Effect of Ampicillin, Streptomycin, Penicillin and Tetracycline on Metal Resistant and Non-Resistant Staphylococcus aureus






Abstract
:1. Introduction
2. Experimental Section
2.1. Cultivation of S. aureus
2.2. Preparation of Resistant Strains of S. aureus
2.3. Chemicals, Preparation of Deionised Water and pH Measurement
2.4. Heavy Metals Ions Preparation
2.5. The Microscopy of the Cells in Ambient Light
2.6. Determination of Growth Curves
2.7. Determination of Cadmium and Lead Ions by Atomic Absorption Spectrometry
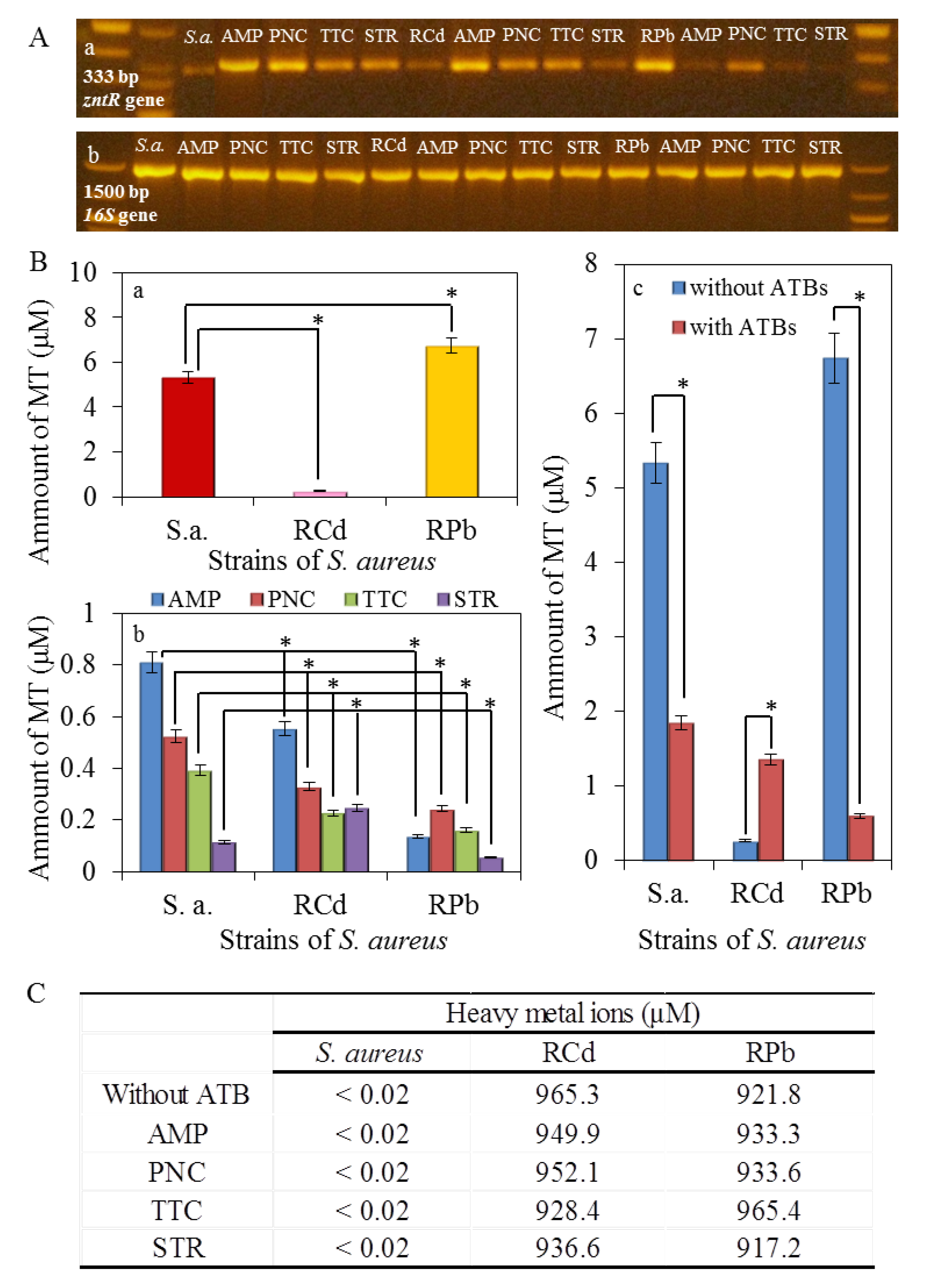
2.8. Interaction of Bacterial DNA Fragment of zntR and 16S Gene with Cadmium and Lead Ions
2.9. Determination of Metallothionein
2.10. Expression of ZntR Gene and 16S Gene
2.10.1. Isolation of RNA
2.10.2. Amplification of mRNA
2.10.3. Amplification of cDNA for zntR Gene
2.10.4. Amplification of cDNA for 16S Gene
2.11. Statistical Analyses
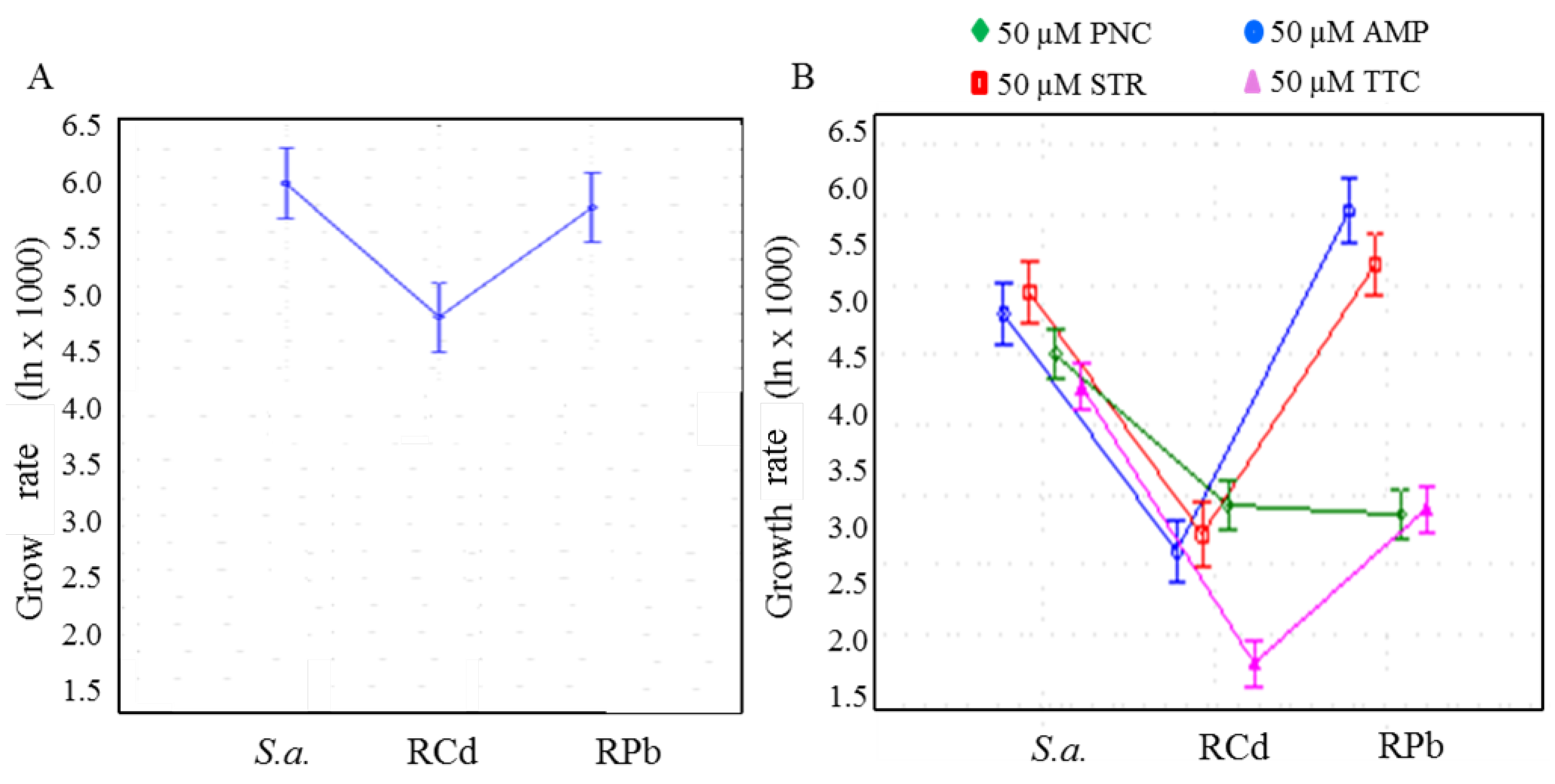
3. Results and Discussion
3.1. Characterization on the Cellular Level
3.1.1. Morphological Characterization

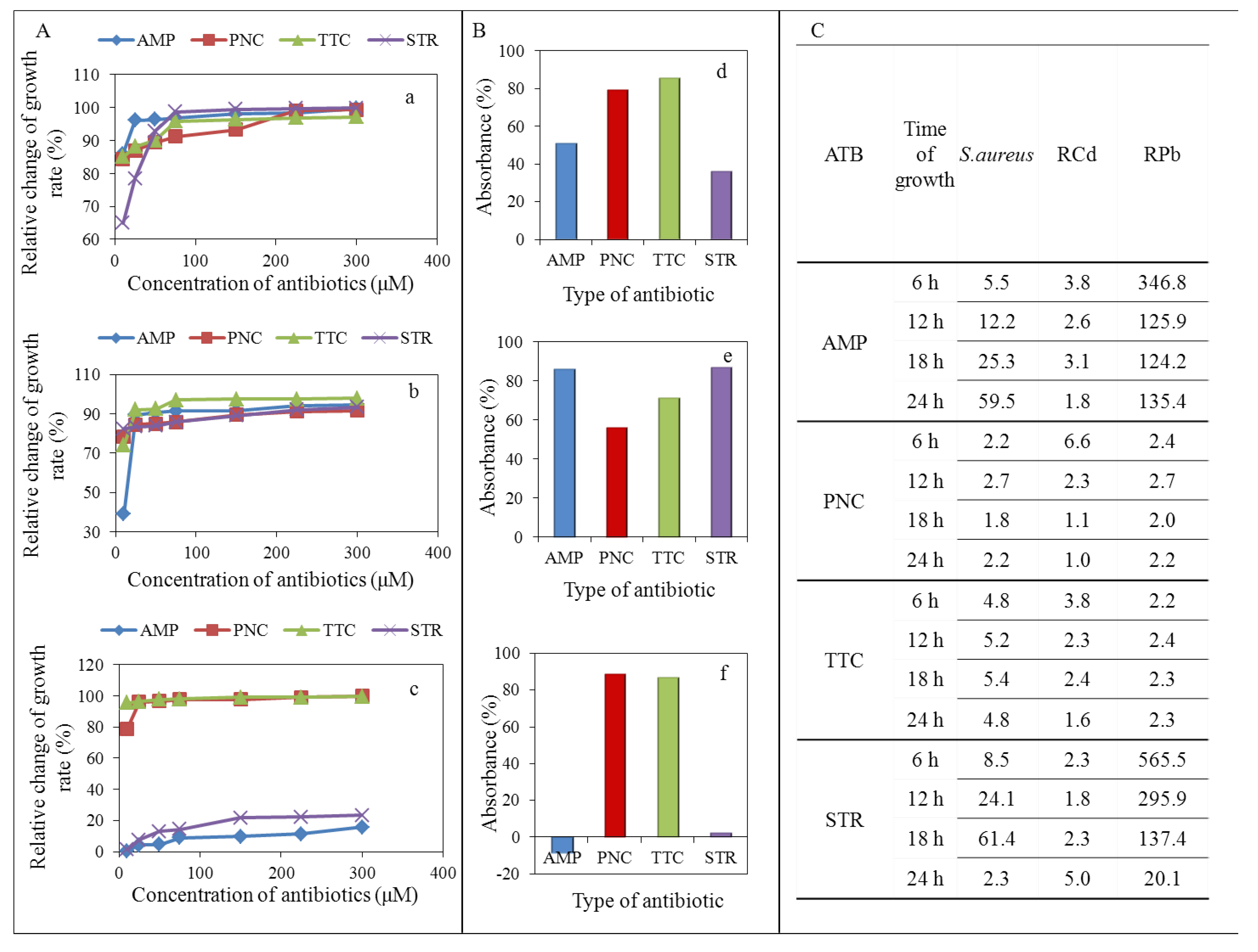
