Neural Tube Defects, Folic Acid and Methylation

Abstract
:1. Introduction
2. Neurulation and Neural Tube Defects
3. NTD Prevention via Folate
3.1. The Folate Story
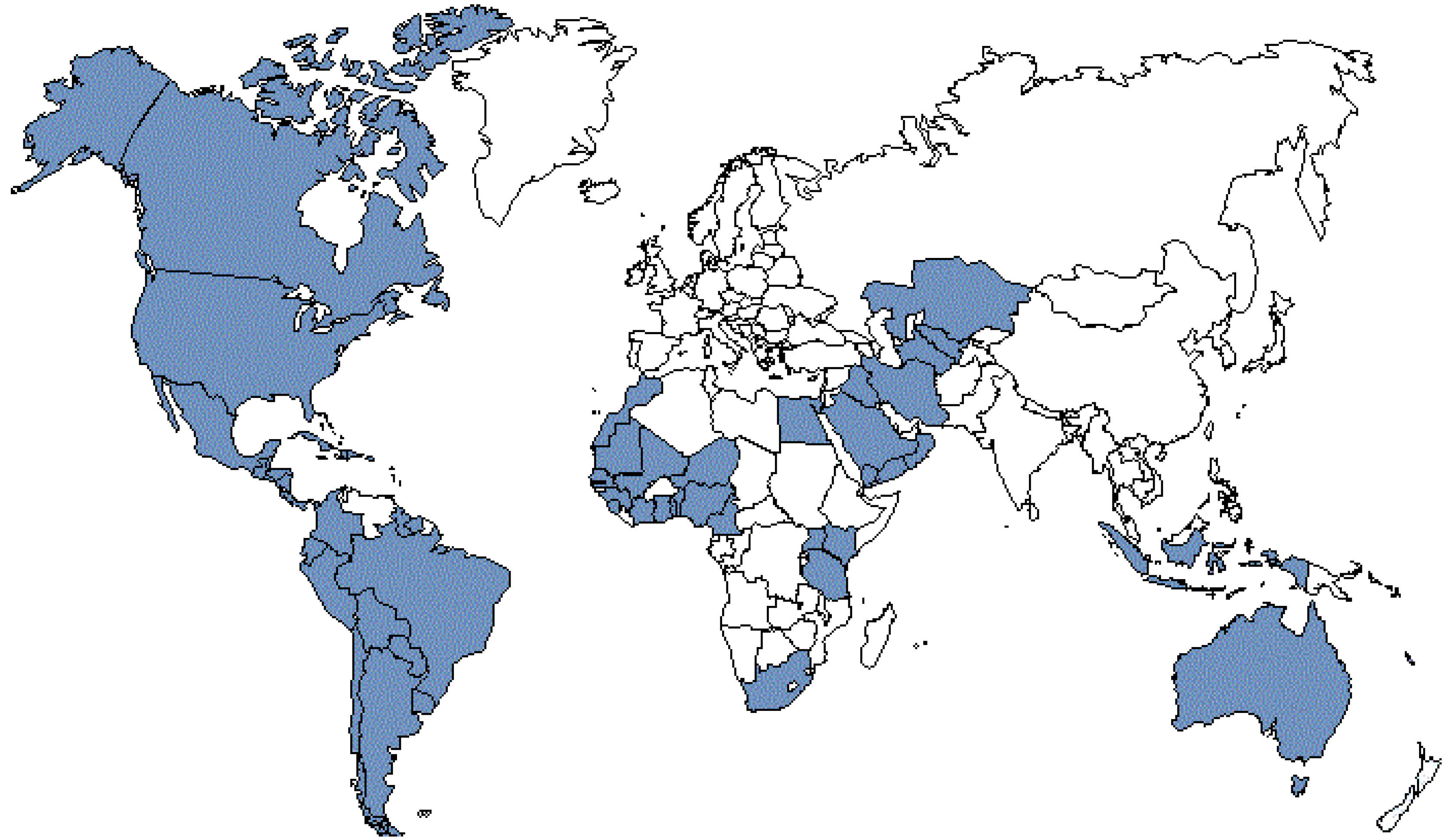
3.2. National Programs for NTDs Prevention


{kind=link}
{kind=link}
{kind=link}
{kind=link}
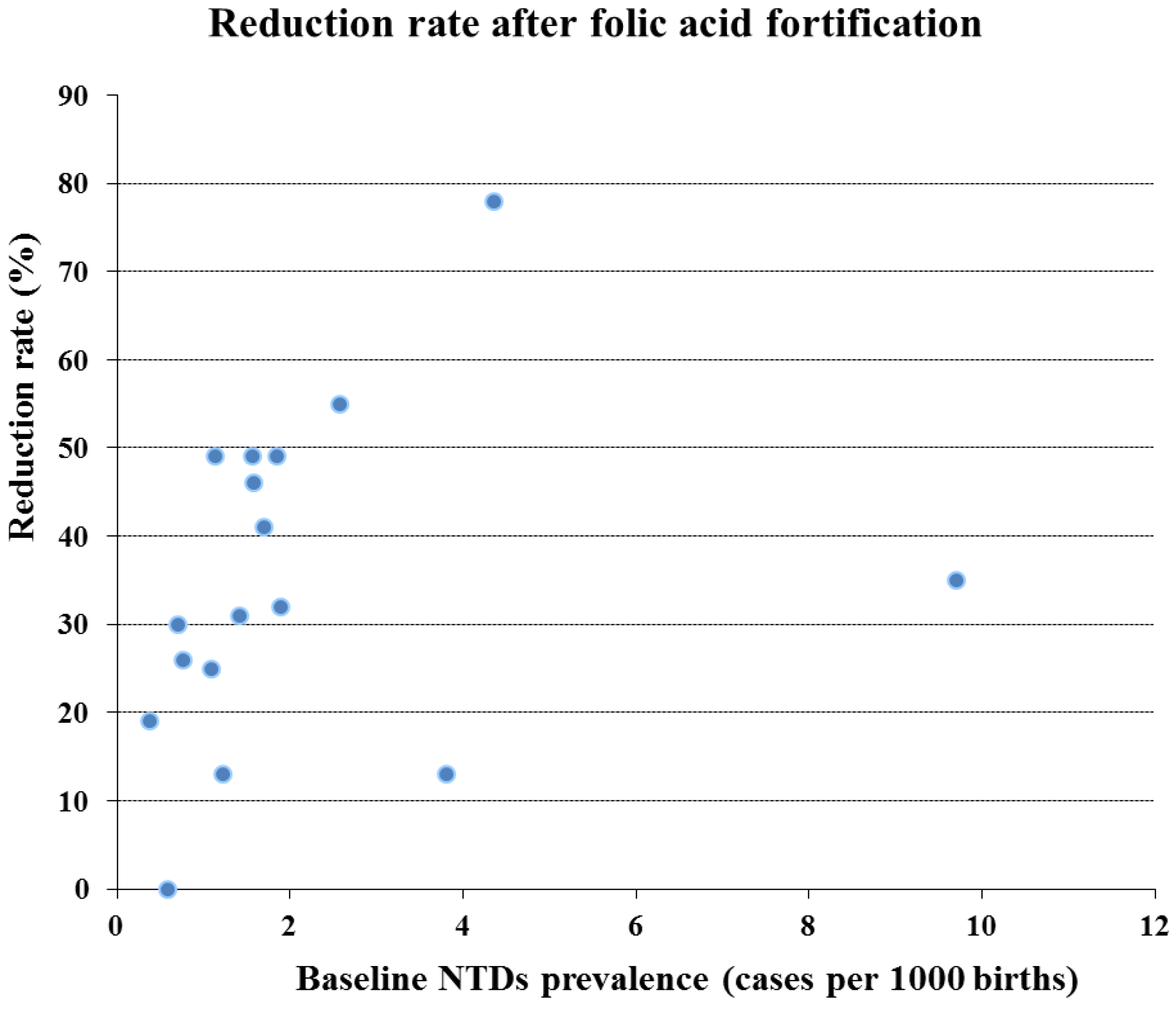
| Reference | Area/State | Period studied | Total NTD prevalence rate (/1,000 births) | % Reduction | ||
|---|---|---|---|---|---|---|
| Before fortification | After fortification | Before fortification | After fortification | |||
| Ray et al. [72] | Ontario | 1994–1997 | 1998–2000 | 1.13 | 0.58 | 49 |
| Honein et al. [73] | USA | 1995–1996 | 1998–1999 | 0.38 | 0.31 | 19 |
| Persad et al. [74] | Nova Scotia | 1991–1997 | 1998–2000 | 2.58 | 1.17 | 55 |
| Williams et al. [75] | USA | 1995–1996 | 1998–1999 | 0.76 | 0.56 | 26 |
| De Wals et al. [76] | Quebec | 1992–1997 | 1998–2000 | 1.89 | 1.28 | 32 |
| Palomaki et al. [77] | Maine | 1993–1996 | 1998–2000 | 1.23 | 1.07 | 13 |
| Lambert-Messerlian et al. [78] | Rhode Island | 1991–1996 | 1998–2000 | 3.8 | 3.3 | 13 |
| Simmons et al. [79] | Arkansas | 1993–1995 | 1999–2000 | 1.09 | 0.82 | 25 |
| Liu et al. [64] | Newfoudland | 1991–1997 | 1998–2001 | 4.36 | 0.96 | 78 |
| Chen et al. [80] | Costa Rica | 1996–1998 | 1999–2000 | 9.7 | 6.3 | 35 |
| Hertrampf et al. [81] | Chile | 1999–2000 | 2001–2002 | 1.70 | 1.01 | 41 |
| Lopez-Camelo et al. [82] | Chile | 1982–1991 | 2001–2002 | 1.57 | 0.80 | 49 |
| Canfield et al. [83] | USA | 1995–1996 | 1999–2000 | 0.71 | 0.5 | 30 |
| De Wals et al. [69] | Canada | 1993–1997 | 2000–2002 | 1.58 | 0.86 | 46 |
| Chen et al. [84] | California | 1989–1996 | 1998–2003 | 0.59 | 0.70 | No decline |
| Sayed et al. [85] | South Africa | 2003–2004 | 2004–2005 | 1.41 | 0.98 | 31 |
| Amarin et al. [86] | Jordan | 2000–2001 | 2005–2006 | 1.85 | 1.07 | 49 |

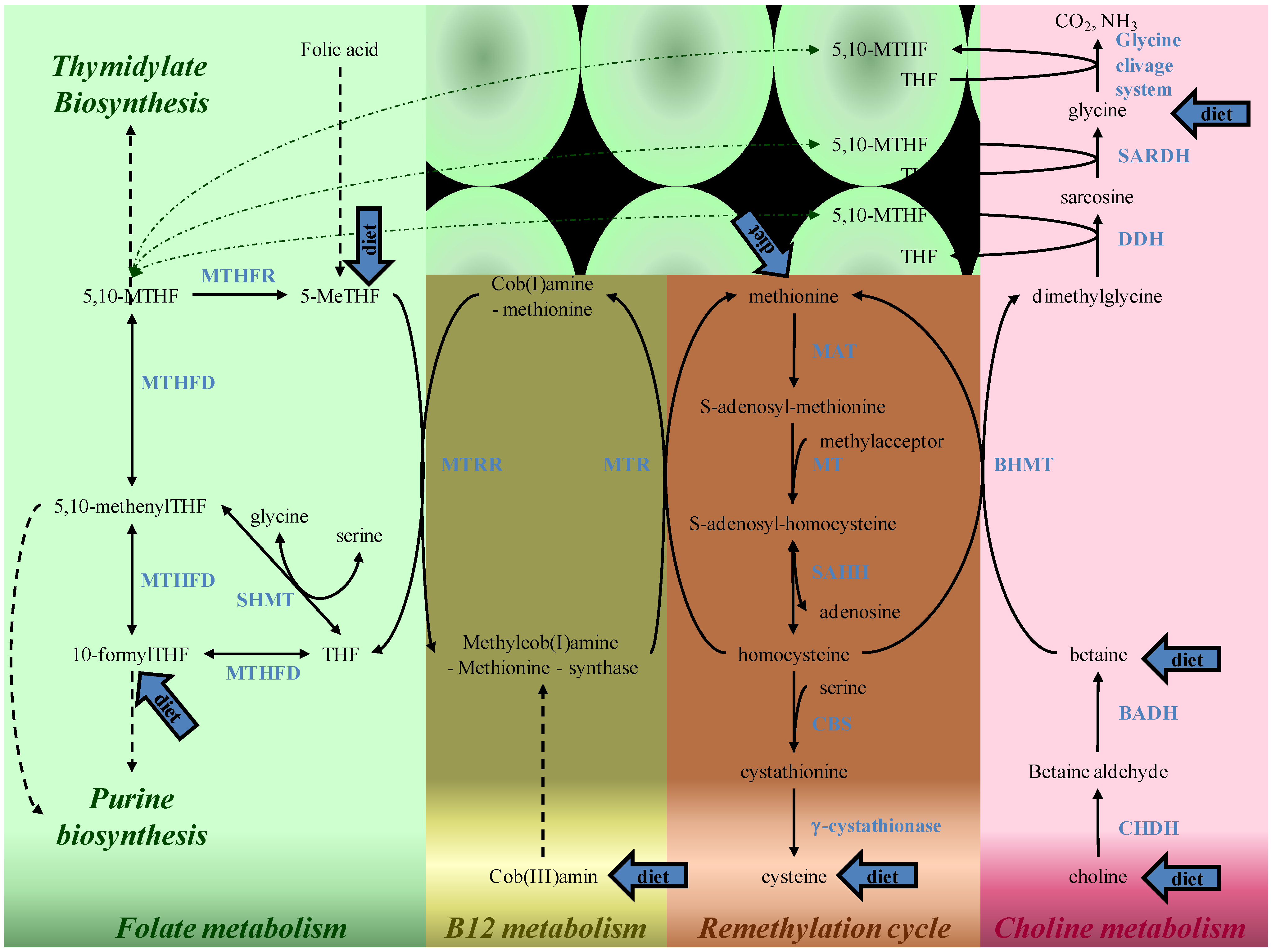
4. Folate and Methylation Metabolism
5. Putative Mechanisms of Folates Action

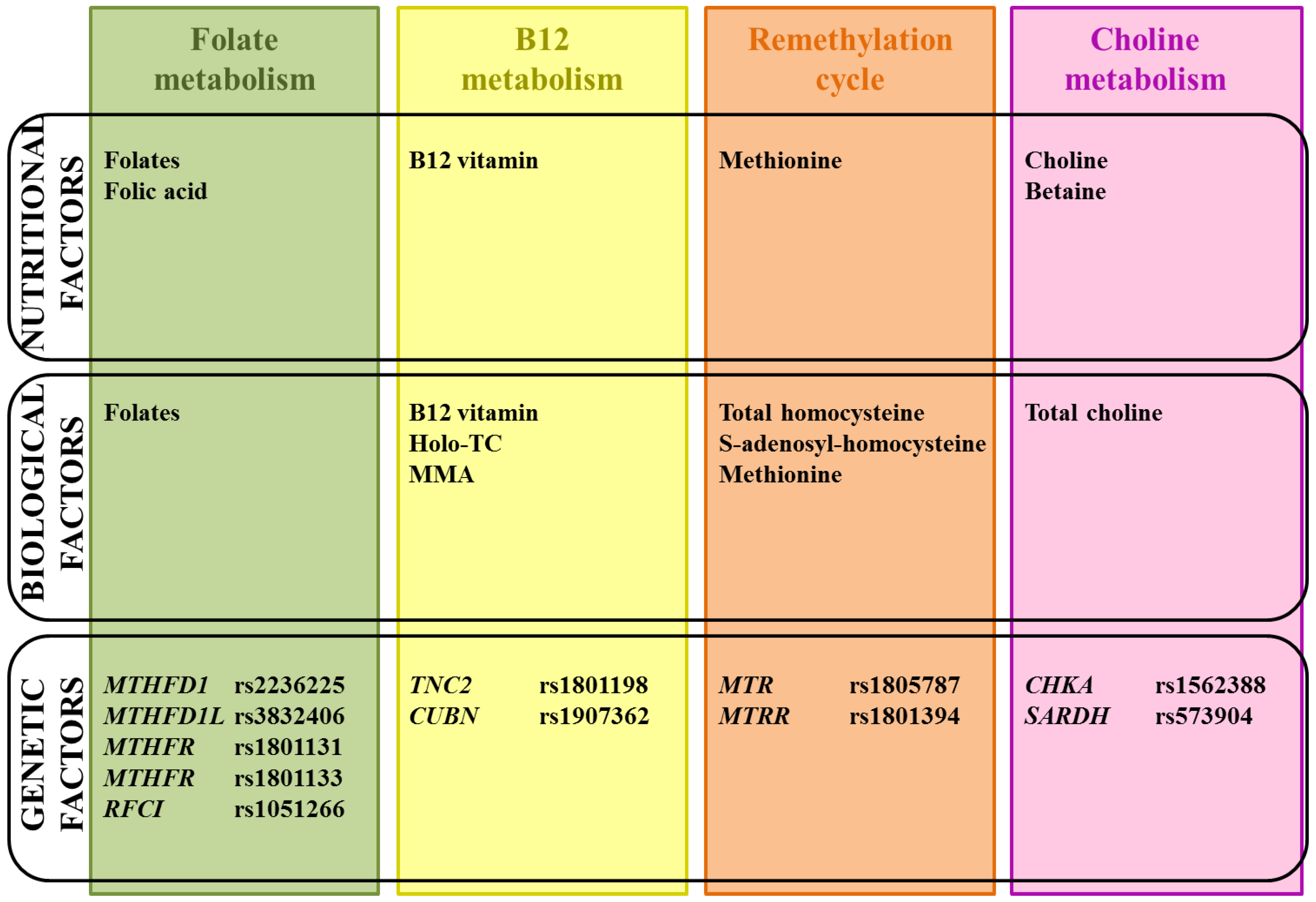
6. Arguments for the Implication of Homocysteine, Vitamin B12 and Choline Pathways
6.1. Biological and Nutritional Aspects
6.1.1. Homocysteine
6.1.2. Vitamin B12
6.1.3. Choline and Betaine
6.2. Genetic Aspects
7. Conclusions
| Gene | Polymorphism(s) studied | Population studied | Effect on NTD risk | Reference | |
|---|---|---|---|---|---|
| B12 Pathway | CUBN | rs1907362 | Children | Decreased risk | Franke et al. [217] |
| rs4748353 | Children | NS | Franke et al. [217] | ||
| TCN2 | rs1801198 | Children | NS | Guéant-Rodriguez et al.; Afman et al.; Swanson et al. [218,219,220] | |
| Mothers | NS | Candito et al.; Afman et al.; Swanson et al. [37,218,219,220] | |||
| Increased risk | Pietrzyk et al. [221] | ||||
| rs96067256 rs4820889 rs9621049 rs1131603 | Mothers and Children | NS | Afman et al.; Swanson et al. [218,219,220] | ||
| Choline pathway | BHMT | rs3733890 | Children | NS | Zhu et al.; Morin et al. [222,223] |
| Mothers | NS | Morin et al. [222] | |||
| BHMT2 | rs626105 | Children | NS | Zhu et al. [223] | |
| CHKA | hCV1562393C | Children | NS | Enaw et al. [199] | |
| rs1562388 | Children | Decreased risk | Enaw et al. [199] | ||
| PCYT1A | rs3772109 | Children | NS | Enaw et al. [199] | |
| SARDH | rs573904 | Children | Increased risk | Franke et al. [217] | |
| Folate pathway | DHFR | 19bp DEL | Fathers | NS | Johnson et al. [224] |
| Children | NS | Johnson et al.; Van der Linden et al.; Doudney et al. [224,225,226] | |||
| Mothers | NS | Johnson et al.; Van der Linden et al. [224,226] | |||
| Receptor Folate | rs651646 | Fathers | NS | Oleary et al. [227] | |
| Children | NS | Oleary et al. [227] | |||
| Mothers | NS | Oleary et al. [227] | |||
| GCPII | rs61886492 | Children | NS | Relton et al. [228] | |
| Mothers | NS | Morin et al.; Relton et al. [228,229] | |||
| MTHFD1 | rs2236225 | Fathers | NS | Brody et al.; De Marco et al. [230,231] | |
| Children | NS | Brody et al.; Hol et al.; Van der Linden et al.; Doudney et al. [225,226,230,232] | |||
| Increased risk | De Marco et al. [231] | ||||
| Mothers | Increased risk | Brody et al.; De Marco et al.; Parle McDermott et al. [230,231,233] | |||
| NS | Van der Linden et al. [234] | ||||
| rs1950902 | Fathers | NS | Brody et al. [230] | ||
| Children | NS | Brody et al. [230] | |||
| Mothers | NS | Brody et al. [230] | |||
| MTHFD1L | rs3832406 | Children and mothers | Increased risk with “allele 1” | Parle Mc Dermott et al. [235] | |
| MTHFR | rs2066462 | Fathers | NS | Morrison et al. [236] | |
| Children | NS | Morrison et al. [236] | |||
| Mothers | NS | Morrison et al. [236] | |||
| rs1801131 | Fathers | NS | Van der Put et al.; Volcik et al.; De Marco et al.; Parle McDermott et al.; Boduroglu et al.; Grandone et al.; Gonzales-Herrera et al.; De Marco et al. [163,216,237,238,239,240,241,242] | ||
| Increased risk | De Marco et al. [239] | ||||
| Children | NS | Van der Put et al.; Volcik et al.; Parle McDermott et al.; Relton et al.; Boduroglu et al.; Felix et al.; Grandone et al.; Gonzales-Herrera et al.; Doudney et al.; Behunova et al.; De Marco et al. [162,163,216,228,237,238,240,241,242,243] | |||
| Increased risk | De Marco et al. [239] | ||||
| Mothers | NS | Van der Put et al.; Volcik et al.; Parle McDermott et al.; Gutierrez-Revilla et al.; Relton et al.; Boduroglu et al.; Grandone et al.; Felix et al.; De Marco et al. [162,163,216,228,237,238,241,242,244] | |||
| Increased risk | De Marco et al.; Gonzales-Herrera et al.; Candito et al. [37,239,240] | ||||
| MTHFR | rs1801133 | Fathers | NS | Papetrou et al.; Van der Put et al.; Morrison et al.; Van der Put et al.; Volcik et al.; Parle McDermott et al.; Rampersaud et al.; Boduroglu et al.; Grandone et al. [163,203,216,236,237,241,242,245,246] | |
| Increased risk | Blom et al. * [12] | ||||
| Children | NS | Papetrou et al.; Mornet et al.; Koch et al.; Morrison et al.; Shaw et al.;Van der Put et al. *; Christensen et al.; Volcik et al.; Gonzales-Herrera et al.; Gutierrez-Revilla et al.; Felix et al.; Boduroglu et al.; Doudney et al.; Behunova et al. [38,162,214,216,225,236,237,242,243,244,245,247,248,249] | |||
| Increased risk | Whitehead et al.; Kirke et al., Ou et al.; Van der Put et al. *; De Franchis et al.; Shields et al.; Botto et al. *; Johanning et al.; Parle McDermott et al.; Johanning et al.; Pietrzyk et al.; Rampersaud et al.; Kirke et al.; Relton et al.; Blom et al. *; Grandone et al.; Wenstrom et al. [12,41,160,163,205,206,241,246,248,250,251,252,253254255256257] | ||||
| Decreased risk | Guéant-Rodriguez et al. [219] | ||||
| Mothers | NS | Papetrou et al.; Morrison et al.; Christensen et al.; Ubbink et al.; Lucock et al.; Volcik et al.; Gonzales-Herrera et al.; Parle McDermott et al., Rampersaud et al.; Gutierrez-Revilla et al.; Relton et al.; Boduroglu et al.; Felix et al.; Grandone et al.; Candito et al.; Shang et al.; Davalos et al. [37,38,162,163,191,215,216,228,236,237,241,244,245,246,247,258,259] | |||
| Increased risk | Van der Put et al. *; Van der Put et al.; Botto et al. *; Martinez et al.; Pietrzyk et al.; Yan et al. *; Blom et al. *; Amorim et al. *; Richter et al. [12,204,205,206,207,252,255,260,261] | ||||
