, J.-M.; Dousset, X.; Haddad, N. Recent Advances in Screening of Anti-Campylobacter Activity in Probiotics for Use in Poultry. Front. Microbiol. 2016, 7, 553. [Google Scholar] [CrossRef] [PubMed]
- Whiley, H.; van den Akker, B.; Giglio, S.; Bentham, R. The Role of Environmental Reservoirs in Human Campylobacteriosis. Int. J. Environ. Res. Public. Health 2013, 10, 5886–5907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krueger, N.A.; Anderson, R.C.; Krueger, W.K.; Horne, W.J.; Wesley, I.V.; Callaway, T.R.; Edrington, T.S.; Carstens, G.E.; Harvey, R.B.; Nisbet, D.J. Prevalence and Concentration of Campylobacter in Rumen Contents and Feces in Pasture and Feedlot-Fed Cattle. Foodborne Pathog. Dis. 2008, 5, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Lazou, T.; Houf, K.; Soultos, N.; Dovas, C.; Iossifidou, E. Campylobacter in small ruminants at slaughter: Prevalence, pulsotypesand antibiotic resistance. Int. J. Food Microbiol. 2014, 173, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Qin, S.-S.; Wu, C.-M.; Jeon, B.; Shen, Z.-Q.; Wang, Y.; Zhnag, Q.; Shen, J.-Z. Antimicrobial resistance in Campylobacter coli isolated from pigs in two provinces of China. Int. J. Food Microbiol. 2011, 146, 94–98. [Google Scholar] [CrossRef] [PubMed]
- Egger, R.; Korczak, B.M.; Niederer, L.; Overesh, G.; Kuhnert, P. Genotypes and antibiotic resistance of Campylobacter coli in fattening pigs. Vet. Microbiol. 2012, 155, 272–278. [Google Scholar] [CrossRef] [PubMed]
- Denis, M.; Henrique, E.; Chidaine, B.; Tricot, A.; Bougeard, S.; Fravalo, P. Campylobacter from sows in farrow-to-finish pig farms: Risk indicators and genetic diversity. Vet. Microbiol. 2011, 154, 163–170. [Google Scholar] [CrossRef] [PubMed]
- Von Altrock, A.; Hamedy, A.; Merle, R.; Waldmann, K.-H. Campylobacter spp.—Prevalence on pig livers and antimicrobial susceptibility. Prev. Vet. Med. 2013, 109, 152–157. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). The Global View of Campylobacteriosis: Report of Expert Consultation, Utrecht, Netherlands, 9 to 11 July 2012; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- Fàbrega, A.; Vila, J. Salmonella enterica Serovar Typhimurium Skills to Succeed in the Host: Virulence and Regulation. Clin. Microbiol. Rev. 2013, 26, 308–341. [Google Scholar] [CrossRef] [PubMed]
- Li, H.P.; Wang, H.; D’Aoust, J.Y.; Maurer, J. Food Microbiology: Fundamentals and Frontiers, 4th ed.; Salmonella species; Doyle, M.P., Buchanan, R., Eds.; ASM Press: Washington, DC, USA, 2013; pp. 225–261. ISBN 9781555818463, 9781628702255. [Google Scholar]
- Cianflone, N.F.C. Salmonellosis and the GI Tract: More than Just Peanut Butter. Curr. Gastroenterol. Rep. 2008, 10, 424–431. [Google Scholar] [CrossRef]
- McSorley, S.J. Immunity to intestinal pathogens: Lessons learned from Salmonella. Immunol. Rev. 2014, 260, 168–182. [Google Scholar] [CrossRef] [PubMed]
- Khan, C.M.A. The Dynamic Interactions between Salmonella and the Microbiota, within the Challenging Niche of the Gastrointestinal Tract. Int. Sch. Res. Not. 2014, 2014, 846049. [Google Scholar] [CrossRef] [PubMed]
- Rönnqvist, M.; Valttila, V.; Ranta, J.; Tuominen, P. Salmonella risk to consumers via pork is related to the Salmonella prevalence in pig feed. Food Microbiol. 2018, 71, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Andino, A.; Hanning, I. Salmonella enterica: Survival, Colonization, and Virulence Differences among Serovars. Sci. World J. 2015, 2015, 520179. [Google Scholar] [CrossRef] [PubMed]
- Razzuoli, E.; Amadori, M.; Lazzara, F.; Bilato, D.; Ferraris, M.; Vito, G.; Ferrari, A. Salmonella serovar-specific interaction with jejunal epithelial cells. Vet. Microbiol. 2017, 207, 219–225. [Google Scholar] [CrossRef] [PubMed]
- Brenner, F.W.; Villar, R.G.; Angulo, F.J.; Tauxe, R.; Swaminathan, B. Salmonella Nomenclature. J. Clin. Microbiol. 2000, 38, 2465–2467. [Google Scholar] [PubMed]
- Agbaje, M.; Begum, R.H.; Oyekunle, M.A.; Ojo, O.E.; Adenubi, O.T. Evolution of Salmonella nomenclature: A critical note. Folia Microbiol. 2011, 56, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-M.; Wang, Y.; Su, L.-H.; Chiu, C.-H. Nontyphoid Infection: Microbiology, Clinical Features, and Antimicrobial Therapy. Pediatr. Neonatol. 2013, 54, 147–152. [Google Scholar] [CrossRef] [PubMed]
- Foley, S.L.; Johnson, T.J.; Ricke, S.C.; Nayak, R.; Danzeisen, J. Salmonella Pathogenicity and Host Adaptation in Chicken-Associated Serovars. Microbiol. Mol. Biol. Rev. 2013, 77, 582–607. [Google Scholar] [CrossRef] [PubMed]
- El-Tayeb, M.; Ibrahim, A.S.S.; Al-Salamah, A.A.; Almaary, K.S.; Elbadawi, Y.B. Prevalence, serotyping and antimicrobials resistance mechanism of Salmonella enterica isolated from clinical and environmental samples in Saudi Arabia. Braz. J. Microbiol. 2017, 48, 499–508. [Google Scholar] [CrossRef] [PubMed]
- Evangelopoulou, G.; Kritas, S.; Christodoulopoulos, G.; Burriel, A.R. The commercial impact of pig Salmonella spp. infections in border-free markets during an economic recession. Vet. World 2015, 8, 257–272. [Google Scholar] [CrossRef] [PubMed]
- Bugarel, M.; Tudor, A.; Loneragan, G.H.; Nightingale, K.K. Molecular detection assay of five Salmonella serotypes of public interest: Typhimurium, Enteritidis, Newport, Heidelberg, and Hadar. J. Microbiol. Methods 2017, 134, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Kurtz, J.R.; Goggins, J.A.; McLachlan, J.B. Salmonella infection: Interplay between the bacteria and host immune system. Immunol. Lett. 2017, 190, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Costa, L.F.; Paixão, T.A.; Tsolis, R.; Bäumler, A.J.; Santos, R. Salmonellosis in cattle: Advantages of being an experimental model. Res. Vet. Sci. 2012, 93, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wiedemann, A.; Virlogeux-Payant, I.; Chaussé, A.-M.; Schikora, A.; Velge, P. Interactions of Salmonella with animals and plants. Front. Microbiol. 2015, 5, 791. [Google Scholar] [CrossRef] [PubMed]
- Hung, Y.-T.; Lay, C.-J.; Wang, C.-L.; Koo, M. Characteristics of nontyphoidal gastroenteritis in Taiwanese children: A 9-year period retrospective medical record review. J. Infect. Public Health 2017, 10, 518–521. [Google Scholar] [CrossRef] [PubMed]
- Bierschenk, D.; Boucher, D.; Schroder, K. Salmonella-induced inflammasome activation in humans. Mol. Immunol. 2017, 86, 38–43. [Google Scholar] [CrossRef] [PubMed]
- Whiley, H.; Ross, K. Salmonella and Eggs: From Production to Plate. Int. J. Environ. Res. Public Health 2015, 12, 2543–2556. [Google Scholar] [CrossRef] [PubMed]
