The Relevance of Marine Chemical Ecology to Plankton and Ecosystem Function: An Emerging Field



 ,
,  and
and
Abstract
:1. Introduction
2. Chemical Cues and Phytoplankton-Zooplankton Interactions
3. Chemical Identification of Allelochemicals in Phytoplankton
4. Can Concepts of Terrestrial Chemical Ecology Be Directly Applied to the Plankton?
5. New Toolboxes to Study Chemical Interactions
6. Conclusions and Future Directions
Acknowledgments
- Samples Availability: Available from the authors.
References
- Beaugrand, G; Reid, PC; Ibañez, F; Lindley, JA; Edwards, M. Reorganisation of North Atlantic marine copepod biodiversity and climate. Science 2002, 296, 1692–1694. [Google Scholar]
- Li, WKW. Macroecological patterns of phytoplankton in the northwestern North Atlantic Ocean. Nature 2002, 419, 154–157. [Google Scholar]
- Azam, F; Malfatti, F. Microbial structuring of marine ecosystems. Nat. Rev. Microbiol 2007, 5, 782–791. [Google Scholar]
- Smetacek, V; Cloern, JE. Oceans—On phytoplankton trends. Science 2008, 319, 1346–1348. [Google Scholar]
- Hay, ME. Marine chemical ecology: Chemical signals and cues structure marine populations, communities, and ecosystems. Annu. Rev. Mar. Sci 2009, 1, 193–212. [Google Scholar]
- Miralto, A; Barone, G; Romano, G; Poulet, SA; Ianora, A; Russo, GL; Buttino, I; Mazzarella, G; Laabir, M; Cabrini, M; et al. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar]
- Wendel, T; Jüttner, F. Lipoxygenase-mediated formation of hydrocarbons and unsaturated aldehydes in freshwater diatoms. Phytochemistry 1996, 41, 1445–1449. [Google Scholar]
- Fontana, A; d’Ippolito, G; Cutignano, A; Romano, R; Lamari, N; Gallucci, AM; Cimino, G; Miralto, A; Ianora, A. LOX-induced peroxidation mechanism responsible for the detrimental effect of marine diatoms on zooplankton grazers. ChemBioChem 2007, 8, 1810–1818. [Google Scholar]
- d’Ippolito, G; Lamari, N; Montresor, M; Romano, G; Cutignano, A; Gerecht, A; Cimino, G; Fontana, A. 15S-Lipoxygenase metabolism in the marine diatom Pseudo-nitzschia delicatissima. New Phytol 2009, 183, 1064–1071. [Google Scholar]
- Jüttner, F; Messina, P; Patalano, C; Zupo, V. Odour compounds of the diatom Cocconeis scutellum: Effects on benthic herbivores living on Posidonia oceanica. Mar. Ecol. Prog. Ser 2010, 400, 63–73. [Google Scholar]
- Ianora, A; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: A review. Ecotoxicology 2010, 19, 493–511. [Google Scholar]
- Pohnert, G. Chemical defense strategies of marine organisms. Top. Curr. Chem 2004, 239, 179–219. [Google Scholar]
- Caldwell, GS. The influence of bioactive oxylipins from marine diatoms on invertebrate reproduction and development. Mar. Drugs 2009, 7, 367–400. [Google Scholar]
- Leflaive, J; Ten-Hage, L. Chemical interactions in diatoms: Role of polyunsaturated aldehydes and precursors. New Phytol 2009, 184, 794–805. [Google Scholar]
- Van Donk, E; Ianora, A; Vos, M. Induced defences in marine and freshwater phytoplankton: A review. Hydrobiologia 2011, 668, 3–19. [Google Scholar]
- Ianora, A; Miralto, A; Poulet, SA; Carotenuto, Y; Buttino, I; Romano, G; Casotti, R; Pohnert, G; Wichard, T; Colucci-D’Amato, L; et al. Aldehyde suppression of copepod recruitment in blooms of a ubiquitous planktonic diatom. Nature 2004, 429, 403–407. [Google Scholar]
- Ribalet, F; Berges, JA; Ianora, A; Casotti, R. Growth inhibition of cultured marine phytoplankton by algal-derived polyunsaturated aldehydes. Aquat. Toxicol 2007, 85, 219–227. [Google Scholar]
- Vardi, A; Formiggini, F; Casotti, R; de Martino, A; Ribalet, F; Miralro, A; Bowler, C. A stress surveillance system based on calcium and nitric oxide in marine diatoms. PLoS Biol 2006, 4, 411–419. [Google Scholar]
