Defense Mechanisms of Two Pioneer Submerged Plants during Their Optimal Performance Period in the Bioaccumulation of Lead: A Comparative Study

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Treatment Conditions
2.2. Quantification of Lead Bioaccumulation
2.3. Analysis of plant growth
2.4. Lipid Peroxidation and Enzyme Activity Assays
2.5. Statistical Analysis
3. Results and Discussion
3.1. Lead Accumulation
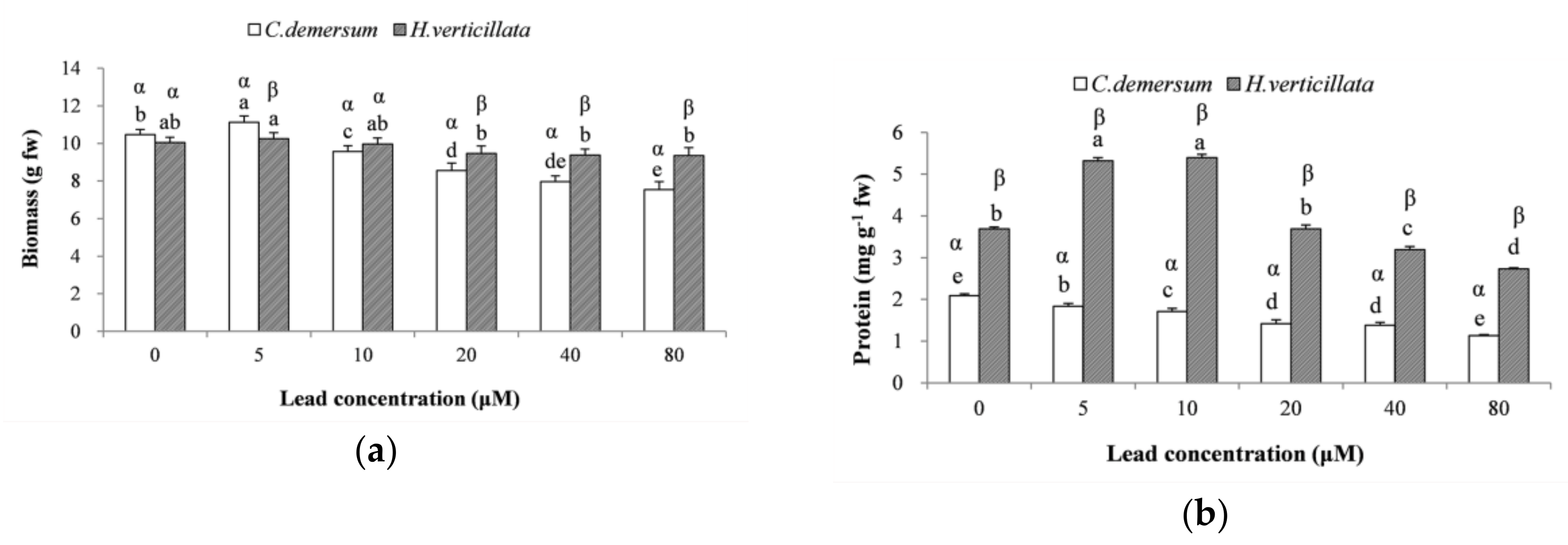
3.2. Growth Status
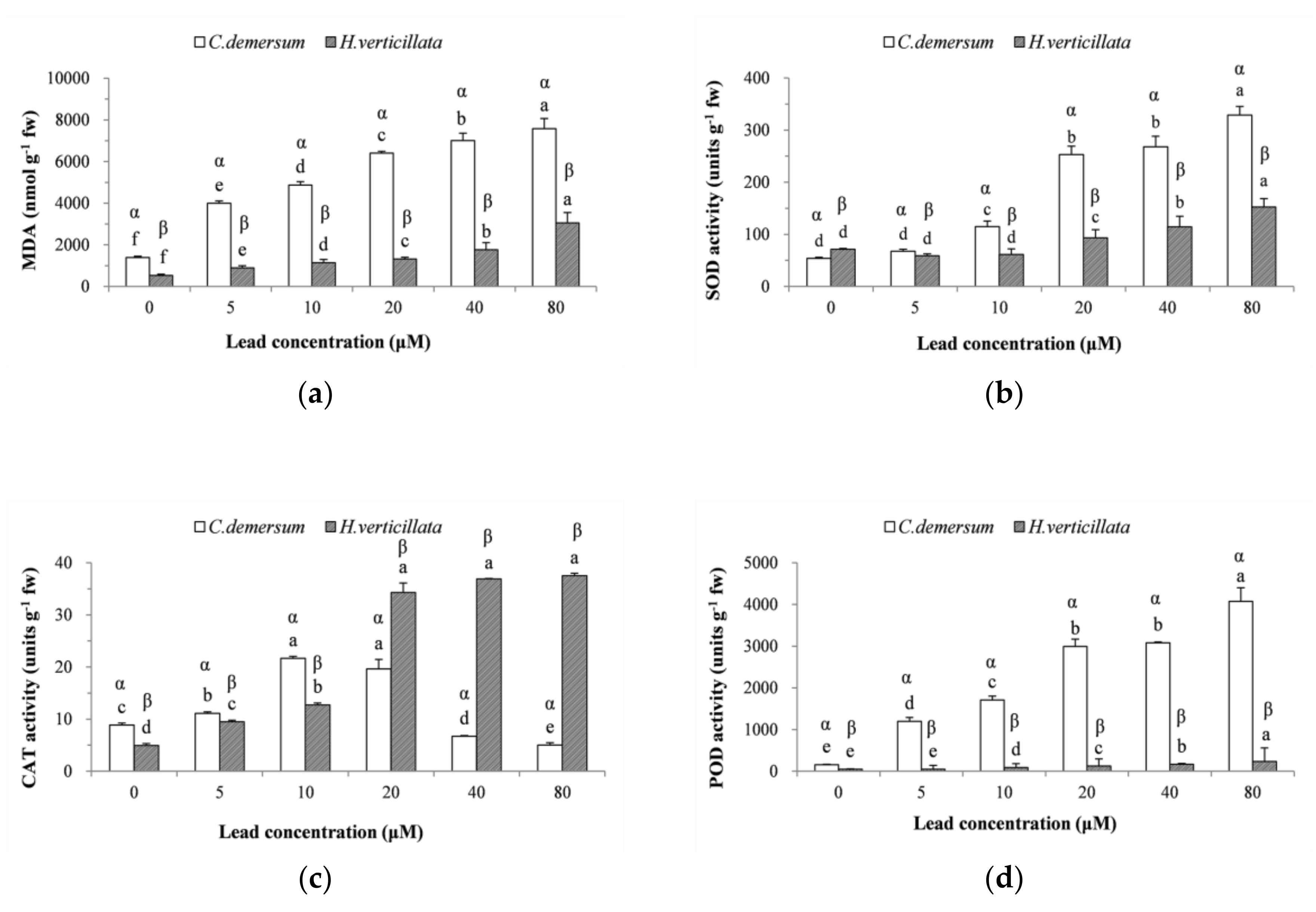
3.3. Lipid Peroxidation Products and Antioxidant Enzymes
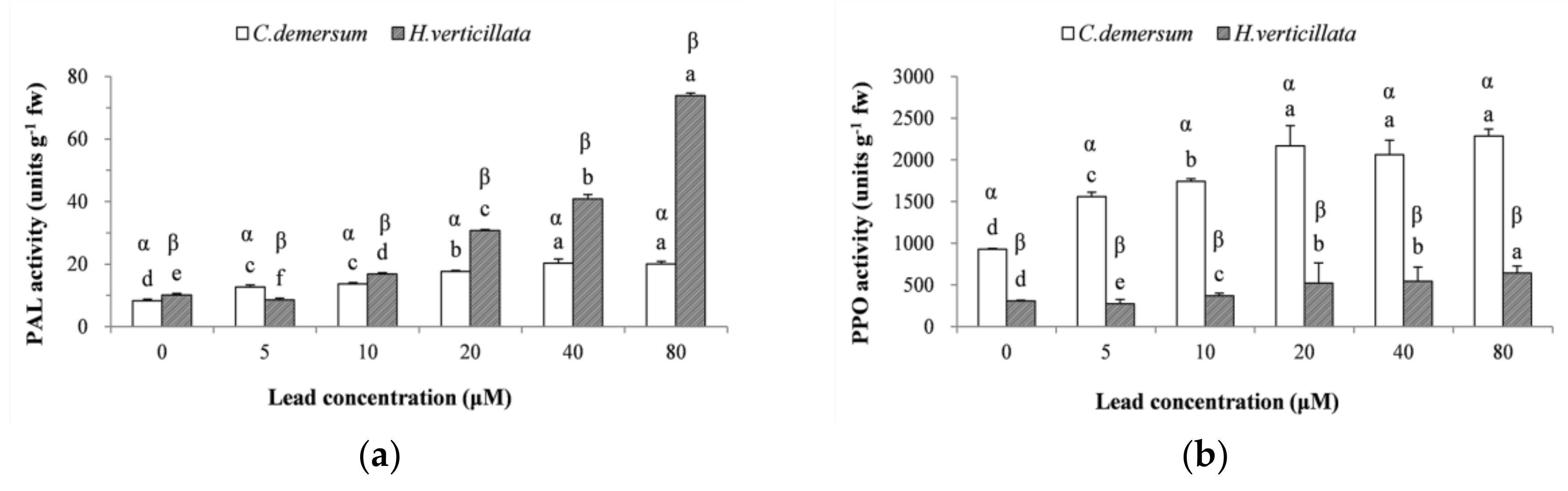
3.4. PAL and PPO Enzyme Activities
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- He, B.; Yun, Z.J.; Shi, J.B.; Jiang, G.B. Research progress of heavy metal pollution in China: Sources, analytical methods, status, and toxicity. Chin. Sci. Bull. 2013, 58, 134–140. [Google Scholar] [CrossRef]
- Gill, M. Heavy metal stress in plants: A review. Int. J. Adv. Res. 2014, 2, 1043–1055. [Google Scholar]
- Kumar, B.; Smita, K.; Flores, L.C. Plant mediated detoxification of mercury and lead. Arab. J. Chem. 2017, 10, S2335–S2342. [Google Scholar] [CrossRef]
- Ali, H.; Khan, E.; Sajad, M.A. Phytoremediation of heavy metals-concepts and applications. Chemosphere 2013, 91, 869–881. [Google Scholar] [CrossRef] [PubMed]
- Placek, A.; Grobelak, A.; Kacprzak, M. Improving the phytoremediation of heavy metals contaminated soil by use of sewage sludge. Int. J. Phytorem. 2013, 18, 605–618. [Google Scholar] [CrossRef] [PubMed]
- Rezania, S.; Taib, S.M.; Din, M.F.M.; Dahalan, F.A.; Kamyab, H. Comprehensive review on phytotechnology: Heavy metals removal by diverse aquatic plants species from wastewater. J. Hazard. Mater. 2016, 318, 587–599. [Google Scholar] [CrossRef] [PubMed]
- Rai, P.K. Heavy metal phytoremediation from aquatic ecosystems with special reference to macrophytes. Crit. Rev. Environ. Sci. Technol. 2009, 39, 697–753. [Google Scholar] [CrossRef]
- Cappa, J.J.; Pilon-Smits, E.A.H. Evolutionary aspects of elemental hyperaccumulation. Planta 2014, 239, 267–275. [Google Scholar] [CrossRef]
