Teratogenic Effects of Diatom Metabolites on Sea Urchin Paracentrotus lividus Embryos




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
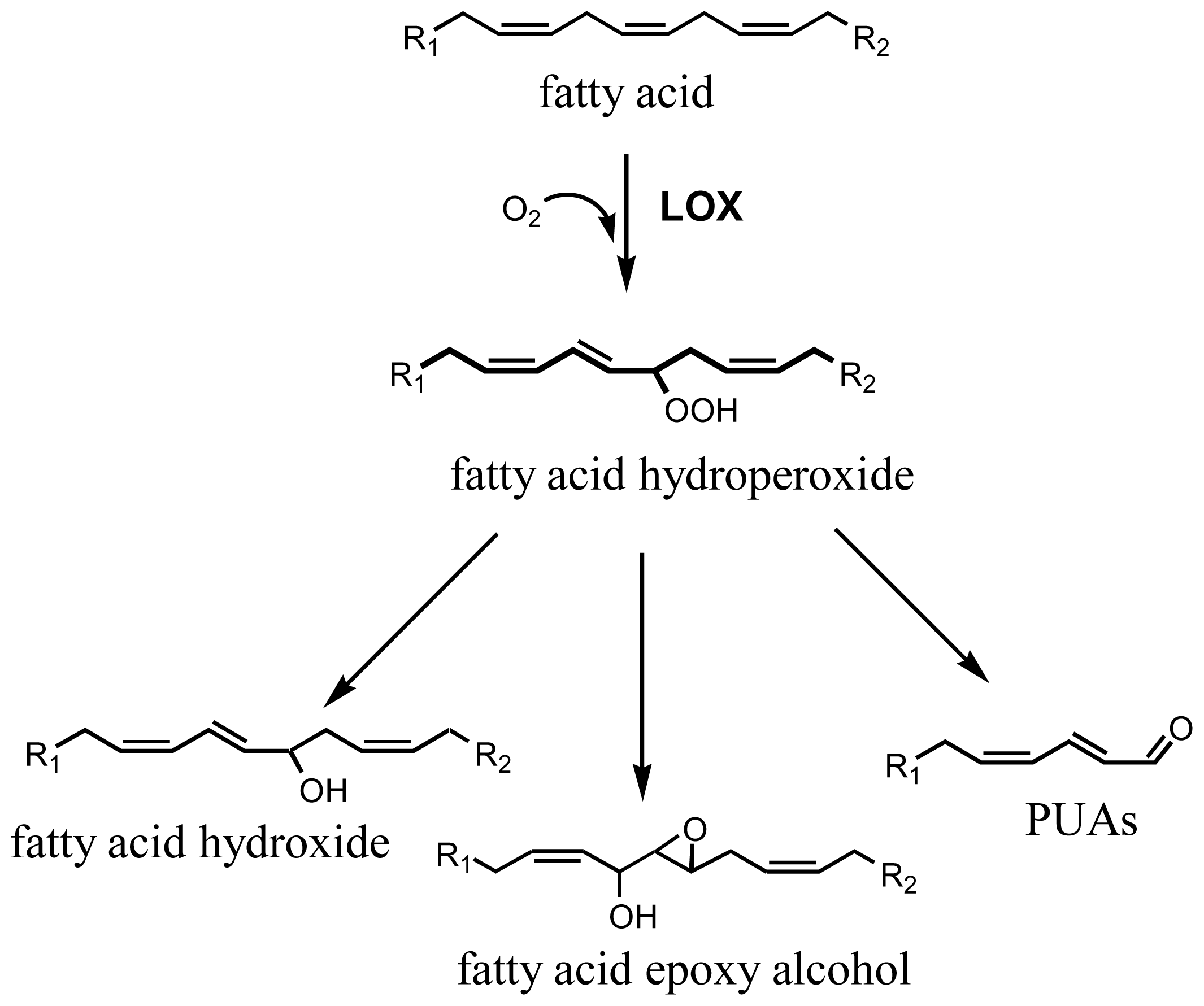
:1. Introduction
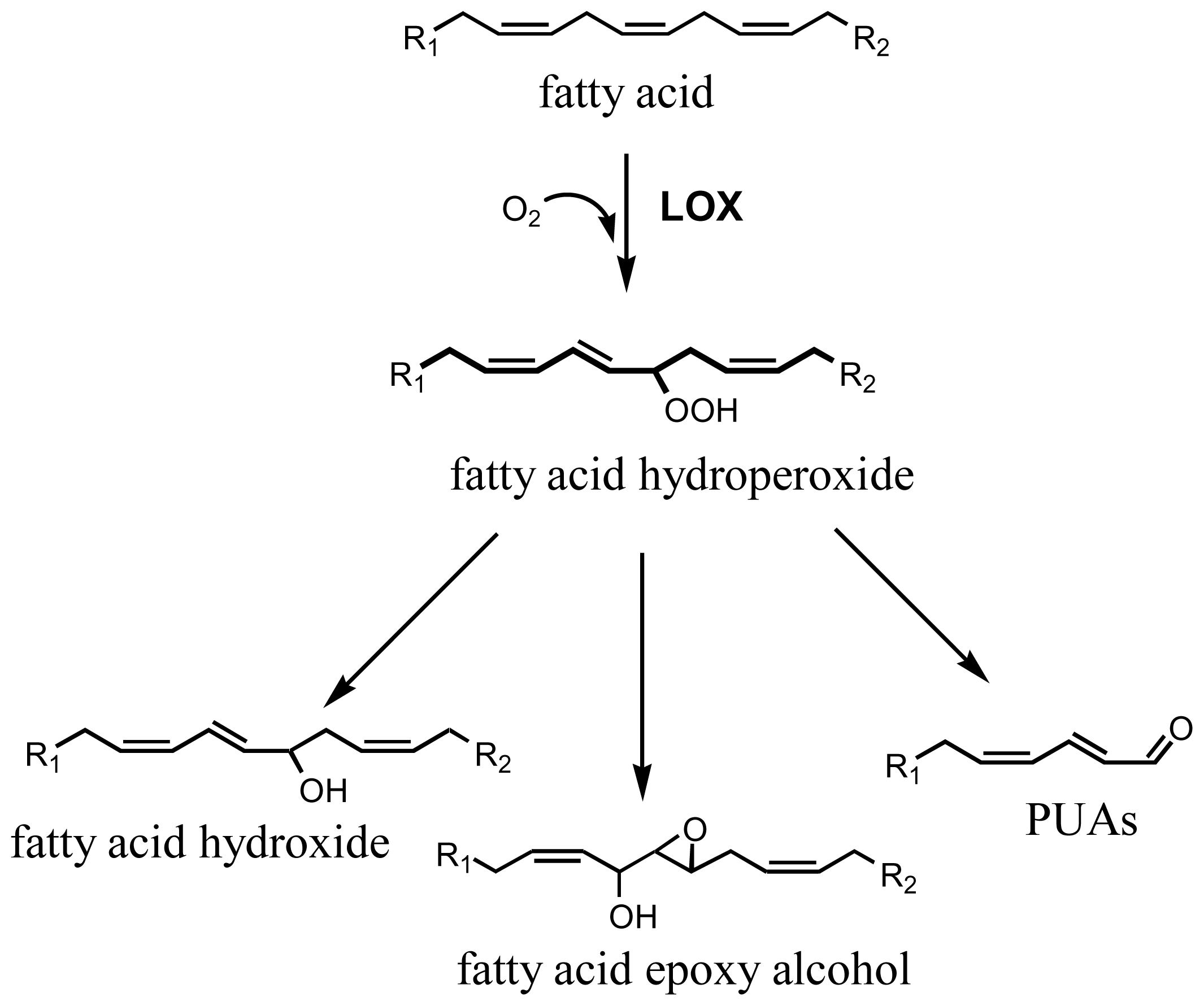
2. Results and Discussion
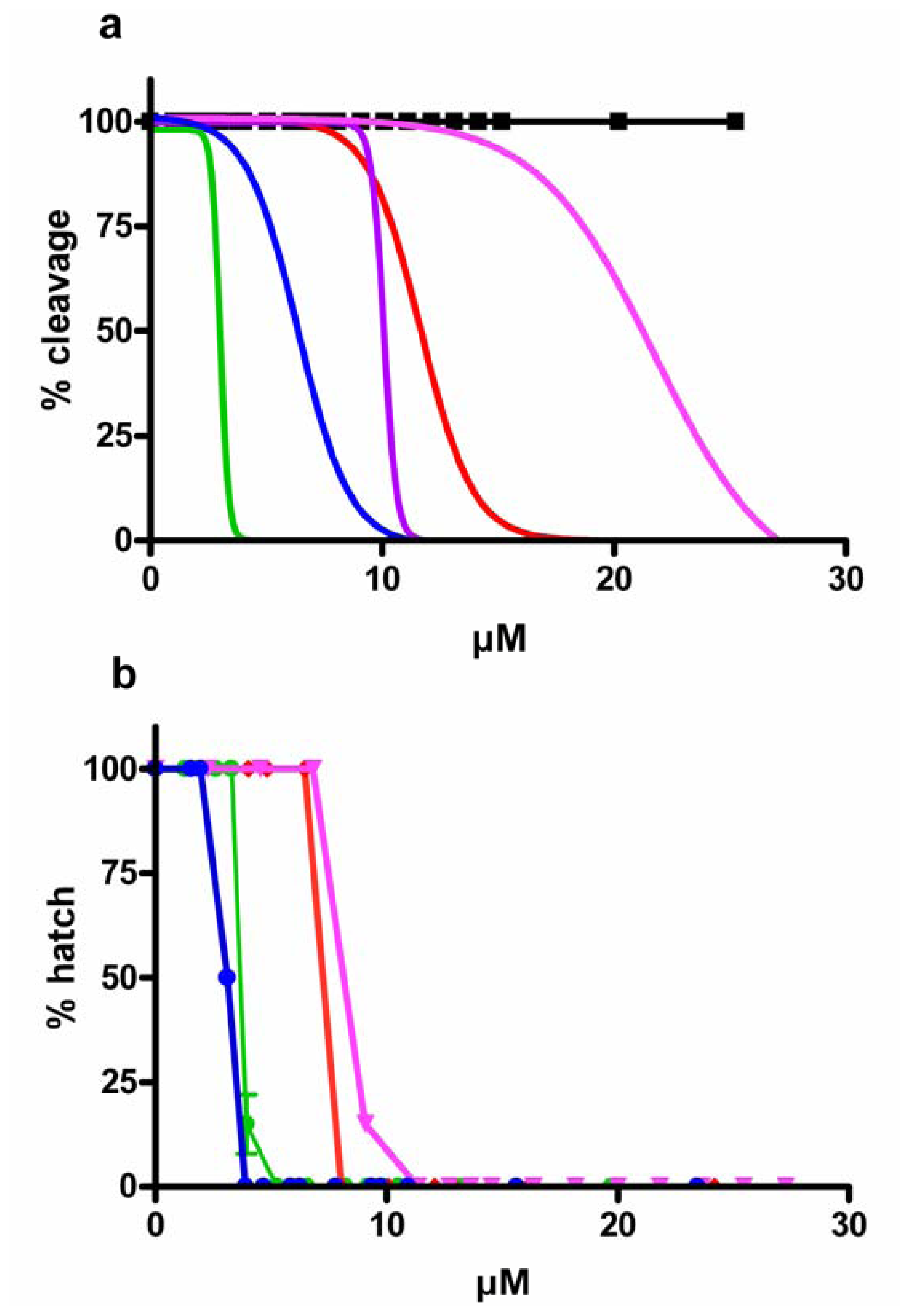
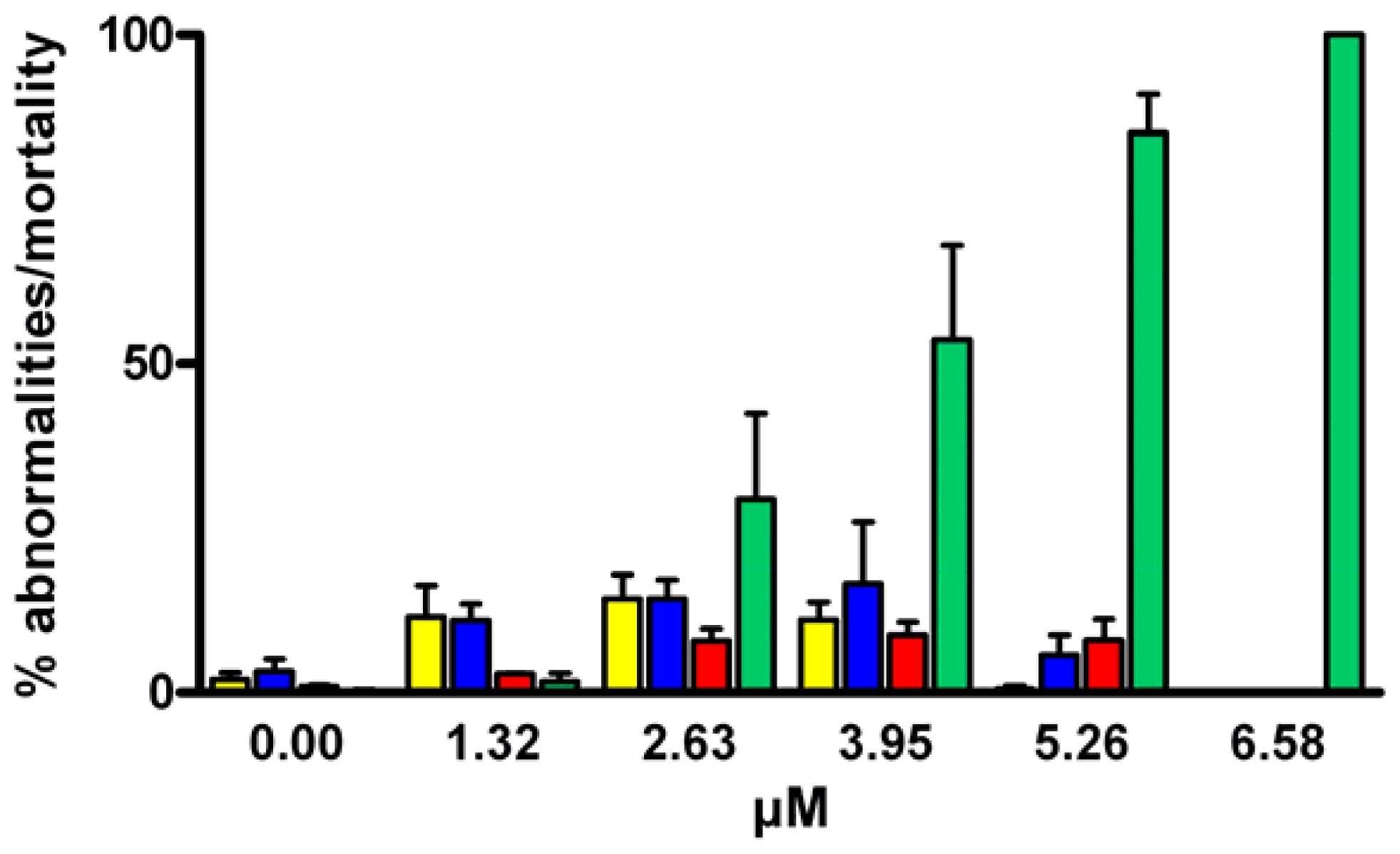
2.1. Effect of PUAs on Sea Urchin Cleavage and Hatching
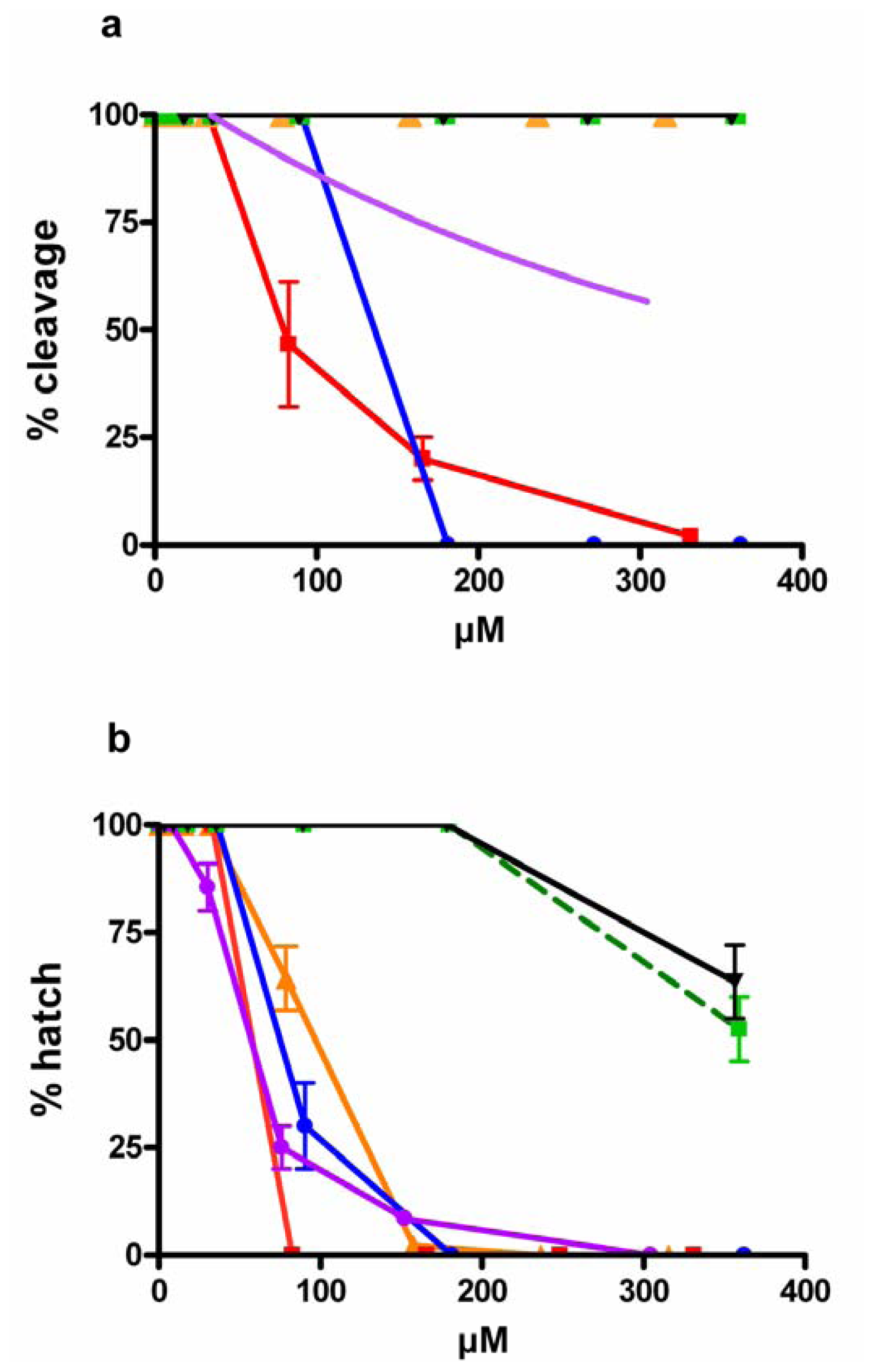
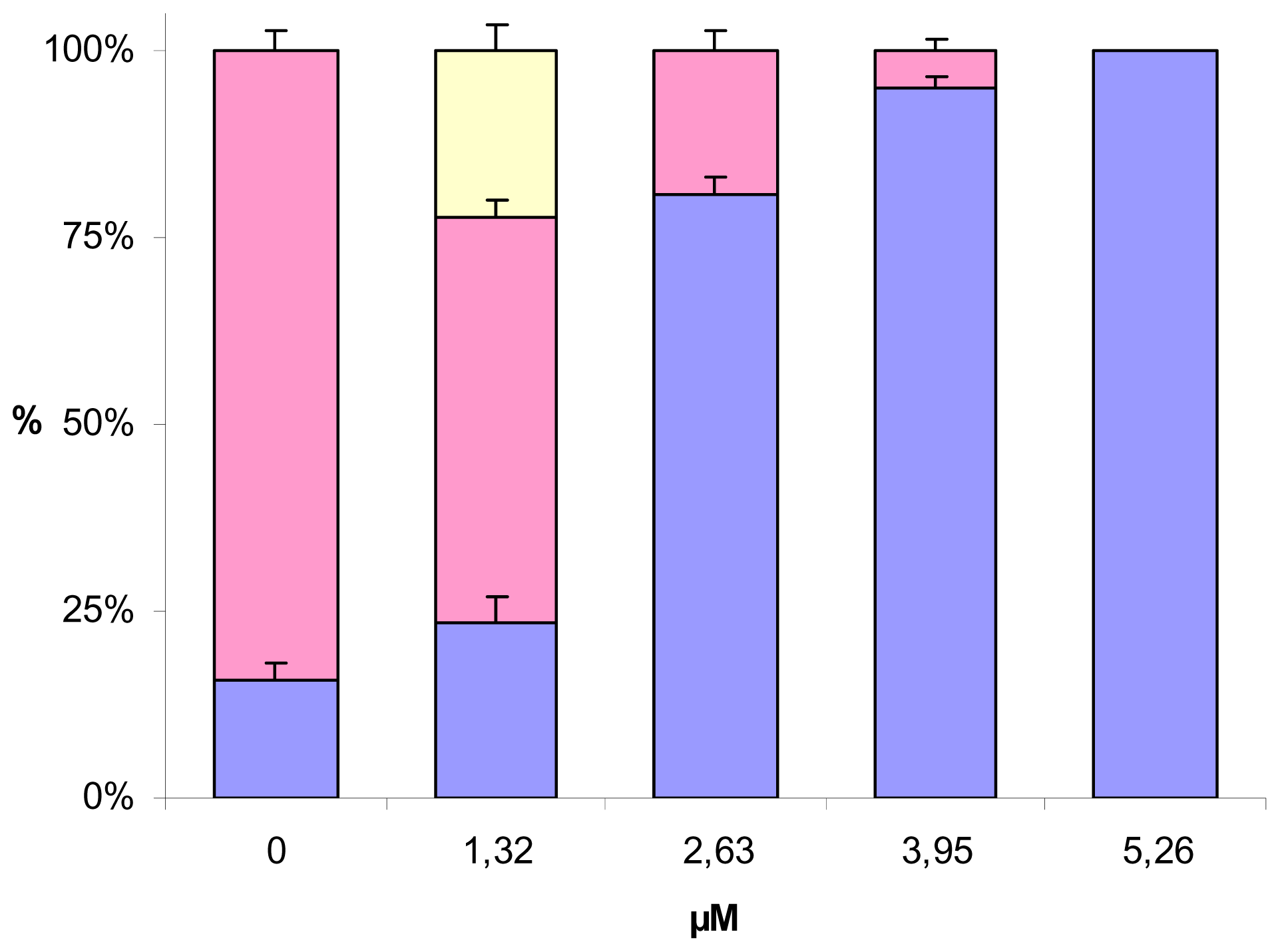
2.2. Effect of PUFAs on Embryo Cleavage and Hatching
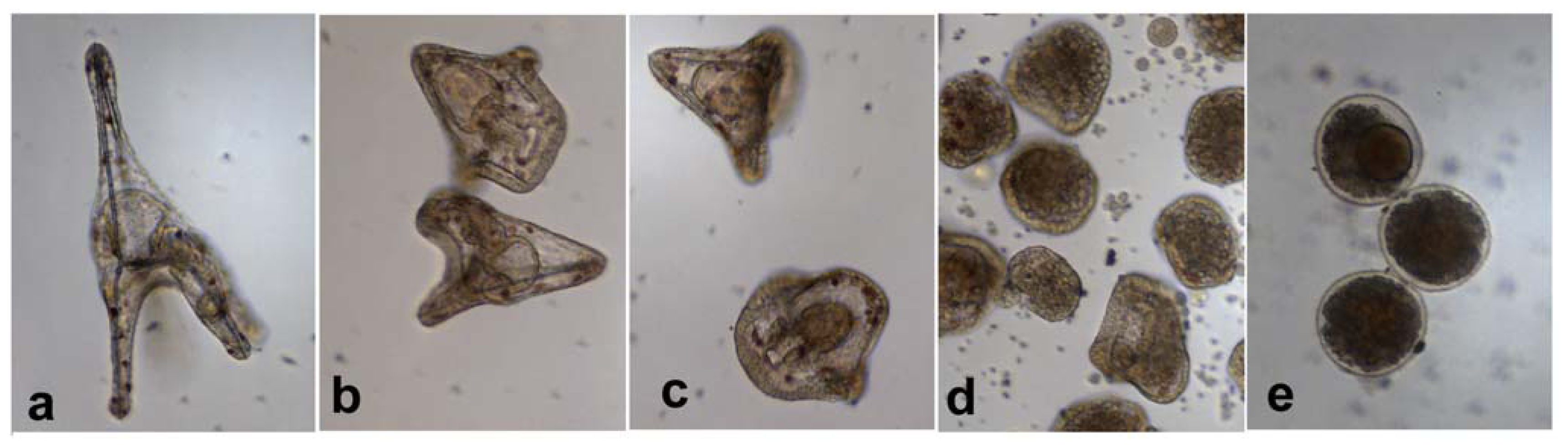
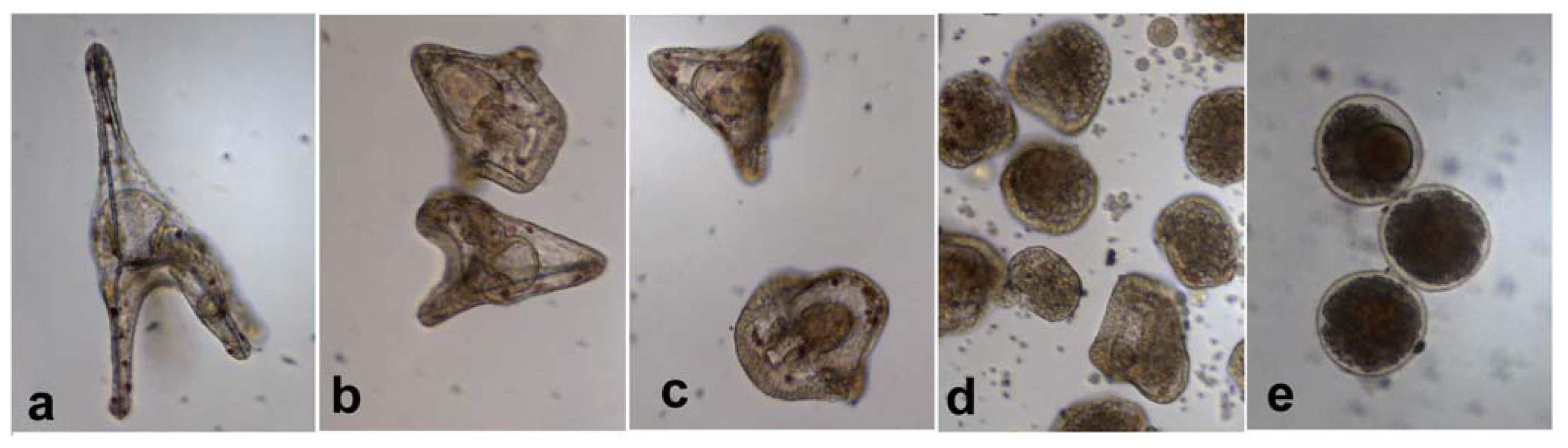
2.3. Effect of PUAs on Paracentrotus lividus Larval Development
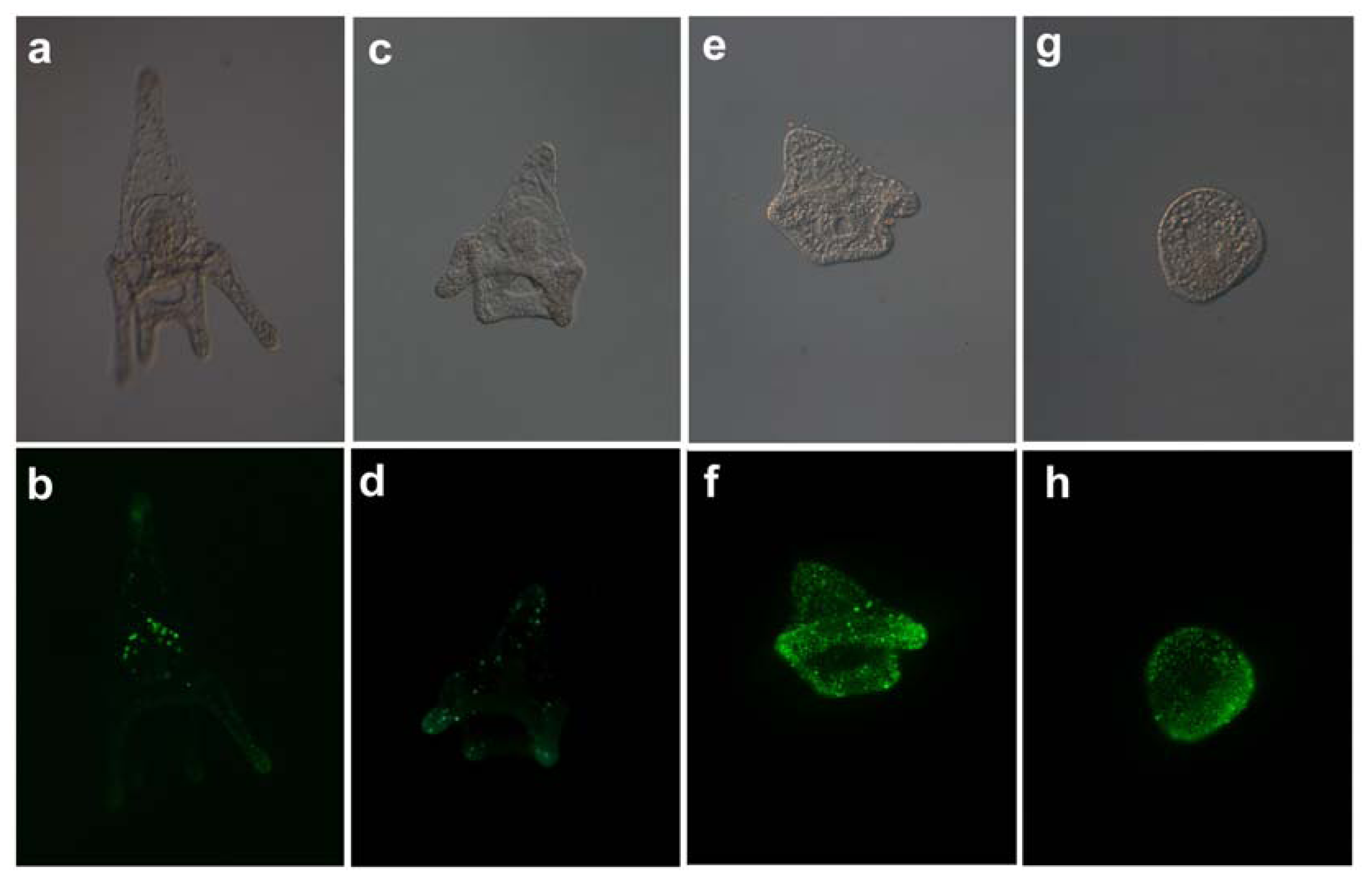
2.4. TUNEL Assay
3. Experimental Section
3.1. Aldehydes and Fatty Acids Stock Solutions
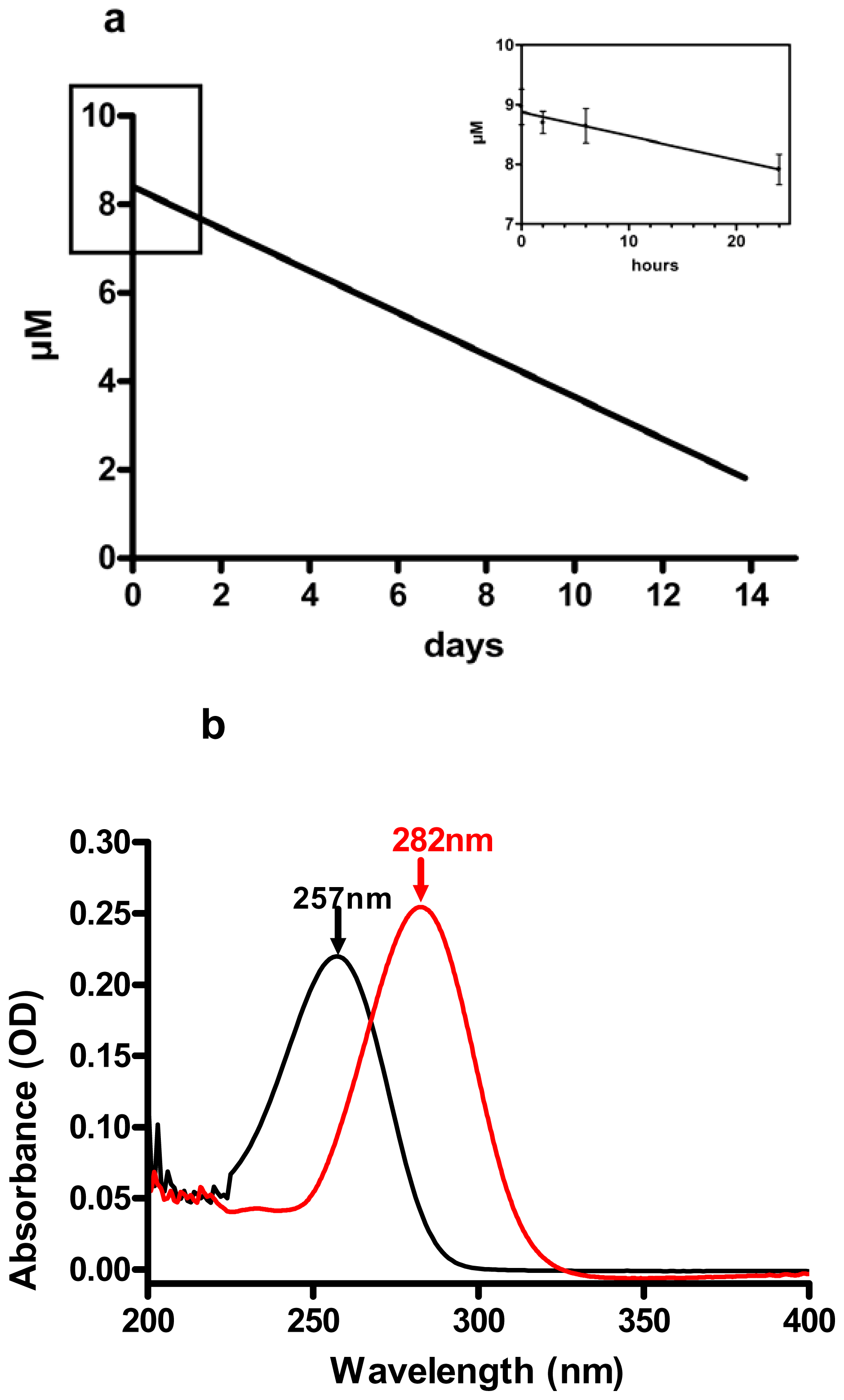
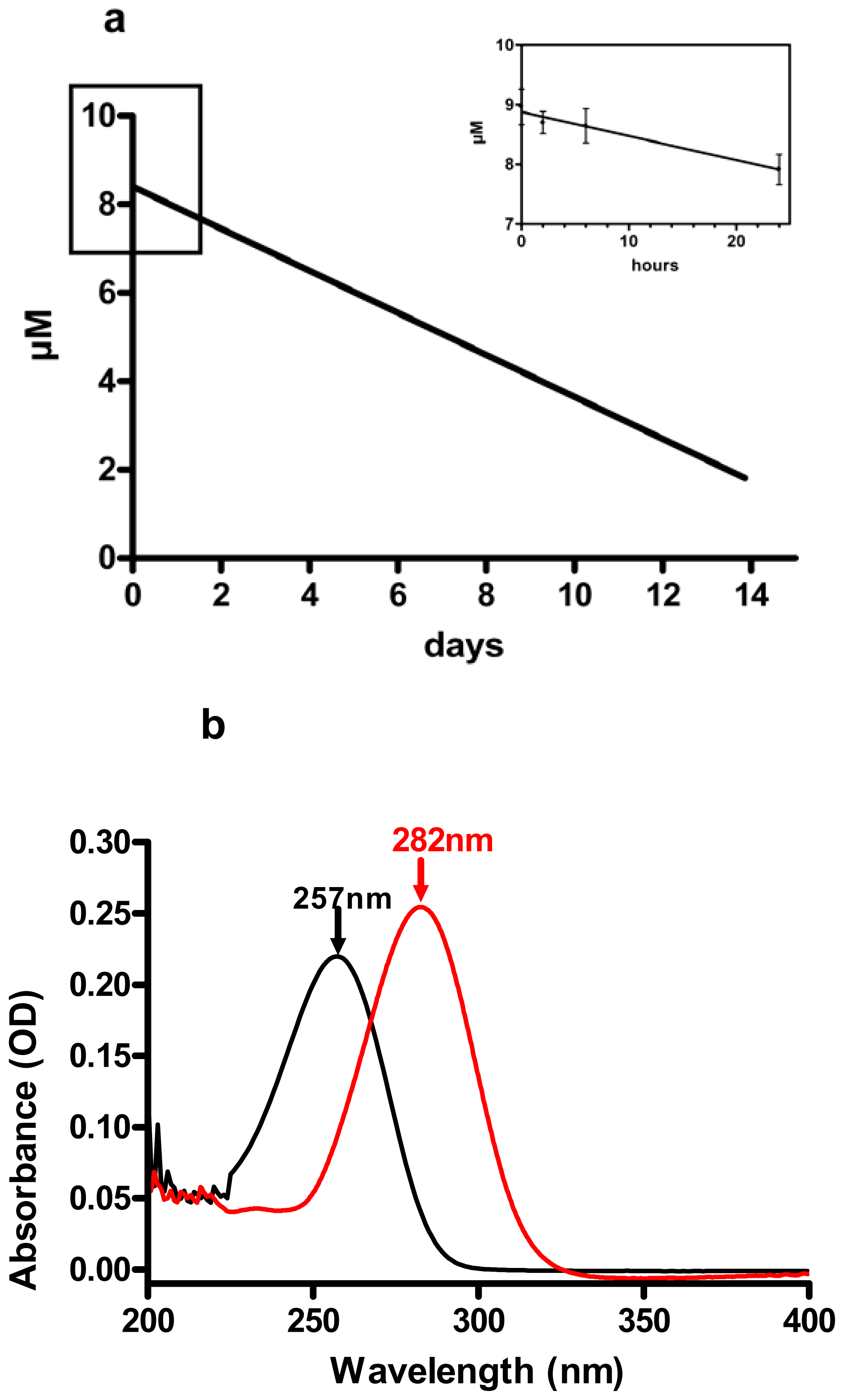
3.2. Decadienal Stability in Nutrient Enriched Sea Water
3.3. Sea Urchin Gamete Collection
3.4. Embryotoxicity Test
3.5. Tunel Fluorescence Labeling (TUNEL)
4. Conclusions
Acknowledgements
- Samples Availability: Available from the authors.
References and Notes
- Falkowski, PG. The role of phytoplankton photosynthesis in global biogeochemical cycles. Photosynth Res 1994, 39, 235–258. [Google Scholar]
