Decreased Capacity for Sperm Production Induced by Perinatal Bisphenol A Exposure Is Associated with an Increased Inflammatory Response in the Offspring of C57BL/6 Male Mice

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Analysis of Sperm Count and Sperm Abnormality Rate
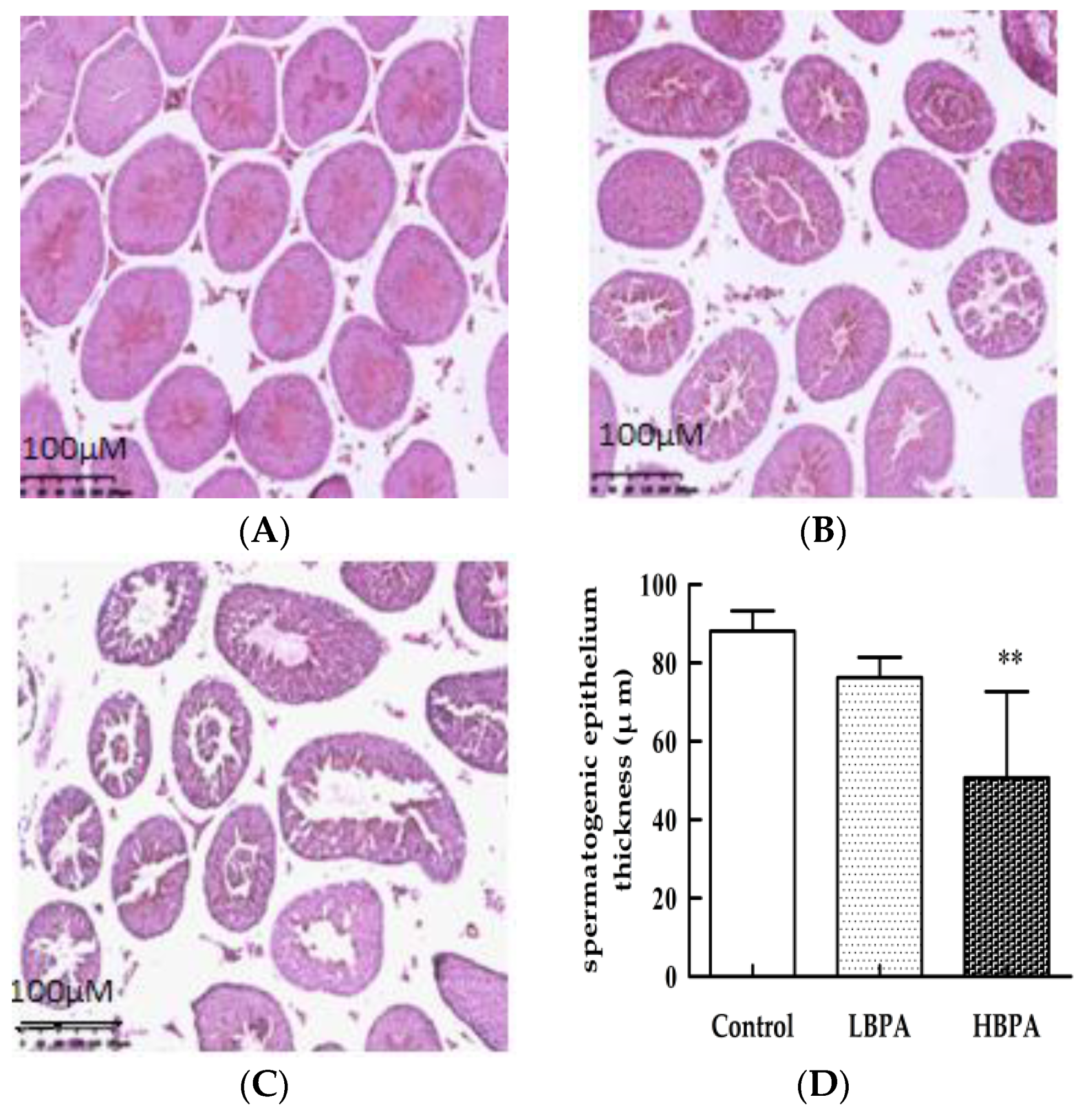
2.3. Histological Change in the Testis
2.4. Biochemical Assays
2.5. Western Blot Analysis
2.6. Statistical Analysis
3. Results
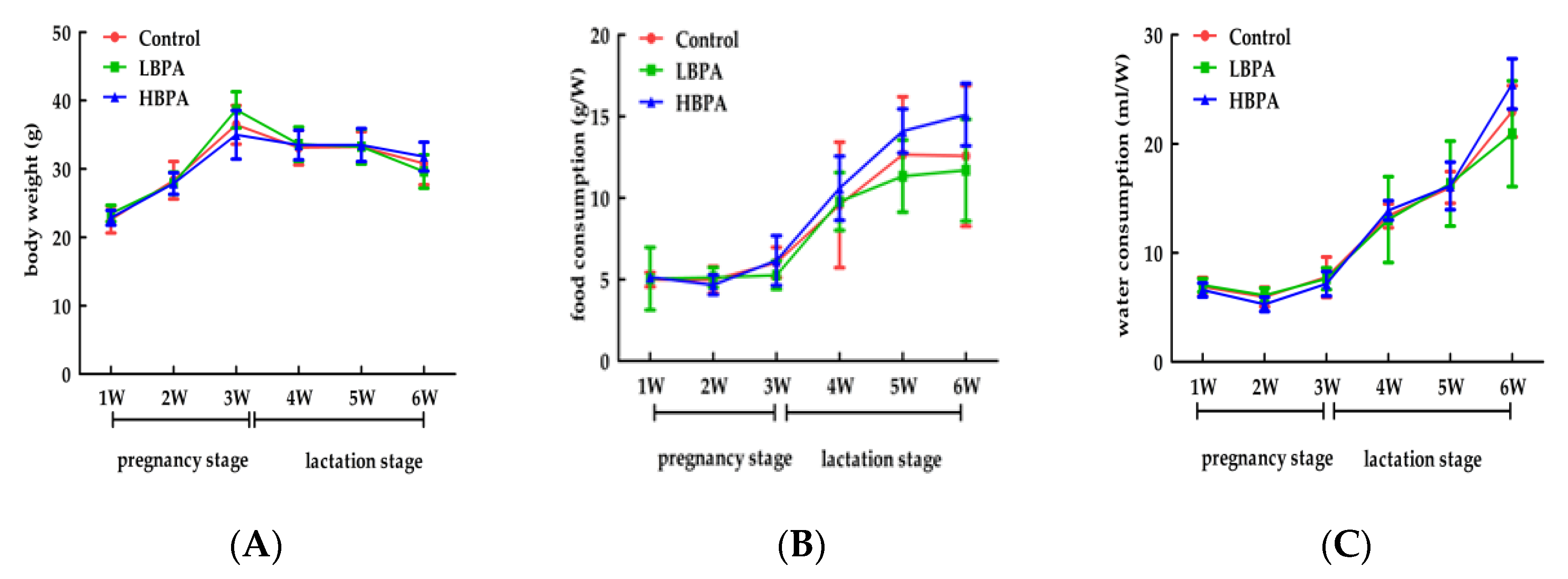
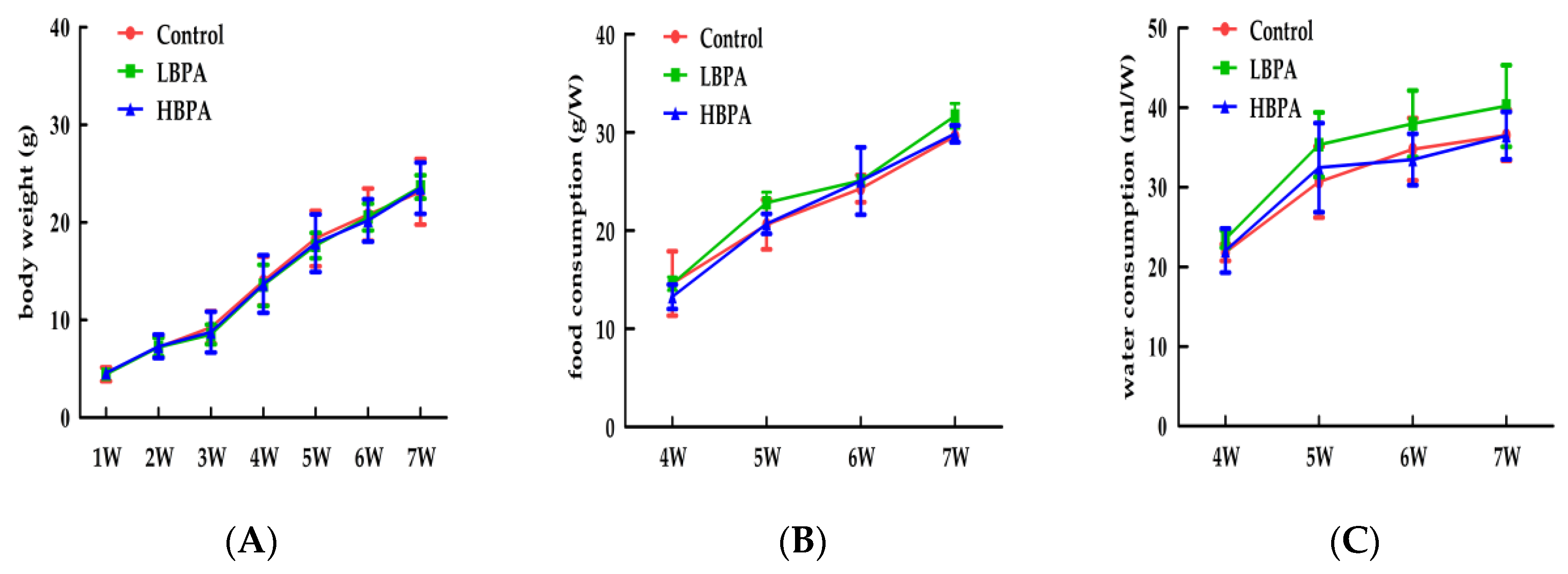
3.1. Effect of Perinatal Bisphenol A Exposurure on Body Weight and Food, Water Intake in Maternal Mice and Their Offspring
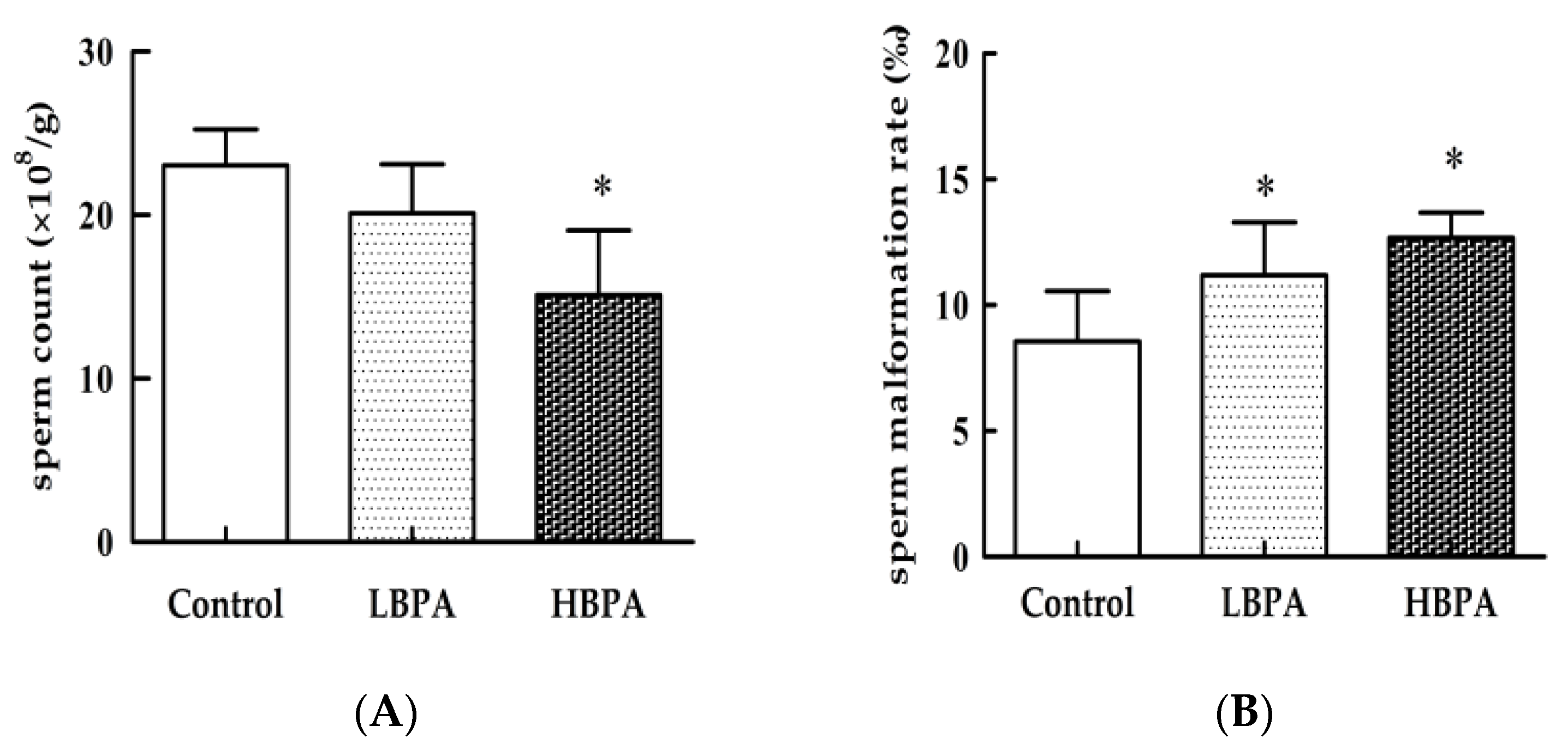
3.2. The Effect of Perinatal Bisphenol A Exposure on Sperm Production Capacity in Male Offspring
3.3. The Effect of Perinatal Exposure to Bisphenol A on Levels of Serum Testosterone and Tumor Necrosis Factor-α in Male Offspring
3.4. The Effect of Perinatal Exposure to Bisphenol A on Inflammatory Factor-Related Protein Expression in Testis Tissue from Male Offspring
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Anifandis, G.; Amiridis, G.; Dafopoulos, K.; Daponte, A.; Dovolou, E.; Gavriil, E.; Gorgogietas, V.; Kachpani, E.; Mamuris, Z.; Messini, C.I.; et al. The in vitro impact of the herbicide roundup on human sperm motility and sperm mitochondria. Toxics 2017, 6, 2. [Google Scholar] [CrossRef] [PubMed]
- Anifandis, G.; Katsanaki, K.; Lagodonti, G.; Messini, C.; Simopoulou, M.; Dafopoulos, K.; Daponte, A. The effect of glyphosate on human sperm motility and sperm DNA fragmentation. Int. J. Environ. Res. Public Health 2018, 15, 1117. [Google Scholar] [CrossRef] [PubMed]
- Costa, N.O.; Vieira, M.L.; Sgarioni, V.; Pereira, M.R.F.; Montagnini, B.G.; de Fátima Paccola Mesquita, S.; Gerardin, D.C.C. Evaluation of the reproductive toxicity of fungicide propiconazole in male rats. Toxicology 2015, 335, 55–61. [Google Scholar] [CrossRef] [PubMed]