- Li, J.; Yu, H.; Luan, Y. Meta-analysis of the Copper, Zinc, and Cadmium absorption capacities of aquatic plants in heavy metal-polluted water. Int. J. Environ. Res. Public Health 2015, 12, 14958–14973. [Google Scholar] [CrossRef]
- Yadav, K.K.; Gupta, N.; Kumar, A.; Reece, L.M.; Singh, N.; Rezania, S.; Khan, S.A. Mechanistic understanding and holistic approach of phytoremediation: A review on application and future prospects. Ecol. Eng. 2018, 120, 274–298. [Google Scholar] [CrossRef]
- Tangahu, B.V.; Abdullah, S.R.; Basri, H.; Idris, M.; Anuar, N.; Mukhlisin, M. Phytoremediation of wastewater containing lead (Pb) in pilot reed bed using Scirpus grossus. Int. J. Phytoremed. 2013, 15, 663–676. [Google Scholar] [CrossRef]
- Sousa, W.T.Z.; Thomaz, S.M.; Murphy, K.J.; Silveira, M.J.; Mormul, R.P. Environmental predictors of the occurrence of exotic Hydrilla verticillata (L.f.) Royle and native Egeria najas Planch. in a sub-tropical river floodplain: The Upper River Paraná, Brazil. Hydrobiologia 2009, 632, 65–78. [Google Scholar] [CrossRef]
- Maleva, M.; Borisova, G.; Chukina, N.; Kumar, A.; Prasad, M.N.V. High dose of urea enhances the nickel and copper toxicity in Brazilian elodea (Egeria densa Planch. Casp.). Braz. J. Bot. 2016, 39, 965–972. [Google Scholar] [CrossRef]
- Jain, M.S.; Kalamdhad, A.S. A review on management of Hydrilla verticillata and its utilization as potential nitrogen-rich biomass for compost or biogas production. Bioresour. Technol. Rep. 2018, 1, 69–78. [Google Scholar] [CrossRef]
- Li, G.X.; Yan, C.Z.; Zhang, D.D.; Zhao, C.; Chen, G.Y. Cadmium(II) biosorption from aqueous solutions using Hydrilla verticillata. Can. J. Chem. Eng. 2013, 91, 1022–1030. [Google Scholar] [CrossRef]
- Nigam, S.; Gopal, K.; Vankar, P.S. Biosorption of arsenic in drinking water by submerged plant: Hydrilla verticilata. Environ. Sci. Pollut. Res. 2013, 20, 4000–4008. [Google Scholar] [CrossRef]