- De Jong, H.K.; Parry, C.M.; van der Poll, T.; Wiersinga, W.J. Host–Pathogen Interaction in Invasive Salmonellosis. PLoS Pathog. 2012, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Anderson, T.C.; Nguyen, T.-A.; Adams, J.K.; Garrett, N.M.; Bopp, C.A.; Baker, J.B.; McNeil, C.; Torres, P.; Ettestad, P.J.; Erdmna, M.N.; et al. Multistate outbreak of human Salmonella Typhimurium infections linked to live poultry from agricultural feed stores and mail-order hatcheries, United States 2013. One Health 2016, 2, 144–149. [Google Scholar] [CrossRef] [PubMed]
- Jiang, C.; Shaw, K.S.; Upperman, C.R.; Blythe, D.; Mitchell, C.; Murtugudde, R.; Sapkota, A.R.; Sapkota, A. Climate change, extreme events and increased risk of salmonellosis in Maryland, USA: Evidence for coastal vulnerability. Environ. Int. 2015, 83, 58–62. [Google Scholar] [CrossRef] [PubMed]
- Chaudhary, J.H.; Nayak, J.B.; Brahmbhatt, M.N.; Makwana, P.P. Virulence genes detection of Salmonella serovars isolated from pork and slaughter house environment in Ahmedabad, Gujarat. Vet. World 2015, 8, 121–124. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, A.; Lahiri, A.; Iyer, N.; Das, P.; Chakravortty, D. Visiting the cell biology of Salmonella infection. Microbes Infect. 2010, 12, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, R.; Card, R.; Nunes, C.; AbuOun, M.; Bagnall, M.C.; Nunez, J.; Mendonça, N.; Anjum, M.F.; da Silva, G.J. Virulence Characterization of Salmonella enterica by a New Microarray: Detection and Evaluation of the Cytolethal Distending Toxin Gene Activity in the Unusual Host S. Typhimurium. PLoS ONE 2015, 10, e0135010. [Google Scholar] [CrossRef] [PubMed]
- Ingram, J.P.; Brodsky, I.E.; Balachandran, S. Interferon-γ in Salmonella pathogenesis: New tricks for an old dog. Cytokine 2017, 98, 27–32. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Morales, F. Impact of Salmonella enterica Type III Secretion System Effectors on the Eukaryotic Host Cell. ISRN Cell Biol. 2012, 2012, 787934. [Google Scholar] [CrossRef]
- Schleker, S.; Sun, J.; Raghavan, B.; Srnec, M.; Müller, N.; Koepfinger, M.; Murthy, L.; Zhao, Z.; Klein-Seetharaman, J. The current Salmonella–host interactome. Proteom. Clin. Appl. 2012, 6, 117–133. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Kamanova, J.; Lara-Tejero, M.; Galán, J.E. A Family of Salmonella Type III Secretion Effector Proteins Selectively Targets the NF-κB Signaling Pathway to Preserve Host Homeostasis. PLoS Pathog. 2016, 12, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Chessa, D.; Spiga, L.; De Riu, N.; Delaconi, P.; Mazzarello, V.; Ganau, G.; Rubino, S. Lipopolysaccharides Belonging to Different Salmonella Serovars Are Differentially Capable of Activating Toll-Like Receptor 4. Infect. Immun. 2014, 82, 4553–4562. [Google Scholar] [CrossRef] [PubMed]
- Giannella, R.A. Salmonella. In Medical Microbiology, 4th ed.; Baron, S., Ed.; University of Texas Medical Branch, Department of Microbiology: Galveston, TX, USA, 1996; Chapter 21. Available online: https://www.ncbi.nlm.nih.gov/books/NBK7627 (accessed on 23 November 2017).
- Kawasaki, K.; Teramoto, M.; Tatsui, R.; Amamoto, S. Lipid A 3’-O-deacylation by Salmonella outer membrane enzyme LpxR modulates the ability of lipid A to stimulate Toll-like receptor 4. Biochem. Biophys. Res. Commun. 2012, 428, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Nobre, T.M.; Martynowycz, M.W.; Andreev, K.; Kuzmenko, I.; Nikaido, H.; Gidalevitz, D. Modification of Salmonella Lipopolysaccharides Prevents the Outer Membrane Penetration of Novobiocin. Biophys. J. 2015, 109, 2537–2545. [Google Scholar] [CrossRef] [PubMed]
- Van Asten, A.J.A.M.; van Dijk, J.E. Distribution of “classic” virulence factors among Salmonella spp. FEMS Immunol. Med. Microbiol. 2005, 44, 251–259. [Google Scholar] [CrossRef] [PubMed]
- Snider, T.A.; Gull, T.; Jackson, T.A.; Martinez-Becerra, F.J.; Picking, D.R.; Picking, W.D.; Picking, W.L. Experimental salmonellosis challenge model in older calves. Vet. Microbiol. 2014, 170, 65–72. [Google Scholar] [CrossRef] [PubMed]
- Kanj, S.S.; Kanafani, Z.A.; Shehab, M.; Sidani, N.; Baban, T.; Baltajian, K.; Dakdouki, G.K.; Zaatari, M.; Araj, G.F.; Wakim, R.H.; et al. Epidemiology, clinical manifestations, and molecular typing of Salmonella typhi isolated from patients with typhoid fever in Lebanon. J. Epidemiol. Glob. Health 2015, 5, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Pham, O.H.; McSorley, S.J. Protective host immune responses to Salmonella infection. Future Microbiol. 2015, 10, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Shukla, J.B.; Goyal, A.; Singh, S.; Chandra, P. Effects of habitat characteristics on the growth of carrier population leading to increased spread of typhoid fever: A model. J. Epidemiol. Glob. Health 2014, 4, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Antillón, M.; Warren, J.L.; Crawford, F.W.; Weinberger, D.M.; Kürüm, E.; Pak, G.D.; Pak, G.D.; Marks, F.; Pitzer, V.E. The burden of typhoid fever in low- and middle-income countries: A meta-regression approach. PLoS Negl. Trop. Dis. 2017, 11, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Antunes, P.; Mourão, J.; Campos, J.; Peixe, L. Salmonellosis: The role of poultry meat. Clin. Microbiol. Infect. 2016, 22, 110–121. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, N.A.; O’Bryan, C.A.; Dawuod, T.M.; Hong Park, S.; Min Kwon, Y.; Crandall, P.G.; Ricke, S.C. An overview of Salmonella thermal destruction during food processing and preparation. Food Control 2016, 68, 280–290. [Google Scholar] [CrossRef]
- Hald, T. Pathogen Updates: Salmonella. In Foodborne Infections and Intoxications, 4th ed.; Morris, G.J., Potter, M., Eds.; Academic Press: San Diego, CA, USA, 2013; pp. 67–97. ISBN 978-0-12-416041-5. [Google Scholar]
- Octavia, S.; Wang, Q.; Tanaka, M.M.; Sintchenko, V.; Lan, R. Genomic Variability of Serial Human Isolates of Salmonella enterica Serovar Typhimurium Associated with Prolonged Carriage. J. Clin. Microbiol. 2015, 53, 3507–3514. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Checkley, S.; Avery, B.; Chalmers, G.; Bohaychuk, V.; Gensler, G.; Reid-Smith, R.; Boerlin, P. Phenotypic and genetic characterization of antimicrobial resistance in Salmonella serovars isolated from retail meats in Alberta, Canada. Food Microbiol. 2012, 32, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Hurtado, A.; Ocejo, M.; Oporto, B. Salmonella spp. and Listeria monocytogenes shedding in domestic ruminants and characterization of potentially pathogenic strains. Vet. Microbiol. 2017, 210, 71–76. [Google Scholar] [CrossRef] [PubMed]