| RFCI | 61bp repeat | Fathers | NS | Oleary et al. [262] | |
| Children | NS | Oleary et al. [262] | |||
| Mothers | NS | Oleary et al. [262] | |||
| rs1051266 | Fathers | NS | De Marco et al.; Viera et al.; De Marco et al.; Pei et al.; Oleary et al. [209,238,262,263,264] | ||
| Children | NS | De Marco et al.; Shaw et al.; Oleary et al.; Doudney et al.; Wang et al. *; Relton et al. [213,225,228,238,262,263,264,265] | |||
| Increased risk | De Marco et al.; Pei et al. [209,263] | ||||
| Decreased risk | Franke et al. [217] | ||||
| Mothers | NS | De Marco et al.; De Marco et al.; Morin et al.; Oleary et al.; Wang et al. *; Relton et al. [209,228,229,238,262,265] | |||
| Increased risk | Pei et al.; Shang et al. [215263] | ||||
| SHMT | rs1979277 | Children | NS | Heil et al.; Relton et al. [41266] | |
| Mothers | NS | Heil et al.; Relton et al. [41266] | |||
| delTCTT1721_1724 | Children | NS | Heil et al. [266] | ||
| Mothers | NS | Heil et al. [266] | |||
| Remethylation pathway | MTR | rs1805087 | Fathers | NS | Morrison et al.; Oleary et al.; De Marco et al. [236,239,267] |
| Children | NS | Van der Put et al.; Morrison et al.; Shaw et al.; Johanning et al.; Zhu et al., Oleary et al.; Doudney et al.; De Marco et al.; Morrison et al. [225,238,251,267,268,269,270,271] | |||
| Decreased risk | Christensen et al. [38] | ||||
| Increased risk | Guéant-Rodriguez et al. [219] | ||||
| Mothers | NS | Van der Put et al.; Morrison et al.; Christensen et al.; Doolin et al.; Zhu et al.; Oleary et al.; Candito et al.; Ouyang et al. *; De Marco et al. [37,38,236,239,267,270,271,272,273] | |||
| MTRR | rs162036 | Fathers | NS | Oleary et al. [267] | |
| Children | NS | Oleary et al. [267] | |||
| Mothers | NS | Oleary et al. [267] | |||
| rs1532268 | Fathers | NS | Oleary et al. [267] | ||
| Children | NS | Oleary et al. [267] | |||
| Mothers | NS | Oleary et al. [267] | |||
| rs1801394 | Fathers | Increased risk | Oleary et al. [267] | ||
| Children | NS | Wilson et al.; Oleary et al.; Van der Linden et al.; Doudney et al. [211214225267] | |||
| Increased risk | Pietrzyk et al.; Zhu et al.; Relton et al. [228255271] | ||||
| Mothers | NS | Wilson et al.; Relton et al.; Oleary et al. [211228267] | |||
| Increased risk | Doolin et al.; Pietrzyk et al.; Zhu et al.; Van der Linden et al.; Candito et al.; Ouyang et al. *, Franke et al. [37,212,217,255,271,272,273] |

Acknowledgments
Conflicts of Interest
References
- Au, K.S.; Ashley-Koch, A.; Northrup, H. Epidemiologic and genetic aspects of spina bifida and other neural tube defects. Dev. Disabil. Res. Rev. 2010, 16, 6–15. [Google Scholar] [CrossRef]
- Northrup, H.; Volcik, K.A. Spina bifida and other neural tube defects. Curr. Probl. Pediatr. 2000, 30, 313–332. [Google Scholar]
- Sadler, T.W. Embryology of neural tube development. Am. J. Med. Genet. C Semin. Med. Genet. 2005, 135C, 2–8. [Google Scholar] [CrossRef]
- Thompson, D.N. Postnatal management and outcome for neural tube defects including spina bifida and encephalocoeles. Prenat. Diagn. 2009, 29, 412–419. [Google Scholar] [CrossRef]
- Waitzman, N.J.; Romano, P.S.; Scheffler, R.M. Estimates of the economic costs of birth defects. Inquiry 1994, 31, 188–205. [Google Scholar]
- Yi, Y.; Lindemann, M.; Colligs, A.; Snowball, C. Economic burden of neural tube defects and impact of prevention with folic acid: A literature review. Eur. J. Pediatr. 2011, 170, 1391–1400. [Google Scholar] [CrossRef]
- Lynch, S.A. Non-multifactorial neural tube defects. Am. J. Med. Genet. C Semin. Med. Genet. 2005, 135C, 69–76. [Google Scholar] [CrossRef]
- Frey, L.; Hauser, W.A. Epidemiology of neural tube defects. Epilepsia 2003, 44, 4–13. [Google Scholar] [CrossRef]
- Greene, N.D.; Stanier, P.; Copp, A.J. Genetics of human neural tube defects. Hum. Mol. Genet. 2009, 18, R113–R129. [Google Scholar] [CrossRef]
- Hibbard, B.M.; Hibbard, E.D.; Jeffcoate, T.N. Folic acid and reproduction. Acta Obstet. Gynecol. Scand. 1965, 44, 375–400. [Google Scholar] [CrossRef]
- Blom, H.J. Folic acid, methylation and neural tube closure in humans. Birth Defects Res. 2009, 85, 295–302. [Google Scholar] [CrossRef]
- Blom, H.J.; Shaw, G.M.; den Heijer, M.; Finnell, R.H. Neural tube defects and folate: Case far from closed. Nat. Rev. Neurosci. 2006, 7, 724–731. [Google Scholar] [CrossRef]
- Blom, H.J.; Smulders, Y. Overview of homocysteine and folate metabolism. With special references to cardiovascular disease and neural tube defects. J. Inherit. Metab. Dis. 2011, 34, 75–81. [Google Scholar] [CrossRef]
- Botto, L.D.; Moore, C.A.; Khoury, M.J.; Erickson, J.D. Neural-tube defects. N. Engl. J. Med. 1999, 341, 1509–1519. [Google Scholar] [CrossRef]
- De Marco, P.; Merello, E.; Cama, A.; Kibar, Z.; Capra, V. Human neural tube defects: Genetic causes and prevention. Biofactors 2011, 37, 261–268. [Google Scholar] [CrossRef]
- Van der Put, N.M.; van Straaten, H.W.; Trijbels, F.J.; Blom, H.J. Folate, homocysteine and neural tube defects: An overview. Exp. Biol. Med. 2001, 226, 243–270. [Google Scholar]
- Copp, A.J.; Greene, N.D.; Murdoch, J.N. The genetic basis of mammalian neurulation. Nat. Rev. Genet. 2003, 4, 784–793. [Google Scholar] [CrossRef]
- Detrait, E.R.; George, T.M.; Etchevers, H.C.; Gilbert, J.R.; Vekemans, M.; Speer, M.C. Human neural tube defects: Developmental biology, epidemiology, and genetics. Neurotoxicol. Teratol. 2005, 27, 515–524. [Google Scholar] [CrossRef]
- Wallingford, J.B. Neural tube closure and neural tube defects: Studies in animal models reveal known knowns and known unknowns. Am. J. Med. Genet. C Semin. Med. Genet. 2005, 135C, 59–68. [Google Scholar] [CrossRef]
- Copp, A.J. Neurulation in the cranial region—Normal and abnormal. J. Anat. 2005, 207, 623–635. [Google Scholar] [CrossRef]
- Padmanabhan, R. Etiology, pathogenesis and prevention of neural tube defects. Congenit. Anom. 2006, 46, 55–67. [Google Scholar] [CrossRef]
- Michie, C.A. Neural tube defects in 18th century. Lancet 1991, 337, 504. [Google Scholar] [CrossRef]
- Smithells, R.W.; Sheppard, S.; Schorah, C.J. Vitamin dificiencies and neural tube defects. Arch. Dis. Child. 1976, 51, 944–950. [Google Scholar] [CrossRef]
- Emery, A.E.; Timson, J.; Watson-Williams, E.J. Pathogenesis of spina bifida. Lancet 1969, 2, 909–910. [Google Scholar]
- Hibbard, B.M. The role of folic acid in pregnancy; With particular reference to anaemia, abruption and abortion. J. Obstet. Gynaecol. Br. Commonw. 1964, 71, 529–542. [Google Scholar] [CrossRef]
- Smithells, R.W. Multivitamins for the prevention of neural tube defects. How convincing is the evidence? Drugs 1989, 38, 849–854. [Google Scholar] [CrossRef]
- Smithells, R.W.; Nevin, N.C.; Seller, M.J.; Sheppard, S.; Harris, R.; Read, A.P.; Fielding, D.W.; Walker, S.; Schorah, C.J.; Wild, J. Further experience of vitamin supplementation for prevention of neural tube defect recurrences. Lancet 1983, 1, 1027–1031. [Google Scholar]
- Smithells, R.W.; Sheppard, S. Possible prevention of neural-tube defects by periconceptional vitamin supplementation. Lancet 1980, 1, 647. [Google Scholar] [CrossRef]