- Ribalet, F; Intertaglia, L; Lebaron, P; Casotti, R. Differential effect of three polyunsaturated aldehydes on marine bacterial isolates. Aquat. Toxicol 2008, 86, 249–255. [Google Scholar]
- Balestra, C; Alonso-Sáez, L; Gasol, JM; Casotti, R. Group-specific effects on coastal bacterioplankton of polyunsaturated aldehydes produced by diatoms. Aquat. Microb. Ecol 2011, 63, 123–131. [Google Scholar] [Green Version]
- Vidoudez, C; Pohnert, G. Growth phase-specific release of polyunsaturated aldehydes by the diatom Skeletonema marinoi. J. Plankton Res 2008, 30, 1305–1313. [Google Scholar]
- Vidoudez, C; Casotti, R; Bastianini, M; Pohnert, G. Quantification of dissolved and particulate polyunsaturated aldehydes in the Adriatic Sea. Mar. Drugs 2011, 9, 500–513. [Google Scholar]
- Wichard, T; Poulet, SA; Halsband-Lenk, C; Albaina, A; Harris, R; Liu, D; Pohnert, G. Survey of the chemical defense potential of diatoms: Screening of 51 species for α,β,λ,δ-unsaturated aldehydes. J. Chem. Ecol 2005, 31, 949–958. [Google Scholar]
- d’Ippolito, G; Iadicicco, I; Romano, G; Fontana, A. Detection of short-chain aldehydes in marine organisms: The diatom Thalassiosira rotula. Tetrahedron Lett 2002, 43, 6137–6140. [Google Scholar]
- Fontana, A; d’Ippolito, G; Cutignano, A; Miralto, A; Ianora, A; Romano, G; Cimino, G. Chemistry of oxylipin pathways in marine diatoms. Pure Appl. Chem 2007, 79, 481–490. [Google Scholar]
- Ianora, A; Romano, G; Carotenuto, Y; Esposito, F; Roncalli, V; Buttino, I; Miralto, A. Impact of the diatom oxylipin 15S-HEPE on the reproductive success of the copepod Temora stylifera. Hydrobiologia 2011, 666, 265–275. [Google Scholar]
- Adolph, S; Bach, S; Blondel, B; Cueff, A; Moreau, M; Pohnert, G; Poulet, SA; Wichard, T; Zuccaro, A. Cytotoxicity of diatom-derived oxylipins in organisms belonging to different phyla. J. Exp. Biol 2004, 207, 2935–2946. [Google Scholar]
- Steinke, M; Stefels, J; Stamhuis, E. Dimethyl sulfide triggers search behavior in copepods. Limnol. Oceanogr 2006, 51, 1925–1930. [Google Scholar]
- Shaw, BA; Andersen, RJ; Harrison, PJ. Feeding deterrence properties of apo-fucoxanthinoids from marine diatoms. 1. Chemical structures of apo-fucoxanthinoids produced by Phaeodactylum tricornutum. Mar. Biol 1995, 124, 467–472. [Google Scholar]
- Shaw, BA; Harrison, PJ; Andersen, RJ. Feeding deterrence properties of apo-fucoxanthinoids from marine diatoms. 2. Physiology of production of apo-fucoxanthinoids by the marine diatoms Phaeodactylum tricornutum and Thalassiosira pseudonana, and their feeding deterrent effects on the copepod Tigriopus californicus. Mar. Biol 1995, 124, 473–481. [Google Scholar]
- Selander, E; Thor, P; Toth, G; Pavia, H. Copepods induce paralytic shellfish toxin production in marine dinoflagellates. Proc. R. Soc. B 2006, 273, 1673–1680. [Google Scholar]
- Waggett, RJ; Tester, PA; Place, AR. Anti-grazing properties of the toxic dinoflagellate Karlodinium veneficum during predator-prey interactions with the copepod Acartia tonsa. Mar. Ecol. Prog. Ser 2008, 366, 31–42. [Google Scholar]
- Ma, H; Krock, B; Tillmann, U; Cembella, A. Preliminary characterization of extracellular allelochemicals of the toxic marine dinoflagellate Alexandrium tamarense using a Rhodomonas salina assay. Mar. Drugs 2009, 7, 497–522. [Google Scholar]
- Wolfe, GV; Steinke, M; Kirst, GO. Grazing-activated chemical defense in a unicellular marine alga. Nature 1997, 387, 894–897. [Google Scholar]
- Fredrickson, KA; Strom, SL. The algal osmolyte DMSP as a microzooplankton grazing deterrent in laboratory and field studies. J. Plankton Res 2009, 31, 135–152. [Google Scholar]