- Nelson, DM; Treguer, P; Brzezinski, MA; Leynaert, A; Queguiner, B. Production and dissolution of biogenic silica in the ocean-revised global estimates, comparison with regional data and relationship to biogenic sedimentation. Global Biogeochem Cycles 1995, 9, 359–372. [Google Scholar]
- Miralto, A; Barone, G; Romano, G; Poulet, SA; Ianora, A; Russo, GL; Buttino, I; Mazzarella, G; Laabir, M; Cabrini, M; Giacobbe, MG. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar]
- d’Ippolito, G; Tucci, S; Cutignano, A; Romano, G; Cimino, G; Miralto, A; Fontana, A. The role of complex lipids in the synthesis of bioactive aldehydes of the marine diatom Skeletonema costatum. Biochim Biophys Acta Mol Cell Biol Lipids 2004, 1686, 100–107. [Google Scholar]
- Fontana, A; d’Ippolito, G; Cutignano, A; Miralto, A; Ianora, A; Romano, G; Cimino, G. Chemistry of oxylipin pathways in marine diatoms. Pure Appl Chem 2007, 79, 481–490. [Google Scholar]
- Fontana, A; d’Ippolito, G; Cutignano, A; Romano, G; Lamari, N; Gallucci, AM; Cimino, G; Miralto, A; Ianora, A. LOX-induced lipid peroxidation mechanism responsible for the detrimental effect of marine diatoms on zooplankton grazers. Chembiochem 2007, 8, 1810–1818. [Google Scholar]
- d’Ippolito, G; Romano, G; Caruso, T; Spinella, A; Cimino, G; Fontana, A. Production of octadienal in the marine diatom Skeletonema costatum. Org Lett 2003, 5, 885–887. [Google Scholar]