- Smithells, R.W.; Sheppard, S.; Schorah, C.J.; Seller, M.J.; Nevin, N.C.; Harris, R.; Read, A.P.; Fielding, D.W. Apparent prevention of neural tube defects by periconceptional vitamin supplementation. Arch. Dis. Child. 1981, 56, 911–918. [Google Scholar] [CrossRef]
- Economides, D.L.; Ferguson, J.; Mackenzie, I.Z.; Darley, J.; Ware, I.I.; Holmes-Siedle, M. Folate and vitamin B12 concentrations in maternal and fetal blood, and amniotic fluid in second trimester pregnancies complicated by neural tube defects. Br. J. Obstet. Gynaecol. 1992, 99, 23–25. [Google Scholar]
- Gardiki-Kouidou, P.; Seller, M.J. Amniotic fluid folate, vitamin B12 and transcobalamins in neural tube defects. Clin. Genet. 1988, 33, 441–448. [Google Scholar] [CrossRef]
- Molloy, A.M.; Kirke, P.; Hillary, I.; Weir, D.G.; Scott, J.M. Maternal serum folate and vitamin B12 concentrations in pregnancies associated with neural tube defects. Arch. Dis. Child. 1985, 60, 660–665. [Google Scholar] [CrossRef]
- Schorah, C.J.; Smithells, R.W.; Scott, J. Vitamin B12 and anencephaly. Lancet 1980, 1, 880. [Google Scholar] [CrossRef]
- Steegers-Theunissen, R.P.; Boers, G.H.; Blom, H.J.; Nijhuis, J.G.; Thomas, C.M.; Borm, G.F.; Eskes, T.K. Neural tube defects and elevated homocysteine levels in amniotic fluid. Am. J. Obstet. Gynecol. 1995, 172, 1436–1441. [Google Scholar] [CrossRef]
- Steen, M.T.; Boddie, A.M.; Fisher, A.J.; Macmahon, W.; Saxe, D.; Sullivan, K.M.; Dembure, P.P.; Elsas, L.J. Neural-tube defects are associated with low concentrations of cobalamin (vitamin B12) in amniotic fluid. Prenat. Diagn. 1998, 18, 545–555. [Google Scholar] [CrossRef]
- Weekes, E.W.; Tamura, T.; Davis, R.O.; Birch, R.; Vaughn, W.H.; Franklin, J.C.; Barganier, C.; Cosper, P.; Finley, S.C.; Finley, W.H. Nutrient levels in amniotic fluid from women with normal and neural tube defect pregnancies. Biol. Neonate 1992, 61, 226–231. [Google Scholar] [CrossRef]
- Candito, M.; Rivet, R.; Herbeth, B.; Boisson, C.; Rudigoz, R.C.; Luton, D.; Journel, H.; Oury, J.F.; Roux, F.; Saura, R.; et al. Nutritional and genetic determinants of vitamin B and homocysteine metabolisms in neural tube defects: A multicenter case-control study. Am. J. Med. Genet. 2008, 146A, 1128–1133. [Google Scholar] [CrossRef]
- Christensen, B.; Arbour, L.; Tran, P.; Leclerc, D.; Sabbaghian, N.; Platt, R.; Gilfix, B.M.; Rosenblatt, D.S.; Gravel, R.A.; Forbes, P.; et al. Genetic polymorphisms in methylenetetrahydrofolate reductase and methionine synthase, folate levels in red blood cells, and risk of neural tube defects. Am. J. Med. Genet. 1999, 84, 151–157. [Google Scholar] [CrossRef]
- Wald, N.J.; Hackshaw, A.D.; Stone, R.; Sourial, N.A. Blood folic acid and vitamin B12 in relation to neural tube defects. Br. J. Obstet. Gynaecol. 1996, 103, 319–324. [Google Scholar] [CrossRef]
- Zhao, W.; Mosley, B.S.; Cleves, M.A.; Melnyk, S.; James, S.J.; Hobbs, C.A. Neural tube defects and maternal biomarkers of folate, homocysteine, and glutathione metabolism. Birth Defects Res. 2006, 76, 230–236. [Google Scholar] [CrossRef]
- Relton, C.L.; Wilding, C.S.; Laffling, A.J.; Jonas, P.A.; Burgess, T.; Binks, K.; Tawn, E.J.; Burn, J. Low erythrocyte folate status and polymorphic variation in folate-related genes are associated with risk of neural tube defect pregnancy. Mol. Genet. Metab. 2004, 81, 273–281. [Google Scholar] [CrossRef]
- Dawson, E.B.; Evans, D.R.; Harris, W.A.; van Hook, J.W. Amniotic fluid B12, calcium, and lead levels associated with neural tube defects. Am. J. Perinatol. 1999, 16, 373–378. [Google Scholar] [CrossRef]
- Pitkin, R.M. Folate and neural tube defects. Am. J. Clin. Nutr. 2007, 85, 285S–288S. [Google Scholar]
- Vergel, R.G.; Sanchez, L.R.; Heredero, B.L.; Rodriguez, P.L.; Martinez, A.J. Primary prevention of neural tube defects with folic acid supplementation: Cuban experience. Prenat. Diagn. 1990, 10, 149–152. [Google Scholar] [CrossRef]
- Bower, C.; Stanley, F.J. Dietary folate as a risk factor for neural-tube defects: Evidence from a case-control study in Western Australia. Med. J. Aust. 1989, 150, 613–619. [Google Scholar]
- McMahon, D.M.; Liu, J.; Zhang, H.; Torres, M.E.; Best, R.G. Maternal obesity, folate intake, and neural tube defects in offspring. Birth Defects Res. 2013, 97, 115–122. [Google Scholar] [CrossRef]
- Milunsky, A.; Jick, H.; Jick, S.S.; Bruell, C.L.; MacLaughlin, D.S.; Rothman, K.J.; Willett, W. Multivitamin/folic acid supplementation in early pregnancy reduces the prevalence of neural tube defects. JAMA 1989, 262, 2847–2852. [Google Scholar] [CrossRef]
- Mulinare, J.; Cordero, J.F.; Erickson, J.D.; Berry, R.J. Periconceptional use of multivitamins and the occurrence of neural tube defects. JAMA 1988, 260, 3141–3145. [Google Scholar]
- Shaw, G.M.; O’Malley, C.D.; Wasserman, C.R.; Tolarova, M.M.; Lammer, E.J. Maternal periconceptional use of multivitamins and reduced risk for conotruncal heart defects and limb deficiencies among offspring. Am. J. Med. Genet. 1995, 59, 536–545. [Google Scholar] [CrossRef]
- Shaw, G.M.; Schaffer, D.; Velie, E.M.; Morland, K.; Harris, J.A. Periconceptional vitamin use, dietary folate, and the occurrence of neural tube defects. Epidemiology 1995, 6, 219–226. [Google Scholar] [CrossRef]
- Werler, M.M.; Shapiro, S.; Mitchell, A.A. Periconceptional folic acid exposure and risk of occurrent neural tube defects. JAMA 1993, 269, 1257–1261. [Google Scholar]
- Mills, J.L.; Rhoads, G.G.; Simpson, J.L.; Cunningham, G.C.; Conley, M.R.; Lassman, M.R.; Walden, M.E.; Depp, O.R.; Hoffman, H.J. The absence of a relation between the periconceptional use of vitamins and neural-tube defects. National institute of child health and human development neural tube defects study group. N. Engl. J. Med. 1989, 321, 430–435. [Google Scholar] [CrossRef]
- Wright, M.E. A case-control study of maternal nutrition and neural tube defects in Northern Ireland. Midwifery 1995, 11, 146–152. [Google Scholar] [CrossRef]
- Laurence, K.M.; James, N.; Miller, M.H.; Tennant, G.B.; Campbell, H. Double-blind randomised controlled trial of folate treatment before conception to prevent recurrence of neural-tube defects. Br. Med. J. 1981, 282, 1509–1511. [Google Scholar] [CrossRef]
- Wald, N.J.; Polani, P.E. Neural-tube defects and vitamins: The need for a randomized clinical trial. Br. J. Obstet. Gynaecol. 1984, 91, 516–523. [Google Scholar] [CrossRef]
- Medical Research Council vitamin study group. Prevention of neural tube defects: Results of the medical research council vitamin study. Lancet 1991, 338, 131–137. [CrossRef]
- Czeizel, A.E.; Dudas, I. Prevention of the first occurrence of neural-tube defects by periconceptional vitamin supplementation. N. Engl. J. Med. 1992, 327, 1832–1835. [Google Scholar] [CrossRef]
- Berry, R.J.; Li, Z.; Erickson, J.D.; Li, S.; Moore, C.A.; Wang, H.; Mulinare, J.; Zhao, P.; Wong, L.Y.; Gindler, J.; et al. Prevention of neural-tube defects with folic acid in China. China-U.S. Collaborative project for neural tube defect prevention. N. Engl. J. Med. 1999, 341, 1485–1490. [Google Scholar] [CrossRef]
- U.S. Department of Health and Human Services Public Health Service Centers for Disease Control. Recommendations for the use of folic acid to reduce the number of cases of spina bifida and other neural tube defects. MMWR Recomm. Rep. 1992, 41, 1–7.