- Konieczna, A.; Rutkowska, A.; Rachoń, D. Health risk of exposure to Bisphenol A (BPA). Roczniki Państwowego Zakładu Higieny 2015, 66, 5–11. [Google Scholar] [PubMed]
- Lehmler, H.J.; Liu, B.; Gadogbe, M.; Bao, W. Exposure to bisphenol A, bisphenol F, and bisphenol S in U.S. adults and children: The national health and nutrition examination survey 2013–2014. ACS Omega 2018, 3, 6523–6532. [Google Scholar] [CrossRef] [PubMed]
- Zimmers, S.M.; Browne, E.P.; O’Keefe, P.W.; Anderton, D.L.; Kramer, L.; Reckhow, D.A.; Arcaro, K.F. Determination of free bisphenol A (BPA) concentrations in breast milk of U.S. women using a sensitive LC/MS/MS method. Chemosphere 2014, 104, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Choi, K.; Park, J.; Moon, H.B.; Choi, G.; Lee, J.J.; Suh, E.; Kim, H.J.; Eun, S.H.; Kim, G.H.; et al. Bisphenol A distribution in serum, urine, placenta, breast milk, and umbilical cord serum in a birth panel of mother-neonate pairs. Sci. Total Environ. 2018, 626, 1494–1501. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Gupta, P.; Chandolia, A.; Alam, I. Bisphenol A: A threat to human health. J. Environ. Health 2015, 77, 20–26. [Google Scholar] [PubMed]
- Li, D.K.; Zhou, Z.; Miao, M.; He, Y.; Wang, J.; Ferber, J.; Herrinton, L.J.; Gao, E.; Yuan, W. Urine bisphenol-A (BPA) level in relation to semen quality. Fertil. Steril. 2011, 95, 625–630.e4. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhou, W.; Zhu, W.; Chen, L.; Wang, W.; Tian, Y.; Shen, L.; Zhang, J. Associations of female exposure to bisphenol A with fecundability: Evidence from a preconception cohort study. Environ. Int. 2018, 117, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Brouard, V.; Guénon, I.; Bouraima-Lelong, H.; Delalande, C. Differential effects of bisphenol A and estradiol on rat spermatogenesis’ establishment. Reprod. Toxicol. 2016, 63, 49–61. [Google Scholar] [CrossRef] [PubMed]