- Dogan, M.; Karatas, M.; Aasim, M. Cadmium and lead bioaccumulation potentials of an aquatic macrophyte Ceratophyllum demersum L.: A laboratory study. Ecotoxicol. Environ. Saf. 2017, 148, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Zhang, L.L.; Li, J.; He, X.J.; Chen, M.; Deng, Y. High-potential accumulation and tolerance in the submerged hydrophyte Hydrilla verticillata (L.f.) Royle for nickel-contaminated water. Ecotoxicol. Environ. Saf. 2018, 161, 553–562. [Google Scholar] [CrossRef]
- Maleva, M.; Borisova, G.; Chukina, N.; Kumar, A. Urea increased nickel and copper accumulation in the leaves of Egeria densa (planch.) Casp. and Ceratophyllum demersum L. during short-term exposure. Ecotoxicol. Environ. Saf. 2018, 148, 152. [Google Scholar] [CrossRef]
- Jain, S.K.; Vasudevan, P.; Jha, N.K. Removal of some heavy metals from polluted waters by aquatic plants: Studies on duckweed and water velvet. Biol. Wastes 1989, 28, 115–126. [Google Scholar] [CrossRef]
- Rai, U.N.; Sinha, S.; Tripathi, P.; Chandra, P. Wastewater treatability potential of some aquatic macrophytes: Removal of heavy metals. Ecol. Eng. 1995, 5, 5–12. [Google Scholar] [CrossRef]
- Abdallah, M.A. Phytoremediation of heavy metals from aqueous solutions by two aquatic macrophytes, Ceratophyllum demersum and Lemna gibba L. Environ. Technol. 2012, 33, 1609–1614. [Google Scholar] [CrossRef] [PubMed]
- Selamat, S.N.; Halmi, M.I.E.B.; Abdullah, S.R.S.; Idris, M.; Hasan, H.A.; Anuar, N. Optimization of lead (pb) bioaccumulation in Melastoma malabathricum L. by response surface methodology (RSM). Rendiconti Lincei Scienze Fisiche E Naturali 2018, 29, 43–51. [Google Scholar] [CrossRef]
- Li, B.; Gu, B.W.; Yang, Z.G.; Zhang, T. The role of submerged macrophytes in phytoremediation of arsenic from contaminated water: A case study on Vallisneria natans (Lour.) Hara. Ecotoxicol. Environ. Saf. 2018, 165, 224–231. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhang, L.L.; Li, J.; He, X.J.; Cai, J.C. Bioaccumulation and tolerance characteristics of a submerged plant (Ceratophyllum demersum, L.) exposed to toxic metal lead. Ecotoxicol. Environ. Saf. 2015, 122, 313–321. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Zhang, L.L.; Tuo, Y.C.; He, X.J.; Li, J.; Song, Y. Treatability thresholds for cadmium-contaminated water in the wetland macrophyte Hydrilla verticillata (L.f.) Royle. Ecol. Eng. 2016, 96, 178–186. [Google Scholar] [CrossRef]