- Stevenson, R.E.; Allen, W.P.; Pai, G.S.; Best, R.; Seaver, L.H.; Dean, J.; Thompson, S. Decline in prevalence of neural tube defects in a high-risk region of the United States. Pediatrics 2000, 106, 677–683. [Google Scholar]
- Food and Drug Administration. Food standards: Amendment of standards of identity for enriched grain products to require addition of folic acid. Fed. Regist. 1996, 61, 8781–8797.
- Bailey, L.B. Folate and vitamin B12 recommended intakes and status in the United States. Nutr. Rev. 2004, 62, S14–S20. [Google Scholar] [CrossRef]
- U.S. Department of Health and Human Services Public Health Service Centers for Disease Control. Folate status in women of childbearing age, by race/ethnicity—United States, 1999–2000. MMWR Morb. Mortal. Wkly. Rep. 2002, 51, 808–810.
- Liu, S.; West, R.; Randell, E.; Longerich, L.; O’Connor, K.S.; Scott, H.; Crowley, M.; Lam, A.; Prabhakaran, V.; McCourt, C. A comprehensive evaluation of food fortification with folic acid for the primary prevention of neural tube defects. BMC Pregnancy Childbirth 2004, 4, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Ray, J.G. Folic acid food fortification in Canada. Nutr. Rev. 2004, 62, S35–S39. [Google Scholar] [CrossRef]
- Wald, N.J.; Law, M.R.; Morris, J.K.; Wald, D.S. Quantifying the effect of folic acid. Lancet 2001, 358, 2069–2073. [Google Scholar] [CrossRef]
- Jacques, P.F.; Selhub, J.; Bostom, A.G.; Wilson, P.W.; Rosenberg, I.H. The effect of folic acid fortification on plasma folate and total homocysteine concentrations. N. Engl. J. Med. 1999, 340, 1449–1454. [Google Scholar] [CrossRef]
- Rader, J.I. Folic acid fortification, folate status and plasma homocysteine. J. Nutr. 2002, 132, 2466S–2470S. [Google Scholar]
- De Wals, P.; Tairou, F.; van Allen, M.I.; Uh, S.H.; Lowry, R.B.; Sibbald, B.; Evans, J.A.; van den Hof, M.C.; Zimmer, P.; Crowley, M.; et al. Reduction in neural-tube defects after folic acid fortification in Canada. N. Engl. J. Med. 2007, 357, 135–142. [Google Scholar] [CrossRef]
- Williams, L.J.; Rasmussen, S.A.; Flores, A.; Kirby, R.S.; Edmonds, L.D. Decline in the prevalence of spina bifida and anencephaly by race/ethnicity: 1995–2002. Pediatrics 2005, 116, 580–586. [Google Scholar] [CrossRef]
- Flour Fortification Initiative. Country Profile. Available online: http://www.ffinetwork.org/country_profiles/index.php (accessed on 1 July 2013).
- Ray, J.G.; Meier, C.; Vermeulen, M.J.; Boss, S.; Wyatt, P.R.; Cole, D.E. Association of neural tube defects and folic acid food fortification in Canada. Lancet 2002, 360, 2047–2048. [Google Scholar] [CrossRef]
- Honein, M.A.; Paulozzi, L.J.; Mathews, T.J.; Erickson, J.D.; Wong, L.Y. Impact of folic acid fortification of the US food supply on the occurrence of neural tube defects. JAMA 2001, 285, 2981–2986. [Google Scholar]
- Persad, V.L.; van den Hof, M.C.; Dube, J.M.; Zimmer, P. Incidence of open neural tube defects in Nova Scotia after folic acid fortification. CMAJ 2002, 167, 241–245. [Google Scholar]
- Williams, L.J.; Mai, C.T.; Edmonds, L.D.; Shaw, G.M.; Kirby, R.S.; Hobbs, C.A.; Sever, L.E.; Miller, L.A.; Meaney, F.J.; Levitt, M. Prevalence of spina bifida and anencephaly during the transition to mandatory folic acid fortification in the United States. Teratology 2002, 66, 33–39. [Google Scholar] [CrossRef]
- De Wals, P.; Rusen, I.D.; Lee, N.S.; Morin, P.; Niyonsenga, T. Trend in prevalence of neural tube defects in Quebec. Birth Defects Res. 2003, 67, 919–923. [Google Scholar] [CrossRef]
- Palomaki, G.E.; Williams, J.; Haddow, J.E. Comparing the observed and predicted effectiveness of folic acid fortification in preventing neural tube defects. J. Med. Screen. 2003, 10, 52–53. [Google Scholar] [CrossRef]
- Lambert-Messerlian, G.; Halliday, J.; Williams, J.; Cain, R.; Msall, M.E.; Palomaki, G.E.; Canick, J.A. Effect of folic acid fortification on prevalence of neural tube defects in Rhode Island. J. Med. Screen. 2004, 11, 106–107. [Google Scholar] [CrossRef]
- Simmons, C.J.; Mosley, B.S.; Fulton-Bond, C.A.; Hobbs, C.A. Birth defects in Arkansas: Is folic acid fortification making a difference? Birth Defects Res. 2004, 70, 559–564. [Google Scholar] [CrossRef]
- Chen, L.T.; Rivera, M.A. The Costa Rican experience: Reduction of neural tube defects following food fortification programs. Nutr. Rev. 2004, 62, S40–S43. [Google Scholar] [CrossRef]
- Hertrampf, E.; Cortes, F. Folic acid fortification of wheat flour: Chile. Nutr. Rev. 2004, 62, S44–S48. [Google Scholar] [CrossRef]
- Lopez-Camelo, J.S.; Orioli, I.M.; da Graca Dutra, M.; Nazer-Herrera, J.; Rivera, N.; Ojeda, M.E.; Canessa, A.; Wettig, E.; Fontannaz, A.M.; Mellado, C.; et al. Reduction of birth prevalence rates of neural tube defects after folic acid fortification in Chile. Am. J. Med. Genet. A 2005, 135, 120–125. [Google Scholar]
- Canfield, M.A.; Collins, J.S.; Botto, L.D.; Williams, L.J.; Mai, C.T.; Kirby, R.S.; Pearson, K.; Devine, O.; Mulinare, J. Changes in the birth prevalence of selected birth defects after grain fortification with folic acid in the United States: Findings from a multi-state population-based study. Birth Defects Res. 2005, 73, 679–689. [Google Scholar] [CrossRef]
- Chen, B.H.; Carmichael, S.L.; Selvin, S.; Abrams, B.; Shaw, G.M. NTD prevalences in central California before and after folic acid fortification. Birth Defects Res. 2008, 82, 547–552. [Google Scholar] [CrossRef]
- Sayed, A.R.; Bourne, D.; Pattinson, R.; Nixon, J.; Henderson, B. Decline in the prevalence of neural tube defects following folic acid fortification and its cost-benefit in South Africa. Birth Defects Res. 2008, 82, 211–216. [Google Scholar] [CrossRef]
- Amarin, Z.O.; Obeidat, A.Z. Effect of folic acid fortification on the incidence of neural tube defects. Paediatr. Perinat. Epidemiol. 2010, 24, 349–351. [Google Scholar] [CrossRef]
- Romano, P.S.; Waitzman, N.J.; Scheffler, R.M.; Pi, R.D. Folic acid fortification of grain: An economic analysis. Am. J. Public Health 1995, 85, 667–676. [Google Scholar]
- Grosse, S.D.; Ouyang, L.; Collins, J.S.; Green, D.; Dean, J.H.; Stevenson, R.E. Economic evaluation of a neural tube defect recurrence-prevention program. Am. J. Prev. Med. 2008, 35, 572–577. [Google Scholar] [CrossRef]
- EFSA, E.F.S.A. Folic Acid: An Update on Scientific Developments. Available online: http://www.efsa.europa.eu/en/home/publication/efsafolicacid.pdf (accessed on 1 July 2013).
- Brent, R.L.; Oakley, G.P., Jr. The food and drug administration must require the addition of more folic acid in “Enriched” Flour and other grains. Pediatrics 2005, 116, 753–755. [Google Scholar] [CrossRef]
- Brent, R.L.; Oakley, G.P., Jr. Triumph and/or tragedy: The present food and drug administration program of enriching grains with folic acid. Pediatrics 2006, 117, 930–932. [Google Scholar] [CrossRef]
- Brent, R.L.; Oakley, G.P., Jr.; Mattison, D.R. The unnecessary epidemic of folic acid-preventable spina bifida and anencephaly. Pediatrics 2000, 106, 825–827. [Google Scholar] [CrossRef]
- Oakley, G.P., Jr. Let’s increase folic acid fortification and include vitamin B-12. Am. J. Clin. Nutr. 1997, 65, 1889–1890. [Google Scholar]
- Daly, L.E.; Kirke, P.N.; Molloy, A.; Weir, D.G.; Scott, J.M. Folate levels and neural tube defects. Implications for prevention. JAMA 1995, 274, 1698–1702. [Google Scholar]
- Moore, L.L.; Bradlee, M.L.; Singer, M.R.; Rothman, K.J.; Milunsky, A. Folate intake and the risk of neural tube defects: An estimation of dose-response. Epidemiology 2003, 14, 200–205. [Google Scholar]
- Wald, N.J. Folic acid and the prevention of neural-tube defects. N. Engl. J. Med. 2004, 350, 101–103. [Google Scholar] [CrossRef]
- Agopian, A.J.; Tinker, S.C.; Lupo, P.J.; Canfield, M.A.; Mitchell, L.E. Proportion of neural tube defects attributable to known risk factors. Birth Defects Res. 2013, 97, 42–46. [Google Scholar] [CrossRef]
- Heseker, H.B.; Mason, J.B.; Selhub, J.; Rosenberg, I.H.; Jacques, P.F. Not all cases of neural-tube defect can be prevented by increasing the intake of folic acid. Br. J. Nutr. 2009, 102, 173–180. [Google Scholar] [CrossRef]
- Crider, K.S.; Bailey, L.B.; Berry, R.J. Folic acid food fortification-its history, effect, concerns, and future directions. Nutrients 2011, 3, 370–384. [Google Scholar] [CrossRef]
- Mills, J.L. Fortification of foods with folic acid—How much is enough? N. Engl. J. Med. 2000, 342, 1442–1445. [Google Scholar] [CrossRef]
- Osterhues, A.; Holzgreve, W.; Michels, K.B. Shall we put the world on folate? Lancet 2009, 374, 959–961. [Google Scholar] [CrossRef]
- Clarke, R.; Halsey, J.; Lewington, S.; Lonn, E.; Armitage, J.; Manson, J.E.; Bonaa, K.H.; Spence, J.D.; Nygard, O.; Jamison, R.; et al. Effects of lowering homocysteine levels with B vitamins on cardiovascular disease, cancer, and cause-specific mortality: Meta-analysis of 8 randomized trials involving 37 485 individuals. Arch. Intern. Med. 2010, 170, 1622–1631. [Google Scholar] [CrossRef]
- Edwards, B.K.; Ward, E.; Kohler, B.A.; Eheman, C.; Zauber, A.G.; Anderson, R.N.; Jemal, A.; Schymura, M.J.; Lansdorp-Vogelaar, I.; Seeff, L.C.; et al. Annual report to the nation on the status of cancer, 1975–2006, featuring colorectal cancer trends and impact of interventions (risk factors, screening, and treatment) to reduce future rates. Cancer 2010, 116, 544–573. [Google Scholar] [CrossRef]
- Marean, A.; Graf, A.; Zhang, Y.; Niswander, L. Folic acid supplementation can adversely affect murine neural tube closure and embryonic survival. Hum. Mol. Genet. 2011, 20, 3678–3683. [Google Scholar] [CrossRef]
- Pickell, L.; Brown, K.; Li, D.; Wang, X.L.; Deng, L.; Wu, Q.; Selhub, J.; Luo, L.; Jerome-Majewska, L.; Rozen, R. High intake of folic acid disrupts embryonic development in mice. Birth Defects Res. 2011, 91, 8–19. [Google Scholar] [CrossRef]
- Botto, L.D.; Lisi, A.; Robert-Gnansia, E.; Erickson, J.D.; Vollset, S.E.; Mastroiacovo, P.; Botting, B.; Cocchi, G.; de Vigan, C.; de Walle, H.; et al. International retrospective cohort study of neural tube defects in relation to folic acid recommendations: Are the recommendations working? BMJ 2005, 330, 571–576. [Google Scholar] [CrossRef]
- EUROCAT Folic Acid Working Group. Prevention of Neural Tube Defects by Periconceptional Folic Acid Supplementation in Europe. Available online: http://www.eurocat-network.eu/content/Special-Report-NTD-3rdEd-Part-I.pdf (accessed on 1 July 2013).