- Park, S.H.; Aydin, M.; Khatiwara, A.; Dolan, M.C.; Gilmore, D.F.; Bouldin, J.L.; Ahn, S.; Ricke, S.C. Current and emerging technologies for rapid detection and characterization of Salmonella in poultry and poultry products. Food Microbiol. 2014, 38, 250–262. [Google Scholar] [CrossRef] [PubMed]
- Pesciaroli, M.; Cucco, L.; De Luca, S.; Massacci, F.R.; Maresca, C.; Medici, L.; Paniccià, M.; Scoccia, E.; Staffolani, M.; Pezzotti, G.; et al. Association between pigs with high caecal Salmonella loads and carcass contamination. Int. J. Food Microbiol. 2017, 242, 82–86. [Google Scholar] [CrossRef] [PubMed]
- Sivaramalingam, T.; McEwen, S.A.; Pearl, D.L.; Ojkic, D.; Guerin, M.T. A temporal study of Salmonella serovars from environmental samples from poultry breeder flocks in Ontario between 1998 and 2008. Can. J. Vet. Res. 2013, 77, 1–11. [Google Scholar] [PubMed]
- Yeh, Y.; Puruschothaman, P.; Gupta, N.; Ragnone, M.; Verma, S.C.; de Mello, A.S. Bacteriophage application on red meats and poultry: Effects on Salmonella population in final ground products. Meat Sci. 2017, 127, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Mani-Lòpez, E.; García, H.S.; Lòpez-Malo, A. Organic acids as antimicrobials to control Salmonella in meat and poultry products. Food Res. Int. 2012, 45, 173–721. [Google Scholar] [CrossRef]
- Kore, K.; Asrade, B.; Demissie, K.; Aragaw, K. Characterization of Salmonella isolated from apparently healthy slaughtered cattle and retail beef in Hawassa, southern Ethiopia. Prev. Vet. Med. 2017, 147, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Dunkley, K.D.; Callaway, T.R.; Chalova, V.I.; McReynolds, J.L.; Hume, M.E.; Dunkley, C.S.; Kubena, L.F.; Nisbet, D.J.; Ricke, S.C. Foodborne Salmonella ecology in the avian gastrointestinal tract. Anaerobe 2009, 15, 26–35. [Google Scholar] [CrossRef] [PubMed]
- Nidaullah, H.; Abirami, N.; Shamila-Syuhada, A.K.; Chuah, L.-O.; Nurul, H.; Tan, T.P.; Abidin, F.W.Z.; Rusul, G. Prevalence of Salmonella in poultry processing environments in wet markets in Penang and Perlis, Malaysia. Vet. World 2017, 10, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Yadav, A.S.; Tripathi, V.; Singh, R.P. Antimicrobial resistance profile of Salmonella present in poultry and poultry environment in north India. Food Control 2013, 33, 545–548. [Google Scholar] [CrossRef]
- Barua, H.; Biswas, P.K.; Olsen, K.E.P.; Shil, S.K.; Christensen, J.P. Molecular Characterization of Motile Serovars of Salmonella enterica from Breeder and Commercial Broiler Poultry Farms in Bangladesh. PLoS ONE 2013, 8, e57811. [Google Scholar] [CrossRef] [PubMed]
- Saravanan, S.; Purushothaman, V.; Murthy, T.R.G.K.; Sukumar, K.; Srinivasan, P.; Gowthaman, V.; Balusamy, M.; Atterbury, R.; Kuchipudi, S.V. Molecular Epidemiology of Nontyphoidal Salmonella in Poultry and Poultry Products in India: Implications for Human Health. Indian J. Microbiol. 2015, 55, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Mir, I.A.; Kashyap, K.; Maherchandani, S. Isolation, serotype diversity and antibiogram of Salmonella enterica isolated from different species of poultry in India. Asian Pac. J. Trop. Biomed. 2015, 5, 561–567. [Google Scholar] [CrossRef]
- Kallapura, G.; Morgan, M.J.; Pumford, N.R.; Bielke, L.R.; Wolfenden, A.D.; Faulkner, O.B.; Latorre, J.D.; Menconi, A.; Hernandez-Velasco, X.; Kuttappan, V.A.; et al. Evaluation of the respiratory route as a viable portal of entry for Salmonella in poultry via intratracheal challenge of Salmonella Enteritidis and Salmonella Typhimurium. Poult. Sci. 2014, 93, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Rajtak, U.; Boland, F.; Leonard, N.; Bolton, D.; Fanning, S. Roles of Diet and the Acid Tolerance Response in Survival of Common Salmonella Serotypes in Feces of Finishing Pigs. Appl. Environ. Microbiol. 2012, 78, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Verbrugghe, E.; Boyen, F.; Van Parys, A.; Van Deun, K.; Croubels, S.; Thompson, A.; Shearer, N.; Leyman, B.; Haesebrouck, F.; Pasmans, F. Stress induced Salmonella Typhimurium recrudescence in pigs coincides with cortisol induced increased intracellular proliferation in macrophages. Vet. Res. 2011, 42, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vidic, B.; Savic, S.; Prica, N.; Suvajdzic, L. Epizootiology and control measures for Salmonella in pigs. Procedia Food Sci. 2015, 5, 312–315. [Google Scholar] [CrossRef]
- Oueslati, W.; Rjeibi, M.R.; Mhadhbi, M.; Jbeli, M.; Zrelli, S.; Ettriqui, A. Prevalence, virulence and antibiotic susceptibility of Salmonella spp. strains, isolated from beef in Greater Tunis (Tunisia). Meat Sci. 2016, 119, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Hoelzer, K.; Moreno Switt, A.I.; Wiedmann, M. Animal contact as a source of human non-typhoidal salmonellosis. Vet. Res. 2011, 42, 34. [Google Scholar] [CrossRef] [PubMed]
- Cummings, K.J.; Warnick, L.D.; Alexander, K.A.; Cripps, C.J.; Gröhn, Y.T.; James, K.I.; McDonough, P.L.; Reed, K.E. The duration of fecal Salmonella shedding following clinical disease among dairy cattle in the northeastern USA. Prev. Vet. Med. 2009, 92, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Janowska, M.; Jędrzejewska, B.; Janowska, J. Jersinioza—Nowe wyzwanie współczesnej medycyny. Med. Og. Nauk. Zdr. 2012, 3, 257–260. (In Polish) [Google Scholar]
- Pechous, R.D.; Sivaraman, V.; Stasulli, N.M.; Goldman, W.E. Pneumonic Plague: The Darker Side of Yersinia pestis. Trends Microbiol. 2016, 24, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Rakin, A.; Garzetti, D.; Bouabe, H.; Sprague, L.D. Yersinia enterocolitica. In Molecular Medical Microbiology, 2nd ed.; Tang, Y.-W., Sussman, M., Liu, D., Poxton, I., Schwartzman, J., Eds.; Academic Press: London, UK; Waltham, MA, USA; San Diego, CA, USA, 2015; Chapter 73; pp. 1319–1344. [Google Scholar]
- Fàbrega, A.; Vila, J. Yersinia enterocolitica: Pathogenesis, virulence and antimicrobial resistance. Enferm. Infecc. Microbiol. Clin. 2012, 30, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Dekker, J.; Frank, K. Salmonella, Shigella, and Yersinia. Clin. Lab. Med. 2015, 35, 225–246. [Google Scholar] [CrossRef] [PubMed]
- Galindo, C.L.; Rosenzweig, J.A.; Kirtley, M.L.; Chopra, A.K. Pathogenesis of Y. enterocolitica and Y. pseudotuberculosis in Human Yersiniosis. J. Pathog. 2011, 2011, 182051. [Google Scholar] [CrossRef] [PubMed]
- Mielczarek, P.; Bagłaj, M. Jersinioza—Rzadko rozpoznawana choroba układu pokarmowego. Gastroenterol. Pol. 2004, 11, 69–74. (In Polish) [Google Scholar]
- Bari, M.L.; Hossain, M.A.; Isshiki, K.; Ukuku, D. Behavior of Yersinia enterocolitica in Foods. J. Pathog. 2011, 2011, 420732. [Google Scholar] [CrossRef] [PubMed]