- Ianora, A; Miralto, A. Toxigenic effects of diatoms on grazers, phytoplankton and other microbes: a review. Ecotoxicology 2009. [Google Scholar] [CrossRef]
- Caldwell, GS. The influence of bioactive oxylipins from marine diatoms on invertebrate reproduction and development. Mar Drugs 2009, 7, 367–400. [Google Scholar]
- Caldwell, GS; Bentley, MG; Olive, PJW. First evidence of sperm motility inhibition by the diatom aldehyde 2E,4E-decadienal. Mar Ecol Progr Ser 2004, 273, 97–108. [Google Scholar]
- Caldwell, GS; Olive, PJW; Bentley, MG. Inhibition of embryonic development and fertilization in broadcast spawning marine invertebrates by water soluble diatom extracts and the diatom toxin 2-trans,4-trans decadienal. Aquat Toxicol 2002, 60, 123–137. [Google Scholar]
- Hansen, E; Even, Y; Geneviere, AM. The alpha,beta,gamma,delta-unsaturated aldehyde 2-trans-4-trans-decadienal disturbs DNA replication and mitotic events in early sea urchin embryos. Toxicol Sci 2004, 81, 190–197. [Google Scholar]
- Adolph, S; Poulet, SA; Pohnert, G. Synthesis and biological activity of alpha,beta,gamma,delta- unsaturated aldehydes from diatoms. Tetrahedron 2003, 59, 3003–3008. [Google Scholar]
