Arsenic-Induced Genotoxic and Cytotoxic Effects in Human Keratinocytes, Melanocytes and Dendritic Cells


Abstract
:Introduction
Materials and Methods
Reagents and Cell Lines
Cell Treatment for Acute Cytotoxicity
Cell Treatment for Chronic Cytotoxicity
Cell Treatment for Genotoxicity
Statistical Analysis
Results
Cytotoxicity Assay

{kind=link}
{kind=link}
{kind=link}
| Cell type | Exposure type | LD10 (ppm) | LD25 (ppm) |
|---|---|---|---|
| Keratinocytes | Acute | 0.38 | 3.00 |
| Chronic | 0.40 | 0.80 | |
| Melanocytes | Acute | 0.19 | 0.38 |
| Chronic | 0.10 | 0.25 | |
| Dendritic cells | Acute | 0.38 | 0.75 |
| Chronic | 0.10 | 1.00 |
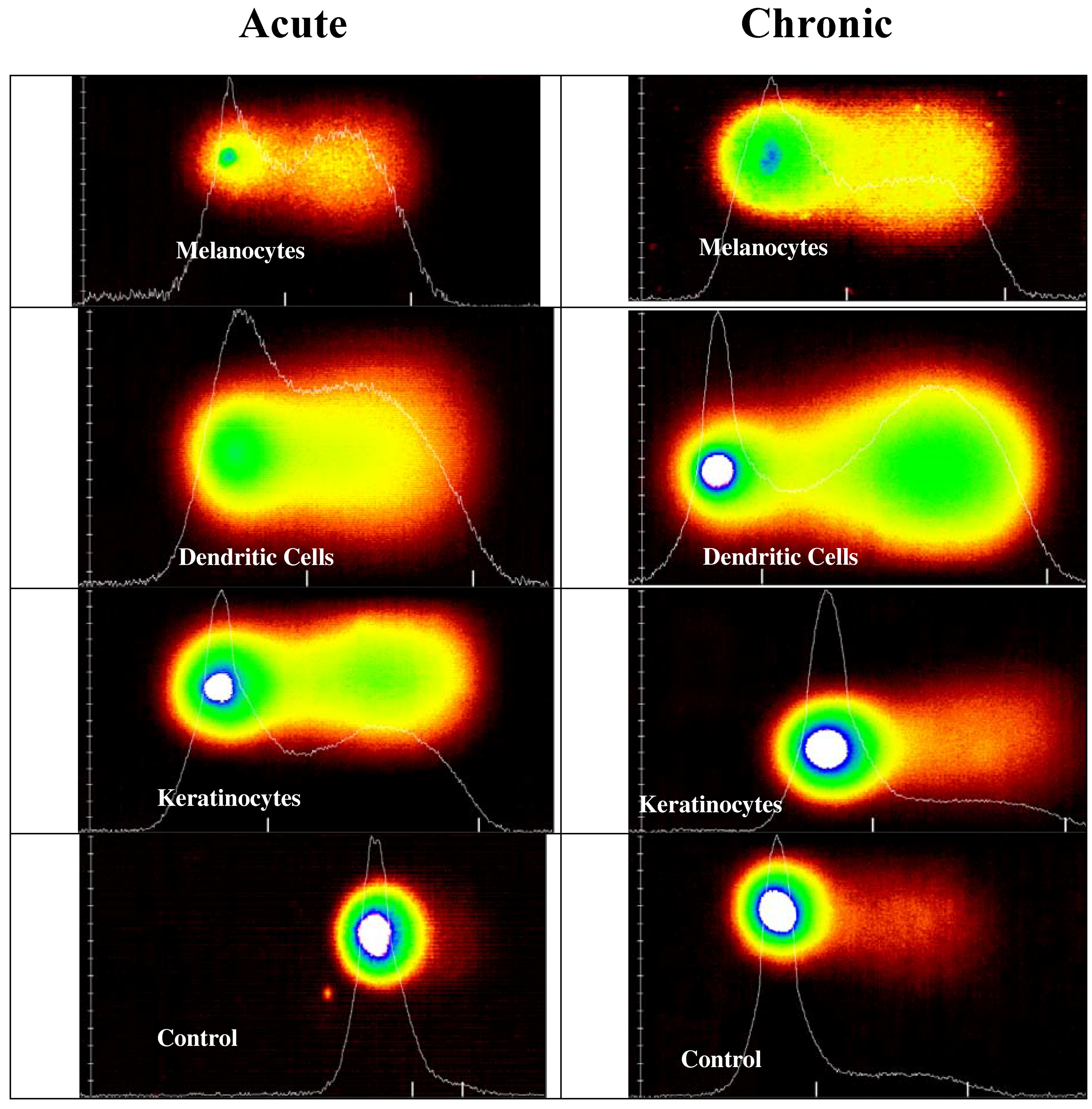
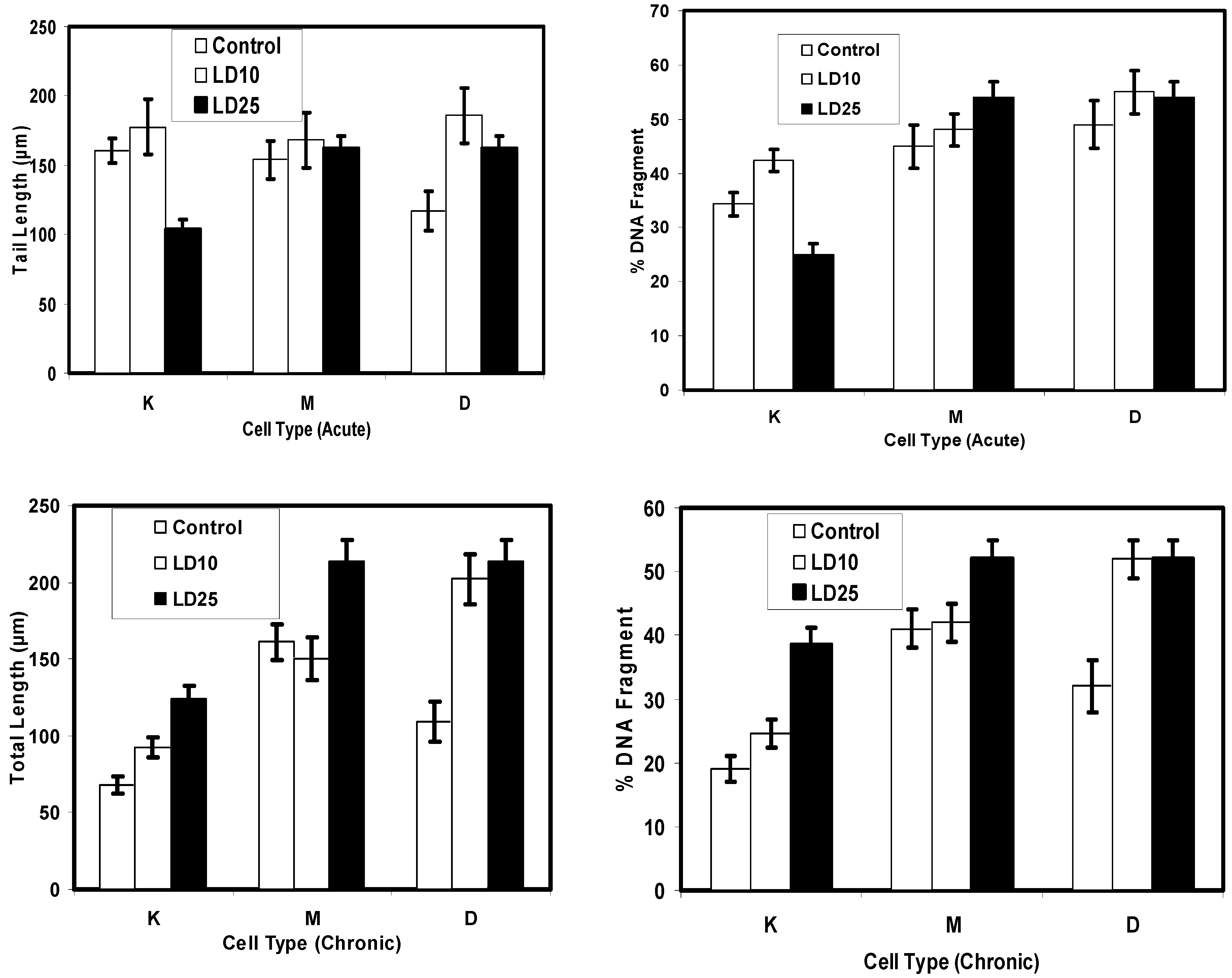
Comet Assay for Genotoxicity
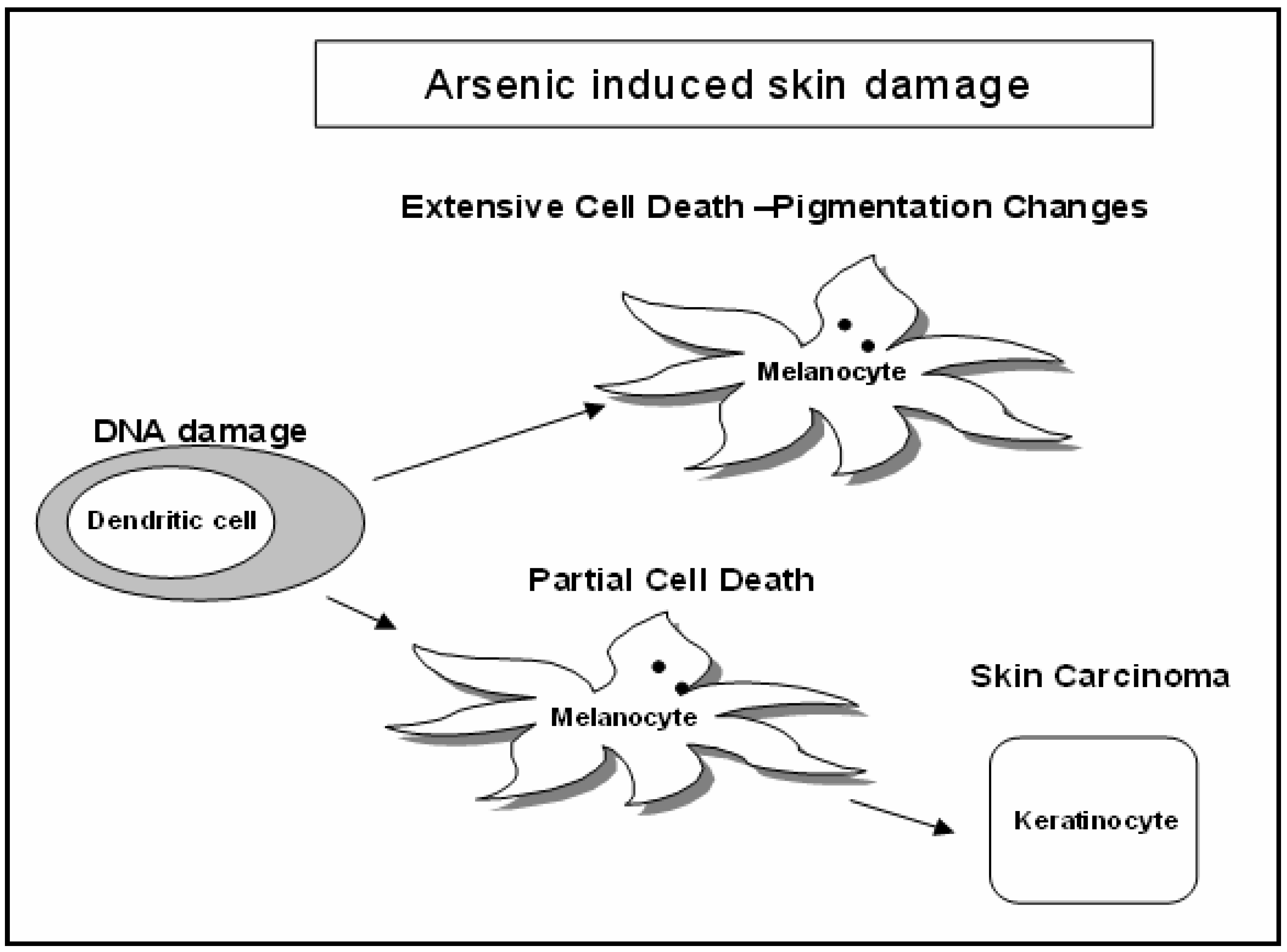
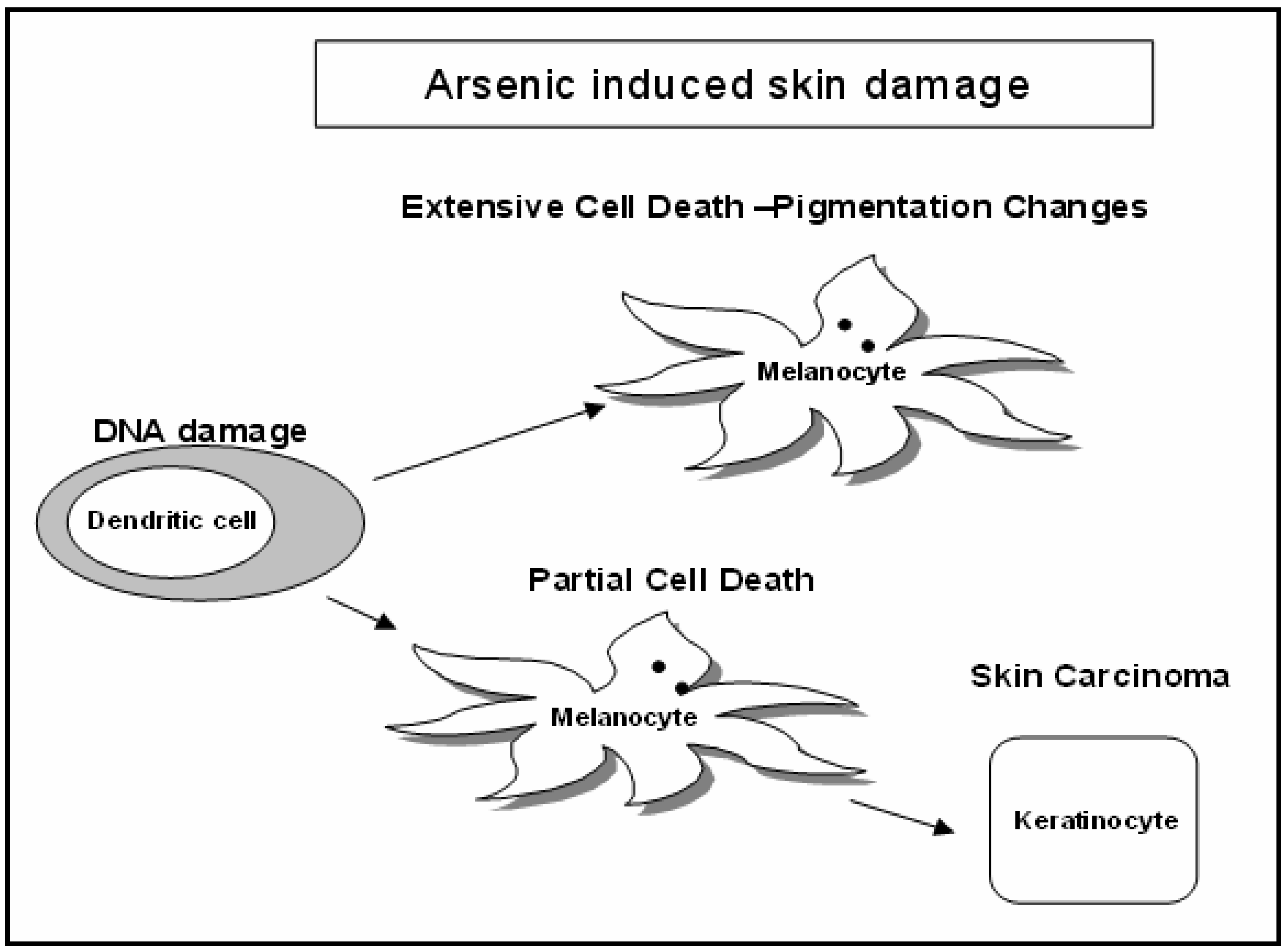
Discussion
Cytotoxicity Assay

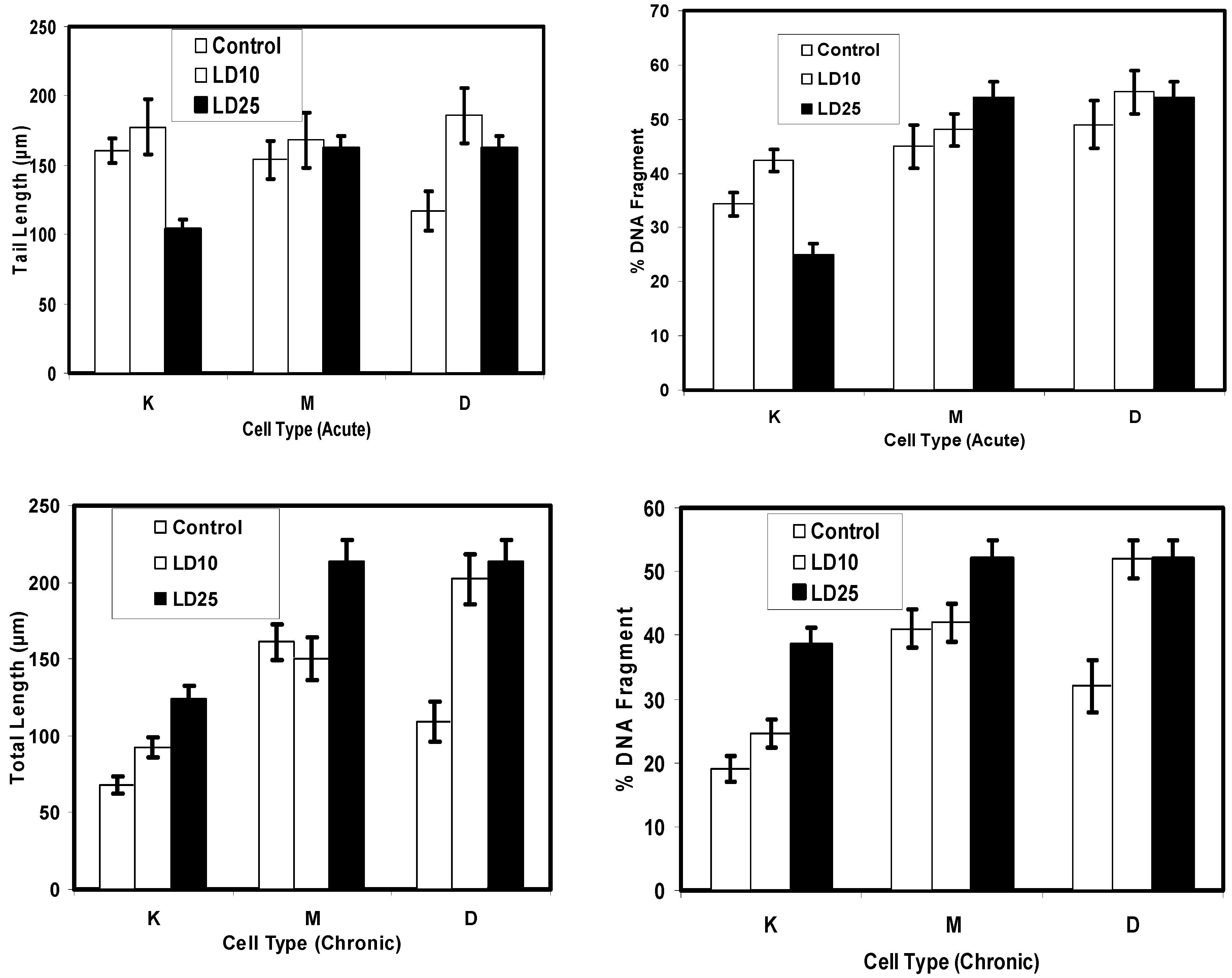
Comet Assay for Genotoxicity
Acknowledgements
References