- Percival, S.L.; Williams, D.W. Yersinia. In Microbiology of Waterborne Diseases, 2nd ed.; Percival, S., Yates, M., Williams, D., Chalmers, R., Gray, N., Eds.; Academic Press: London, UK, 2014; Chapter 13; pp. 249–259. ISBN 978-0-12-415846-7. [Google Scholar]
- Söderqvist, K.; Boqvist, S.; Wauters, G.; Vågsholm, I.; Thisted-Lambertz, S. Yersinia enterocolitica in sheep—A high frequency of biotype 1A. Acta Vet. Scand. 2012, 54, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Bhagat, N.; Virdi, J.S. The Enigma of Yersinia enterocolitica biovar 1A. Crit. Rev. Microbiol. 2010, 37, 1–15. [Google Scholar] [CrossRef]
- Sihvonen, L.M.; Jalkanen, K.; Huovinen, E.; Toivonen, S.; Corander, J.; Kuusi, M.; Skurnik, M.; Siitonen, A.; Haukka, K. Clinical isolates of Yersinia enterocolitica Biotype 1A represent two phylogenetic lineages with differing pathogenicity-related properties. BMC Microbiol. 2012, 12, 208. [Google Scholar] [CrossRef] [PubMed]
- Garzetti, D.; Susen, R.; Fruth, A.; Tietze, E.; Heesemann, J.; Rakin, A. A molecular scheme for Yersinia enterocolitica patho-serotyping derived from genome-wide analysis. Int. J. Med. Microbiol. 2014, 304, 275–283. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Bonny, T.S.; Stonsaovapak, S.; Ananchaipattana, C. Yersinia enterocolitica: Epidemiological Studies and Outbreaks. J. Pathog. 2011, 2011, 239391. [Google Scholar] [CrossRef] [PubMed]
- Schaake, J.; Drees, A.; Grüning, P.; Uliczka, F.; Pisano, F.; Thiermann, T.; Von Altrock, A.; Seehusen, F.; Valentin-Weigand, P.; Dersch, P. Essential Role of Invasin for Colonization and Persistence of Yersinia enterocolitica in Its Natural Reservoir Host, the Pig. Infect. Immun. 2014, 82, 960–969. [Google Scholar] [CrossRef] [PubMed]
- Bancerz-Kisiel, A.; Szewda, W. Yersiniosis—A zoonotic foodborne disease of relevance to public health. Ann. Agric. Environ. Med. 2015, 22, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Fàbrega, A.; Ballesté-Delpierre, C.; Vila, J. Antimicrobial Resistance in Yersinia enterocolitica. In Antimicrobial Resistance and Food Safety; Chen, C.-Y., Yan, X., Jackson, C.R., Eds.; Academic Press: London, UK; San Diego, CA, USA; Waltham, MA, USA; Oxford, UK, 2015; Chapter 5; pp. 77–104. ISBN 978-0-12-801214-7. [Google Scholar]
- Atkinson, S.; Williams, P. Yersinia virulence factors—A sophisticated arsenal for combating host defences. F1000Res 2016, 5. [Google Scholar] [CrossRef] [PubMed]
- Bottone, E.J. Yersinia enterocolitica: Revisitation of an Enduring Human Pathogen. Clin. Microbiol. Newslett. 2015, 37, 1–8. [Google Scholar] [CrossRef]
- Chauhan, N.; Wrobel, A.; Skurnik, M.; Leo, J.C. Yersinia adhesins: An arsenal for infection. Proteom. Clin. Appl. 2016, 10, 949–963. [Google Scholar] [CrossRef] [PubMed]
- Dhar, M.S.; Virdi, J.S. Strategies used by Yersinia enterocolitica to evade killing by the host: Thinking beyond Yops. Microbes Infect. 2014, 16, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Gierczyński, R. Filogeneza, chorobotwórczość i zróżnicowanie pałeczek Yersinia enterocolitica. Med. Dosw. Mikrobiol. 2012, 64, 159–181. (In Polish) [Google Scholar] [PubMed]
- Heroven, A.K.; Dersch, P. Coregulation of host-adapted metabolism and virulence by pathogenic Yersiniae. Front. Cell. Infect. Microbiol. 2014, 4, 146. [Google Scholar] [CrossRef] [PubMed]
- Lawrenz, M.B.; Lenz, J.D.; Miller, V.L. A Novel Autotransporter Adhesin Is Required for Efficient Colonization during Bubonic Plague. Infect. Immun. 2009, 77, 317–326. [Google Scholar] [CrossRef] [PubMed]
- Diepold, A.; Amstutz, M.; Abel, S.; Sorg, I.; Jenal, U.; Cornelis, G.R. Deciphering the assembly of the Yersinia type III secretion injectisome. EMBO J. 2010, 29, 1928–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dewoody, R.S.; Merritt, P.M.; Marketon, M.M. Regulation of the Yersinia type III secretion system: Traffic control. Front. Cell. Infect. Microbiol. 2013, 3, 4. [Google Scholar] [CrossRef] [PubMed]
- Kopaskie, K.S.; Ligtenberg, K.G.; Schneewind, O. Translational Regulation of Yersinia enterocolitica mRNA Encoding a Type III Secretion Substrate. J. Biol. Chem. 2013, 288, 35478–35488. [Google Scholar] [CrossRef] [PubMed]
- Uliczka, F.; Pisano, F.; Schaake, J.; Stolz, T.; Rohde, M.; Fruth, A.; Strauch, E.; Skurnik, M.; Batzilla, J.; Rakin, A.; et al. Unique Cell Adhesion and Invasion Properties of Yersinia enterocolitica O:3, the Most Frequent Cause of Human Yersiniosis. PLoS Pathog. 2011, 7, e1002117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imori, P.F.M.; Passaglia, J.; Souza, R.A.; Rocha, L.B.; Falcão, J.P. Virulence-related genes, adhesion and invasion of some Yersinia enterocolitica-like strains suggests its pathogenic potential. Microb. Pathog. 2017, 104, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Kechagia, N.; Nicolaou, C.; Ioannidou, V.; Kourti, E.; Ioannidis, A.; Legakis, N.J.; Chatzipanagiotou, S. Detection of chromosomal and plasmid—Encoded virulence determinants in Yersinia enterocolitica and other Yersinia spp. isolated from food animals in Greece. Int. J. Food Microbiol. 2007, 118, 326–331. [Google Scholar] [CrossRef] [PubMed]
- Schneeberger, M.; Brodard, I.; Overesch, G. Virulence-associated gene pattern of porcine and human Yersinia enterocolitica biotype 4 isolates. Int. J. Food. Microbiol. 2015, 198, 70–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielińska, D.; Ołdak, A.; Rzepkowska, A.; Kołożyn-Krajewska, D. Psychotrofy w chłodniczym przechowywaniu żywności. Przem. Spoż. 2015, 1, 18–22. [Google Scholar] [CrossRef]
- Chung, L.K.; Bliska, J.B. Yersinia versus host immunity: How a pathogen evades or triggers a protective response. Curr. Opin. Microbiol. 2016, 29, 56–62. [Google Scholar] [CrossRef] [PubMed]
- Rosner, B.M.; Stark, K.; Werber, D. Epidemiology of reported Yersinia enterocolitica infections in Germany, 2001–2008. BMC Public Health 2010, 10, 337. [Google Scholar] [CrossRef] [PubMed]
- Kasprzak, J.; Domańska, E.; Małkińska-Horodyska, M. Sytuacja epidemiologiczna jersiniozy (zakażenie jelitowe) w województwie kujawsko-pomorskim w latach 2011–2013. Probl. Hig. Epidemio. 2014, 95, 844–853. (In Polish) [Google Scholar]
- Frazão, M.R.; Andrade, L.N.; Darini, A.L.C.; Falcão, J.P. Antimicrobial resistance and plasmid replicons in Yersinia enterocolitica strains isolated in Brazil in 30 years. Braz. J. Infect. Dis. 2017, 21, 477–480. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Yersinia enterocolitica (Yersiniosis). 2016. Available online: https://www.cdc.gov/yersinia/ (accessed on 11 November 2017).