3.1.2. Determination of Antimicrobial Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotics | S. aureus strains | N | Absorbance (mean ± S.D × 103) | Group different at p < 0.05 |
| Penicillin (50 µM) | ||||
| RPb | 48 | 26.01 ± 0.76 | **** | |
| RCd | 46 | 28.78 ± 0.9 | **** | |
| S. a. | 48 | 90.81 ± 0.93 | **** | |
| Streptomycin (50 µM) | ||||
| RCd | 46 | 21.83 ± 0.22 | **** | |
| S. a. | 48 | 121.62 ± 1.26 | **** | |
| RPb | 48 | 254.09 ± 0.83 | **** | |
| Ampicillin (50 µM) | ||||
| RCd | 46 | 23.33 ± 3.21 | **** | |
| S. a. | 48 | 175.22 ± 4.95 | **** | |
| RPb | 48 | 323.05 ± 5.15 | **** | |
| Tetracycline (50 µM) | ||||
| RCd | 46 | 9.86 ± 0.83 | **** | |
| RPb | 48 | 30.04 ± 0.35 | **** | |
| S. a. | 48 | 71.91 ± 0.58 | **** | |
3.2. Characterization on the Molecular Level
 0,
0,  60,
60,  240 and
240 and  950 μM): (A) Interaction of 16S gene with cadmium ions, (a) decrease of absorbance of 16S gene with cadmium ions in 260 nm. (B) Interaction of 16S gene with lead ions, (b) decrease of absorbance of 16S gene with lead ions in 260 nm. (C) Interaction of zntR gene with cadmium ions, (c) decrease of absorbance of zntR gene with cadmium ions in 260 nm. (D) Interaction of zntR gene with lead ions, (d) decrease of absorbance of zntR gene with lead ions in 260 nm.
0, 60, 240 and 950 μM): (A) Interaction of 16S gene with cadmium ions, (a) decrease of absorbance of 16S gene with cadmium ions in 260 nm. (B) Interaction of 16S gene with lead ions, (b) decrease of absorbance of 16S gene with lead ions in 260 nm. (C) Interaction of zntR gene with cadmium ions, (c) decrease of absorbance of zntR gene with cadmium ions in 260 nm. (D) Interaction of zntR gene with lead ions, (d) decrease of absorbance of zntR gene with lead ions in 260 nm.
950 μM): (A) Interaction of 16S gene with cadmium ions, (a) decrease of absorbance of 16S gene with cadmium ions in 260 nm. (B) Interaction of 16S gene with lead ions, (b) decrease of absorbance of 16S gene with lead ions in 260 nm. (C) Interaction of zntR gene with cadmium ions, (c) decrease of absorbance of zntR gene with cadmium ions in 260 nm. (D) Interaction of zntR gene with lead ions, (d) decrease of absorbance of zntR gene with lead ions in 260 nm.