- Okada, A.; Kai, O. Effects of estradiol-17 beta and bisphenol A administered chronically to mice throughout pregnancy and lactation on the male pups’ reproductive system. Asian J. Androl. 2008, 10, 271–276. [Google Scholar] [CrossRef] [PubMed]
- Peretz, J.; Vrooman, L.; Ricke, W.A.; Hunt, P.A.; Ehrlich, S.; Hauser, R.; Padmanabhan, V.; Taylor, H.S.; Swan, S.H.; VandeVoort, C.A.; et al. Bisphenol a and reproductive health: Update of experimental and human evidence, 2007–2013. Environ. Health Perspect. 2014, 122, 775–786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, S.; Shi, W.; Wang, X.; Song, P.; Zhong, X. Bisphenol A exposure during pregnancy alters the mortality and levels of reproductive hormones and genes in offspring mice. Biomed. Res. Int. 2017, 2017, 3585809. [Google Scholar] [CrossRef] [PubMed]
- Lemmen, J.G.; Arends, R.J.; Van der Saag, P.T.; Van der Burg, B. In vivo imaging of activated estrogen receptors in utero by estrogens and bisphenol A. Environ. Health Perspect. 2004, 112, 1544–1549. [Google Scholar] [CrossRef] [PubMed]
- Nanjappa, M.K.; Simon, L.; Akingbemi, B.T. The industrial chemical bisphenol A (BPA) interferes with proliferative activity and development of steroidogenic capacity in rat Leydig cells. Biol. Reprod. 2012, 86, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Lau, S.W.; Fan, Y.; Wu, R.S.S.; Ge, W. Juvenile exposure to bisphenol A promotes ovarian differentiation but suppresses its growth-potential involvement of pituitary follicle-stimulating hormone. Aquat. Toxicol. 2017, 193, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ding, Z.; Shi, Q.M.; Ge, X.; Wang, H.X.; Li, M.X.; Chen, G.; Wang, Q.; Ju, Q.; Zhang, J.P.; et al. Anti-androgenic mechanisms of bisphenol A involve androgen receptor signaling pathway. Toxicology 2017, 387, 10–16. [Google Scholar] [CrossRef] [PubMed]