- Arrausi-Subiza, M.; Gerrikagoitia, X.; Alvarez, V.; Ibabe, J.C.; Barral, M. Prevalence of Yersinia enterocolitica and Yersinia pseudotuberculosis in wild boars in the Basque Country, northern Spain. Acta Vet. Scand. 2016, 58, 4. [Google Scholar] [CrossRef] [PubMed]
- Nesbakken, T. Surveillance and Control of Enteric Yersinioses. In Yersinia: System Biology and Control; Carniel, E., Hinnebusch, B.J., Eds.; Caister Academic Press: Norfolk, UK, 2012; pp. 217–238. ISBN 9781908230058. [Google Scholar]
- Bursová, Š.; Necidová, L.; Haruštiaková, D.; Janštová, B. Growth potential of Yersinia enterocolitica in pasteurized cow’s and goat’s milk stored at 8 °C and 24 °C. Food Control 2017, 73, 1415–1419. [Google Scholar] [CrossRef]
- Ye, Q.; Wu, Q.; Hu, H.; Zhang, J.; Huang, H. Prevalence and characterization of Yersinia enterocolitica isolated from retail foods in China. Food Control 2016, 61, 20–27. [Google Scholar] [CrossRef]
- Le Guern, A.-S.; Martin, L.; Savin, C.; Carniel, E. Yersiniosis in France: Overview and potential sources of infection. Int. J. Infect. Dis. 2016, 46, 1–7. [Google Scholar] [CrossRef]
- Tan, L.K.; Ooi, P.T.; Thong, K.L. Prevalence of Yersinia enterocolitica from food and pigs in selected states of Malaysia. Food Control 2014, 35, 94–100. [Google Scholar] [CrossRef]
- Van Damme, I.; De Zutter, L.; Jacxsens, L.; Nauta, M.J. Control of human pathogenic Yersinia enterocolitica in minced meat: Comparative analysis of different interventions using a risk assessment approach. Food Microbiol. 2017, 64, 93–95. [Google Scholar] [CrossRef] [PubMed]
- Donovan, S. Listeriosis: A Rare but Deadly Disease. Clin. Microbiol. Newslett. 2015, 37, 135–140. [Google Scholar] [CrossRef]
- Schäfer, D.F.; Steffens, J.; Barbosa, J.; Zeni, J.; Paroul, N.; Valduga, E.; Junges, A.; Backes, G.T.; Cansian, R.L. Monitoring of contamination sources of Listeria monocytogenes in a poultry slaughterhouse. LWT Food Sci. Technol. 2017, 86, 393–398. [Google Scholar] [CrossRef]
- De Noordhout, C.M.; Devleesschauwer, B.; Angulo, F.J.; Verbeke, G.; Haagsma, J.; Kirk, M.; Havelaar, A.; Speybroeck, N. The global burden of listeriosis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 1073–1082. [Google Scholar] [CrossRef]
- McNeill, C.; Sisson, W.; Jarrett, A. Listeriosis: A Resurfacing Menace. J. Nurse Pract. 2017, 13, 647–654. [Google Scholar] [CrossRef]
- Orsi, R.H.; Wiedmann, M. Characteristics and distribution of Listeria spp., including Listeria species newly described since 2009. Appl. Microbiol. Biotechnol. 2016, 100, 5273–5287. [Google Scholar] [CrossRef] [PubMed]
- Den Bakker, H.C.; Warchocki, S.; Wright, E.M.; Allred, A.F.; Ahlstorm, C.; Manuel, C.S.; Stasiewicz, M.J.; Burrell, A.; Roof, S.; Strawn, L.K.; et al. Listeria floridensis sp. nov., Listeria aquatic sp. nov., Listeria cornellensis sp. nov., Listeria riparia sp. nov. and Listeria grandensis sp. nov., from agricultural and natural environments. Int. J. Syst. Evol. Microbiol. 2014, 64, 1882–1889. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.; McLauchlin, J. Biology. In Handbook of Listeria monocytogenes; Liu, D., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 3–26. ISBN 9781420051407. [Google Scholar]
- Jurkiewicz, A.; Oleszczak-Momot, W. Listeria monocytogenes jako problem zdrowia publicznego. Med. Og. Nauk. Zdr. 2015, 21, 29–32. [Google Scholar] [CrossRef]
- Osimani, A.; Clementi, F. The occurrence of Listeria monocytogenes in mass catering: An overview in the European Union. Int. J. Hosp. Manag. 2016, 57, 9–17. [Google Scholar] [CrossRef]
- Muskalska, K.B.; Szymczak, B. Postęp badań na bakteriami rodzaju Listeria. Post. Mikrobiol. 2015, 54, 123–132. (In Polish) [Google Scholar]
- Lotfollahi, L.; Chaharbalesh, A.; Rezaee, M.A.; Hasani, A. Prevalence, antimicrobial susceptibility and multiplex PCR-serotyping of Listeria monocytogenes isolated from humans, foods and livestock in Iran. Microb. Pathog. 2017, 107, 425–429. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.X.Y.; Brooks, B.W.; Huang, H.; Pagotto, F.; Lin, M. Identification of Surface Protein Biomarkers of Listeria monocytogenes via Bioinformatics and Antibody-Based Protein Detection Tools. Appl. Environ. Microbiol. 2016, 82, 5465–5476. [Google Scholar] [CrossRef] [PubMed]
- Lechowicz, J.; Krawczyk-Balska, A. An update on the transport and metabolism of iron in Listeria monocytogenes: The role of proteins involved in pathogenicity. Biometals 2015, 28, 587–603. [Google Scholar] [CrossRef] [PubMed]
- Seveau, S. Multifaceted Activity of Listeriolysin O, the Cholesterol-Dependent Cytolysin of Listeria monocytogenes. In MACPF/CDC Proteins—Agents of Defence, Attack and Invasion. Subcellular Biochemistry; Anderluh, G., Gilbert, R., Eds.; Springer: Dordrecht, The Netherlands, 2014; pp. 161–195. ISBN 978-94-017-8881-6. [Google Scholar]
- Carvalho, F.; Sousa, S.; Cabanes, D. How Listeria monocytogenes organizes its surface for virulence. Front. Cell. Infect. Microbiol. 2014, 4, 48. [Google Scholar] [CrossRef] [PubMed]
- Prokop, A.; Gouin, E.; Villiers, V.; Nahori, M.-A.; Vincentelli, R.; Duval, M.; Cossart, P.; Dussurget, O. OrfX, a Nucleomodulin Required for Listeria monocytogenes Virulence. mBio 2017, 8, e01550-17. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Bhunia, A. Secreted Listeria adhesion protein (Lap) influences Lap-mediated Listeria monocytogenes paracellular translocation through epithelial barrier. Gut Pathog. 2013, 5, 16. [Google Scholar] [CrossRef] [PubMed]
- Koo, O.K.; Amalaradjou, M.A.R.; Bhunia, A.K. Recombinant Probiotic Expressing Listeria Adhesion Protein Attenuates Listeria monocytogenes Virulence In Vitro. PLoS ONE 2012, 7, e29277. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.; Akgul, A.; Attila, K.; Abdelhamed, H.; Wills, R.W.; Lawrence, M.L. The role of Listeria monocytogenes cell wall surface anchor protein LapB in virulence, adherence, and intracellular replication. Microb. Pathog. 2016, 92, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Camejo, A.; Buchrieser, C.; Couvé, E.; Carvalho, F.; Reis, O.; Ferreira, P.; Sousa, S.; Cossart, P.; Cabanes, D. In Vivo Transcriptional Profiling of Listeria monocytogenes and Mutagenesis Identify New Virulence Factors Involved in Infection. PLoS Pathog. 2009, 5, e1000449. [Google Scholar] [CrossRef] [PubMed]
- Gilmartin, N.; Gião, M.S.; Keevil, C.W.; O’Kennedy, R. Differential internalin A levels in biofilms of Listeria monocytogenes grown on different surfaces and nutrient conditions. Int. J. Microbiol. 2016, 219, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Neves, D.; Job, V.; Dortet, L.; Cossart, P.; Dessen, A. Structure of Internalin InlK from the Human Pathogen Listeria monocytogenes. J. Mol. Biol. 2013, 425, 4520–4529. [Google Scholar] [CrossRef] [PubMed]
- Popowska, M.; Krawczyk-Balska, A.; Ostrowski, R.; Desvaux, M. InlL from Listeria monocytogenes Is Involved in Biofilm Formation and Adhesion to Mucin. Front. Microbiol. 2017, 8, 660. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cao, G.; Xu, X.; Allard, M.; Li, P.; Brown, E.; Yang, X.; Pan, H.; Meng, J. Evolution and Diversity of Listeria monocytogenes from Clinical and Food Samples in Shanghai, China. Front. Microbiol. 2016, 7, 1138. [Google Scholar] [CrossRef] [PubMed]
- Hadjilouka, A.; Molfeta, C.; Panagiotopoulou, O.; Paramithiotis, S.; Mataragas, M.; Drosinos, E.H. Expression of Listeria monocytogenes key virulence genes during growth in liquid medium, on rocket and melon at 4, 10 and 30 °C. Food Microbiol. 2016, 55, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Hamon, M.A.; Ribet, D.; Stavru, F.; Cossart, P. Listeriolysin O: The Swiss army knife of Listeria. Trends Microbiol. 2012, 20, 360–368. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Teng, Z.; Wang, J.; Lu, G.; Deng, X.; Li, L. Inhibition of listeriolysin O oligomerization by lutein prevents Listeria monocytogenes infection. Fitoterapia 2017, 116, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Tattoli, I.; Sorbara, M.T.; Yang, C.; Tooze, S.A.; Philpott, D.J.; Girardin, S.E. Listeria phospholipases subvert host autophagic defenses by stalling pre-autophagosomal structures. EMBO J. 2013, 32, 3066–3078. [Google Scholar] [CrossRef] [PubMed]
- Travier, L.; Lecuit, M. Listeria monocytogenes ActA: A new funtion for “classic” virulence factor. Curr. Opin. Microbiol. 2014, 17, 53–60. [Google Scholar] [CrossRef] [PubMed]
- Travier, L.; Guadagnini, S.; Gouin, E.; Dufour, A.; Chenal-Francisque, V.; Cossart, P.; Olivo-Marin, J.-C.; Ghigo, J.-M.; Disson, O.; Lecuit, M. ActA Promotes Listeria monocytogenes Aggregation, Intestinal Colonization and Carriage. PLoS Pathog. 2013, 9, e1003131. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Q.; Gooneratne, R.; Hussain, M.A. Listeria monocytogenes in Fresh Produce: Outbreaks, Prevalence and Contamination Levels. Foods 2017, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Milian, A.; Payeras-Cifre, A. What Is New in Listeriosis? BioMed Res. Int. 2014, 2014, 358051. [Google Scholar] [CrossRef] [PubMed]
- Mateus, T.; Silva, J.; Maia, R.L.; Teixeira, P. Listeriosis during Pregnancy: A Public Health Concern. ISRN Obstet. Gynecol. 2013, 2013, 851712. [Google Scholar] [CrossRef] [PubMed]
- Gómez, D.; Azón, E.; Marco, N.; Carramiñana, J.J.; Rota, C.; Ariño, A.; Yangüela, J. Antimicrobial resistance of Listeria monocytogenes and Listeria innocua from meat products and meat-processing environment. Food Microbiol. 2014, 42, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Rothrock, M.J.; Davis, M.L.; Locatelli, A.; Bodie, A.; McIntosh, T.G.; Donaldson, J.R.; Ricke, S.C. Listeria Occurrence in Poultry Flocks: Detection and Potential Implications. Front. Vet. Sci. 2017, 4, 125. [Google Scholar] [CrossRef] [PubMed]
- Tahoun, A.B.M.B.; Abou Elez, R.M.M.; Abdelfatah, E.N.; Elsohaby, I.; El-Gedawy, A.A.; Elmoslemany, A.M. Listeria monocytogenes in raw milk, milking equipment and dairy workers: Molecular characterization and antimicrobial resistance patterns. J. Glob. Antimicrob. Resist. 2017, 10, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention. Listeria (Listeriosis): People at Risk. 2017. Available online: https://www.cdc.gov/listeria/risk.html (accessed on 9 December 2017).