0, 60, 240 and 950 μM): (A) Interaction of 16S gene with cadmium ions, (a) decrease of absorbance of 16S gene with cadmium ions in 260 nm. (B) Interaction of 16S gene with lead ions, (b) decrease of absorbance of 16S gene with lead ions in 260 nm. (C) Interaction of zntR gene with cadmium ions, (c) decrease of absorbance of zntR gene with cadmium ions in 260 nm. (D) Interaction of zntR gene with lead ions, (d) decrease of absorbance of zntR gene with lead ions in 260 nm.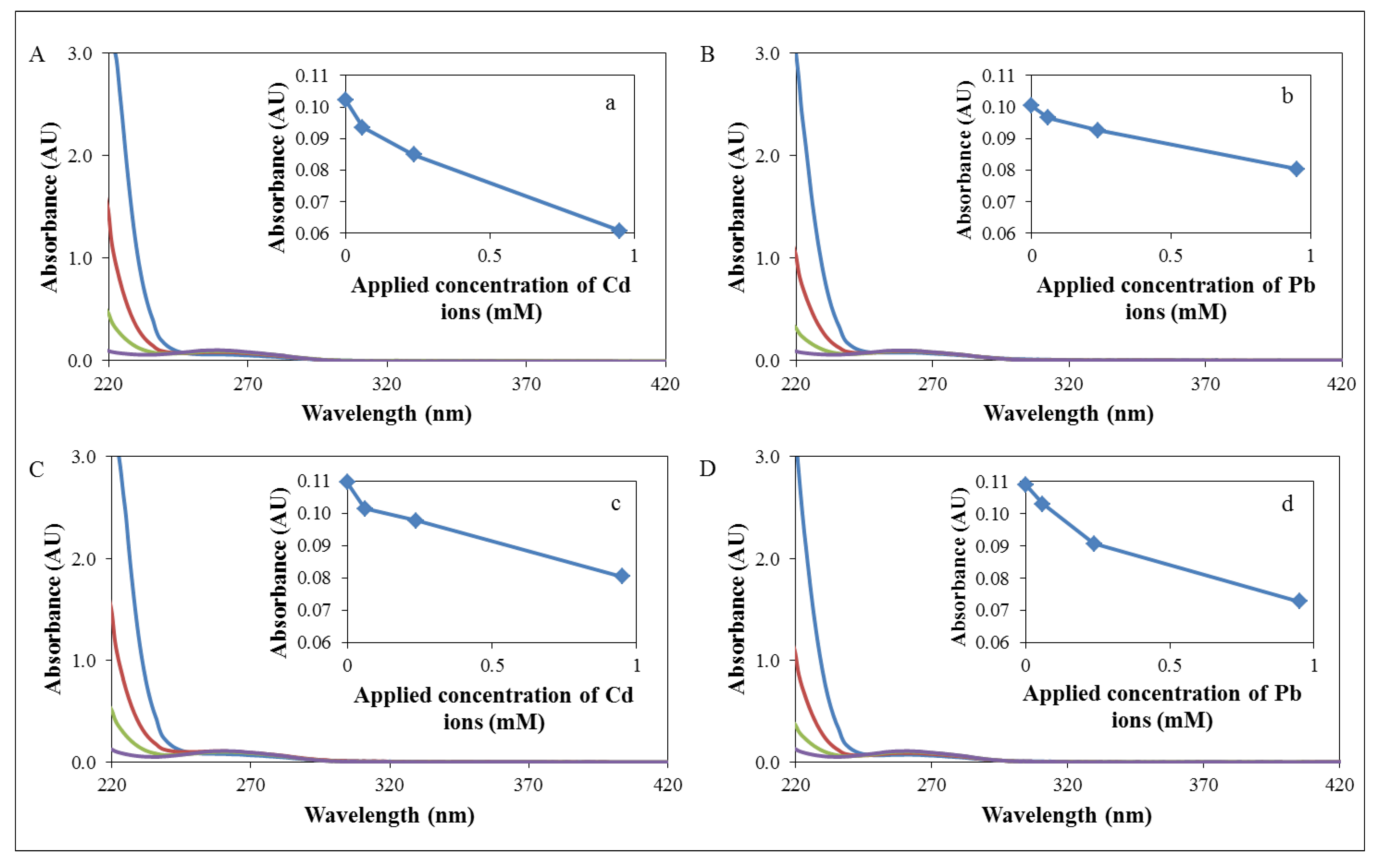
4. Conclusions
Acknowledgments
Authors Contributions
Conflicts of Interest
References
- Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: Causes, challenges and responses. Nat. Med. 2004, 10, S122–S129. [Google Scholar] [CrossRef]
- Zhou, F.; Wang, Y. Characteristics of antibiotic resistance of airborne Staphylococcus isolated from metro stations. Int. J. Environ. Res. Public Health 2013, 10, 2412–2426. [Google Scholar] [CrossRef]
- Gatica, J.; Cytryn, E. Impact of treated wastewater irrigation on antibiotic resistance in the soil microbiome. Environ. Sci. Pollut. Res. 2013, 20, 3529–3538. [Google Scholar] [CrossRef]
- Stone, N.D.; Lewis, D.R.; Lowery, H.K.; Darrow, L.A.; Kroll, C.M.; Gaynes, R.P.; Jernigan, J.A.; McGowan, J.E.; Tenover, F.C.; Richards, C.L. Importance of bacterial burden among methicillin-resistant Staphylococcus aureus carriers in a long-term care facility. Infect. Control Hosp. Epidemiol. 2008, 29, 143–148. [Google Scholar] [CrossRef]
- Bastug, A.; Yilmaz, G.R.; Kayaaslan, B.; Akinci, E.; Bodur, H. Risk factors for mortality in patients with nosocomial Staphylococcus aureus bacteremia. Turk. J. Med. Sci. 2012, 42, 1222–1229. [Google Scholar]
- Tacconelli, E.; Pop-Vicas, A.E.; D’Agata, E.M.C. Increased mortality among elderly patients with meticillin-resistant Staphylococcus aureus bacteraemia. J. Hosp. Infect. 2006, 64, 251–256. [Google Scholar] [CrossRef]
- Cosgrove, S.E.; Qi, Y.L.; Kaye, K.S.; Harbarth, S.; Karchmer, A.W.; Carmeli, Y. The impact of methicillin-resistance in Staphylococcus aureus bacteremia on patient outcomes: Mortality, length of stay, and hospital charges. Infect. Control Hosp. Epidemiol. 2005, 26, 166–174. [Google Scholar]
- Ammerlaan, H.S.M.; Harbarth, S.; Buiting, A.G.M.; Crook, D.W.; Fitzpatrick, F.; Hanberger, H.; Herwaldt, L.A.; van Keulen, P.H.J.; Kluytmans, J.; Kola, A.; et al. Secular trends in nosocomial bloodstream infections: Antibiotic-resistant bacteria increase the total burden of infection. Clin. Infect. Dis. 2013, 56, 798–805. [Google Scholar] [CrossRef]
- Ye, Y.; Li, S.L.; Li, Y.J.; Ren, T.S.; Liu, K.G. Mycoplasma pneumoniae 23S rRNA Gene Mutations and Mechanisms of Macrolide Resistance. Labmedicine 2013, 44, 63–68. [Google Scholar]
- Aktan, Y.; Tan, S.; Icgen, B. Characterization of lead-resistant river isolate Enterococcus faecalis and assessment of its multiple metal and antibiotic resistance. Environ. Monit. Assess. 2013, 185, 5285–5293. [Google Scholar] [CrossRef]
- Hellweger, F.L. Simple model of tetracycline antibiotic resistance in aquatic environment: Accounting for metal coselection. J. Environ. Eng.-ASCE 2013, 139, 913–921. [Google Scholar] [CrossRef]
- My, N.H.; Hirao, H.; Van, D.U.; Morokuma, K. Computational studies of bacterial resistance to beta-lactam antibiotics: Mechanism of covalent inhibition of the penicillin-binding protein 2a (PBP2a). J. Chem. Inf. Model. 2011, 51, 3226–3234. [Google Scholar] [CrossRef]