- Hoagland, D.R.; Arnon, D.I. The water-culture method for growing plants without soil. Calif. Agric. Exp. Stn. Circ. 1950, 347, 1–32. [Google Scholar]
- Sedmak, J.J.; Grossberg, S.E. A rapid, sensitive, and versatile assay for protein using Coomassie brilliant blue G250. Anal. Biochem. 1977, 79, 544–552. [Google Scholar] [CrossRef]
- Beauchamp, C.; Fridovich, I. Superoxide dis-mutase: Improved assays and an assay applicable to acrylamide gels. Anal. Biochem. 1971, 44, 276–287. [Google Scholar] [CrossRef]
- Aebi, H. Catalase. In Methods of Enzymatic Analysis; Bergmeyer, H.U., Ed.; Verlag Chemie Weinheim Academic Press: New York, NY, USA, 1974; p. 680. [Google Scholar]
- Zhang, J.X.; Cui, S.P.; Li, J.M.; Kirkham, M.B. Protoplasmic factors, antoxidant responses, and chilling resistance in maize. Plant Physiol. Biochem. 1995, 33, 567–575. [Google Scholar]
- Hahlbrock, K.; Ragg, H. Light-induced changes of enzyme activities in parsley cell suspension cultures: Effects of inhibitors of RNA and protein synthesis. Arch. Biochem. Biophys. 1975, 166, 41–46. [Google Scholar] [CrossRef]
- Gao, S.; Chen, L.H.; Xu, Z.F.; Yuan, H. Changes of polyphenol oxidase activity during Jatropha curcas seed germination. J. Anim. Plant Sci. 2013, 18, 2659–2667. [Google Scholar]
- Ent, A.V.D.; Baker, A.J.M.; Reeves, R.D.; Pollard, A.J.; Schat, H. Hyperaccumulators of metal and metalloid trace elements: Facts and fiction. Plant Soil 2013, 362, 319–334. [Google Scholar] [CrossRef]
- Goolsby, E.W.; Mason, C.M. Response: Commentary: Toward a more physiologically and evolutionarily relevant definition of metal hyperaccumulation in plants. Front. Plant Sci. 2016, 6, 1252. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.B.; Li, D.F.; Tang, T.; Cai, Q.H. Studies on fractal character of macrophytes’ leaves. Acta Hydrobiol. Sin. 2004, 28, 23–28. (In Chinese) [Google Scholar]
- Pourrut, B.; Shahid, M.; Douay, F.; Dumat, C.; Pinelli, E. Molecular Mechanisms Involved in Lead Uptake, Toxicity and Detoxification in Higher Plants. In Heavy Metal Stress in Plants; Gupta, D.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2013; pp. 121–147. [Google Scholar]