- Food Standards Australia New Zealand. Listeria: Food Safety and Recalls. 2017. Available online: http://www.foodstandards.gov.au/consumer/safety/listeria/Pages/default.aspx (accessed on 9 December 2017).
- Stea, E.C.; Purdue, L.M.; Jamieson, R.C.; Yost, C.K.; Truelstrup Hansen, L. Comparison of the Prevalences and Diversities of Listeria Species and Listeria monocytogenes in an Urban and a Rural Agricultural Watershed. Appl. Environ. Microbiol. 2015, 81, 3812–3822. [Google Scholar] [CrossRef] [PubMed]
- McIntyre, L.; Wilcott, L.; Naus, M. Listeriosis Outbreaks in British Columbia, Canada, Caused by Soft Ripened Cheese Contaminated from Environmental Sources. BioMed Res. Int. 2015, 2015, 131623. [Google Scholar] [CrossRef] [PubMed]
- Gambarin, P.; Magnabosco, C.; Losio, M.N.; Pavoni, E.; Gattuso, A.; Arcangeli, G.; Favretti, M. Listeria monocytogenes in Ready-to-Eat Seafood and Potential Hazards for the Consumers. Int. J. Microbiol. 2012, 2012, 497635. [Google Scholar] [CrossRef] [PubMed]
- Wałecka-Zacharska, E.; Kosek-Paszkowska, K.; Bania, J.; Staroniewicz, Z.; Bednarski, M.; Wieliczko, A. Invasiveness of Listeria monocytogenes strains isolated from animals in Poland. Pol. J. Vet. Sci. 2015, 18, 697–702. [Google Scholar] [CrossRef] [PubMed]
- Dreyer, M.; Thomann, A.; Böttcher, S.; Frey, J.; Oevermann, A. Outbreak investigation identifies a single Listeria monocytogenes strain in sheep with different clinical manifestations, soil and water. Vet. Microbiol. 2015, 179, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Esteban, J.I.; Oporto, B.; Aduriz, G.; Juste, R.A.; Hurtado, A. Faecal shedding and strain diversity of Listeria monocytogenes in healthy ruminants and swine in Northern Spain. BMC Vet. Res. 2009, 5, 2. [Google Scholar] [CrossRef] [PubMed]
- Bundrant, B.N.; Hutchins, T.; den Bakker, H.C.; Fortes, E.; Wiedmann, M. Listeriosis outbreak in dairy cattle coused by an unusual Listeria monocytogenes serotype 4b strain. J. Vet. Diagn. Investig. 2011, 23, 155–158. [Google Scholar] [CrossRef] [PubMed]
- Oevermann, A.; Zurbriggen, A.; Vandevelde, M. Rhombencephalitis Caused by Listeria monocytogenes in Humans and Ruminants: A Zoonosis on the Rise? Interdiscip. Perspect. Infect. Dis. 2010, 2010, 632513. [Google Scholar] [CrossRef] [PubMed]
- Gliński, Z.; Kostro, K. Listerioza współczesnym zagrożeniem. Życie Weter. 2012, 87, 577–581. (In Polish) [Google Scholar]