- Yuan, W.C.; Hu, Q.W.; Cheng, H.; Shang, W.L.; Liu, N.; Hua, Z.Y.; Zhu, J.M.; Hu, Z.; Yuan, J.Z.; Zhang, X.; et al. Cell wall thickening is associated with adaptive resistance to amikacin in methicillin-resistant Staphylococcus aureus clinical isolates. J. Antimicrob. Chemother. 2013, 68, 1089–1096. [Google Scholar] [CrossRef]
- Anaya-Lopez, J.L.; Lopez-Meza, J.E.; Ochoa-Zarzosa, A. Bacterial resistance to cationic antimicrobial peptides. Crit. Rev. Microbiol. 2013, 39, 180–195. [Google Scholar] [CrossRef]
- Majzlik, P.; Strasky, A.; Adam, V.; Nemec, M.; Trnkova, L.; Zehnalek, J.; Hubalek, J.; Provaznik, I.; Kizek, R. Influence of zinc(II) and copper(II) ions on Streptomyces bacteria revealed by electrochemistry. Int. J. Electrochem. Sci. 2011, 6, 2171–2191. [Google Scholar]
- Sobrova, P.; Zehnalek, J.; Adam, V.; Beklova, M.; Kizek, R. The effects on soil/water/plant/animal systems by platinum group elements. Cent. Eur. J. Chem. 2012, 10, 1369–1382. [Google Scholar] [CrossRef]
- Krizkova, S.; Huska, D.; Beklova, M.; Hubalek, J.; Adam, V.; Trnkova, L.; Kizek, R. Protein-based electrochemical biosensor for detection of silver(I) ions. Environ. Toxicol. Chem. 2010, 29, 492–496. [Google Scholar] [CrossRef]
- Pereira, P.M.; Filipe, S.R.; Tomasz, A.; Pinho, M.G. Fluorescence ratio imaging microscopy shows decreased access of vancomycin to cell wall synthetic sites in vancomycin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2007, 51, 3627–3633. [Google Scholar] [CrossRef]
- Summers, A.O. Genetic linkage and horizontal gene transfer, the roots of the antibiotic multi-resistance problem. Anim. Biotechnol. 2006, 17, 125–135. [Google Scholar] [CrossRef]
- Resende, J.A.; Silva, V.L.; Fontes, C.O.; Souza, J.A.; de Oliveira, T.L.R.; Coelho, C.M.; Cesar, D.E.; Diniz, C.G. Multidrug-resistance and toxic metal tolerance of medically important bacteria isolated from an aquaculture system. Microbes Environ. 2012, 27, 449–455. [Google Scholar] [CrossRef]
- Ji, X.L.; Shen, Q.H.; Liu, F.; Ma, J.; Xu, G.; Wang, Y.L.; Wu, M.H. Antibiotic resistance gene abundances associated with antibiotics and heavy metals in animal manures and agricultural soils adjacent to feedlots in Shanghai, China. J. Hazard. Mater. 2012, 235, 178–185. [Google Scholar]
- Malik, A.; Aleem, A. Incidence of metal and antibiotic resistance in Pseudomonas spp. from the river water, agricultural soil irrigated with wastewater and groundwater. Environ. Monit. Assess. 2011, 178, 293–308. [Google Scholar] [CrossRef]
- Adam, V.; Fabrik, I.; Kohoutkova, V.; Babula, P.; Hubalek, J.; Vrba, R.; Trnkova, L.; Kizek, R. Automated electrochemical analyzer as a new tool for detection of thiols. Int. J. Electrochem. Sci. 2010, 5, 429–447. [Google Scholar]
- Adam, V.; Baloun, J.; Fabrik, I.; Trnkova, L.; Kizek, R. An electrochemical detection of metallothioneins at the zeptomole level in nanolitre volumes. Sensors 2008, 8, 2293–2305. [Google Scholar] [CrossRef]
- Adam, V.; Blastik, O.; Krizkova, S.; Lubal, P.; Kukacka, J.; Prusa, R.; Kizek, R. Application of the Brdicka reaction in determination of metallothionein in patients with tumours. Chem. Listy 2008, 102, 51–58. [Google Scholar]
- Maret, W. Fluorescent probes for the structure and function of metallothionein. J. Chromatogr. B 2009, 877, 3378–3383. [Google Scholar] [CrossRef]