- Botto, L.D.; Lisi, A.; Bower, C.; Canfield, M.A.; Dattani, N.; de Vigan, C.; de Walle, H.; Erickson, D.J.; Halliday, J.; Irgens, L.M.; et al. Trends of selected malformations in relation to folic acid recommendations and fortification: An international assessment. Birth Defects Res. 2006, 76, 693–705. [Google Scholar] [CrossRef]
- Stoll, C.; Alembik, Y.; Dott, B. Are the recommendations on the prevention of neural tube defects working? Eur. J. Med. Genet. 2006, 49, 461–465. [Google Scholar] [CrossRef]
- Rofail, D.; Colligs, A.; Abetz, L.; Lindemann, M.; Maguire, L. Factors contributing to the success of folic acid public health campaigns. J. Public Health 2012, 34, 90–99. [Google Scholar]
- Bitzer, J.; von Stenglin, A.; Bannemerschult, R. Women’s awareness and periconceptional use of folic acid: Data from a large European survey. Int. J. Womens Health 2013, 5, 201–213. [Google Scholar]
- Michie, C.A.; Narang, I.; Rogers, J.; Robinson, A. Folate supplementation and neural-tube defects. Lancet 2000, 335, 147. [Google Scholar] [CrossRef]
- Ren, A.; Zhang, L.; Li, Z.; Hao, L.; Tian, Y. Awareness and use of folic acid, and blood folate concentrations among pregnant women in northern China—An area with a high prevalence of neural tube defects. Reprod. Toxicol. 2006, 22, 431–436. [Google Scholar] [CrossRef]
- Ray, J.G.; Singh, G.; Burrows, R.F. Evidence for suboptimal use of periconceptional folic acid supplements globally. BJOG 2004, 111, 399–408. [Google Scholar] [CrossRef]
- Abramsky, L.; Dolk, H. Should Europe fortify a staple food with folic acid? Lancet 2007, 369, 641–642. [Google Scholar] [CrossRef]
- Czeizel, A.E. Folic acid: A public-health challenge. Lancet 2006, 367, 2056. [Google Scholar] [CrossRef]
- Mastroiacovo, P.; Leoncini, E. More folic acid, the five questions: Why, who, when, how much, and how. Biofactors 2011, 37, 272–279. [Google Scholar] [CrossRef]
- Taruscio, D.; Carbone, P.; Granata, O.; Baldi, F.; Mantovani, A. Folic acid and primary prevention of birth defects. Biofactors 2011, 37, 280–284. [Google Scholar] [CrossRef]
- Beaudin, A.E.; Stover, P.J. Insights into metabolic mechanisms underlying folate-responsive neural tube defects: A minireview. Birth Defects Res. 2009, 85, 274–284. [Google Scholar] [CrossRef]
- Stover, P.J. Physiology of folate and vitamin B12 in health and disease. Nutr. Rev. 2004, 62, S3–S12. [Google Scholar] [CrossRef]
- Appling, D.R. Compartmentation of folate-mediated one-carbon metabolism in eukaryotes. FASEB J. 1991, 5, 2645–2651. [Google Scholar]
- Stover, P.J.; Field, M.S. Trafficking of intracellular folates. Adv. Nutr. 2011, 2, 325–331. [Google Scholar] [CrossRef]
- Holm, P.I.; Ueland, P.M.; Vollset, S.E.; Midttun, O.; Blom, H.J.; Keijzer, M.B.; den Heijer, M. Betaine and folate status as cooperative determinants of plasma homocysteine in humans. Arterioscler. Thromb. Vasc. Biol. 2005, 25, 379–385. [Google Scholar]
- Imbard, A.; Smulders, Y.M.; Barto, R.; Smith, D.E.; Kok, R.M.; Jakobs, C.; Blom, H.J. Plasma choline and betaine correlate with serum folate, plasma S-adenosyl-methionine and S-adenosyl-homocysteine in healthy volunteers. Clin. Chem. Lab. Med. 2012, 51, 683–692. [Google Scholar]
- Melse-Boonstra, A.; Holm, P.I.; Ueland, P.M.; Olthof, M.; Clarke, R.; Verhoef, P. Betaine concentration as a determinant of fasting total homocysteine concentrations and the effect of folic acid supplementation on betaine concentrations. Am. J. Clin. Nutr. 2005, 81, 1378–1382. [Google Scholar]
- Finkelstein, J.D. Metabolic regulatory properties of S-adenosylmethionine and S-adenosylhomocysteine. Clin. Chem. Lab. Med. 2007, 45, 1694–1699. [Google Scholar] [CrossRef]
- Burren, K.A.; Savery, D.; Massa, V.; Kok, R.M.; Scott, J.M.; Blom, H.J.; Copp, A.J.; Greene, N.D. Gene-environment interactions in the causation of neural tube defects: Folate deficiency increases susceptibility conferred by loss of Pax3 function. Hum. Mol. Genet. 2008, 17, 3675–3685. [Google Scholar] [CrossRef]
- Heid, M.K.; Bills, N.D.; Hinrichs, S.H.; Clifford, A.J. Folate deficiency alone does not produce neural tube defects in mice. J. Nutr. 1992, 122, 888–894. [Google Scholar]
- Rothenberg, S.P.; da Costa, M.P.; Sequeira, J.M.; Cracco, J.; Roberts, J.L.; Weedon, J.; Quadros, E.V. Autoantibodies against folate receptors in women with a pregnancy complicated by a neural-tube defect. N. Engl. J. Med. 2004, 350, 134–142. [Google Scholar] [CrossRef]
- Obican, S.G.; Finnell, R.H.; Mills, J.L.; Shaw, G.M.; Scialli, A.R. Folic acid in early pregnancy: A public health success story. FASEB J. 2010, 24, 4167–4174. [Google Scholar] [CrossRef]
- Piedrahita, J.A.; Oetama, B.; Bennett, G.D.; van Waes, J.; Kamen, B.A.; Richardson, J.; Lacey, S.W.; Anderson, R.G.; Finnell, R.H. Mice lacking the folic acid-binding protein Folbp1 are defective in early embryonic development. Nat. Genet. 1999, 23, 228–232. [Google Scholar] [CrossRef]
- Taparia, S.; Gelineau-van Waes, J.; Rosenquist, T.H.; Finnell, R.H. Importance of folate-homocysteine homeostasis during early embryonic development. Clin. Chem. Lab. Med. 2007, 45, 1717–1727. [Google Scholar]
- Spiegelstein, O.; Mitchell, L.E.; Merriweather, M.Y.; Wicker, N.J.; Zhang, Q.; Lammer, E.J.; Finnell, R.H. Embryonic development of folate binding protein-1 (Folbp1) knockout mice: Effects of the chemical form, dose, and timing of maternal folate supplementation. Dev. Dyn. 2004, 231, 221–231. [Google Scholar] [CrossRef]
- Zhao, R.; Russell, R.G.; Wang, Y.; Liu, L.; Gao, F.; Kneitz, B.; Edelmann, W.; Goldman, I.D. Rescue of embryonic lethality in reduced folate carrier-deficient mice by maternal folic acid supplementation reveals early neonatal failure of hematopoietic organs. J. Biol. Chem. 2001, 276, 10224–10228. [Google Scholar]
- Barber, R.C.; Lammer, E.J.; Shaw, G.M.; Greer, K.A.; Finnell, R.H. The role of folate transport and metabolism in neural tube defect risk. Mol. Genet. Metab. 1999, 66, 1–9. [Google Scholar] [CrossRef]
- Fleming, A.; Copp, A.J. Embryonic folate metabolism and mouse neural tube defects. Science 1998, 280, 2107–2109. [Google Scholar] [CrossRef]
- Wlodarczyk, B.J.; Tang, L.S.; Triplett, A.; Aleman, F.; Finnell, R.H. Spontaneous neural tube defects in splotch mice supplemented with selected micronutrients. Toxicol. Appl. Pharmacol. 2006, 213, 55–63. [Google Scholar] [CrossRef]
- Ichi, S.; Nakazaki, H.; Boshnjaku, V.; Singh, R.M.; Mania-Farnell, B.; Xi, G.; McLone, D.G.; Tomita, T.; Mayanil, C.S. Fetal neural tube stem cells from Pax3 mutant mice proliferate, differentiate, and form synaptic connections when stimulated with folic acid. Stem. Cells Dev. 2012, 21, 321–330. [Google Scholar] [CrossRef]
- Beaudin, A.E.; Abarinov, E.V.; Malysheva, O.; Perry, C.A.; Caudill, M.; Stover, P.J. Dietary folate, but not choline, modifies neural tube defect risk in SHMT1 knockout mice. Am. J. Clin. Nutr. 2012, 95, 109–114. [Google Scholar] [CrossRef]
- Beaudin, A.E.; Abarinov, E.V.; Noden, D.M.; Perry, C.A.; Chu, S.; Stabler, S.P.; Allen, R.H.; Stover, P.J. SHMT1 and de novo thymidylate biosynthesis underlie folate-responsive neural tube defects in mice. Am. J. Clin. Nutr. 2011, 93, 789–798. [Google Scholar] [CrossRef]
- Christensen, K.E.; Deng, L.; Leung, K.Y.; Arning, E.; Bottiglieri, T.; Malysheva, O.V.; Caudill, M.A.; Krupenko, N.I.; Greene, N.D.; Jerome-Majewska, L.; et al. A novel mouse model for genetic variation in 10-formyltetrahydrofolate synthetase exhibits disturbed purine synthesis with impacts on pregnancy and embryonic development. Hum. Mol. Genet. 2013, 22, 3705–3719. [Google Scholar] [CrossRef]
- Barbera, J.P.; Rodriguez, T.A.; Greene, N.D.; Weninger, W.J.; Simeone, A.; Copp, A.J.; Beddington, R.S.; Dunwoodie, S. Folic acid prevents exencephaly in Cited2 deficient mice. Hum. Mol. Genet. 2002, 11, 283–293. [Google Scholar] [CrossRef]
- Dunlevy, L.P.; Burren, K.A.; Chitty, L.S.; Copp, A.J.; Greene, N.D. Excess methionine suppresses the methylation cycle and inhibits neural tube closure in mouse embryos. FEBS Lett. 2006, 580, 2803–2807. [Google Scholar] [CrossRef]
- Dunlevy, L.P.; Burren, K.A.; Mills, K.; Chitty, L.S.; Copp, A.J.; Greene, N.D. Integrity of the methylation cycle is essential for mammalian neural tube closure. Birth Defects Res. 2006, 76, 544–552. [Google Scholar] [CrossRef]
- Afman, L.A.; Blom, H.J.; Drittij, M.J.; Brouns, M.R.; van Straaten, H.W. Inhibition of transmethylation disturbs neurulation in chick embryos. Brain Res. Dev. Brain Res. 2005, 158, 59–65. [Google Scholar] [CrossRef]
- Essien, F.B.; Wannberg, S.L. Methionine but not folinic acid or vitamin B-12 alters the frequency of neural tube defects in axd mutant mice. J. Nutr. 1993, 123, 27–34. [Google Scholar]
- Narisawa, A.; Komatsuzaki, S.; Kikuchi, A.; Niihori, T.; Aoki, Y.; Fujiwara, K.; Tanemura, M.; Hata, A.; Suzuki, Y.; Relton, C.L.; et al. Mutations in genes encoding the glycine cleavage system predispose to neural tube defects in mice and humans. Hum. Mol. Genet. 2012, 21, 1496–1503. [Google Scholar] [CrossRef]
- Greene, N.D.; Stanier, P.; Moore, G.E. The emerging role of epigenetic mechanisms in the etiology of neural tube defects. Epigenetics 2011, 6, 875–883. [Google Scholar] [CrossRef]
- Chang, H.; Zhang, T.; Zhang, Z.; Bao, R.; Fu, C.; Wang, Z.; Bao, Y.; Li, Y.; Wu, L.; Zheng, X.; et al. Tissue-specific distribution of aberrant DNA methylation associated with maternal low-folate status in human neural tube defects. J. Nutr. Biochem. 2011, 22, 1172–1177. [Google Scholar] [CrossRef]
- Okano, M.; Bell, D.W.; Haber, D.A.; Li, E. DNA methyltransferases DNMT3a and DNMT3b are essential for de novo methylation and mammalian development. Cell 1999, 99, 247–257. [Google Scholar] [CrossRef]
- Van der Linden, I.J.; Heil, S.G.; van Egmont Petersen, M.; van Straaten, H.W.; den Heijer, M.; Blom, H.J. Inhibition of methylation and changes in gene expression in relation to neural tube defects. Birth Defects Res. 2008, 82, 676–683. [Google Scholar] [CrossRef]
- Chen, Z.; Karaplis, A.C.; Ackerman, S.L.; Pogribny, I.P.; Melnyk, S.; Lussier-Cacan, S.; Chen, M.F.; Pai, A.; John, S.W.; Smith, R.S.; et al. Mice deficient in methylenetetrahydrofolate reductase exhibit hyperhomocysteinemia and decreased methylation capacity, with neuropathology and aortic lipid deposition. Hum. Mol. Genet. 2001, 10, 433–443. [Google Scholar] [CrossRef]
- Bjorklund, N.K.; Gordon, R. A hypothesis linking low folate intake to neural tube defects due to failure of post-translation methylations of the cytoskeleton. Int. J. Dev. Biol. 2006, 50, 135–141. [Google Scholar] [CrossRef]
- Moephuli, S.R.; Klein, N.W.; Baldwin, M.T.; Krider, H.M. Effects of methionine on the cytoplasmic distribution of actin and tubulin during neural tube closure in rat embryos. Proc. Natl. Acad. Sci. USA 1997, 94, 543–548. [Google Scholar]