- Smith, B.; Larsson, J.T.; Lisby, M.; Müller, L.; Madsen, S.B.; Engberg, J.; Bangsborg, J.; Ethelberg, S.; Kemp, M. Outbreak of listeriosis caused by infected beef meat from a meals-on-wheels delivery in Denmark 2009. Clin. Microbiol. Infect. 2011, 17, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Gebretsadik, S.; Kassa, T.; Alemayehu, H.; Huruy, K.; Kebede, N. Isolation and characterization of Listeria monocytogenes and other Listeria species in foods of animal origin in Addis Ababa, Ethiopia. J. Infect. Public Health 2011, 4, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.J. Bacterial Zoonoses: An Overview. In Molecular Medical Microbiology, 2nd ed.; Tang, Y.-W., Sussman, M., Liu, D., Poxton, I., Schwartzman, J., Eds.; Academic Press: Boston, MA, USA, 2015; Chapter 100; Volume 3, pp. 1771–1780. ISBN 978-0-12-397169-2. [Google Scholar]
- Layton, D.S.; Choudhary, A.; Bean, A.G.D. Breaking the chain of zoonoses through biosecurity in livestock. Vaccine 2017, 35, 5967–5973. [Google Scholar] [CrossRef] [PubMed]
- Angelos, J.; Arens, A.; Johnson, H.; Cadriel, J.; Osburn, B. One Health in food safety and security education: A curricular framework. Comp. Immunol. Microbiol. Infect. Dis. 2016, 44, 29–33. [Google Scholar] [CrossRef] [PubMed]

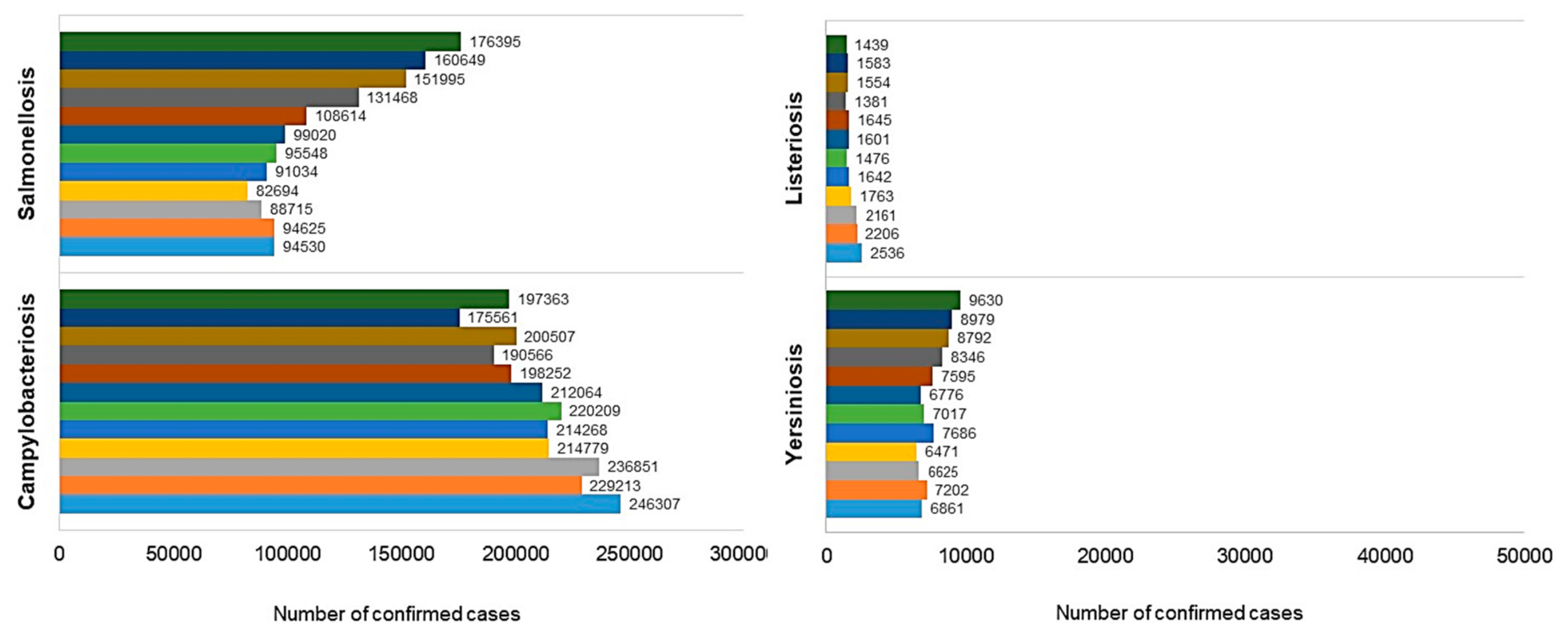


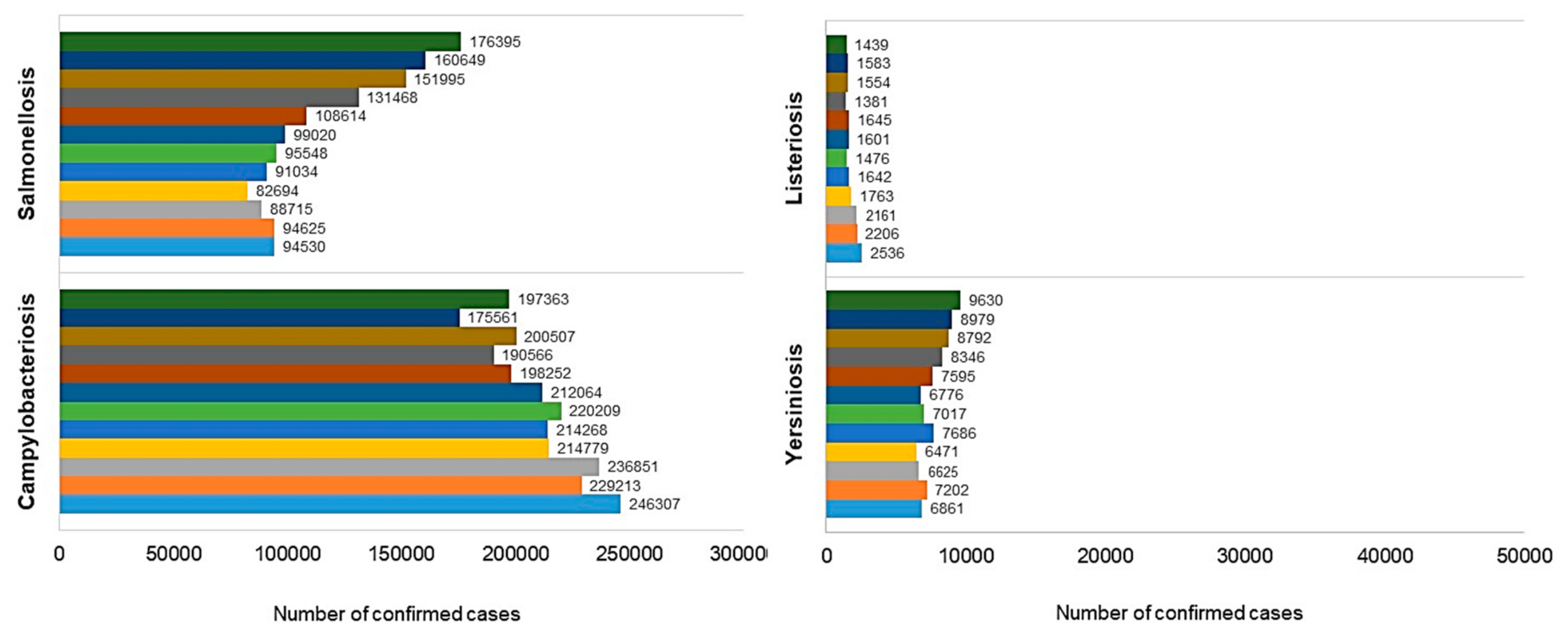
{kind=link}
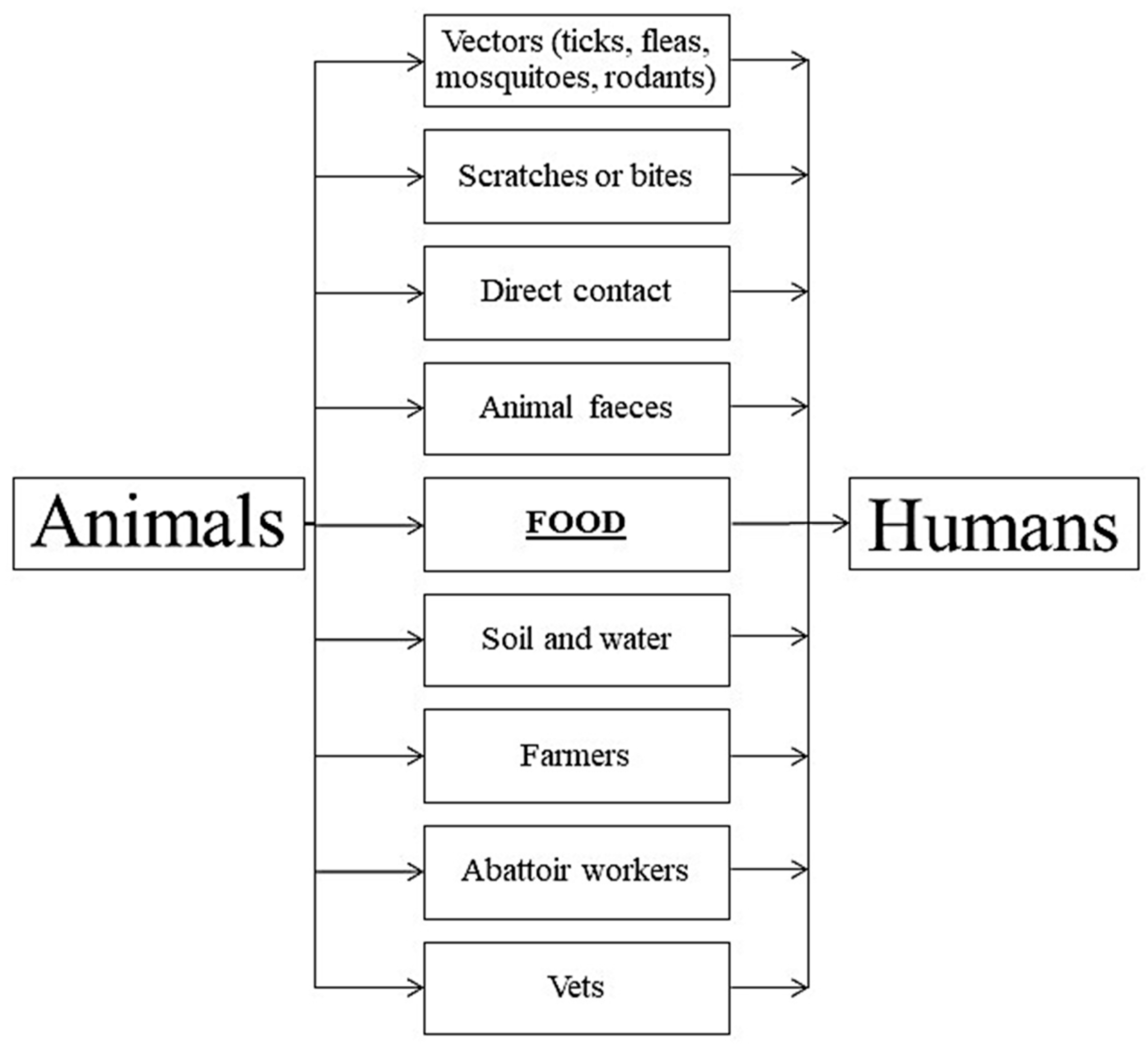
{kind=link}
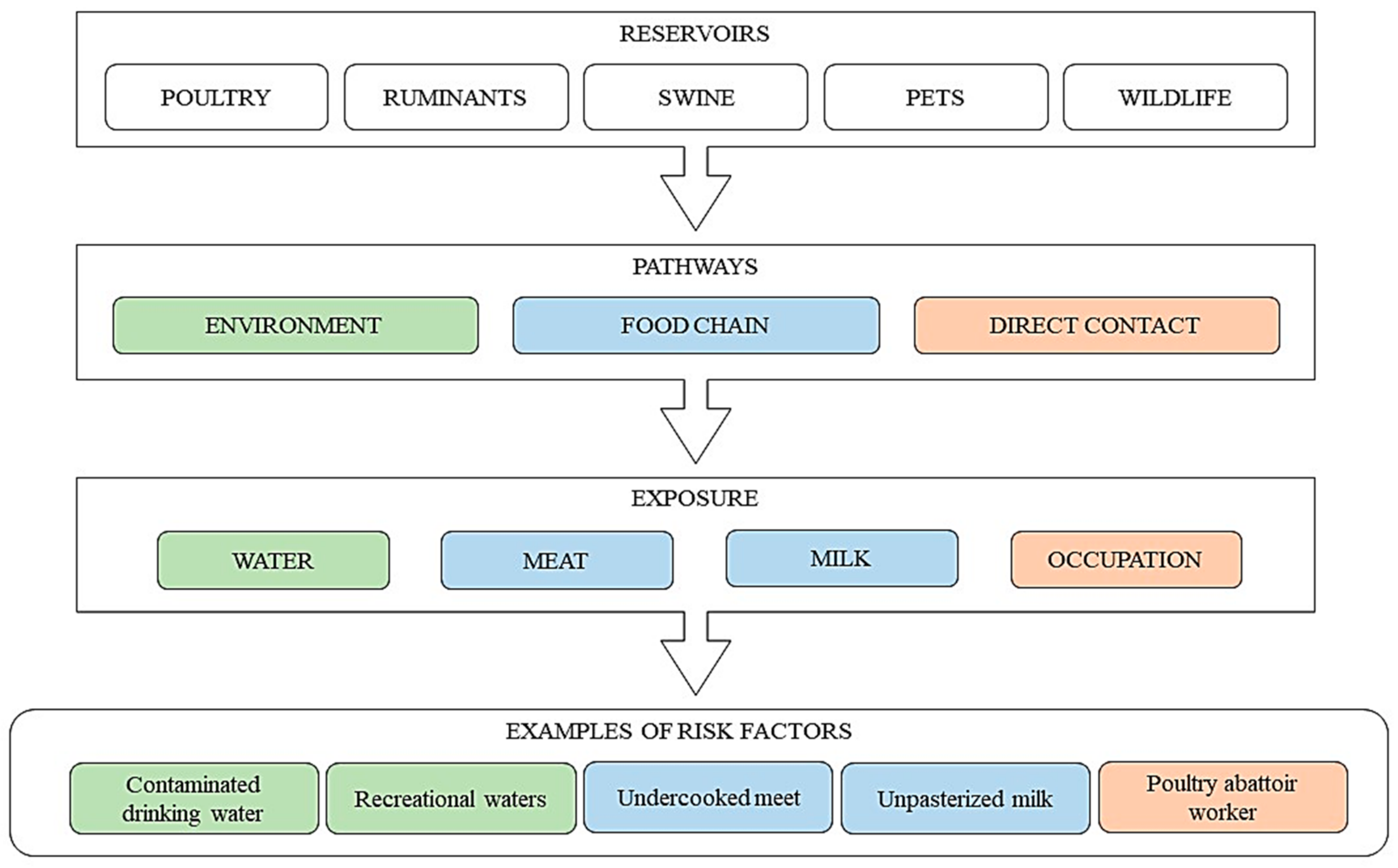{kind=link}
| The Mechanism of Survival/Virulence | Description | References |
|---|---|---|
| Mobility |
| [40,41,42,43] |
| Drug resistance |
| [27,40,44,45,46,47] |
| Adherence to host’s epithelial cells |
| [40,41,48] |
| Invasion of host’s cells |
| [39,41] |
| Production of toxins—cytolethal distending toxin (CDT) |
| [28,40,49] |
| The Mechanism of Virulence | Description | References |
|---|---|---|
| Adherence to host’s cells | Adherence modulated by:
| [78,84,93] |
| Invasion and replication inside host’s cells |
| [86,93,95,96,97] |
| Polysaccharide coating |
| [98,99,100,101,102] |
| Production of toxins |
| [99,102] |
| Year | Animal Reservoirs | |||
|---|---|---|---|---|
| Broiler | Cattle | Pigs | Turkey | |
| Salmonella Serotypes | ||||
| 2014 | S. Typhimurium S. Infantis S. Enteritidis S. Mbandaka S. Livingstone S. Kedougou S. Enftenberg S. Kentucky S. Typhimurium Copenhagen S. Brandenburg | S. Typhimurium S. Enteritidis S. Dublin S. Mbandaka S. Coeln S. Give S. Montevideo S. Anatum S. Bredeney S. Typhimurium Copenhagen | S. Typhimurium S. Infantis S. Derby S. Typhimurium monophasic S. Typhimurium Copenhagen S. Rissen S. London S. Muenchen S. Livingstone ar14 | S. Typhimurium S. Infantis S. Derby S. Enteritidis S. Newport S. Hadar S. Stanley S. Saintpaul S. Virchow S. Kottbus |