- Singh, A.; Kumar, C.S.; Agarwal, A. Effect of lead and cadmium on aquatic plant Hydrilla verticillata. J. Environ. Biol. 2013, 34, 1027–1031. [Google Scholar] [PubMed]
- Emamverdian, A.; Ding, Y.; Mokhberdoran, F.; Xie, Y. Heavy metal stress and some mechanisms of plant defense response. Sci. World J. 2015, 2015, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Tripathi, R.D.; Dwivedi, S.; Kumar, A.; Trivedi, P.K.; Chakrabarty, D. Lead bioaccumulation potential of an aquatic macrophyte Najas indica are related to antioxidant system. Bioresour. Technol. 2010, 101, 3025–3032. [Google Scholar] [CrossRef] [PubMed]
- El-Khatib, A.A.; Hegazy, A.K.; Abo-El-Kassem, A.M. Bioaccumulation potential and physiological responses of aquatic macrophytes to Pb pollution. Int. J. Phytoremed. 2014, 16, 29–45. [Google Scholar] [CrossRef]
- Mishra, S.; Srivastava, S.; Tripathi, R.D.; Kumar, R.; Seth, C.S.; Gupta, D.K. Lead detoxification by coontail (Ceratophyllum demersum L.) involves induction of phytochelatins and antioxidant system in response to its accumulation. Chemosphere 2006, 65, 1027–1039. [Google Scholar] [CrossRef]
- Romero-Puertas, M.C.; Palma, J.M.; Gómez, M.; Río, L.A.D.; Sandalio, L.M. Cadmium causes the oxidative modification of proteins in pea plants. Plant Cell Environ. 2002, 25, 677–686. [Google Scholar] [CrossRef] [Green Version]
- Shahid, M.; Dumat, C.; Silvestre, J.; Pinelli, E. Effect of fulvic acids on lead-induced oxidative stress to metal sensitive Vicia faba, L. plant. Biol. Fertil. Soils 2012, 48, 689–697. [Google Scholar] [CrossRef]
- Ojuederie, O.B.; Babalola, O.O. Microbial and plant-assisted bioremediation of heavy metal polluted environments: A review. Int. J. Environ. Res. Public Health 2017, 14, 1504. [Google Scholar] [CrossRef] [PubMed]
- Gupta, D.K.; Huang, H.G.; Corpas, F.J. Lead tolerance in plants: Strategies for phytoremediation. Environ. Sci. Pollut. Res. 2013, 20, 2150–2161. [Google Scholar] [CrossRef] [PubMed]