- Pohnert, G; Lumineau, O; Cueff, A; Adolph, S; Cordevant, C; Lange, M; Poulet, S. Are volatile unsaturated aldehydes from diatoms the main line of chemical defence against copepods. Mar Ecol Progr Ser 2002, 245, 33–45. [Google Scholar]
- Romano, G; Russo, GL; Buttino, I; Ianora, A; Miralto, A. A marine diatom-derived aldehyde induces apoptosis in copepod and sea urchin embryos. J Exp Biol 2003, 206, 3487–3494. [Google Scholar]
- d’Ippolito, G; Iadicicco, I; Romano, G; Fontana, A. Detection of short-chain aldehydes in marine organisms: the diatom Thalassiosira rotula. Tetrahedron Lett 2002, 43, 6137–6140. [Google Scholar]
- d’Ippolito, G; Romano, G; Iadicicco, O; Miralto, A; Ianora, A; Cimino, G; Fontana, A. New birth-control aldehydes from the marine diatom Skeletonema costatum: characterization and biogenesis. Tetrahedron Lett 2002, 43, 6133–6136. [Google Scholar]
- Sellem, F; Pesando, D; Bodennec, G; El Abed, A; Girard, JP. Toxic effects of Gymnodinium cf. mikimotoi unsaturated fatty acids to gametes and embryos of the sea urchin Paracentrotus lividus. Water Res 2000, 34, 550–556. [Google Scholar]
- Pesando, D; Graillet, C; Braekman, JC; Dubreuil, A; Girard, JP; Puiseuxdao, S. The use of sea-urchin eggs as a model to investigate the effects of crassolide, a diterpene isolated from a soft coral. Toxicol in Vitro 1991, 5, 395–401. [Google Scholar]