- Wolfe, GV; Strom, SL; Holmes, JL; Radzio, T; Olson, MB. Dimethylsulfoniopropionate cleavage by marine phytoplankton in response to mechanical, chemical, or dark stress. J. Phycol 2002, 38, 948–960. [Google Scholar]
- Nejstgaard, JC; Gismervik, I; Solberg, PT. Feeding and reproduction by Calanus finmarchicus, and microzooplankton grazing during mesocosm blooms of diatoms and the coccolithophore Emiliania huxleyi. Mar. Ecol. Prog. Ser 1997, 147, 197–217. [Google Scholar]
- Strom, SL; Brainard, MA; Holmes, JL; Olson, MB. Phytoplankton blooms are strongly impacted by microzooplankton grazing in coastal North Pacific. Mar. Biol 2003, 138, 355–368. [Google Scholar]
- Tang, KW; Visscher, PT; Dam, HG. DMSP-consuming bacteria associated with the calanoid copepod Acartia tonsa (Dana). J. Exp. Mar. Biol. Ecol 2001, 256, 185–198. [Google Scholar]
- Turner, JT; Ianora, A; Esposito, F; Carotenuto, Y; Miralto, A. Zooplankton feeding ecology: Does a diet of Phaeocystis support good copepod grazing, survival, egg production and egg hatching success? J. Plankton Res 2002, 24, 1185–1195. [Google Scholar]
- Steinke, M; Malin, G; Liss, PS. Trophic interactions in the sea: An ecological role for climate relevant volatiles? J. Phycol 2002, 38, 630–638. [Google Scholar]
- Pohnert, G; Steinke, M; Tollrian, R. Chemical cues, defense metabolites and the shaping of pelagic interspecific interactions. Trends Ecol. Evol 2007, 22, 198–204. [Google Scholar]
- Nevitt, GA. Olfactory foraging by Antarctic procellariiform seabirds: Life at high Reynolds numbers. Biol. Bull 2000, 198, 245–253. [Google Scholar]
- Wolfe, GV. The chemical defense ecology of marine unicellular plankton: Constraints, mechanisms, and impacts. Biol. Bull 2000, 198, 225–244. [Google Scholar]
- Hansen, E; Ernsten, A; Eilersten, HC. Isolation and characterisation of cytotoxic polyunsaturated aldehyde from the marine phytoplankter Phaeocystis pouchetii (Hariot) Lagerheim. Toxicology 2004, 199, 207–217. [Google Scholar]
- Sykes, PF; Huntley, ME. Acute physiological reactions of Calanus pacificus to selected dinoflagellates–direct observations. Mar. Biol 1987, 94, 19–24. [Google Scholar]
- Teegarden, GJ; Cembella, AD. Grazing of toxic dinoflagellates, Alexandrium spp., by adult copepods of coastal Maine: Implications for the fate of paralytic shellfish toxins in marine food webs. J. Exp. Mar. Biol. Ecol 1996, 196, 145–176. [Google Scholar]
- Guisande, C; Frangopulos, M; Carotenuto, Y; Maneiro, I; Riveiro, I. Fate of paralytic shellfish poisoning toxins ingested by the copepod Acartia clausi. Mar. Ecol. Prog. Ser 2002, 240, 105–115. [Google Scholar]
- Frangopulos, M; Guisande, C; Maneiro, I; Riveiro, I; Franco, J. Short-term and long-term effects of the toxic dinoflagellate Alexandrium minutum on the copepod Acartia clausi. Mar. Ecol. Prog. Ser 2000, 203, 161–169. [Google Scholar]
- Turner, JT; Tester, PA. Toxic marine phytoplankton, zooplankton grazers, and pelagic food webs. Limnol. Oceanogr 1997, 42, 1203–1214. [Google Scholar]
- Turriff, N; Runge, JA; Cembella, AD. Toxin accumulation and feeding behavior of the planktonic copepod Calanus finmarchicus exposed to the red tide dinoflagellate Alexandrium excavatum. Mar. Biol 1995, 123, 55–64. [Google Scholar]
- Cembella, AD. Chemical ecology of eukaryotic microalgae in marine ecosystems. Phycologia 2003, 42, 420–447. [Google Scholar]
- Bergkvist, J; Selander, E; Pavia, H. Induction of toxin production in dinoflagellates: The grazer makes a difference. Oecologia 2008, 156, 147–154. [Google Scholar]