- Sobrova, P.; Vyslouzilova, L.; Stepankova, O.; Ryvolova, M.; Anyz, J.; Trnkova, L.; Adam, V.; Hubalek, J.; Kizek, R. Tissue specific electrochemical fingerprinting. PLoS One 2012, 7, e49654. [Google Scholar] [CrossRef]
- Brissonnoel, A.; Trieucuot, P.; Courvalin, P. Mechanisms of action of spiramycin and other macrolides. J. Antimicrob. Chemother. 1988, 22, 13–23. [Google Scholar]
- Jenkins, R.; Burton, N.; Cooper, R. Manuka honey inhibits cell division in methicillin-resistant Staphylococcus aureus. J. Antimicrob. Chemother. 2011, 66, 2536–2542. [Google Scholar] [CrossRef]
- Belley, A.; Harris, R.; Beveridge, T.; Parr, T.; Moeck, G. Ultrastructural effects of oritavancin on methicillin-resistant Staphylococcus aureus and vancomycin-resistant Enterococcus. Antimicrob. Agents Chemother. 2009, 53, 800–804. [Google Scholar] [CrossRef]
- Webster, D.; Rennie, R.P.; Brosnikoff, C.L.; Chui, L.; Brown, C. Methicillin-resistant Staphylococcus aureus with reduced susceptibility to vancomycin in Canada. Diagn. Microbiol. Infect. Dis. 2007, 57, 177–181. [Google Scholar] [CrossRef]
- Rouch, D.A.; Lee, B.T.O.; Morby, A.P. Understanding cellular-responses to toxic agents—A model for mechanism-choice in bacterial metal resistance. J. Ind. Microbiol. 1995, 14, 132–141. [Google Scholar] [CrossRef]
- Trajanovska, S.; Britz, M.L.; Bhave, M. Detection of heavy metal ion resistance genes in Gram-positive and Gram-negative bacteria isolated from a lead-contaminated site. Biodegradation 1997, 8, 113–124. [Google Scholar] [CrossRef]
- Nies, D.H.; Silver, S. Ion efflux systems involved in bacterial metal resistances. J. Ind. Microbiol. 1995, 14, 186–199. [Google Scholar] [CrossRef]
- Nies, D.H. Microbial heavy-metal resistance. Appl. Microbiol. Biotechnol. 1999, 51, 730–750. [Google Scholar] [CrossRef]
- Choudhury, R.; Srivastava, S. Zinc resistance mechanisms in bacteria. Curr. Sci. 2001, 81, 768–775. [Google Scholar]
- Naik, M.M.; Dubey, S.K.; Khanolkar, D.; D’Costa, B. P-type ATPase and MdrL efflux pump-mediated lead and multi-drug resistence in estuarine bacterial isolates. Curr. Sci. 2013, 105, 1366–1372. [Google Scholar]
- Blaszak, M.; Bienkowska, D. Effect of soil pollution on bacterial resistance to lead ions: An experimental approach. Pol. J. Ecol. 2009, 57, 555–560. [Google Scholar]
- Rensing, C.; Sun, Y.; Mitra, B.; Rosen, B.P. Pb(II)-translocating P-type ATPases. J. Biol. Chem. 1998, 273, 32614–32617. [Google Scholar] [CrossRef]
- Levinson, H.S.; Mahler, I.; Blackwelder, P.; Hood, T. Lead resistance and sensitivity in Staphylococcus aureus. FEMS Microbiol. Lett. 1996, 145, 421–425. [Google Scholar] [CrossRef]
- Deb, S.; Ahmed, S.F.; Basu, M. Metal accumulation in cell wall: A possible mechanism of cadmium resistance by Pseudomonas stutzeri. Bull. Environ. Contam. Toxicol. 2013, 90, 323–328. [Google Scholar] [CrossRef]
- Baker-Austin, C.; Wright, M.S.; Stepanauskas, R.; McArthur, J.V. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006, 14, 176–182. [Google Scholar] [CrossRef]
- Chapman, J.S. Disinfectant resistance mechanisms, cross-resistance, and co-resistance. Int. Biodeterior. Biodegrad. 2003, 51, 271–276. [Google Scholar] [CrossRef]
- Jensen, S.O.; Kwong, S.M.; Lyon, B.R.; Firth, N. Evolution of multiple drug resistance in Staphylococci. Microbiol. Aust. 2008, 29, 120–123. [Google Scholar]
- Jensen, S.O.; Lyon, B.R. Genetics of antimicrobial resistance in Staphylococcus aureus. Future Microbiol. 2009, 4, 565–582. [Google Scholar]