- Gamberucci, A; Fulceri, R; Bygrave, FL; Benedetti, A. Unsaturated fatty acids mobilize intracellular calcium independent of IP3 generation and VIA insertion at the plasma membrane. Biochem Biophys Res Commun 1997, 241, 312–316. [Google Scholar]
- Guillard, RRL. Chanley, MH, Smith, WL, Eds.; Culture of phytoplankton for feeding marine invertebrates. In Culture of Marine Invertebrate Animals; Plenum Press: New York, NY, USA, 1975; pp. 26–60. [Google Scholar]
- Pohnert, G. Phospholipase A(2) activity triggers the wound-activated chemical defense in the diatom Thalassiosira rotula. Plant Physiol 2002, 129, 103–111. [Google Scholar]
- Starr, M; Himmelman, JH; Therriault, JC. Direct coupling of marine invertebrate spawning with phytoplankton blooms. Science 1990, 247, 1071–1074. [Google Scholar]
- Vidoudez, C; Pohnert, G. Growth phase-specific release of polyunsaturated aldehydes by the diatom Skeletonema marinoi. J Plankton Res 2008, 30, 1305–1313. [Google Scholar]
- Ribalet, F; Berges, JA; Ianora, A; Casotti, R. Growth inhibition of cultured marine. phytoplankton by toxic algal-derived polyunsaturated aldehydes. Aquat Toxicol 2007, 85, 219–227. [Google Scholar]
- Tosti, E; Romano, G; Buttino, I; Cuomo, A; Ianora, A; Miralto, A. Bioactive aldehydes from diatoms block the fertilization current in asciadian oocytes. Mol Reprod Dev 2003, 66, 72–80. [Google Scholar]