- Selander, E; Cervin, G; Pavia, H. Effects of nitrate and phosphate on grazer-induced toxin production in Alexandrium minitum. Limnol. Oceanogr 2008, 53, 523–530. [Google Scholar]
- Selander, E; Jakobsen, HH; Lombard, F; Kiørboe, T. Grazer cues induce stealth behavior in marine dinoflagellates. Proc. Natl. Acad. Sci. USA 2011, 108, 4030–4034. [Google Scholar]
- Wohirab, S; Iversen, MH; John, U. A molecular and co-evolutionary context for grazer induced toxin production in Alexandrium tamarense. PLoS One 2010, 5, e15039. [Google Scholar]
- Yang, I; Selander, E; Pavia, H; John, U. Grazer-induced toxin formation in dinoflagellates: A transcriptomic model study. Eur. J. Phycol 2011, 46, 66–73. [Google Scholar]
- Baden, DG. Brevetoxins: Unique polyether dinoflagellate toxins. FASEB J 1989, 3, 1807–1817. [Google Scholar]
- Prince, EK; Lettieri, L; McCurdy, KJ; Kubanek, J. Fitness consequences for copepods feeding on a red tide dinoflagellate: Deciphering the effects of nutritional value, toxicity, and feeding behavior. Oecologia 2006, 147, 479–488. [Google Scholar]
- Kubanek, J; Snell, TW; Pirkle, C. Chemical defense of the red tide dinoflagellate Karenia brevis against rotifer grazing. Limnol. Oceanogr 2007, 52, 1026–1035. [Google Scholar]
- Manning, SR; La Claire, JW. Prymnesins: Toxic metabolites of the golden alga, Prymnesium parvum Carter (Haptophyta). Mar. Drugs 2010, 8, 678–704. [Google Scholar]
- Sopanen, S; Koski, M; Kuuppo, P; Uronen, P; Legrand, C; Tamminen, T. Toxic haptophyte Prymnesium parvum affects grazing, survival, egestion and egg production of the calanoid copepods Eurytemora affinis and Acartia bifilosa. Mar. Ecol. Prog. Ser 2006, 327, 223–232. [Google Scholar]
- Sopanen, S; Koski, M; Uronen, P; Kuuppo, P; Lehtinen, S; Legrand, C; Tamminen, T. Prymnesium parvum exotoxins affect the grazing and viability of the calanoid copepod Eurytemora affinis. Mar. Ecol. Prog. Ser 2008, 361, 191–202. [Google Scholar]
- Poulson, KL; Sieg, D; Kubanek, J. Chemical ecology of the marine plankton. Nat. Prod. Rep 2009, 26, 729–745. [Google Scholar]
- Sieg, RD; Poulson-Ellestad, KL; Kubanek, J. Chemical ecology of marine plankton. Nat. Prod. Rep 2011, 28, 388–399. [Google Scholar]
- Lürling, M; Scheffer, M. Info-disruption: Pollution and the transfer of chemical information between organisms. Trends Ecol. Evol 2007, 22, 374–379. [Google Scholar]
- Vogt, M; Steinke, M; Turner, S; Paulino, A; Meyerhöfer, M; Riebesell, U; LeQuéré, C; Liss, P. Dynamics of dimethylsulphoniopropionate and dimethylsulphide under different CO2 concentrations during a mesocosm experiment. Biogeosciences 2008, 5, 407–419. [Google Scholar]
- Sotka, EE; Whalen, KE. Herbivore Offense in the Sea: The Detoxification and Transport of Secondary Metabolites. In Algal Chemical Ecology; Amsler, CD, Ed.; Springer-Verlag: Berlin, Germany, 2008; pp. 203–228. [Google Scholar]
- Colin, SP; Dam, HG. Effects of the toxic dinoflagellate Alexandrium fundyense on the copepod Acartia hudsonica: A test of the mechanisms that reduce ingestion rates. Mar. Ecol. Prog. Ser 2003, 248, 55–65. [Google Scholar]
- Colin, SP; Dam, HG. Testing for resistance of pelagic marine copepods to a toxic dinoflagellate. Evol. Ecol 2005, 18, 355–377. [Google Scholar]
- Tester, PA; Turner, JT; Shea, D. Vectorial transport of toxins from the dinoflagellate Gymnodinium breve through copepods to fish. J. Plankton Res 2000, 22, 47–61. [Google Scholar]
- Doucette, GJ; Turner, JT; Powell, CL; Keafer, BA; Anderson, DM. Trophic accumulation of PSP toxins in zooplankton during Alexandrium fundyense blooms in Casco Bay, Gulf of Maine, April-June 1998. 1. Toxin level in A. fundyense and zooplankton size fractions. Deep Sea Res. II Trop. Stud. Oceanogr 2005, 52, 19–21. [Google Scholar]