- Novick, R.P.; Roth, C. Plasmid-linked resistance to inorganic salts in Staphylococcus aureus. J. Bacteriol. 1968, 95, 1335–1342. [Google Scholar]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef]
- Lacey, R.W.; Chopra, I. Evidence for mutation to streptomycin resistance in clinical strains of Staphylococcus aureus. J. Gen. Microbiol. 1972, 73, 175–180. [Google Scholar] [CrossRef]
- Derbise, A.; de Cespedes, G.; el Solh, N. Nucleotide sequence of the Staphylococcus aureus transposon, Tn5405, carrying aminoglycosides resistance genes. J. Basic Microbiol. 1997, 37, 379–384. [Google Scholar] [CrossRef]
- Singh, V.K.; Xiong, A.M.; Usgaard, T.R.; Chakrabarti, S.; Deora, R.; Misra, T.K.; Jayaswal, R.K. ZntR is an autoregulatory protein and negatively regulates the chromosomal zinc resistance operon znt of Staphylococcus aureus. Mol. Microbiol. 1999, 33, 200–207. [Google Scholar] [CrossRef]
- Yoon, K.P.; Misra, T.K.; Silver, S. Regulation of the cadA cadmium resistance determinant of Staphylococcus aureus plasmid PI258. J. Bacteriol. 1991, 173, 7643–7649. [Google Scholar]
- Yoon, K.P.; Silver, S. A 2nd gene in the Staphylococcus aureus cadA cadmium resistance determinant of plasmid-PI258. J. Bacteriol. 1991, 173, 7636–7642. [Google Scholar]
- Okkeri, J.; Haltia, T. Expression and mutagenesis of ZntA, a zinc-transporting P-type ATPase from Escherichia coli. Biochemistry 1999, 38, 14109–14116. [Google Scholar] [CrossRef]
- Stomeo, F.; Valverde, A.; Pointing, S.B.; McKay, C.P.; Warren-Rhodes, K.A.; Tuffin, M.I.; Seely, M.; Cowan, D.A. Hypolithic and soil microbial community assembly along an aridity gradient in the Namib Desert. Extremophiles 2013, 17, 329–337. [Google Scholar] [CrossRef]
- Xiong, A.M.; Jayaswal, R.K. Molecular characterization of a chromosomal determinant conferring resistance to zinc and cobalt ions in Staphylococcus aureus. J. Bacteriol. 1998, 180, 4024–4029. [Google Scholar]
- Thelwell, C.; Robinson, N.J.; Turner-Cavet, J.S. An SmtB-like repressor from Synechocystis PCC 6803 regulates a zinc exporter. Proc. Natl. Acad. Sci. USA 1998, 95, 10728–10733. [Google Scholar] [CrossRef]
- Cook, W.J.; Kar, S.R.; Taylor, K.B.; Hall, L.M. Crystal structure of the cyanobacterial metallothionein repressor SmtB: A model for metalloregulatory proteins. J. Mol. Biol. 1998, 275, 337–346. [Google Scholar] [CrossRef]
- Shi, W.P.; Dong, J.; Scott, R.A.; Ksenzenko, M.Y.; Rosen, B.P. The role of arsenic thiol interactions in metalloregulation of the ars operon. J. Biol. Chem. 1996, 271, 9291–9297. [Google Scholar]
- Prusa, R.; Kizek, R.; Trnkova, L.; Vacek, J.; Zehnalek, J. Study of relationship between metallothionein and heavy metals by CPSA method. Clin. Chem. 2004, 50, A28–A29. [Google Scholar]
- Vasak, M. Advances in metallothionein structure and functions. J. Trace Elem. Med. Biol. 2005, 19, 13–17. [Google Scholar] [CrossRef]
- Skalickova, S.; Zitka, O.; Nejdl, L.; Krizkova, S.; Sochor, J.; Janu, L.; Ryvolova, M.; Hynek, D.; Zidkova, J.; Zidek, V.; et al. Study of interaction between metallothionein and CdTe quantum dots. Chromatographia 2013, 76, 345–353. [Google Scholar] [CrossRef]
- Huckle, J.W.; Morby, A.P.; Turner, J.S.; Robinson, N.J. Isolation of a prokaryotic metallothionein locus and analysis of transcriptional control by trace-metal ions. Mol. Microbiol. 1993, 7, 177–187. [Google Scholar] [CrossRef]