- Caldwell, GS; Lewis, C; Olive, PJW; Bentley, MG. Exposure to 2,4-decadienal negatively impacts upon marine invertebrate larval fitness. Mar Environ Res 2005, 59, 405–417. [Google Scholar]
- Ianora, A; Romano, G; Carotenuto, Y; Esposito, F; Roncalli, V; Buttino, I; Miralto, A. Impact of the diatom oxylipin 15S-HEPE on the reproductive success of the copepod Temora stylifera. Hydrobiologia 2010. submitted. [Google Scholar]
- Zakeri, Z; Lockshin, RA. Cell death during development. J Immunol Methods 2002, 265, 3–20. [Google Scholar]
- Agnello, M; Roccheri, M. Apoptosis: focus on sea urchin development. Apoptosis 2009, 15, 322–330. [Google Scholar]
- Juttner, F. Evidence that polyunsaturated aldehydes of diatoms are repellents for pelagic crustacean grazers. Aquat Ecol 2005, 39, 271–282. [Google Scholar]
- Evans, WG. Oscillatoria sp. (Cyanophyta) mat metabolites implicated in habitat selection in Bembidion obtusidens (Coleoptera: Carabidae). J Chem Ecol 1982, 8, 671–678. [Google Scholar]
- Hockelmann, C; Moens, T; Juttner, F. Odor compounds from cyanobacterial biofilms acting as attractants and repellents for free-living nematodes. Limnol Oceanogr 2004, 49, 1809–1819. [Google Scholar]
- Juttner, F; Durst, U. High lipoxygenase activities in epilithic biofilms of diatoms. Archiv Fur Hydrobiologie 1997, 138, 451–463. [Google Scholar]