- Blanco, J; Reyero, I; Franco, J. Kinetics of accumulation and transformation of paralytic shellfish toxins in the blue mussel Mytilus galloprovincialis. Toxicon 2003, 42, 777–784. [Google Scholar]
- Buttino, I; de Rosa, G; Carotenuto, Y; Mazzella, M; Ianora, A; Esposito, F; Vitiello, V; Quaglia, F; La Rotonda, MI; Miralto, A. Aldehyde-encapsulated liposomes impair marine grazer survivorship. J. Exp. Biol 2008, 211, 1426–1433. [Google Scholar]
- Caldwell, GS; Watson, SB; Bentley, MG. How to assess toxin ingestion and postingestion partitioning in zooplankton? J. Plankton Res 2004, 26, 1369–1377. [Google Scholar]
- Legrand, C; Rengefors, K; Fistarol, GO; Granéli, E. Allelopathy in phytoplankton—Biochemical, ecological and evolutionary aspects. Phycologia 2003, 42, 406–419. [Google Scholar]
- Prince, EK; Poulson, KL; Myers, TL; Sieg, RD; Kubanek, J. Characterization of allelopathic compounds from the red tide dinoflagellate Karenia brevis. Harmful Algae 2010, 10, 39–48. [Google Scholar]
- Tillmann, U; John, U. Toxic effects of Alexandrium spp. on heterotrophic dinoflagellates: An allochemical defence mechanism independent of PSP-toxin content. Mar. Ecol. Prog. Ser 2002, 230, 47–58. [Google Scholar]
- Tillmann, U; John, U; Cembella, A. On the allelochemical potency of the marine dinoflagellate Alexandrium ostenfeldii against heterotrophic and autotrophic protists. J. Plankton Res 2007, 29, 527–543. [Google Scholar]
- Tillmann, U; Hansen, PJ. Allelopathic effects of Alexandrium tamarense on other algae: Evidence from mixed growth experiments. Aquat. Microb. Ecol 2009, 57, 101–112. [Google Scholar]
- Van Wagoner, RM; Deeds, JR; Satake, M; Ribeiro, AA; Place, AR; Wright, JLC. Isolation and characterization of karlotoxin 1, a new amphipathic toxin from Karlodinium veneficum. Tetrahedron Lett 2008, 49, 6457–6461. [Google Scholar]
- Yamasaki, Y; Shikata, T; Nukata, A; Ichiki, S; Nagasoe, S; Matsubara, T; Shimasaki, Y; Nakao, M; Yamaguchi, K; Oshima, Y; et al. Extracellular polysaccharide-protein complexes of a harmful alga mediate the allelopathic control it exerts within the phytoplankton community. ISME J 2009, 3, 808–817. [Google Scholar]
- Kubanek, J; Hicks, MK; Naar, J; Villareal, TA. Does the red tide dinoflagellate Karenia brevis use allelopathy to outcompete other phytoplankton? Limnol. Oceanogr 2005, 50, 883–895. [Google Scholar]
- Prince, EK; Myers, TL; Kubanek, J. Effects of harmful algal blooms on competitors: Allelopathic mechanisms of the red tide dinoflagellate Karenia brevis. Limnol. Oceanogr 2008, 53, 531–541. [Google Scholar]
- Prince, EK; Myers, TL; Naar, J; Kubanek, J. Competing phytoplankton undermines allelopathy of a bloom-forming dinoflagellate. Proc. R. Soc. B 2008, 275, 2733–2741. [Google Scholar]
- Poulson, KL; Sieg, RD; Prince, EK; Kubanek, J. Allelopathic compounds of a red tide dinoflagellate have species-specific and context-dependent impacts on phytoplankton. Mar. Ecol. Prog. Ser 2010, 416, 69–78. [Google Scholar]
- Shilo, M. The Toxic Principles of Prymnesium parvum. In The Water Environment: Algal Toxins and Health; Carmichael, WW, Ed.; Plenum Press: New York, NY, USA, 1981; Volume 20, pp. 37–47. [Google Scholar]
- Fistarol, GO; Legrand, C; Granéli, E. Allelopathic effect of Prymnesium parvum on a natural plankton community. Mar. Ecol. Prog. Ser 2003, 255, 115–125. [Google Scholar]
- Igarashi, T; Satake, M; Yasumoto, T. Prymnesin-2: A potent ichtyotoxic and hemolytic glycoside isolated from the red tide alga Prymnesium parvum. J. Am. Chem. Soc 1996, 118, 479–480. [Google Scholar]