- Malar, S.; Vikram, S.S.; Favas, P.J.; Perumal, V. Lead heavy metal toxicity induced changes on growth and antioxidative enzymes level in water hyacinths [Eichhornia crassipes (Mart.)]. Bot. Stud. 2014, 55, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Prasad, S.M. Antioxidant enzyme responses to the oxidative stress due to chlorpyrifos, dimethoate and dieldrin stress in palak (Spinacia oleracea L.) and their toxicity alleviation by soil amendments in tropical cropland. Sci. Total Environ 2017, 35, 985–999. [Google Scholar] [CrossRef] [PubMed]
- Willekens, H.; Chamnongpol, S.; Davey, M.; Schraudner, M.; Langebartels, C.; Van, M.M.; Inzé, D.; Van, C.W. Catalase is a sink for H2O2 and is indispensable for stress defense in C3 plants. Eur. Mol. Biol. Org. 1997, 16, 4806–4816. [Google Scholar] [CrossRef]
- Asgari, L.B.; Ghorbanpour, M.; Nikabadi, S. Heavy metals in contaminated environment: Destiny of secondary metabolite biosynthesis, oxidative status and phytoextraction in medicinal plants. Ecotoxicol. Environ. Saf. 2017, 145, 377–390. [Google Scholar] [CrossRef]
- Parlak, K.U.; Yilmaz, D.D. Ecophysiological tolerance of Lemna gibba L. exposed to cadmium. Ecotoxicol. Environ. Saf. 2013, 91, 79–85. [Google Scholar] [CrossRef]
- Li, F.T.; Qi, J.M.; Zhang, G.Y.; Lin, L.H.; Fang, P.P.; Tao, A.F.; Xu, J.T. Effect of cadmium stress on the growth antioxidative enzymes and lipid peroxidation in two kenaf (Hibiscus cannabinus L.) plant seedlings. J. Integr. Agric. 2013, 12, 610–620. [Google Scholar] [CrossRef]
- Macdonald, J.M.J.; D’Cunha, G.B.D.B. Erratum: A modern view of phenylalanine ammonia lyase. Biochem. Cell Biol. 2007, 85, 759–759. [Google Scholar] [CrossRef]
- Lavid, N.; Tel-Or, E. The involvement of polyphenols and peroxidase activities in heavy-metal accumulation by epidermal glands of the waterlily (nymphaeaceae). Planta 2001, 212, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Kováčik, J.; Bačkor, M. Phenylalanine ammonialyase and phenolic compounds in chamomile tolerance to cadmium and copper excess. Water Air Soil Pollut. 2007, 185, 1–4. [Google Scholar] [CrossRef]