- Felkner, M.; Suarez, L.; Canfield, M.A.; Brender, J.D.; Sun, Q. Maternal serum homocysteine and risk for neural tube defects in a Texas-Mexico border population. Birth Defects Res. 2009, 85, 574–581. [Google Scholar] [CrossRef]
- Gaber, K.R.; Farag, M.K.; Soliman, S.E.; El-Bassyouni, H.T.; El-Kamah, G. Maternal vitamin B12 and the risk of fetal neural tube defects in Egyptian patients. Clin. Lab. 2007, 53, 69–75. [Google Scholar]
- Gu, Q.; Li, Y.; Cui, Z.L.; Luo, X.P. Homocysteine, folate, vitamin B12 and B6 in mothers of children with neural tube defects in Xinjiang, China. Acta Paediatr. 2012, 101, e486–e490. [Google Scholar] [CrossRef]
- Mills, J.L.; McPartlin, J.M.; Kirke, P.N.; Lee, Y.J.; Conley, M.R.; Weir, D.G.; Scott, J.M. Homocysteine metabolism in pregnancies complicated by neural-tube defects. Lancet 1995, 345, 149–151. [Google Scholar]
- Van der Put, N.M.; Thomas, C.M.; Eskes, T.K.; Trijbels, F.J.; Steegers-Theunissen, R.P.; Mariman, E.C.; de Graaf-Hess, A.; Smeitink, J.A.; Blom, H.J. Altered folate and vitamin B12 metabolism in families with spina bifida offspring. QJM 1997, 90, 505–510. [Google Scholar] [CrossRef]
- Wenstrom, K.D.; Johanning, G.L.; Owen, J.; Johnston, K.E.; Acton, S.; Cliver, S.; Tamura, T. Amniotic fluid homocysteine levels, 5,10-methylenetetrahydrafolate reductase genotypes, and neural tube closure sites. Am. J. Med. Genet. 2000, 90, 6–11. [Google Scholar] [CrossRef]
- Ceyhan, S.T.; Beyan, C.; Atay, V.; Yaman, H.; Alanbay, I.; Kaptan, K.; Baser, I. Serum vitamin B12 and homocysteine levels in pregnant women with neural tube defect. Gynecol. Endocrinol. 2010, 26, 578–581. [Google Scholar] [CrossRef]
- Felix, T.M.; Leistner, S.; Giugliani, R. Metabolic effects and the methylenetetrahydrofolate reductase (MTHFR) polymorphism associated with neural tube defects in southern Brazil. Birth Defects Res. 2004, 70, 459–463. [Google Scholar] [CrossRef]
- Grandone, E.; Corrao, A.M.; Colaizzo, D.; Vecchione, G.; di Girgenti, C.; Paladini, D.; Sardella, L.; Pellegrino, M.; Zelante, L.; Martinelli, P.; et al. Homocysteine metabolism in families from southern Italy with neural tube defects: Role of genetic and nutritional determinants. Prenat. Diagn. 2006, 26, 1–5. [Google Scholar] [CrossRef]
- Groenen, P.M.; van Rooij, I.A.; Peer, P.G.; Gooskens, R.H.; Zielhuis, G.A.; Steegers-Theunissen, R.P. Marginal maternal vitamin B12 status increases the risk of offspring with spina bifida. Am. J. Obstet. Gynecol. 2004, 191, 11–17. [Google Scholar] [CrossRef]
- Ratan, S.K.; Rattan, K.N.; Pandey, R.M.; Singhal, S.; Kharab, S.; Bala, M.; Singh, V.; Jhanwar, A. Evaluation of the levels of folate, vitamin B12, homocysteine and fluoride in the parents and the affected neonates with neural tube defect and their matched controls. Pediatr. Surg. Int. 2008, 24, 803–808. [Google Scholar] [CrossRef]
- Zhang, H.Y.; Luo, G.A.; Liang, Q.L.; Wang, Y.; Yang, H.H.; Wang, Y.M.; Zheng, X.Y.; Song, X.M.; Chen, G.; Zhang, T.; et al. Neural tube defects and disturbed maternal folate- and homocysteine-mediated one-carbon metabolism. Exp. Neurol. 2008, 212, 515–521. [Google Scholar] [CrossRef]
- Graham, A.; Brender, J.D.; Sharkey, J.R.; Zhu, L.; Felkner, M.; Suarez, L.; Canfield, M.A. Dietary methionine intake and neural tube defects in Mexican-American women. Birth Defects Res. 2010, 88, 451–457. [Google Scholar] [CrossRef]
- Shaw, G.M.; Velie, E.M.; Schaffer, D.M. Is dietary intake of methionine associated with a reduction in risk for neural tube defect-affected pregnancies? Teratology 1997, 56, 295–299. [Google Scholar] [CrossRef]
- Shoob, H.D.; Sargent, R.G.; Thompson, S.J.; Best, R.G.; Drane, J.W.; Tocharoen, A. Dietary methionine is involved in the etiology of neural tube defect-affected pregnancies in humans. J. Nutr. 2001, 131, 2653–2658. [Google Scholar]
- Rosenquist, T.H.; Finnell, R.H. Genes, folate and homocysteine in embryonic development. Proc. Nutr. Soc. 2001, 60, 53–61. [Google Scholar]
- Van Mil, N.H.; Oosterbaan, A.M.; Steegers-Theunissen, R.P. Teratogenicity and underlying mechanisms of homocysteine in animal models: A review. Reprod. Toxicol. 2010, 30, 520–531. [Google Scholar] [CrossRef]
- Afman, L.A.; Blom, H.J.; van der Put, N.M.; van Straaten, H.W. Homocysteine interference in neurulation: A chick embryo model. Birth Defects Res. 2003, 67, 421–428. [Google Scholar] [CrossRef]
- Rosenquist, T.H.; Ratashak, S.A.; Selhub, J. Homocysteine induces congenital defects of the heart and neural tube: Effect of folic acid. Proc. Natl. Acad. Sci. USA 1996, 93, 15227–15232. [Google Scholar] [CrossRef]
- Vanaerts, L.A.; Blom, H.J.; Deabreu, R.A.; Trijbels, F.J.; Eskes, T.K.; Copius Peereboom-Stegeman, J.H.; Noordhoek, J. Prevention of neural tube defects by and toxicity of L-homocysteine in cultured postimplantation rat embryos. Teratology 1994, 50, 348–360. [Google Scholar] [CrossRef]
- Bennett, G.D.; Vanwaes, J.; Moser, K.; Chaudoin, T.; Starr, L.; Rosenquist, T.H. Failure of homocysteine to induce neural tube defects in a mouse model. Birth Defects Res. B 2006, 77, 89–94. [Google Scholar] [CrossRef]
- Greene, N.D.; Dunlevy, L.E.; Copp, A.J. Homocysteine is embryotoxic but does not cause neural tube defects in mouse embryos. Anat. Embryol. 2003, 206, 185–191. [Google Scholar]
- Watanabe, M.; Osada, J.; Aratani, Y.; Kluckman, K.; Reddick, R.; Malinow, M.R.; Maeda, N. Mice deficient in cystathionine beta-synthase: Animal models for mild and severe homocyst(e)inemia. Proc. Natl. Acad. Sci. USA 1995, 92, 1585–1589. [Google Scholar]
- Carmichael, S.L.; Yang, W.; Shaw, G.M. Periconceptional nutrient intakes and risks of neural tube defects in California. Birth Defects Res. 2010, 88, 670–678. [Google Scholar] [CrossRef]
- Magnus, P.; Magnus, E.M.; Berg, K. Increased levels of apo-transcobalamins I and II in amniotic fluid from pregnant women with previous neural tube defect offspring. Clin. Genet. 1986, 30, 167–172. [Google Scholar] [CrossRef]
- Li, F.; Watkins, D.; Rosenblatt, D.S. Vitamin B(12) and birth defects. Mol. Genet. Metab. 2009, 98, 166–172. [Google Scholar] [CrossRef]
- Cech, I.; Burau, K.D. Serological differences in folate/vitamin B12 in pregnancies affected by neural tube defects. South. Med. J. 2010, 103, 419–424. [Google Scholar] [CrossRef]
- Kirke, P.N.; Molloy, A.M.; Daly, L.E.; Burke, H.; Weir, D.G.; Scott, J.M. Maternal plasma folate and vitamin B12 are independent risk factors for neural tube defects. Q. J. Med. 1993, 86, 703–708. [Google Scholar]
- Molloy, A.M.; Kirke, P.N.; Troendle, J.F.; Burke, H.; Sutton, M.; Brody, L.C.; Scott, J.M.; Mills, J.L. Maternal vitamin B12 status and risk of neural tube defects in a population with high neural tube defect prevalence and no folic acid fortification. Pediatrics 2009, 123, 917–923. [Google Scholar] [CrossRef]
- Song, Y.Z.; Li, B.X.; Hao, H.; Xin, R.L.; Zhang, T.; Zhang, C.H.; Kobayashi, K.; Wang, Z.N.; Zheng, X.Y. Selective screening for inborn errors of metabolism and secondary methylmalonic aciduria in pregnancy at high risk district of neural tube defects: A human metabolome study by GC-MS in China. Clin. Biochem. 2008, 41, 616–620. [Google Scholar] [CrossRef]
- Suarez, L.; Hendricks, K.; Felkner, M.; Gunter, E. Maternal serum B12 levels and risk for neural tube defects in a Texas-Mexico border population. Ann. Epidemiol. 2003, 13, 81–88. [Google Scholar] [CrossRef]
- Zhang, T.; Xin, R.; Gu, X.; Wang, F.; Pei, L.; Lin, L.; Chen, G.; Wu, J.; Zheng, X. Maternal serum vitamin B12, folate and homocysteine and the risk of neural tube defects in the offspring in a high-risk area of China. Public Health Nutr. 2009, 12, 680–686. [Google Scholar] [CrossRef]
- Afman, L.A.; van der Put, N.M.; Thomas, C.M.; Trijbels, J.M.; Blom, H.J. Reduced vitamin B12 binding by transcobalamin II increases the risk of neural tube defects. QJM 2001, 94, 159–166. [Google Scholar] [CrossRef]
- Luo, X.; Zhang, L.; Wei, H.; Liu, W.; Wang, M.; Ning, Q. Methylmalonic acid in amniotic fluid and maternal urine as a marker for neural tube defects. J. Huazhong Univ. Sci. Technol. Med. Sci. 2004, 24, 166–169. [Google Scholar] [CrossRef]
- Mills, J.L.; Tuomilehto, J.; Yu, K.F.; Colman, N.; Blaner, W.S.; Koskela, P.; Rundle, W.E.; Forman, M.; Toivanen, L.; Rhoads, G.G. Maternal vitamin levels during pregnancies producing infants with neural tube defects. J. Pediatr. 1992, 120, 863–871. [Google Scholar] [CrossRef]
- Mobasheri, E.; Keshtkar, A.; Golalipour, M.J. Maternal folate and vitamin B(12) status and neural tube defects in northern iran: A case control study. Iran. J. Pediatr. 2010, 20, 167–173. [Google Scholar]
- Ubbink, J.B.; Christianson, A.; Bester, M.J.; van Allen, M.I.; Venter, P.A.; Delport, R.; Blom, H.J.; van der Merwe, A.; Potgieter, H.; Vermaak, W.J. Folate status, homocysteine metabolism, and methylene tetrahydrofolate reductase genotype in rural South African blacks with a history of pregnancy complicated by neural tube defects. Metabolism 1999, 48, 269–274. [Google Scholar] [CrossRef]
- Ray, J.G.; Blom, H.J. Vitamin B12 insufficiency and the risk of fetal neural tube defects. QJM 2003, 96, 289–295. [Google Scholar] [CrossRef]
- Wang, Z.P.; Shang, X.X.; Zhao, Z.T. Low maternal vitamin B(12) is a risk factor for neural tube defects: A meta-analysis. J. Matern. Fetal Neonatal Med. 2012, 25, 389–394. [Google Scholar] [CrossRef]
- Magnus, P.; Magnus, E.M.; Berg, K. Transcobalamins in the etiology of neural tube defects. Clin. Genet. 1991, 39, 309–310. [Google Scholar] [CrossRef]
- Ray, J.G.; Wyatt, P.R.; Thompson, M.D.; Vermeulen, M.J.; Meier, C.; Wong, P.Y.; Farrell, S.A.; Cole, D.E. Vitamin B12 and the risk of neural tube defects in a folic-acid-fortified population. Epidemiology 2007, 18, 362–366. [Google Scholar] [CrossRef]
- Adams, M.J., Jr.; Khoury, M.J.; Scanlon, K.S.; Stevenson, R.E.; Knight, G.J.; Haddow, J.E.; Sylvester, G.C.; Cheek, J.E.; Henry, J.P.; Stabler, S.P.; et al. Elevated midtrimester serum methylmalonic acid levels as a risk factor for neural tube defects. Teratology 1995, 51, 311–317. [Google Scholar]
- Shaw, G.M.; Carmichael, S.L.; Yang, W.; Selvin, S.; Schaffer, D.M. Periconceptional dietary intake of choline and betaine and neural tube defects in offspring. Am. J. Epidemiol. 2004, 160, 102–109. [Google Scholar] [CrossRef]
- Chandler, A.L.; Hobbs, C.A.; Mosley, B.S.; Berry, R.J.; Canfield, M.A.; Qi, Y.P.; Siega-Riz, A.M.; Shaw, G.M. Neural tube defects and maternal intake of micronutrients related to one-carbon metabolism or antioxidant activity. Birth Defects Res. 2012, 94, 864–874. [Google Scholar] [CrossRef]
- Enaw, J.O.; Zhu, H.; Yang, W.; Lu, W.; Shaw, G.M.; Lammer, E.J.; Finnell, R.H. Chka and Pcyt1a gene polymorphisms, choline intake and spina bifida risk in a California population. BMC Med. 2006, 4, 36–41. [Google Scholar] [CrossRef]