- Schug, KA; Skingel, TR; Spencer, SE; Serrano, CA; Le, CQ; Schug, CA; Valenti, TW; Brooks, BW; Mydlarz, LD; Grover, JP. Hemolysis, fish mortality, and LC-ESI-MS of cultured crude and fractionated golden alga (Prymnesium parvum). J. Am. Water Resour. Assoc 2010, 46, 33–44. [Google Scholar]
- Ribalet, F; Wichard, T; Pohnert, G; Ianora, A; Miralto, A; Casotti, R. Age and nutrient limitation enhance polyunsaturated aldehyde production in marine diatoms. Phytochemistry 2007, 68, 2059–2067. [Google Scholar]
- Gross, EM. Allelopathy of aquatic autotrophs. Crit. Rev. Plant Sci 2003, 22, 313–339. [Google Scholar]
- Hamm, CE; Merkel, R; Springer, O; Jurkojc, P; Maier, C; Prechtel, K; Smetacek, V. Diatom cells are mechanically protected by their strong, lightweight, silica shells. Nature 2003, 421, 841–843. [Google Scholar]
- Smetacek, V. A watery arms race. Nature 2001, 411, 745. [Google Scholar]
- Steppuhn, A; Baldwin, IT. Induced Defenses and the Cost-benefit Paradigm. In Induced Plant Resistance to Herbivory; Schaller, A, Ed.; Springer: Dordrecht, The Netherlands, 2008; pp. 61–83. [Google Scholar]
- Bryant, JP; Chapin, FS; Klein, DR. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 1983, 40, 357–368. [Google Scholar]
- Ianora, A; Boersma, M; Casotti, R; Fontana, A; Harder, J; Hoffmann, F; Pavia, H; Potin, P; Poulet, SA; Toth, G. The H.T. odum synthesis essay: New trends in marine chemical ecology. Estuaries Coasts 2006, 29, 531–551. [Google Scholar]
- Granéli, E; Johansson, N. Increase in the production of allelopathic substances by Prymnesium parvum cells grown under N- or P-deficient conditions. Harmful Algae 2003, 2, 135–145. [Google Scholar]
- Uronen, P; Lehtinen, S; Legrand, C; Kuuppo, P; Tamminen, T. Haemolytic activity and allelopathy of the haptophyte Prymnesium parvum in nutrient-limited and balanced growth conditions. Mar. Ecol. Prog. Ser 2005, 299, 137–148. [Google Scholar]
- Wells, ML; Trick, CG; Cochlan, WP; Hughes, MP; Trainer, VL. Domoic acid: The synergy of iron, copper, and the toxicity of diatoms. Limnol. Oceanogr 2005, 50, 1908–1917. [Google Scholar]
- Coley, PD; Bryant, JP; Chapin, FS. Resource availability and plant antiherbivore defense. Science 1985, 230, 895–899. [Google Scholar]
- Loomis, WE. Growth and Differentiation—An Introduction and Summary. In Growth and Differentiation in Plants; Wareing, PF, Phillips, IDJ, Eds.; Pergamon Press: New York, NY, USA, 1981; pp. 1–17. [Google Scholar]
- Gentien, P; Lunven, M; Lazure, P; Youenou, A; Crassous, MP. Motility and autotoxicity in Karenia mikimotoi (Dinophyceae). Philos. Trans. R. Soc. B 2007, 362, 1937–1946. [Google Scholar]
- Karolyi, G; Neufeld, Z; Scheuring, I. Rock-scissors-paper game in a chaotic flow: The effect of dispersion on the cyclic competition of microorganisms. J. Theor. Biol 2005, 236, 12–20. [Google Scholar]
- An, M; Liu, DL; Wul, H; Liu, YH. Allelopathy from A Mathematical Modeling Perspective. In Allelopathy in Sustainable Agriculture and Forestry; Zeng, RS, Mallik, AU, Ming, LS, Eds.; Springer: New York, NY, USA, 2008; pp. 169–186. [Google Scholar]
- Sole, J; Garcia-Ladona, E; Ruardij, P; Estrada, M. Modelling allelopathy among marine algae. Ecol. Model 2005, 183, 373–384. [Google Scholar]
- Roy, S; Bhattacharya, S; Das, P; Chattopadhyay, J. Interaction among non-toxic phytoplankton, toxic phytoplankton and zooplankton: Inferences from field observations. J. Biol. Phys 2007, 33, 1–17. [Google Scholar]
- Mukhopadhyay, A; Chattopadadhyay, J; Tapaswi, PK. A delay differential equations model of plankton allelopathy. Math. Biosci 1998, 149, 167–189. [Google Scholar]