- Shi, M.; Sekulovski, N.; MacLean, J.A.; Hayashi, K. Effects of bisphenol A analogues on reproductive functions in mice. Reprod. Toxicol. 2017, 73, 280–291. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Duan, W.; Li, R.; Xu, S.; Zhang, L.; Chen, C.; He, M.; Lu, Y.; Wu, H.; Pi, H.; et al. Exposure to bisphenol A disrupts meiotic progression during spermatogenesis in adult rats through estrogen-like activity. Cell Death Dis. 2013, 4, e676. [Google Scholar] [CrossRef] [PubMed]
- Koike, E.; Yanagisawa, R.; Win-Shwe, T.T.; Takano, H. Exposure to low-dose bisphenol A during the juvenile period of development disrupts the immune system and aggravates allergic airway inflammation in mice. Int. J. Immunopathol. Pharmacol. 2018, 32, 2058738418774897. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Park, J.; Bui, P.T.C.; Choi, K.; Gye, M.C.; Hong, Y.C.; Kim, J.H.; Lee, Y.J. Bisphenol A induces COX-2 through the mitogen-activated protein kinase pathway and is associated with levels of inflammation-related markers in elderly populations. Environ. Res. 2017, 158, 490–498. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, P.; Romano, R.M.; Kizys, M.M.; Oliveira, K.C.; Kasamatsu, T.; Giannocco, G.; Chiamolera, M.I.; Dias-da-Silva, M.R.; Romano, M.A. Adult exposure to bisphenol A (BPA) in Wistar rats reduces sperm quality with disruption of the hypothalamic-pituitary-testicular axis. Toxicology 2015, 329, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xie, M.; Bu, P.; Li, F.; Lan, S.; Wu, H.; Yuan, L.; Wang, Y. Neonatal bisphenol A exposure induces meiotic arrest and apoptosis of spermatogenic cells. Oncotarget 2016, 7, 10606–10615. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eladak, S.; Moison, D.; Guerquin, M.J.; Matilionyte, G.; Kilcoyne, K.; N’Tumba-Byn, T.; Messiaen, S.; Deceuninck, Y.; Pozzi-Gaudin, S.; Benachi, A.; et al. Effects of environmental Bisphenol A exposures on germ cell development and Leydig cell function in the human fetal testis. PLoS ONE 2018, 13, e0191934. [Google Scholar] [CrossRef] [PubMed]