- Klaassen, C.D.; Liu, J.; Diwan, B.A. Metallothionein protection of cadmium toxicity. Toxicol. Appl. Pharmacol. 2009, 238, 215–220. [Google Scholar] [CrossRef]
- Templeton, D.M.; Cherian, M.G. Toxicological significance of metallothionein. Method Enzymol. 1991, 205, 11–24. [Google Scholar] [CrossRef]
- Sochor, J.; Pohanka, M.; Ruttkay-Nedecky, B.; Zitka, O.; Hynek, D.; Mares, P.; Zeman, L.; Adam, V.; Kizek, R. Effect of selenium in organic and inorganic form on liver, kidney, brain and muscle of Wistar rats. Cent. Eur. J. Chem. 2012, 10, 1442–1451. [Google Scholar] [CrossRef]
- Coyle, P.; Philcox, J.C.; Carey, L.C.; Rofe, A.M. Metallothionein: The multipurpose protein. Cell. Mol. Life Sci. 2002, 59, 627–647. [Google Scholar] [CrossRef]
- Henkel, G.; Krebs, B. Metallothioneins: Zinc, cadmium, mercury, and copper thiolates and selenolates mimicking protein active site features—Structural aspects and biological implications. Chem. Rev. 2004, 104, 801–824. [Google Scholar] [CrossRef]
- Adam, V.; Fabrik, I.; Eckschlager, T.; Stiborova, M.; Trnkova, L.; Kizek, R. Vertebrate metallothioneins as target molecules for analytical techniques. TRAC-Trends Anal. Chem. 2010, 29, 409–418. [Google Scholar] [CrossRef]
- Krizkova, S.; Ryvolova, M.; Hrabeta, J.; Adam, V.; Stiborova, M.; Eckschlager, T.; Kizek, R. Metallothioneins and zinc in cancer diagnosis and therapy. Drug Metab. Rev. 2012, 44, 287–301. [Google Scholar] [CrossRef]
- Naik, M.M.; Shamim, K.; Dubey, S.K. Biological characterization of lead-resistant bacteria to explore role of bacterial metallothionein in lead resistance. Curr. Sci. 2012, 103, 426–429. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Chudobova, D.; Dostalova, S.; Blazkova, I.; Michalek, P.; Ruttkay-Nedecky, B.; Sklenar, M.; Nejdl, L.; Kudr, J.; Gumulec, J.; Tmejova, K.; et al. Effect of Ampicillin, Streptomycin, Penicillin and Tetracycline on Metal Resistant and Non-Resistant Staphylococcus aureus. Int. J. Environ. Res. Public Health 2014, 11, 3233-3255. https://doi.org/10.3390/ijerph110303233
Chudobova D, Dostalova S, Blazkova I, Michalek P, Ruttkay-Nedecky B, Sklenar M, Nejdl L, Kudr J, Gumulec J, Tmejova K, et al. Effect of Ampicillin, Streptomycin, Penicillin and Tetracycline on Metal Resistant and Non-Resistant Staphylococcus aureus. International Journal of Environmental Research and Public Health. 2014; 11(3):3233-3255. https://doi.org/10.3390/ijerph110303233
Chicago/Turabian StyleChudobova, Dagmar, Simona Dostalova, Iva Blazkova, Petr Michalek, Branislav Ruttkay-Nedecky, Matej Sklenar, Lukas Nejdl, Jiri Kudr, Jaromir Gumulec, Katerina Tmejova, and et al. 2014. "Effect of Ampicillin, Streptomycin, Penicillin and Tetracycline on Metal Resistant and Non-Resistant Staphylococcus aureus" International Journal of Environmental Research and Public Health 11, no. 3: 3233-3255. https://doi.org/10.3390/ijerph110303233
APA StyleChudobova, D., Dostalova, S., Blazkova, I., Michalek, P., Ruttkay-Nedecky, B., Sklenar, M., Nejdl, L., Kudr, J., Gumulec, J., Tmejova, K., Konecna, M., Vaculovicova, M., Hynek, D., Masarik, M., Kynicky, J., Kizek, R., & Adam, V. (2014). Effect of Ampicillin, Streptomycin, Penicillin and Tetracycline on Metal Resistant and Non-Resistant Staphylococcus aureus. International Journal of Environmental Research and Public Health, 11(3), 3233-3255. https://doi.org/10.3390/ijerph110303233