- Brown, SP; Inglis, RF; Taddei, F. Evolutionary ecology of microbial wars: Within-host competition and (incidental) virulence. Evol. Appl 2009, 2, 32–39. [Google Scholar]
- Durrett, R; Levin, S. Allelopathy in spatially distributed populations. J. Theor. Biol 1997, 185, 165–171. [Google Scholar]
- Gardner, A; West, SA; Buckling, A. Bactericins, spite and virulence. Proc. R. Soc. B 2004, 271, 1529–1535. [Google Scholar]
- Hibbing, ME; Fuqua, C; Parsek, MR; Peterson, SB. Bacterial competition: Surviving and thriving in the microbial jungle. Nat. Rev. Microbiol 2010, 8, 15–25. [Google Scholar]
- Karolyi, G; Neufeld, Z; Scheuring, I. Rock-scissors-paper game in a chaotic flow: The effects of dispersion on the cyclic competition of microorganisms. J. Theor. Biol 2005, 236, 12–20. [Google Scholar]
- Tirichine, L; Bowler, C. Decoding algal genomes: Tracing back the history of photosynthetic life on Earth. Plant J 2011, 66, 45–57. [Google Scholar]
- Armbrust, EV; Berges, JA; Bowler, C; Green, BR; Martinez, D; Putnam, NH; Zhou, S; Allen, AE; Apt, KE; Bechner, M; et al. The genome of the diatom Thalassiosira pseudonana: Ecology, evolution, and metabolism. Science 2004, 306, 79–86. [Google Scholar]
- Bowler, C; Allen, AE; Badger, JH; Grimwood, J; Jabbari, K; Kuo, A; Maheswari, U; Martens, C; Maumus, F; Otillar, RP; et al. The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 2008, 456, 239–244. [Google Scholar]
- Vardi, A; Bidie, K; Kwityn, C; Hirsh, DJ; Thompson, SM; Callow, JA; Falkowski, P; Bowler, C. A diatom gene regulating nitric-oxide signaling and susceptibility to diatom-derived aldehydes. Curr. Biol 2008, 18, 895–899. [Google Scholar]
- Siaut, M; Heijde, M; Mangogna, M; Montsant, M; Coesel, S; Allen, A; Manfredonia, A; Falciatore, A; Bowler, C. Molecular toolbox for studying diatom biology in Phaeodactylum tricornutum. Gene 2007, 406, 23–35. [Google Scholar]
- Yang, I; John, U; Beszteri, S; Glöckner, G; Krock, B; Goesmann, A; Cembella, AD. Comparative gene expression in toxic versus non-toxic strains of the marine dinoflagellate Alexandrium minutum. BMC Genomics 2010, 11, 248. [Google Scholar]
- Van Dolah, FM; Lidie, KB; Monroe, EA; Bhattacharya, D; Campbell, L; Doucette, GJ; Kamykowski, D. The Forida red tide dinoflagellate Karenia brevis: New insights into cellular and molecular processes underlying bloom dynamics. Harmful Algae 2009, 8, 562–572. [Google Scholar]
- Alpermann, TJ; Tillmann, U; Beszteri, B; Cembella, AD; John, U. Phenotypic variation and genotypic diversity in a planktonic population of the toxigenic marine dinoflagellate Alexandrium tamarense (Dinophyceae). J. Phycol 2010, 46, 18–32. [Google Scholar]
- Barofsky, A; Vidoudez, C; Pohnert, G. Metabolic profiling reveals growth stage variability in diatom exudates. Limnol. Oceanogr. Methods 2009, 7, 382–390. [Google Scholar]
- Turnbaugh, PJ; Gordon, JI. An invitation to the marriage of metagenomics and metabolomics. Cell 2008, 134, 708–713. [Google Scholar]
- Bruce, TJA; Matthes, MC; Chamberlain, K; Woodcock, CM; Mohib, A; Webster, B; Smart, LE; Birkett, MA; Pickett, JA; Napier, JA. cis-Jasmone induces Arabidopsis genes that affect the chemical ecology of multitrophic interactions with aphids and their parasitoids. Proc. Natl. Acad. Sci. USA 2008, 105, 4553–4558. [Google Scholar]
- Ferrer, RP; Zimmer, RK. Chemical neuroecology and community dynamics. Ann. N.Y. Acad. Sci 2009, 1170, 450–455. [Google Scholar]
- Zimmer, RK; Ferrer, RP. Neuroecology, chemical defense, and the keystone species concept. Biol. Bull 2007, 213, 208–225. [Google Scholar]
- Wink, M; Schimmer, O. Modes of Action of Defensive Secondary Metabolites. In Functions of Plant Secondary Metabolites and Their Exploitation in Biotechnology; Wink, M, Ed.; Sheffield Academic Press: Sheffield, UK, 1999; pp. 17–133. [Google Scholar]