- Shaw, G.M.; Finnell, R.H.; Blom, H.J.; Carmichael, S.L.; Vollset, S.E.; Yang, W.; Ueland, P.M. Choline and risk of neural tube defects in a folate-fortified population. Epidemiology 2009, 20, 714–719. [Google Scholar] [CrossRef]
- Fisher, M.C.; Zeisel, S.H.; Mar, M.H.; Sadler, T.W. Inhibitors of choline uptake and metabolism cause developmental abnormalities in neurulating mouse embryos. Teratology 2001, 64, 114–122. [Google Scholar] [CrossRef]
- Kang, S.S.; Zhou, J.; Wong, P.W.; Kowalisyn, J.; Strokosch, G. Intermediate homocysteinemia: A thermolabile variant of methylenetetrahydrofolate reductase. Am. J. Hum. Genet. 1988, 43, 414–421. [Google Scholar]
- Van der Put, N.M.; Steegers-Theunissen, R.P.; Frosst, P.; Trijbels, F.J.; Eskes, T.K.; van den Heuvel, L.P.; Mariman, E.C.; den Heyer, M.; Rozen, R.; Blom, H.J. Mutated methylenetetrahydrofolate reductase as a risk factor for spina bifida. Lancet 1995, 346, 1070–1071. [Google Scholar] [CrossRef]
- Amorim, M.R.; Lima, M.A.; Castilla, E.E.; Orioli, I.M. Non-Latin European descent could be a requirement for association of NTDs and MTHFR variant 677C > T: A meta-analysis. Am. J. Med. Genet. A 2007, 143A, 1726–1732. [Google Scholar] [CrossRef]
- Botto, L.D.; Yang, Q. 5,10-methylenetetrahydrofolate reductase gene variants and congenital anomalies: A huge review. Am. J. Epidemiol. 2000, 151, 862–877. [Google Scholar] [CrossRef]
- Van der Put, N.M.; Eskes, T.K.; Blom, H.J. Is the common 677C > T mutation in the methylenetetrahydrofolate reductase gene a risk factor for neural tube defects? A meta-analysis. QJM 1997, 90, 111–115. [Google Scholar] [CrossRef]
- Yan, L.; Zhao, L.; Long, Y.; Zou, P.; Ji, G.; Gu, A.; Zhao, P. Association of the maternal mthfr C677T polymorphism with susceptibility to neural tube defects in offsprings: Evidence from 25 case-control studies. PLoS One 2012, 7, e41689. [Google Scholar] [CrossRef]
- Boyles, A.L.; Billups, A.V.; Deak, K.L.; Siegel, D.G.; Mehltretter, L.; Slifer, S.H.; Bassuk, A.G.; Kessler, J.A.; Reed, M.C.; Nijhout, H.F.; et al. Neural tube defects and folate pathway genes: Family-based association tests of gene-gene and gene-environment interactions. Environ. Health Perspect. 2006, 114, 1547–1552. [Google Scholar] [CrossRef]
- De Marco, P.; Calevo, M.G.; Moroni, A.; Merello, E.; Raso, A.; Finnell, R.H.; Zhu, H.; Andreussi, L.; Cama, A.; Capra, V. Reduced folate carrier polymorphism (80A > G) and neural tube defects. Eur. J. Hum. Genet. 2003, 11, 245–252. [Google Scholar] [CrossRef]
- Speer, M.C.; Nye, J.; McLone, D.; Worley, G.; Melvin, E.C.; Viles, K.D.; Franklin, A.; Drake, C.; Mackey, J.; George, T.M. Possible interaction of genotypes at cystathionine beta-synthase and methylenetetrahydrofolate reductase (MTHFR) in neural tube defects. NTD collaborative group. Clin. Genet. 1999, 56, 142–144. [Google Scholar] [CrossRef]
- Wilson, A.; Platt, R.; Wu, Q.; Leclerc, D.; Christensen, B.; Yang, H.; Gravel, R.A.; Rozen, R. A common variant in methionine synthase reductase combined with low cobalamin (vitamin B12) increases risk for spina bifida. Mol. Genet. Metab. 1999, 67, 317–323. [Google Scholar] [CrossRef]
- Van der Linden, I.J.; den Heijer, M.; Afman, L.A.; Gellekink, H.; Vermeulen, S.H.; Kluijtmans, L.A.; Blom, H.J. The methionine synthase reductase 66A > G polymorphism is a maternal risk factor for spina bifida. J. Mol. Med. 2006, 84, 1047–1054. [Google Scholar] [CrossRef]
- Shaw, G.M.; Lammer, E.J.; Zhu, H.; Baker, M.W.; Neri, E.; Finnell, R.H. Maternal periconceptional vitamin use, genetic variation of infant reduced folate carrier (A80G), and risk of spina bifida. Am. J. Med. Genet. 2002, 108, 1–6. [Google Scholar] [CrossRef]
- Shaw, G.M.; Rozen, R.; Finnell, R.H.; Wasserman, C.R.; Lammer, E.J. Maternal vitamin use, genetic variation of infant methylenetetrahydrofolate reductase, and risk for spina bifida. Am. J. Epidemiol. 1998, 148, 30–37. [Google Scholar] [CrossRef]
- Shang, Y.; Zhao, H.; Niu, B.; Li, W.I.; Zhou, R.; Zhang, T.; Xie, J. Correlation of polymorphism of MTHFRS and RFC-1 genes with neural tube defects in China. Birth Defects Res. 2008, 82, 3–7. [Google Scholar] [CrossRef]
- Volcik, K.A.; Blanton, S.H.; Tyerman, G.H.; Jong, S.T.; Rott, E.J.; Page, T.Z.; Romaine, N.K.; Northrup, H. Methylenetetrahydrofolate reductase and spina bifida: Evaluation of level of defect and maternal genotypic risk in Hispanics. Am. J. Med. Genet. 2000, 95, 21–27. [Google Scholar] [CrossRef]
- Franke, B.; Vermeulen, S.H.; Steegers-Theunissen, R.P.; Coenen, M.J.; Schijvenaars, M.M.; Scheffer, H.; den Heijer, M.; Blom, H.J. An association study of 45 folate-related genes in spina bifida: Involvement of cubilin (CUBN) and trna aspartic acid methyltransferase 1 (TRDMT1). Birth Defects Res. 2009, 85, 216–226. [Google Scholar] [CrossRef]
- Afman, L.A.; Lievers, K.J.; van der Put, N.M.; Trijbels, F.J.; Blom, H.J. Single nucleotide polymorphisms in the transcobalamin gene: Relationship with transcobalamin concentrations and risk for neural tube defects. Eur. J. Hum. Genet. 2002, 10, 433–438. [Google Scholar] [CrossRef]
- Gueant-Rodriguez, R.M.; Rendeli, C.; Namour, B.; Venuti, L.; Romano, A.; Anello, G.; Bosco, P.; Debard, R.; Gerard, P.; Viola, M.; et al. Transcobalamin and methionine synthase reductase mutated polymorphisms aggravate the risk of neural tube defects in humans. Neurosci. Lett. 2003, 344, 189–192. [Google Scholar] [CrossRef]
- Swanson, D.A.; Pangilinan, F.; Mills, J.L.; Kirke, P.N.; Conley, M.; Weiler, A.; Frey, T.; Parle-McDermott, A.; O’Leary, V.B.; Seltzer, R.R.; et al. Evaluation of transcobalamin II polymorphisms as neural tube defect risk factors in an Irish population. Birth Defects Res. 2005, 73, 239–244. [Google Scholar] [CrossRef]
- Pietrzyk, J.J.; Bik-Multanowski, M. 776C > G polymorphism of the transcobalamin II Gene as a risk factor for spina bifida. Mol. Genet. Metab. 2003, 80, 364. [Google Scholar] [CrossRef]
- Morin, I.; Platt, R.; Weisberg, I.; Sabbaghian, N.; Wu, Q.; Garrow, T.A.; Rozen, R. Common variant in betaine-homocysteine methyltransferase (BHMT) and risk for spina bifida. Am. J. Med. Genet. 2003, 119A, 172–176. [Google Scholar] [CrossRef]
- Zhu, H.; Curry, S.; Wen, S.; Wicker, N.J.; Shaw, G.M.; Lammer, E.J.; Yang, W.; Jafarov, T.; Finnell, R.H. Are the betaine-homocysteine methyltransferase (BHMT and BHMT2) genes risk factors for spina bifida and orofacial clefts? Am. J. Med. Genet. A 2005, 135, 274–277. [Google Scholar]
- Johnson, W.G.; Stenroos, E.S.; Spychala, J.R.; Chatkupt, S.; Ming, S.X.; Buyske, S. New 19 bp deletion polymorphism in intron-1 of dihydrofolate reductase (DHFR): A risk factor for spina bifida acting in mothers during pregnancy? Am. J. Med. Genet. A 2004, 124A, 339–345. [Google Scholar] [CrossRef]
- Doudney, K.; Grinham, J.; Whittaker, J.; Lynch, S.A.; Thompson, D.; Moore, G.E.; Copp, A.J.; Greene, N.D.; Stanier, P. Evaluation of folate metabolism gene polymorphisms as risk factors for open and closed neural tube defects. Am. J. Med. Genet. A 2009, 149A, 1585–1589. [Google Scholar] [CrossRef]
- Van der Linden, I.J.; Nguyen, U.; Heil, S.G.; Franke, B.; Vloet, S.; Gellekink, H.; den Heijer, M.; Blom, H.J. Variation and expression of dihydrofolate reductase (DHFR) in relation to spina bifida. Mol. Genet. Metab. 2007, 91, 98–103. [Google Scholar] [CrossRef]
- O’Leary, V.B.; Mills, J.L.; Kirke, P.N.; Parle-McDermott, A.; Swanson, D.A.; Weiler, A.; Pangilinan, F.; Conley, M.; Molloy, A.M.; Lynch, M.; et al. Analysis of the human folate receptor beta gene for an association with neural tube defects. Mol. Genet. Metab. 2003, 79, 129–133. [Google Scholar] [CrossRef]
- Relton, C.L.; Wilding, C.S.; Pearce, M.S.; Laffling, A.J.; Jonas, P.A.; Lynch, S.A.; Tawn, E.J.; Burn, J. Gene-gene interaction in folate-related genes and risk of neural tube defects in a UK population. J. Med. Genet. 2004, 41, 256–260. [Google Scholar] [CrossRef]
- Morin, I.; Devlin, A.M.; Leclerc, D.; Sabbaghian, N.; Halsted, C.H.; Finnell, R.; Rozen, R. Evaluation of genetic variants in the reduced folate carrier and in glutamate carboxypeptidase II for spina bifida risk. Mol. Genet. Metab. 2003, 79, 197–200. [Google Scholar] [CrossRef]
- Brody, L.C.; Conley, M.; Cox, C.; Kirke, P.N.; McKeever, M.P.; Mills, J.L.; Molloy, A.M.; O’Leary, V.B.; Parle-McDermott, A.; Scott, J.M.; et al. A polymorphism, r653q, in the trifunctional enzyme methylenetetrahydrofolate dehydrogenase/methenyltetrahydrofolate cyclohydrolase/formyltetrahydrofolate synthetase is a maternal genetic risk factor for neural tube defects: Report of the birth defects research group. Am. J. Hum. Genet. 2002, 71, 1207–1215. [Google Scholar] [CrossRef]
- De Marco, P.; Merello, E.; Calevo, M.G.; Mascelli, S.; Raso, A.; Cama, A.; Capra, V. Evaluation of a methylenetetrahydrofolate-dehydrogenase 1958G > A polymorphism for neural tube defect risk. J. Hum. Genet. 2006, 51, 98–103. [Google Scholar] [CrossRef]
- Hol, F.A.; van der Put, N.M.; Geurds, M.P.; Heil, S.G.; Trijbels, F.J.; Hamel, B.C.; Mariman, E.C.; Blom, H.J. Molecular genetic analysis of the gene encoding the trifunctional enzyme mthfd (methylenetetrahydrofolate-dehydrogenase, methenyltetrahydrofolate-cyclohydrolase, formyltetrahydrofolate synthetase) in patients with neural tube defects. Clin. Genet. 1998, 53, 119–125. [Google Scholar]
- Parle-McDermott, A.; Kirke, P.N.; Mills, J.L.; Molloy, A.M.; Cox, C.; O’Leary, V.B.; Pangilinan, F.; Conley, M.; Cleary, L.; Brody, L.C.; et al. Confirmation of the R653Q polymorphism of the trifunctional C1-synthase enzyme as a maternal risk for neural tube defects in the Irish population. Eur. J. Hum. Genet. 2006, 14, 768–772. [Google Scholar] [CrossRef]
- Van der Linden, I.J.; Heil, S.G.; Kouwenberg, I.C.; den Heijer, M.; Blom, H.J. The methylenetetrahydrofolate dehydrogenase (MTHFD1) 1958G > A variant is not associated with spina bifida risk in the Dutch population. Clin. Genet. 2007, 72, 599–600. [Google Scholar] [CrossRef]
- Parle-McDermott, A.; Pangilinan, F.; O’Brien, K.K.; Mills, J.L.; Magee, A.M.; Troendle, J.; Sutton, M.; Scott, J.M.; Kirke, P.N.; Molloy, A.M.; et al. A common variant in MTHFD1L is associated with neural tube defects and MRNA splicing efficiency. Hum. Mutat. 2009, 30, 1650–1656. [Google Scholar] [CrossRef]
- Morrison, K.; Papapetrou, C.; Hol, F.A.; Mariman, E.C.; Lynch, S.A.; Burn, J.; Edwards, Y.H. Susceptibility to spina bifida; An association study of five candidate genes. Ann. Hum. Genet. 1998, 62, 379–396. [Google Scholar] [CrossRef]
- Boduroglu, K.; Alanay, Y.; Alikasifoglu, M.; Aktas, D.; Tuncbilek, E. Analysis of MTHFR 1298A > C in addition to MTHFR 677C > T polymorphism as a risk factor for neural tube defects in the Turkish population. Turk. J. Pediatr. 2005, 47, 327–333. [Google Scholar]
- De Marco, P.; Calevo, M.G.; Moroni, A.; Arata, L.; Merello, E.; Cama, A.; Finnell, R.H.; Andreussi, L.; Capra, V. Polymorphisms in genes involved in folate metabolism as risk factors for ntds. Eur. J. Pediatr. Surg. 2001, 11, S14–S17. [Google Scholar] [CrossRef]