- Agency for Toxic Substances and Disease Registry (ATSDR). Toxfacts for arsenic; (Update); U.S. Department of Health and Human Services, Public Health Service: Atlanta, GA, 2000. [Google Scholar]
- Tchounwou, P. B.; Centeno, J, A.; Patlolla, A. K. Arsenic toxicity, mutagenesis and carcinogenesis: a health risk assessment and management approach. Mol. Cell Biochem. 2004, 255, 47–55. [Google Scholar] [CrossRef]
- Ali, A.; Lock, J. Physiological and toxicological changes in the skin result from the action and interaction of metal ions. Crit. Rev. Toxicol. 1995, 25, 397–462. [Google Scholar] [CrossRef]
- Chiang, M. C.; Yih, L. H.; Haung, R. N.; Peck, K.; Lee, T. C. Tumor formation of immortalized HaCaT cells in nude mice by long term exposure to sodium arsenite at non toxic doses. Toxicol. 2001, 164, 95(P2A21). [Google Scholar]
- Tchou, I.; Sabido, O.; Lambert, C.; Misery, L.; Garraud, O.; Genin, C. Technique for obtaining highly enriched, quiescent immature Langerhans cells suitable for ex vivo assays. Immunol. Lett. 2003, 86((1)), 7–14. [Google Scholar]