- Wink, M. Introduction: Biochemistry, Role and Biotechnology of Secondary Metabolites. In Functions of Plant Secondary Metabolites and Their Exploitation in Biotechnology; Wink, M, Ed.; Sheffield Academic Press: Sheffield, UK, 1999; pp. 1–16. [Google Scholar]
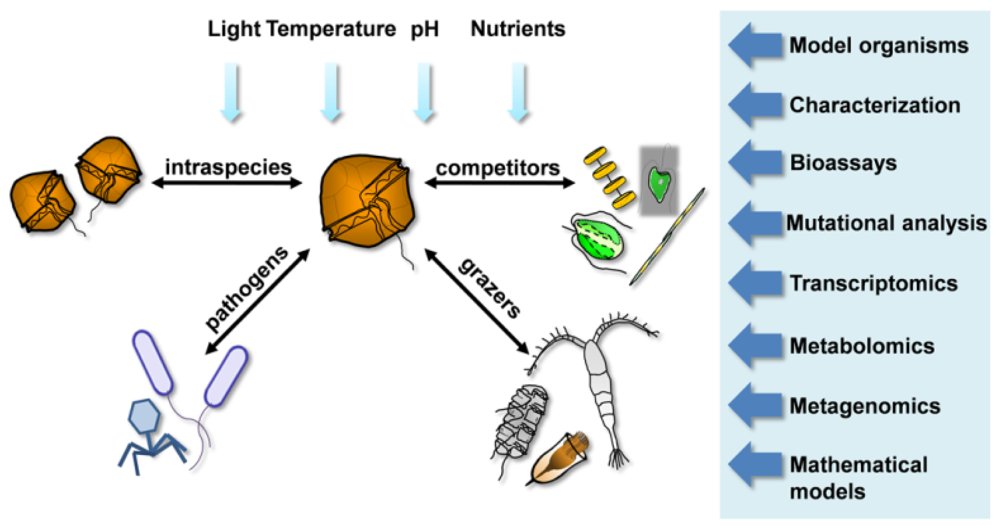

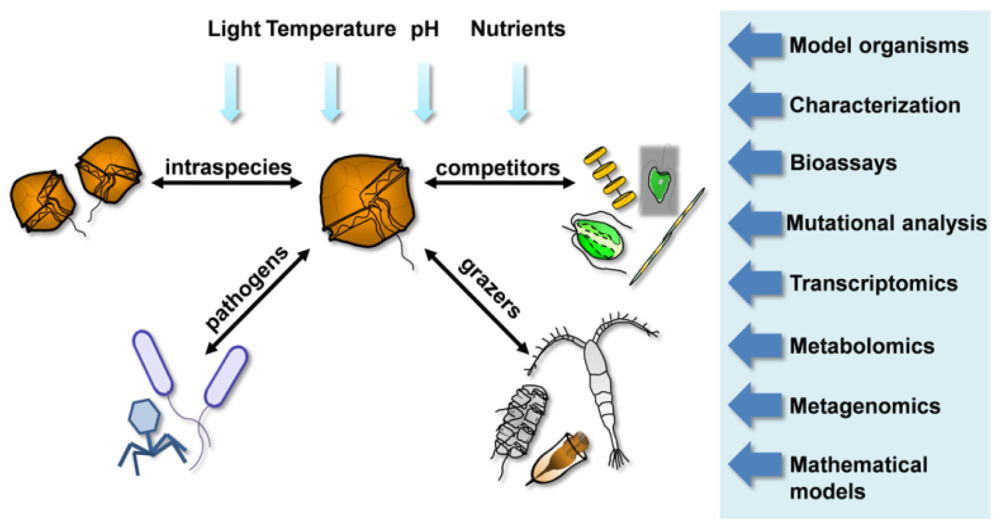{kind=link}
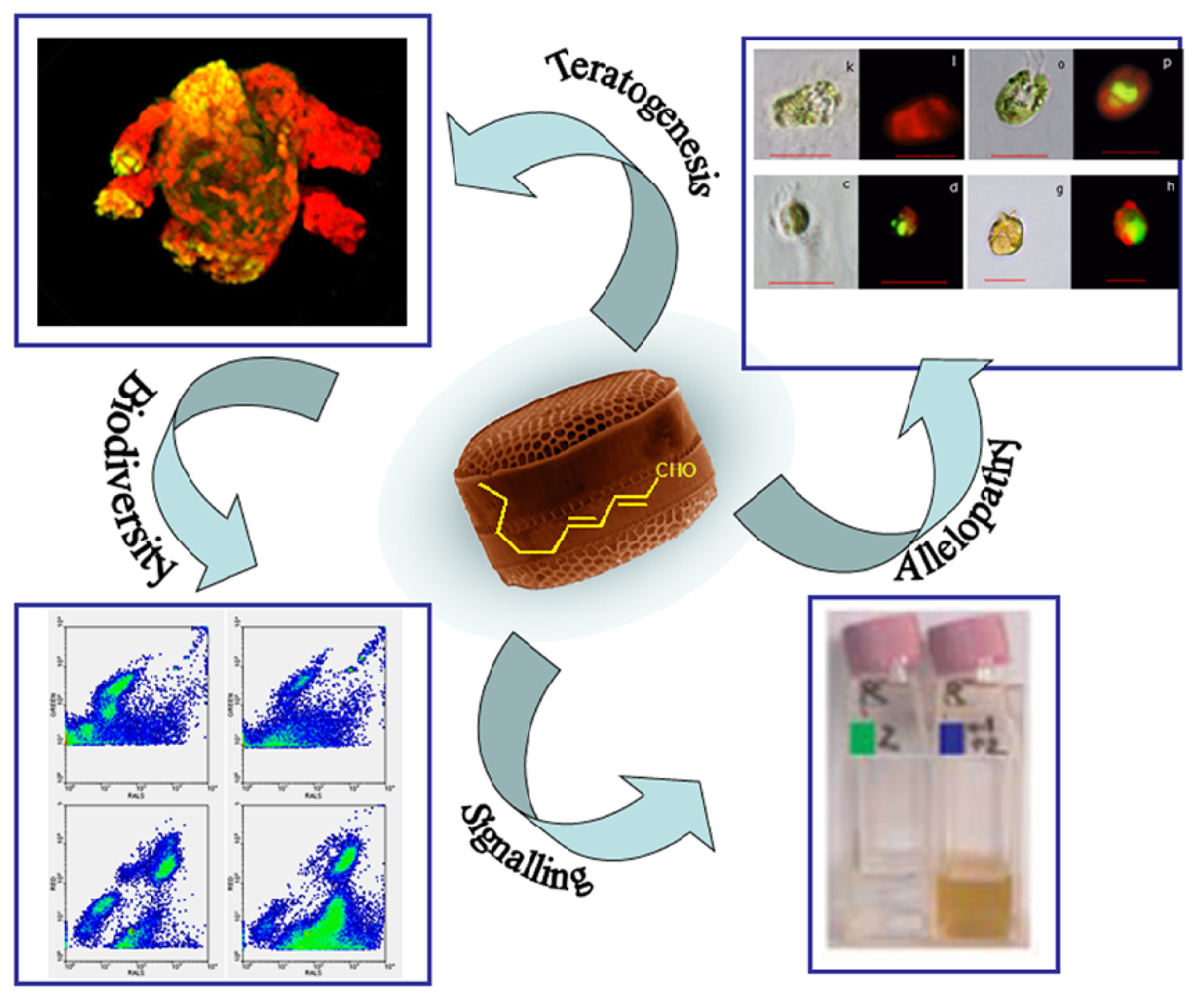
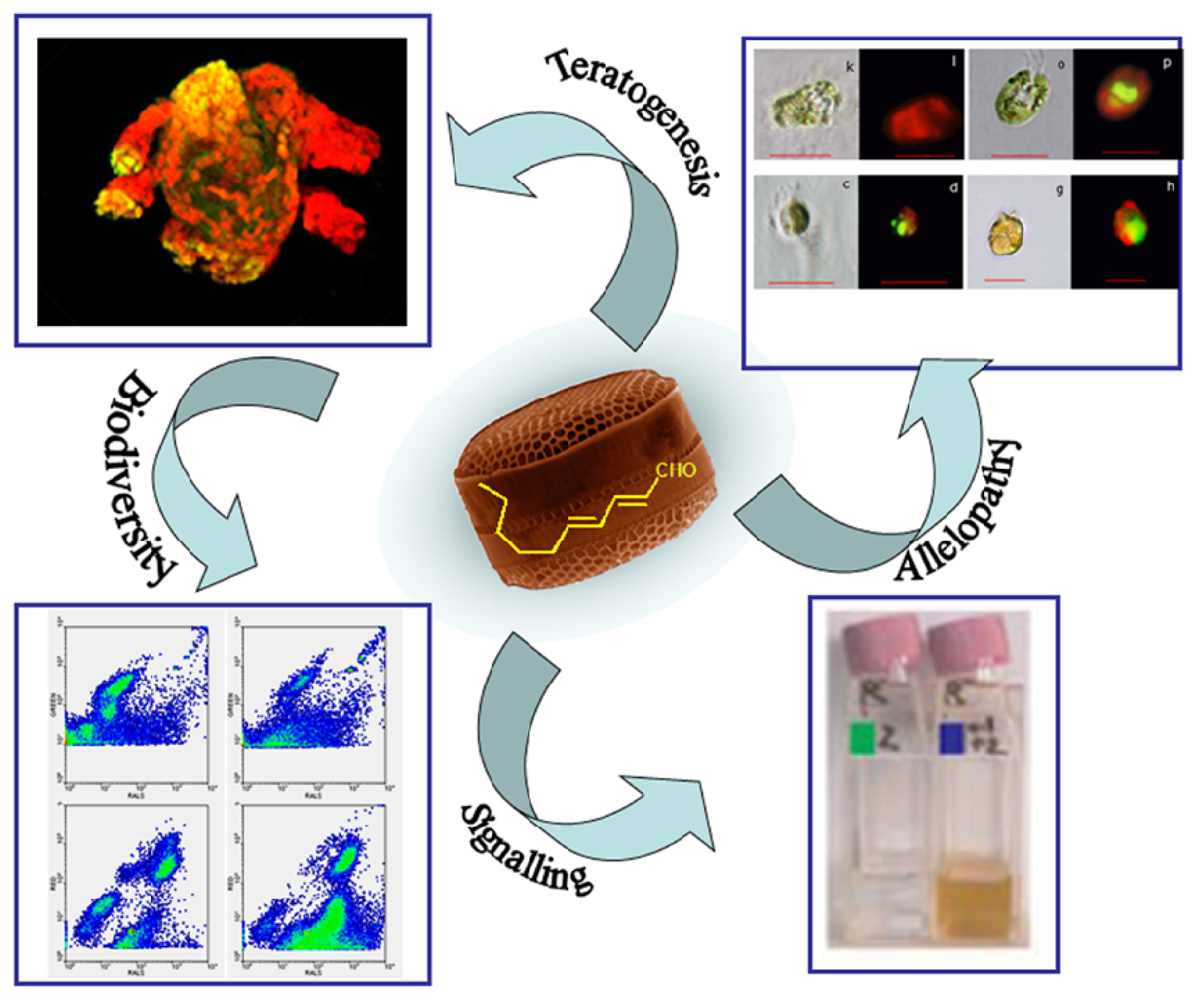{kind=link}
| Compounds | Producer | Target organism | Effect | Mode of action | References | |
|---|---|---|---|---|---|---|
| PUAs | Decatrienal | Thalassiosira rotula and other Thalassiosira species | Copepods Phytoplankton Bacteria | Reduced hatching Abnormal nauplii Growth inhibition | Anti-mitotic Apoptosis | [6,7,17,19] |
| Octatrienal Heptadienal | T. rotula Skeletonema marinoi | Copepods Phytoplankton | Reduced hatching Abnormal nauplii Growth inhibition | Anti-mitotic Apoptosis | [23,24] | |
| Hydroxy- and epoxy-Fatty acids | T. rotula S. marinoi Pseudo-nitzschia delicatissima Chaetoceros affinis C. socialis | Copepods | Reduced hatching Teratogenic effect Growth inhibition | Anti-mitotic Apoptosis | [8,25,26] | |
| Fatty acid hydroperoxydes | T. rotula S. marinoi P. delicatissima C. affinis C. socialis | Copepods | Reduced hatching Teratogenic effect Growth inhibition | Anti-mitotic Apoptosis | [8,25] | |
| Oxoacids | P. delicatissima | Copepods | Reduced hatching Teratogenic effect | Anti-mitotic Apoptosis | [9,27] | |
| DMSP: acrylate DMS | Emiliana huxleyi | Protistan Crustacean | Feeding deterrence Search behaviour | Unknown | [28] | |
| Apo-fucoxanthins | T. weissfloggi Phaeodactilum tricornutum | Copepods | Feeding deterrence | Unknown | [29,30] | |
| Saxitoxins | Alexandrium sp. | Copepods | Feeding deterrence | Unknown | [31] | |
| Karlotoxin | Karlodinium veneficum | Copepods Cryptophytes | Feeding deterrence Prey capture | Unknown | [32] | |
| Lytic compound (s) | Alexandrium tamarense | Rhodomonas baltica | Cell membrane lyis | Unknown | [33] | |
© 2011 by the authors; licensee MDPI, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Ianora, A.; Bentley, M.G.; Caldwell, G.S.; Casotti, R.; Cembella, A.D.; Engström-Öst, J.; Halsband, C.; Sonnenschein, E.; Legrand, C.; Llewellyn, C.A.; et al. The Relevance of Marine Chemical Ecology to Plankton and Ecosystem Function: An Emerging Field. Mar. Drugs 2011, 9, 1625-1648. https://doi.org/10.3390/md9091625
Ianora A, Bentley MG, Caldwell GS, Casotti R, Cembella AD, Engström-Öst J, Halsband C, Sonnenschein E, Legrand C, Llewellyn CA, et al. The Relevance of Marine Chemical Ecology to Plankton and Ecosystem Function: An Emerging Field. Marine Drugs. 2011; 9(9):1625-1648. https://doi.org/10.3390/md9091625
Chicago/Turabian StyleIanora, Adrianna, Matthew G. Bentley, Gary S. Caldwell, Raffaella Casotti, Allan D. Cembella, Jonna Engström-Öst, Claudia Halsband, Eva Sonnenschein, Catherine Legrand, Carole A. Llewellyn, and et al. 2011. "The Relevance of Marine Chemical Ecology to Plankton and Ecosystem Function: An Emerging Field" Marine Drugs 9, no. 9: 1625-1648. https://doi.org/10.3390/md9091625
APA StyleIanora, A., Bentley, M. G., Caldwell, G. S., Casotti, R., Cembella, A. D., Engström-Öst, J., Halsband, C., Sonnenschein, E., Legrand, C., Llewellyn, C. A., Paldavičienë, A., Pilkaityte, R., Pohnert, G., Razinkovas, A., Romano, G., Tillmann, U., & Vaiciute, D. (2011). The Relevance of Marine Chemical Ecology to Plankton and Ecosystem Function: An Emerging Field. Marine Drugs, 9(9), 1625-1648. https://doi.org/10.3390/md9091625