- Ahsan, N.; Lee, D.G.; Alam, I.; Kim, P.J. Comparative proteomic study of arsenic-induced differentially expressed proteins in rice roots reveals glutathione plays a central role during as stress. Proteomics 2008, 8, 3561–3576. [Google Scholar] [CrossRef] [PubMed]
- Constabel, C.P.; Ryan, C.A. A survey of wound- and methyl jasmonate-induced leaf polyphenol oxidase in crop plants. Phytochemistry 1998, 47, 507–511. [Google Scholar] [CrossRef]
- Figueira, E.; Freitas, R.; Guasch, H.; Almeida, S.F. Efficiency of cadmium chelation by phytochelatins in Nitzschia palea (Kutzing) W. Smith. Ecotoxicology 2014, 23, 285–292. [Google Scholar] [CrossRef]
- Kovacik, J.; Dresler, S.; Peterkova, V.; Babula, P. Metal-induced oxidative stress in terrestrial macrolichens. Chemosphere 2018, 203, 402–409. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Concentration (μM) | Pb Accumulation (mg kg−1 dw) | |
|---|---|---|
| C. demersum | H. verticillata | |
| 0 | ND | ND |
| 5 | 626.0 d ± 36.7 | 222.1 e ± 12.2 |
| 10 | 1510.0 c ± 135.0 | 305.6 d ± 19.4 |
| 20 | 1653.8 c ± 77.7 | 680.6 c ± 36.7 |
| 40 | 1992.9 b ± 110.3 | 1000.4 b ± 23.5 |
| 80 | 2462.7 a ± 58.4 | 1792.0 a ± 80.5 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, D.; Zhang, L.; Chen, M.; He, X.; Li, J.; An, R. Defense Mechanisms of Two Pioneer Submerged Plants during Their Optimal Performance Period in the Bioaccumulation of Lead: A Comparative Study. Int. J. Environ. Res. Public Health 2018, 15, 2844. https://doi.org/10.3390/ijerph15122844
Li D, Zhang L, Chen M, He X, Li J, An R. Defense Mechanisms of Two Pioneer Submerged Plants during Their Optimal Performance Period in the Bioaccumulation of Lead: A Comparative Study. International Journal of Environmental Research and Public Health. 2018; 15(12):2844. https://doi.org/10.3390/ijerph15122844
Chicago/Turabian StyleLi, Dian, Linglei Zhang, Min Chen, Xiaojia He, Jia Li, and Ruidong An. 2018. "Defense Mechanisms of Two Pioneer Submerged Plants during Their Optimal Performance Period in the Bioaccumulation of Lead: A Comparative Study" International Journal of Environmental Research and Public Health 15, no. 12: 2844. https://doi.org/10.3390/ijerph15122844