- Chen, C-S. J.; Siegel, D. M. Arsenical Keratosis. eMedicine J. 2002, 2((6)). [Google Scholar]
- www.mgs.bionet.nsc.ru/mgs/gnw/trrd/theraurus/Sk/skin.html.
- Gradecka, D.; Palus, J.; Wasowicz, W. Selected mechanisms of genotoxic effects of inorganic arsenic compounds. Int. J. Occup. Med. Environ. Health 2001, 14((4)), 317–328. [Google Scholar]
- Hei, T. Free Radicals Mediate Arsenic’s Harmful Effects. 2002; Online at hsdnews@columbia.edu. [Google Scholar]
- Gebel, T. W. Genotoxicity of arsenical compounds. Int J Hyg Environ Health 2001, 203((3)), 249–62. [Google Scholar] [CrossRef]
- Waclavicek, M.; Berer, A.; Oehler, L.; Stockl, J.; Schloegl, E.; Majdic, O.; Knapp, W. Calcium ionophore: a single reagent for the differentiation of primary human acute myelogenous leukaemia cells towards dendritic cells. Br J Haematol 2001, 114((2)), 466–73. [Google Scholar]
- Graham-Evans, B.; Tchounwou, Paul B.; Cohly, Hari H. P. Cytotoxicity and Proliferation Studies with Arsenic in Established Human Cell Lines: Keratinocytes, Melanocytes, Dendritic Cells, Dermal Fibroblasts, Microvascular Endothelial Cells, Monocytes and T-Cells. Int. J. Mol. Sci. 2003, 4, 13–21. [Google Scholar]
- Bae, D. S.; Gennings, C.; Carter, W. H., Jr.; Yang, R. S.; Campain, J. A. Toxicological interactions among arsenic, cadmium, chromium, and lead in human keratinocytes. Toxicol. Sci. 2001, 63, 132–142. [Google Scholar] [CrossRef]
- Bau, D. T.; Wang, T. S.; Chung, C. H.; Wang, A. S.; Wang, A. S.; Jan, K. Y. Oxidative DNA adducts and DNA-protein cross-links are the major DNA lesions induced by arsenite. Environ. Health Perspect. 2002, 110((5)), 753–756. [Google Scholar]
- Bernstam, L.; Lan, C. H.; Lee, J.; Nriagu, J. O. Effects of arsenic on human keratinocytes, morphological, physiological, and precursor incorporation studies. Environ. Res. 2002, 3, 220–235. [Google Scholar] [CrossRef]