- De Marco, P.; Calevo, M.G.; Moroni, A.; Arata, L.; Merello, E.; Finnell, R.H.; Zhu, H.; Andreussi, L.; Cama, A.; Capra, V. Study of MTHFR and ms polymorphisms as risk factors for ntd in the Italian population. J. Hum. Genet. 2002, 47, 319–324. [Google Scholar] [CrossRef]
- Gonzalez-Herrera, L.; Castillo-Zapata, I.; Garcia-Escalante, G.; Pinto-Escalante, D. A1298c polymorphism of the MTHFR gene and neural tube defects in the state of Yucatan, Mexico. Birth Defects Res. 2007, 79, 622–626. [Google Scholar] [CrossRef]
- Parle-McDermott, A.; Mills, J.L.; Kirke, P.N.; O’Leary, V.B.; Swanson, D.A.; Pangilinan, F.; Conley, M.; Molloy, A.M.; Cox, C.; Scott, J.M.; et al. Analysis of the MTHFR 1298A > C and 677C > T polymorphisms as risk factors for neural tube defects. J. Hum. Genet. 2003, 48, 190–193. [Google Scholar] [CrossRef]
- Van der Put, N.M.; Gabreels, F.; Stevens, E.M.; Smeitink, J.A.; Trijbels, F.J.; Eskes, T.K.; van den Heuvel, L.P.; Blom, H.J. A second common mutation in the methylenetetrahydrofolate reductase gene: An additional risk factor for neural-tube defects? Am. J. Hum. Genet. 1998, 62, 1044–1051. [Google Scholar] [CrossRef]
- Behunova, J.; Klimcakova, L.; Zavadilikova, E.; Potocekova, D.; Sykora, P.; Podracka, L. Methylenetetrahydrofolate reductase gene polymorphisms and neural tube defects epidemiology in the Slovak population. Birth Defects Res. 2010, 88, 695–700. [Google Scholar] [CrossRef]
- Gutierrez Revilla, J.I.; Perez Hernandez, F.; Calvo Martin, M.T.; Tamparillas Salvador, M.; Gracia Romero, J. C677T and A1298C MTHFR polymorphisms in the etiology of neural tube defects in Spanish population. Med. Clin. 2003, 120, 441–445. [Google Scholar]
- Papapetrou, C.; Lynch, S.A.; Burn, J.; Edwards, Y.H. Methylenetetrahydrofolate reductase and neural tube defects. Lancet 1996, 348, 58. [Google Scholar] [CrossRef]
- Rampersaud, E.; Melvin, E.C.; Siegel, D.; Mehltretter, L.; Dickerson, M.E.; George, T.M.; Enterline, D.; Nye, J.S.; Speer, M.C. Updated investigations of the role of methylenetetrahydrofolate reductase in human neural tube defects. Clin. Genet. 2003, 63, 210–214. [Google Scholar] [CrossRef]
- Gonzalez-Herrera, L.; Garcia-Escalante, G.; Castillo-Zapata, I.; Canto-Herrera, J.; Ceballos-Quintal, J.; Pinto-Escalante, D.; Diaz-Rubio, F.; del Angel, R.M.; Orozco-Orozco, L. Frequency of the thermolabile variant C677T in the MTHFR gene and lack of association with neural tube defects in the state of Yucatan, Mexico. Clin. Genet. 2002, 62, 394–398. [Google Scholar] [CrossRef]
- Koch, M.C.; Stegmann, K.; Ziegler, A.; Schroter, B.; Ermert, A. Evaluation of the MTHFR C677T allele and the MTHFR gene locus in a German spina bifida population. Eur. J. Pediatr. 1998, 157, 487–492. [Google Scholar]
- Mornet, E.; Muller, F.; Lenvoise-Furet, A.; Delezoide, A.L.; Col, J.Y.; Simon-Bouy, B.; Serre, J.L. Screening of the C677T mutation on the methylenetetrahydrofolate reductase gene in french patients with neural tube defects. Hum. Genet. 1997, 100, 512–514. [Google Scholar]
- De Franchis, R.; Buoninconti, A.; Mandato, C.; Pepe, A.; Sperandeo, M.P.; del Gado, R.; Capra, V.; Salvaggio, E.; Andria, G.; Mastroiacovo, P. The C677T mutation of the 5,10-methylenetetrahydrofolate reductase gene is a moderate risk factor for spina bifida in Italy. J. Med. Genet. 1998, 35, 1009–1013. [Google Scholar]
- Johanning, G.L.; Tamura, T.; Johnston, K.E.; Wenstrom, K.D. Comorbidity of 5,10-methylenetetrahydrofolate reductase and methionine synthase gene polymorphisms and risk for neural tube defects. J. Med. Genet. 2000, 37, 949–951. [Google Scholar] [CrossRef]
- Kirke, P.N.; Mills, J.L.; Molloy, A.M.; Brody, L.C.; O’Leary, V.B.; Daly, L.; Murray, S.; Conley, M.; Mayne, P.D.; Smith, O.; et al. Impact of the MTHFR C677T polymorphism on risk of neural tube defects: Case-control study. BMJ 2004, 328, 1535–1536. [Google Scholar] [CrossRef]
- Kirke, P.N.; Mills, J.L.; Whitehead, A.S.; Molloy, A.; Scott, J.M. Methylenetetrahydrofolate reductase mutation and neural tube defects. Lancet 1996, 348, 1037–1038. [Google Scholar]
- Ou, C.Y.; Stevenson, R.E.; Brown, V.K.; Schwartz, C.E.; Allen, W.P.; Khoury, M.J.; Rozen, R.; Oakley, G.P., Jr.; Adams, M.J., Jr. 5,10 methylenetetrahydrofolate reductase genetic polymorphism as a risk factor for neural tube defects. Am. J. Med. Genet. 1996, 63, 610–614. [Google Scholar] [CrossRef]
- Pietrzyk, J.J.; Bik-Multanowski, M.; Sanak, M.; Twardowska, M. Polymorphisms of the 5,10-methylenetetrahydrofolate and the methionine synthase reductase genes as independent risk factors for spina bifida. J. Appl. Genet. 2003, 44, 111–113. [Google Scholar]
- Shields, D.C.; Kirke, P.N.; Mills, J.L.; Ramsbottom, D.; Molloy, A.M.; Burke, H.; Weir, D.G.; Scott, J.M.; Whitehead, A.S. The thermolabile variant of methylenetetrahydrofolate reductase and neural tube defects: An evaluation of genetic risk and the relative importance of the genotypes of the embryo and the mother. Am. J. Hum. Genet. 1999, 64, 1045–1055. [Google Scholar] [CrossRef]
- Whitehead, A.S.; Gallagher, P.; Mills, J.L.; Kirke, P.N.; Burke, H.; Molloy, A.M.; Weir, D.G.; Shields, D.C.; Scott, J.M. A genetic defect in 5,10 methylenetetrahydrofolate reductase in neural tube defects. QJM 1995, 88, 763–766. [Google Scholar]
- Davalos, I.P.; Olivares, N.; Castillo, M.T.; Cantu, J.M.; Ibarra, B.; Sandoval, L.; Moran, M.C.; Gallegos, M.P.; Chakraborty, R.; Rivas, F. The C677T polymorphism of the methylenetetrahydrofolate reductase gene in Mexican mestizo neural-tube defect parents, control mestizo and native populations. Ann. Genet. 2000, 43, 89–92. [Google Scholar]
- Lucock, M.; Daskalakis, I.; Briggs, D.; Yates, Z.; Levene, M. Altered folate metabolism and disposition in mothers affected by a spina bifida pregnancy: Influence of 677C > T methylenetetrahydrofolate reductase and 2756A > G methionine synthase genotypes. Mol. Genet. Metab. 2000, 70, 27–44. [Google Scholar] [CrossRef]
- Martinez de Villarreal, L.E.; Delgado-Enciso, I.; Valdez-Leal, R.; Ortiz-Lopez, R.; Rojas-Martinez, A.; Limon-Benavides, C.; Sanchez-Pena, M.A.; Ancer-Rodriguez, J.; Barrera-Saldana, H.A.; Villarreal-Perez, J.Z. Folate levels and N(5)-N(10)-methylenetetrahydrofolate reductase genotype (MTHFR) in mothers of offspring with neural tube defects: A case-control study. Arch. Med. Res. 2001, 32, 277–282. [Google Scholar] [CrossRef]
- Richter, B.; Stegmann, K.; Roper, B.; Boddeker, I.; Ngo, E.T.; Koch, M.C. Interaction of folate and homocysteine pathway genotypes evaluated in susceptibility to neural tube defects (NTD) in a German population. J. Hum. Genet. 2001, 46, 105–109. [Google Scholar] [CrossRef]
- O’Leary, V.; Pangilinan, F.; Cox, C.; Parle-McDermott, A.; Conley, M.; Molloy, A.M.; Kirke, P.N.; Mills, J.L.; Brody, L.C.; Scott, J.M. Reduced folate carrier polymorphisms and neural tube defect risk. Mol. Genet. Metab. 2006, 87, 364–369. [Google Scholar] [CrossRef]
- Pei, L.; Zhu, H.; Ren, A.; Li, Z.; Hao, L.; Finnell, R.H. Reduced folate carrier gene is a risk factor for neural tube defects in a Chinese population. Birth Defects Res. 2005, 73, 430–433. [Google Scholar] [CrossRef]
- Vieira, A.R.; Murray, J.C.; Trembath, D.; Orioli, I.M.; Castilla, E.E.; Cooper, M.E.; Marazita, M.L.; Lennon-Graham, F.; Speer, M. Studies of reduced folate carrier 1 (RFC1) A80G and 5,10-methylenetetrahydrofolate reductase (MTHFR) C677T polymorphisms with neural tube and orofacial cleft defects. Am. J. Med. Genet. 2005, 135, 220–223. [Google Scholar]
- Wang, H.G.; Wang, J.L.; Zhang, J.; Zhao, L.X.; Zhai, G.X.; Xiang, Y.Z.; Chang, P. Reduced folate carrier A80G polymorphism and susceptibility to neural tube defects: A meta-analysis. Gene 2012, 510, 180–184. [Google Scholar] [CrossRef]
- Heil, S.G.; van der Put, N.M.; Waas, E.T.; den Heijer, M.; Trijbels, F.J.; Blom, H.J. Is mutated serine hydroxymethyltransferase (SHMT) involved in the etiology of neural tube defects? Mol. Genet. Metab. 2001, 73, 164–172. [Google Scholar] [CrossRef]
- O’Leary, V.B.; Mills, J.L.; Pangilinan, F.; Kirke, P.N.; Cox, C.; Conley, M.; Weiler, A.; Peng, K.; Shane, B.; Scott, J.M.; et al. Analysis of methionine synthase reductase polymorphisms for neural tube defects risk association. Mol. Genet. Metab. 2005, 85, 220–227. [Google Scholar] [CrossRef]
- Morrison, K.; Edwards, Y.H.; Lynch, S.A.; Burn, J.; Hol, F.; Mariman, E. Methionine synthase and neural tube defects. J. Med. Genet. 1997, 34, 958–960. [Google Scholar]
- Shaw, G.M.; Todoroff, K.; Finnell, R.H.; Lammer, E.J.; Leclerc, D.; Gravel, R.A.; Rozen, R. Infant methionine synthase variants and risk for spina bifida. J. Med. Genet. 1999, 36, 86–87. [Google Scholar]
- Van der Put, N.M.; van der Molen, E.F.; Kluijtmans, L.A.; Heil, S.G.; Trijbels, J.M.; Eskes, T.K.; van Oppenraaij-Emmerzaal, D.; Banerjee, R.; Blom, H.J. Sequence analysis of the coding region of human methionine synthase: Relevance to hyperhomocysteinaemia in neural-tube defects and vascular disease. QJM 1997, 90, 511–517. [Google Scholar] [CrossRef]
- Zhu, H.; Wicker, N.J.; Shaw, G.M.; Lammer, E.J.; Hendricks, K.; Suarez, L.; Canfield, M.; Finnell, R.H. Homocysteine remethylation enzyme polymorphisms and increased risks for neural tube defects. Mol. Genet. Metab. 2003, 78, 216–221. [Google Scholar] [CrossRef]
- Doolin, M.T.; Barbaux, S.; McDonnell, M.; Hoess, K.; Whitehead, A.S.; Mitchell, L.E. Maternal genetic effects, exerted by genes involved in homocysteine remethylation, influence the risk of spina bifida. Am. J. Hum. Genet. 2002, 71, 1222–1226. [Google Scholar] [CrossRef]
- Ouyang, S.; Li, Y.; Liu, Z.; Chang, H.; Wu, J. Association between MTR A2756G and MTRR A66G polymorphisms and maternal risk for neural tube defects: A meta-analysis. Gene 2013, 515, 308–312. [Google Scholar] [CrossRef]
- Selhub, J.; Paul, L. Folic acid fortification: Why not vitamin B12 also? Biofactors 2011, 37, 269–271. [Google Scholar] [CrossRef]
- Benevenga, N.J. Consideration of betaine and one-carbon sources of N5-methyltetrahydrofolate for use in homocystinuria and neural tube defects. Am. J. Clin. Nutr. 2007, 85, 946–949. [Google Scholar]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Imbard, A.; Benoist, J.-F.; Blom, H.J. Neural Tube Defects, Folic Acid and Methylation. Int. J. Environ. Res. Public Health 2013, 10, 4352-4389. https://doi.org/10.3390/ijerph10094352
Imbard A, Benoist J-F, Blom HJ. Neural Tube Defects, Folic Acid and Methylation. International Journal of Environmental Research and Public Health. 2013; 10(9):4352-4389. https://doi.org/10.3390/ijerph10094352
Chicago/Turabian StyleImbard, Apolline, Jean-François Benoist, and Henk J. Blom. 2013. "Neural Tube Defects, Folic Acid and Methylation" International Journal of Environmental Research and Public Health 10, no. 9: 4352-4389. https://doi.org/10.3390/ijerph10094352