| 2015 | S. Typhimurium S. Infantis S. Enteritidis S. Derby S. Typhimurium monophasic S. Livingstone S. Mbandaka S. Cerro S. Thompson S. Kedougou | S. Typhimurium S. Infantis S. Enteritidis S. Dublin S. Typhimurium monophasic S. Mbandaka S. Newport S. Goldcoast S. Brandenburg | S. Typhimurium S. Infantis S. Enteritidis S. Derby S. Typhimurium monophasic S. Goldcoast S. Rissen S. Brandenburg S. London | S. Typhimurium S. Infantis S. Enteritids S. Derby S. Typhimurium monophasic S. Newport S. Kedougou S. Branderburg |
| 2016 | S. Enteritidis S. Typhimurium S. Typhimurium monophasic S. Infantis S. Derby | S. Enteritidis S. Typhimurium S. Typhimurium monophasic S. Infantis S. Derby | S. Enteritidis S. Typhimurium S. Typhimurium monophasic S. Infantis S. Derby | S. Enteritidis S. Typhimurium S. Typhimurium monophasic S. Infantis S. Derby |
| Virulence Mechanisms/Factors | Description | References |
|---|---|---|
| Adhesion and invasion of host’s cells | Adherence modulated by:
| [136,138,150,151,152,153,154,155,156] |
| Secretion system type III |
| [137,138,149,157,158,159] |
| Mobility |
| [136,160] |
| Lipopolysaccharides |
| [135,136] |
| Thermostable enterotoxin Yst |
| [135,136,144,161,162,163,164] |
| Production of urease |
| [153,155] |
| Avoiding the host’s immunological response |
| [153,155,165] |
| Virulence Factors | Description | References |
|---|---|---|
| The adhesive protein (LAP) |
| [193,194,195] |
| Internalines |
| [190,196,197,198,199,200] |
| Listeriolysin O |
| [190,201,202,203] |
| Secretion systems |
| [191] |
| Phospholipases C PlcA and PlcB |
| [189,201,204] |
| Superficial proteins ActaA |
| [201,205,206] |
| The OrfX protein |
| [192] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chlebicz, A.; Śliżewska, K. Campylobacteriosis, Salmonellosis, Yersiniosis, and Listeriosis as Zoonotic Foodborne Diseases: A Review. Int. J. Environ. Res. Public Health 2018, 15, 863. https://doi.org/10.3390/ijerph15050863
Chlebicz A, Śliżewska K. Campylobacteriosis, Salmonellosis, Yersiniosis, and Listeriosis as Zoonotic Foodborne Diseases: A Review. International Journal of Environmental Research and Public Health. 2018; 15(5):863. https://doi.org/10.3390/ijerph15050863
Chicago/Turabian StyleChlebicz, Agnieszka, and Katarzyna Śliżewska. 2018. "Campylobacteriosis, Salmonellosis, Yersiniosis, and Listeriosis as Zoonotic Foodborne Diseases: A Review" International Journal of Environmental Research and Public Health 15, no. 5: 863. https://doi.org/10.3390/ijerph15050863
APA StyleChlebicz, A., & Śliżewska, K. (2018). Campylobacteriosis, Salmonellosis, Yersiniosis, and Listeriosis as Zoonotic Foodborne Diseases: A Review. International Journal of Environmental Research and Public Health, 15(5), 863. https://doi.org/10.3390/ijerph15050863