- Latinwo, L.; Ikediobi, C.; Singh, N. P.; Sponholtz, G.; Fasanya, C.; Riley, L. Comparative studies of in vivo genotoxic effects of cadmium chloride in rat brain, kidney and liver cells. Cellular and Molecular Biology 1997, 43((2)), 203–210. [Google Scholar]
- Hamadeh, H. K.; Trouba, K. J.; Amin, R. P.; Afshari, C. A.; Germolec, D. Coordination of altered DNA repair and damage pathways in arsenite-exposed keratinocytes. Toxicol Sci 2002, 69((2)), 306–16. [Google Scholar]
- Chen, G. Q.; Zhu, J.; Shi, X. G.; Ni, J. H.; Zhong, H. J.; Si, G. Y.; Jin, X. L.; Tang, W.; Li, X. S.; Xong, S. M.; Shen, Z. X.; Sun, G. L.; Ma, J.; Zhang, P.; Zhang, T. D.; Gazin, C.; Naoe, T.; Chen, S. J.; Wang, Z. Y.; Chen, Z. In vitro studies on cellular and molecular mechanisms of arsenic trioxide (As2O3) in the treatment of acute promyelocytic leukemia: As2O3 induces NB4 cell apoptosis with down regulation of Bcl-2 expression and modulation of PML-RAR alpha/PML proteins. Blood 1996, 88, 1052–1061. [Google Scholar]