- Pippen, E; Nonaka, M. Notes-A convenient method for synthesizing normal aliphatic 2,4-dienals. J Org Chem 1958, 23, 1580–1582. [Google Scholar]
- Pawson, B; Cheung, H; Gurbaxani, S; Saucy, G. Synthesis of natural(+)-juvabione, its enantiomer(−)-juvabione, and their diastereoisomers (+)- and (−)-epijuvabione. J Am Chem Soc 1970, 92, 336–343. [Google Scholar]
- Pagano, G; Cipollaro, M; Corsale, G; Esposito, A; Ragucci, E; Giordano, G; Trieff, N. The Sea Urchin: Bioassay for the Assessment of Damage from Environmental Contaminants; American Society for Testing and Materials: Philadelphia, PA, USA, 1986; pp. 67–92. [Google Scholar]
 ), aldehyde mix (see text) (
), aldehyde mix (see text) (  ), 2-trans,4-trans-octadienal (
), 2-trans,4-trans-octadienal (  ), 2-trans,4-trans,7-octatrienal (
), 2-trans,4-trans,7-octatrienal (  ), 2-trans,4-trans-heptadienal (
), 2-trans,4-trans-heptadienal (  ) and tridecanal (▪). Values (means ± S.D.; N = 600) are the results of three different experiments.
), aldehyde mix (see text) ( ), 2-trans,4-trans-octadienal ( ), 2-trans,4-trans,7-octatrienal ( ), 2-trans,4-trans-heptadienal ( ) and tridecanal (▪). Values (means ± S.D.; N = 600) are the results of three different experiments.
) and tridecanal (▪). Values (means ± S.D.; N = 600) are the results of three different experiments.