- Nesan, D.; Sewell, L.C.; Kurrasch, D.M. Opening the black box of endocrine disruption of brain development: Lessons from the characterization of Bisphenol A. Horm. Behav. 2018, 101, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Valentino, R.; D’Esposito, V.; Passaretti, F.; Liotti, A.; Cabaro, S.; Longo, M.; Perruolo, G.; Oriente, F.; Beguinot, F.; Formisano, P. Bisphenol-A impairs insulin action and up-regulates inflammatory pathways in human subcutaneous adipocytes and 3T3-L1 cells. PLoS ONE 2013, 8, e82099. [Google Scholar] [CrossRef] [PubMed]
- Rochefort, H. Endocrine disruptors (EDs) and hormone-dependent cancers: Correlation or causal relationship? C. R. Biol. 2017, 340, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Valentino, R.; D’Esposito, V.; Ariemma, F.; Cimmino, I.; Beguinot, F.; Formisano, P. Bisphenol A environmental exposure and the detrimental effects on human metabolic health: Is it necessary to revise the risk assessment in vulnerable population? J. Endocrinol. Investig. 2016, 39, 259–263. [Google Scholar] [CrossRef] [PubMed]
- Izumi, Y.; Yamaguchi, K.; Ishikawa, T.; Ando, M.; Chiba, K.; Hashimoto, H.; Shiotani, M.; Fujisawa, M. Molecular changes induced by bisphenol-A in rat Sertoli cell culture. Syst. Biol. Reprod. Med. 2011, 57, 228–232. [Google Scholar] [CrossRef] [PubMed]
- Ogo, F.M.; de Lion Siervo, G.E.M.; Staurengo-Ferrari, L.; de Oliveira Mendes, L.; Luchetta, N.R.; Vieira, H.R.; Fattori, V.; Verri, W.A., Jr.; Scarano, W.R.; Fernandes, G.S.A. Bisphenol A exposure impairsepididymal development during the peripubertal period of rats: Inflammatory profile and tissue changes. Basic Clin. Pharmacol. Toxicol. 2018, 122, 262–270. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.P.; Bradfield, C.A. The search for endogenous activators of the aryl hydrocarbon receptor. Chem. Res. Toxicol. 2008, 21, 102–116. [Google Scholar] [CrossRef] [PubMed]