- Fischer, A. B.; Buchet, J. P.; Lauwerys, R. R. Arsenic uptake, cytotoxicity and detoxification studied in mammalian cells in culture. Arch. Toxicol. 1985, 57((3)), 168–72. [Google Scholar] [CrossRef]
- National Academy Press. Arsenic in Drinking Water: 2001 Update, 2001; Online at: www.nap.edu.
- Saleha Banu, B.; Danadevi, K.; Jamil, K.; Ahuja, Y. R.; Visweswara; Rao, K.; Ishaq, M. In vivo genotoxic effect of arsenic trioxide in mice using comet assay. Toxicology 2001, 162((3)), 171–7. [Google Scholar]
- Nelms, E. Measuring Apoptosis in Individual Cells with the Comet Assay. Promega Notes. 1997, 64, 13. [Google Scholar]
© 2004 MDPI. All rights reserved.
Share and Cite
Graham-Evans, B.; Cohly, H.H.P.; Yu, H.; Tchounwou, P.B. Arsenic-Induced Genotoxic and Cytotoxic Effects in Human Keratinocytes, Melanocytes and Dendritic Cells. Int. J. Environ. Res. Public Health 2004, 1, 83-89. https://doi.org/10.3390/ijerph2004020083
Graham-Evans B, Cohly HHP, Yu H, Tchounwou PB. Arsenic-Induced Genotoxic and Cytotoxic Effects in Human Keratinocytes, Melanocytes and Dendritic Cells. International Journal of Environmental Research and Public Health. 2004; 1(2):83-89. https://doi.org/10.3390/ijerph2004020083
Chicago/Turabian StyleGraham-Evans, Barbara, Hari H. P. Cohly, Hongtao Yu, and Paul B. Tchounwou. 2004. "Arsenic-Induced Genotoxic and Cytotoxic Effects in Human Keratinocytes, Melanocytes and Dendritic Cells" International Journal of Environmental Research and Public Health 1, no. 2: 83-89. https://doi.org/10.3390/ijerph2004020083
APA StyleGraham-Evans, B., Cohly, H. H. P., Yu, H., & Tchounwou, P. B. (2004). Arsenic-Induced Genotoxic and Cytotoxic Effects in Human Keratinocytes, Melanocytes and Dendritic Cells. International Journal of Environmental Research and Public Health, 1(2), 83-89. https://doi.org/10.3390/ijerph2004020083