), aldehyde mix (see text) ( ), 2-trans,4-trans-octadienal ( ), 2-trans,4-trans,7-octatrienal ( ), 2-trans,4-trans-heptadienal ( ) and tridecanal (▪). Values (means ± S.D.; N = 600) are the results of three different experiments.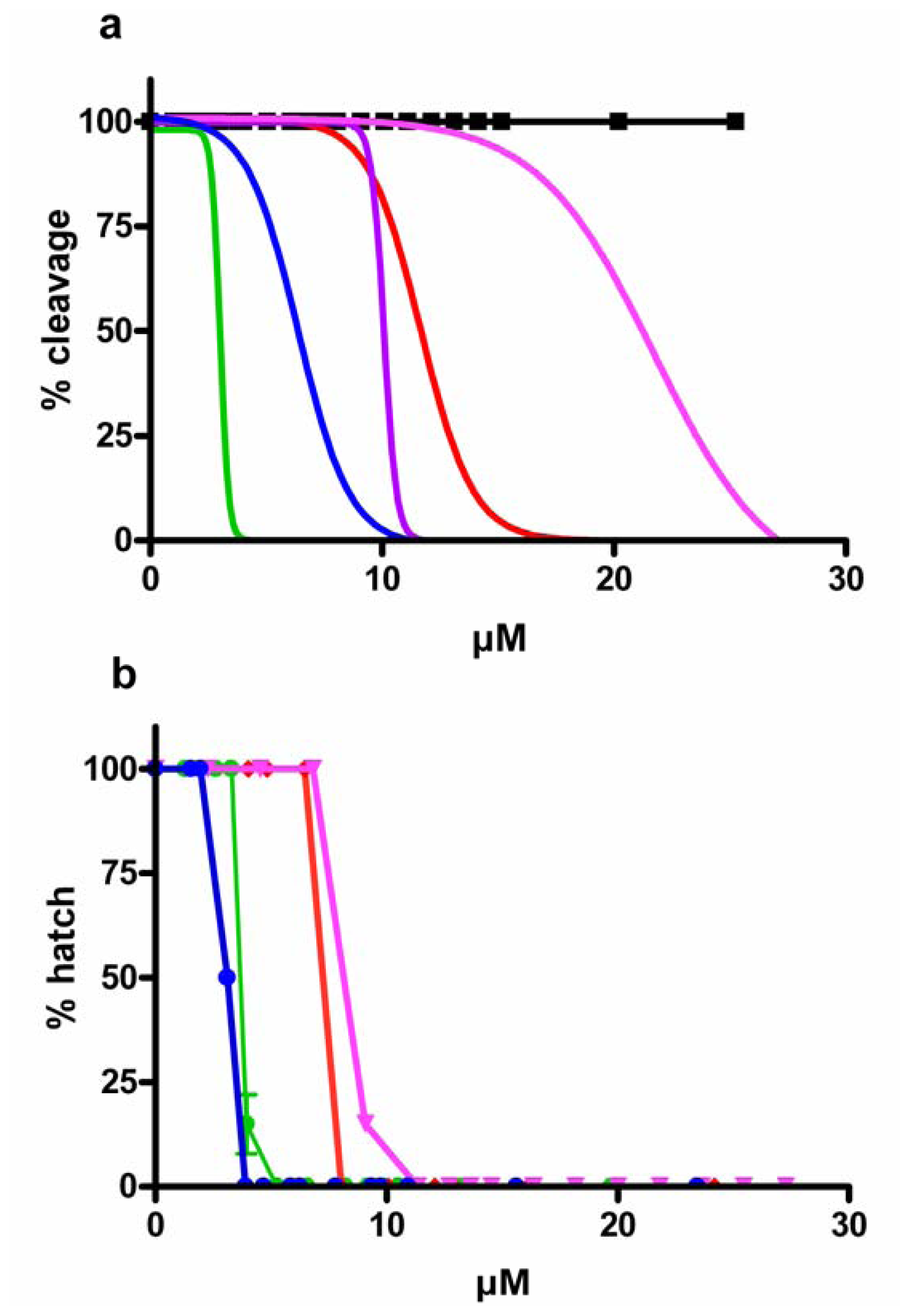
 ), EPA-ME (
), EPA-ME (  ), DHA (
), DHA (  ), stearidonic ( ), linoleic (▾) and γ-linolenic (
), stearidonic ( ), linoleic (▾) and γ-linolenic (  ) acids. Values (means ± S.D.; N = 600) are the results of three different experiments.
), EPA-ME ( ), DHA ( ), stearidonic ( ), linoleic (▾) and γ-linolenic ( ) acids. Values (means ± S.D.; N = 600) are the results of three different experiments.
) acids. Values (means ± S.D.; N = 600) are the results of three different experiments.
), EPA-ME ( ), DHA ( ), stearidonic ( ), linoleic (▾) and γ-linolenic ( ) acids. Values (means ± S.D.; N = 600) are the results of three different experiments.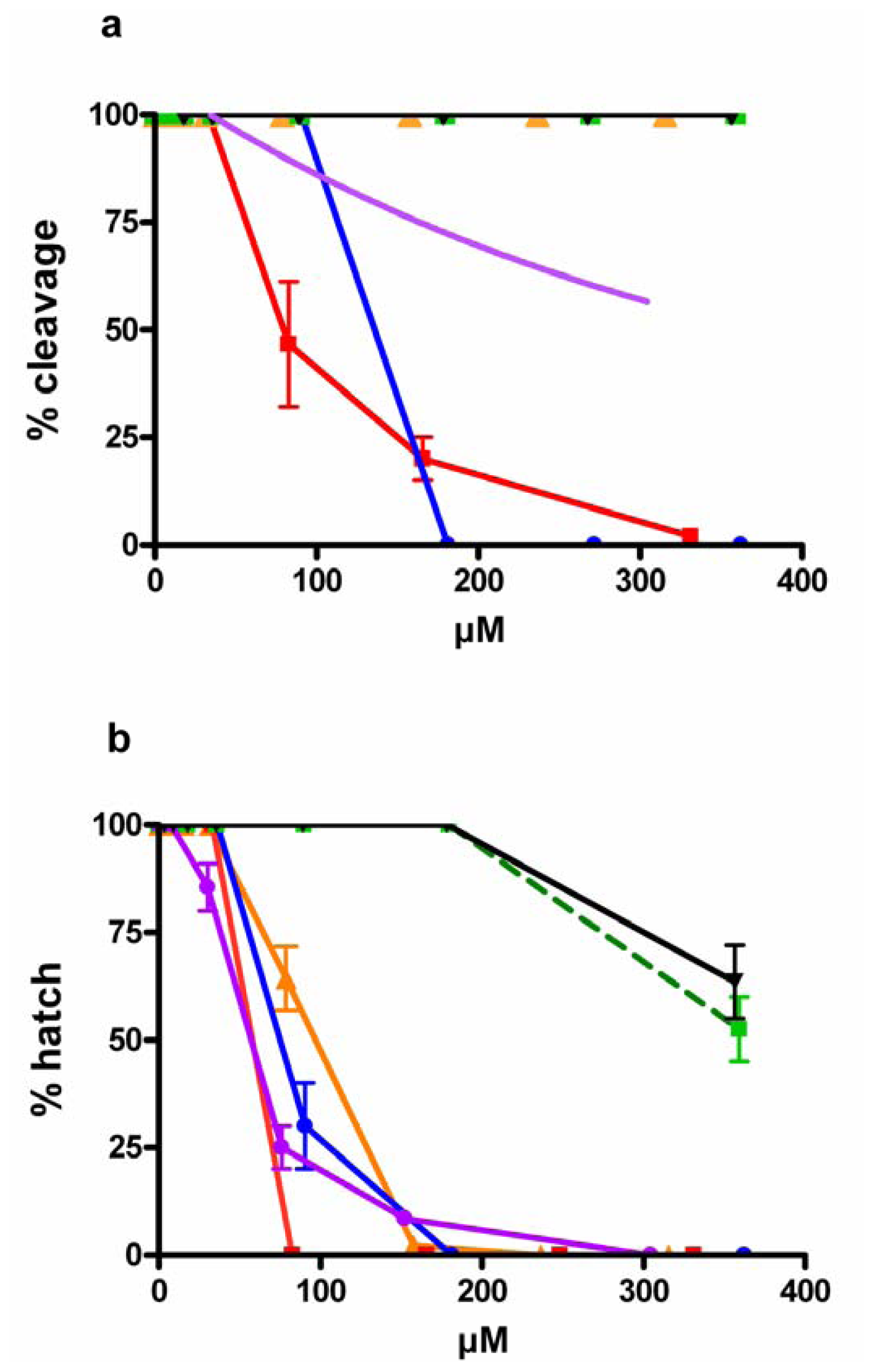


© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Romano, G.; Miralto, A.; Ianora, A. Teratogenic Effects of Diatom Metabolites on Sea Urchin Paracentrotus lividus Embryos. Mar. Drugs 2010, 8, 950-967. https://doi.org/10.3390/md8040950
Romano G, Miralto A, Ianora A. Teratogenic Effects of Diatom Metabolites on Sea Urchin Paracentrotus lividus Embryos. Marine Drugs. 2010; 8(4):950-967. https://doi.org/10.3390/md8040950
Chicago/Turabian StyleRomano, Giovanna, Antonio Miralto, and Adrianna Ianora. 2010. "Teratogenic Effects of Diatom Metabolites on Sea Urchin Paracentrotus lividus Embryos" Marine Drugs 8, no. 4: 950-967. https://doi.org/10.3390/md8040950