- Baricza, E.; Tamási, V.; Marton, N.; Buzás, E.I.; Nagy, G. The emerging role of aryl hydrocarbon receptor in the activation and differentiation of Th17 cells. Cell Mol. Life Sci. 2016, 73, 95–117. [Google Scholar] [CrossRef] [PubMed]
- La-Rocca, C.; Tait, S.; Guerranti, C.; Busani, L.; Ciardo, F.; Bergamasco, B.; Perra, G.; Mancini, F.R.; Marci, R.; Bordi, G.; et al. Exposure to endocrine disruptors and nuclear receptors gene expression in infertile and fertile men from Italian Areas with different environmental features. Int. J. Environ. Res. Public Health 2015, 12, 12426–12445. [Google Scholar] [CrossRef] [PubMed]
- La-Rocca, C.; Tait, S.; Guerranti, C.; Busani, L.; Ciardo, F.; Bergamasco, B.; Stecca, L.; Perra, G.; Mancini, F.R.; Marci, R.; et al. Exposure to endocrine disrupters and nuclear receptor gene expression in infertile and fertile women from different Italian areas. Int. J. Environ. Res. Public Health 2014, 11, 10146–10164. [Google Scholar] [CrossRef] [PubMed]
- Ziv-Gal, A.; Craig, Z.R.; Wang, W.; Flaws, J.A. Bisphenol A inhibits cultured mouse ovarian follicle growth partially via the aryl hydrocarbon receptor signaling pathway. Reprod. Toxicol. 2013, 42, 58–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, E.A.; Mezrich, J.D.; Bradfield, C.A. The aryl hydrocarbon receptor: A perspective on potential roles in the immune system. Immunology 2009, 127, 299–311. [Google Scholar] [CrossRef] [PubMed]
- Vogel, C.F.; Li, W.; Wu, D.; Miller, J.K.; Sweeney, C.; Lazennec, G.; Fujisawa, Y.; Matsumura, F. Interaction of aryl hydrocarbon receptor and NF-κB subunit RelB in breast cancer is associated with interleukin-8 overexpression. Arch. Biochem. Biophys. 2011, 512, 78–86. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Testis | Epididymis | ||
|---|---|---|---|---|
| Weight (g) | Coefficient (%) | Weight (g) | Coefficient (%) | |
| Control | 0.15 ± 0.03 | 0.65 ± 0.06 | 0.08 ± 0.02 | 0.37 ± 0.03 |
| LBPA | 0.16 ± 0.02 | 0.70 ± 0.09 | 0.09 ± 0.01 | 0.39 ± 0.05 |
| HBPA | 0.16 ± 0.03 | 0.68 ± 0.08 | 0.08 ± 0.01 | 0.35 ± 0.04 |
| Group | n | Testosterone (ng/mL) | TNF-α (ng/L) |
|---|---|---|---|
| Control | 7 | 13.47 ± 1.12 | 3.31 ± 0.58 |
| LBPA | 7 | 12.93 ± 0.98 | 4.26 ± 0.35 * |
| HBPA | 7 | 10.69 ± 1.04 ** | 4.46 ± 0.37 * |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, Y.; Lin, R.; Wu, F.; Sun, Q.; Jia, L. Decreased Capacity for Sperm Production Induced by Perinatal Bisphenol A Exposure Is Associated with an Increased Inflammatory Response in the Offspring of C57BL/6 Male Mice. Int. J. Environ. Res. Public Health 2018, 15, 2158. https://doi.org/10.3390/ijerph15102158
Meng Y, Lin R, Wu F, Sun Q, Jia L. Decreased Capacity for Sperm Production Induced by Perinatal Bisphenol A Exposure Is Associated with an Increased Inflammatory Response in the Offspring of C57BL/6 Male Mice. International Journal of Environmental Research and Public Health. 2018; 15(10):2158. https://doi.org/10.3390/ijerph15102158
Chicago/Turabian StyleMeng, Yuan, Ren Lin, Fengjuan Wu, Qi Sun, and Lihong Jia. 2018. "Decreased Capacity for Sperm Production Induced by Perinatal Bisphenol A Exposure Is Associated with an Increased Inflammatory Response in the Offspring of C57BL/6 Male Mice" International Journal of Environmental Research and Public Health 15, no. 10: 2158. https://doi.org/10.3390/ijerph15102158
APA StyleMeng, Y., Lin, R., Wu, F., Sun, Q., & Jia, L. (2018). Decreased Capacity for Sperm Production Induced by Perinatal Bisphenol A Exposure Is Associated with an Increased Inflammatory Response in the Offspring of C57BL/6 Male Mice. International Journal of Environmental Research and Public Health, 15(10), 2158. https://doi.org/10.3390/ijerph15102158