Effect of Elatol, Isolated from Red Seaweed Laurencia dendroidea, on Leishmania amazonensis

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
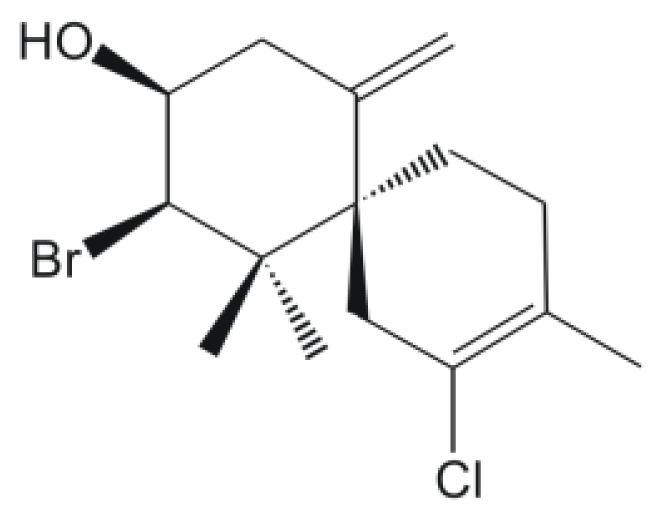
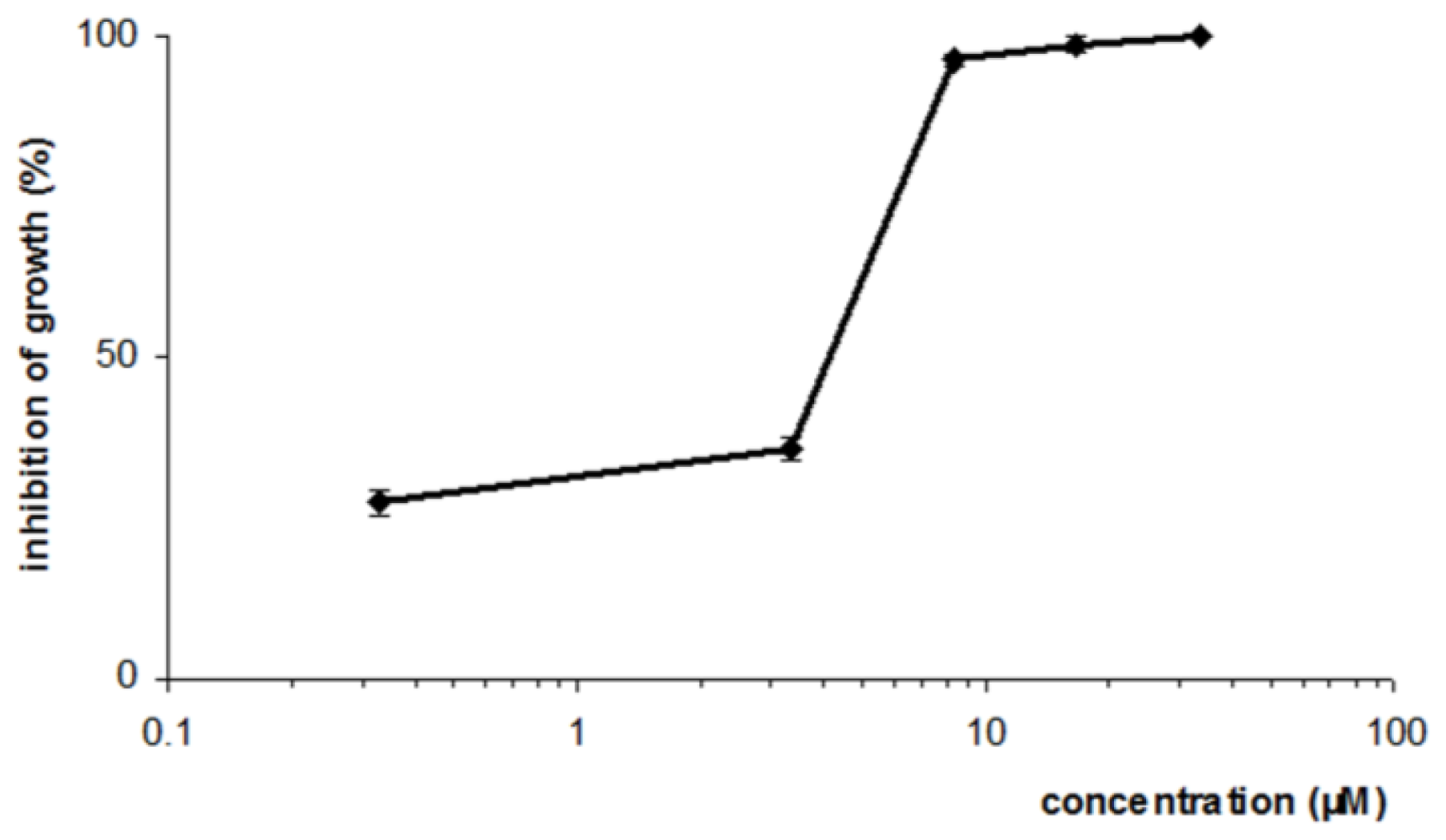
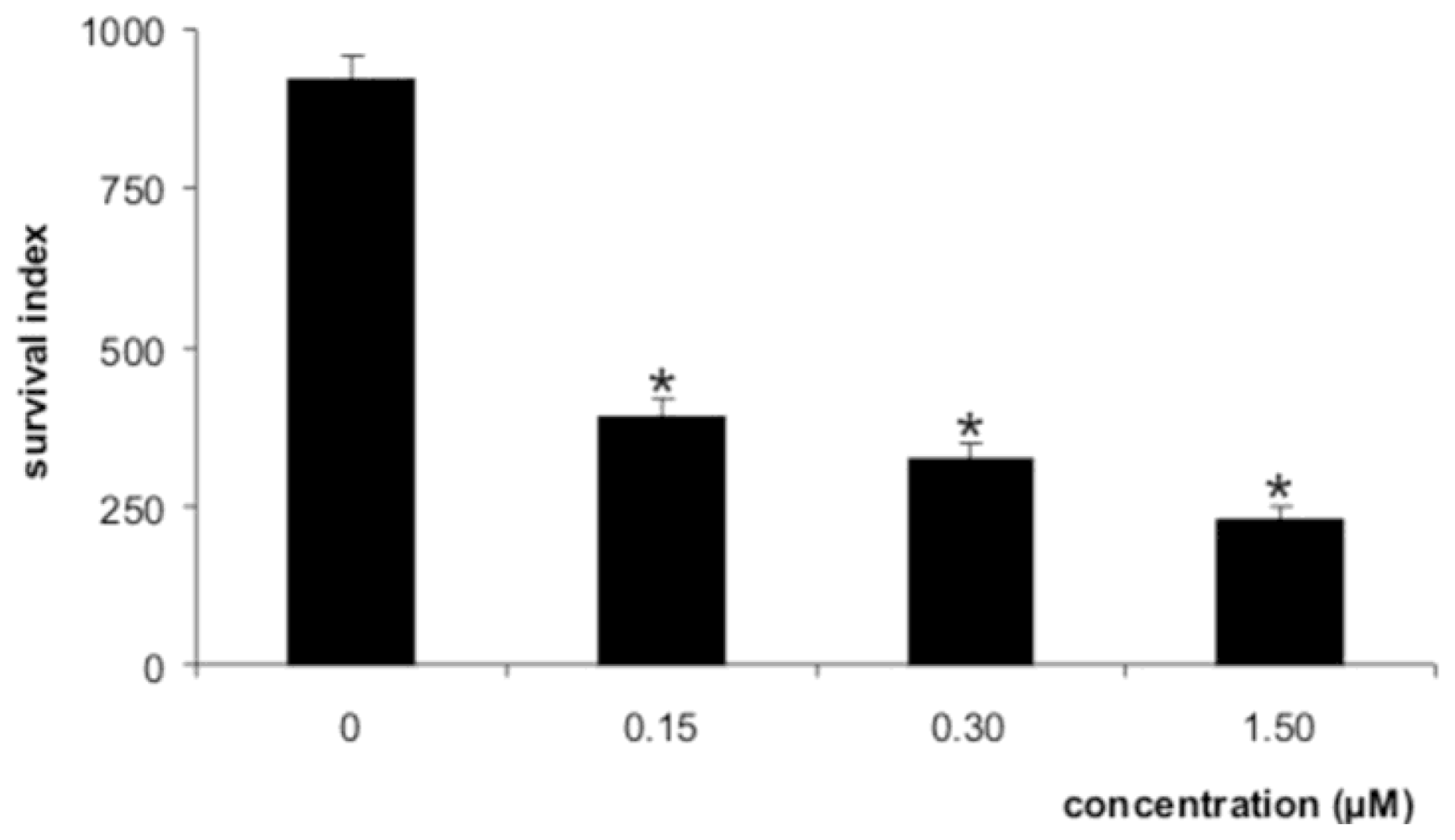
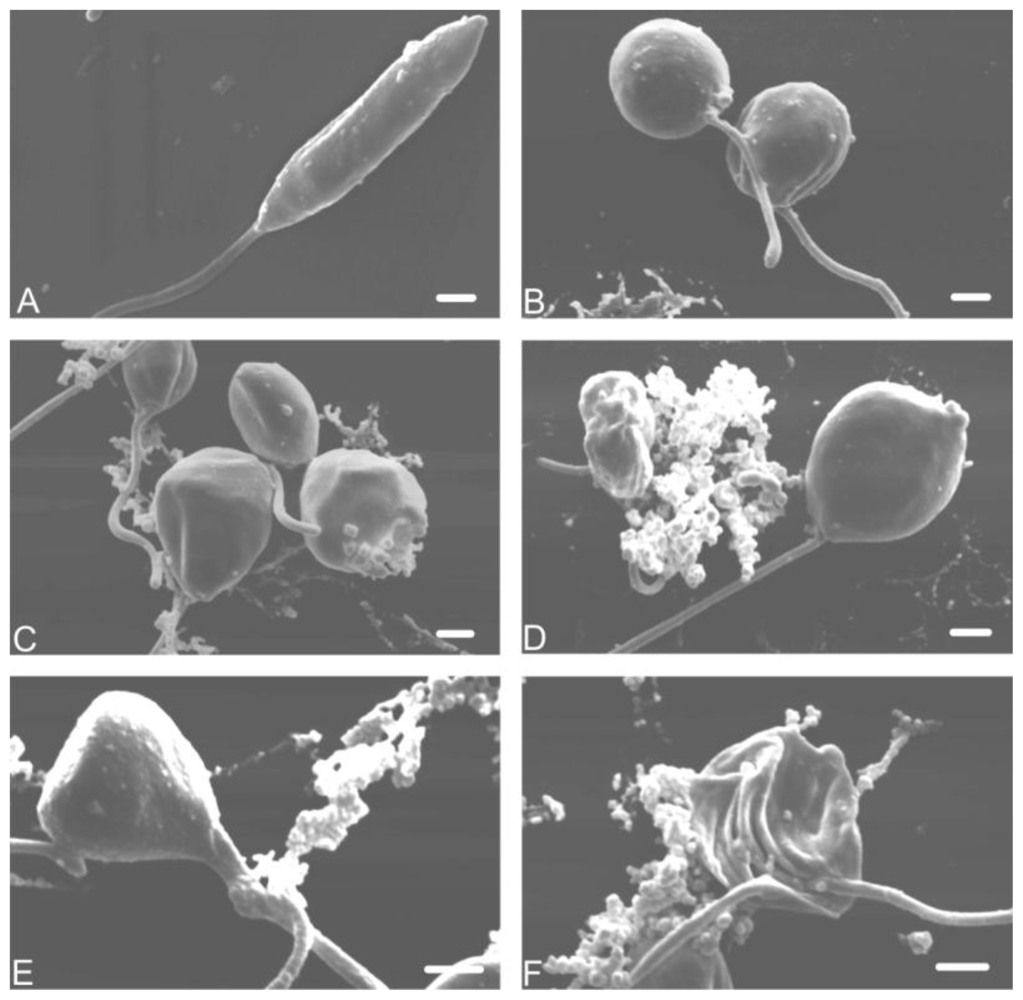
2. Results and Discussion
3. Experimental Section
3.1. Plant material and extraction procedures
3.2. Extraction procedures and elatol isolation
3.3. Spectroscopic data
3.4. Parasite and cell culture
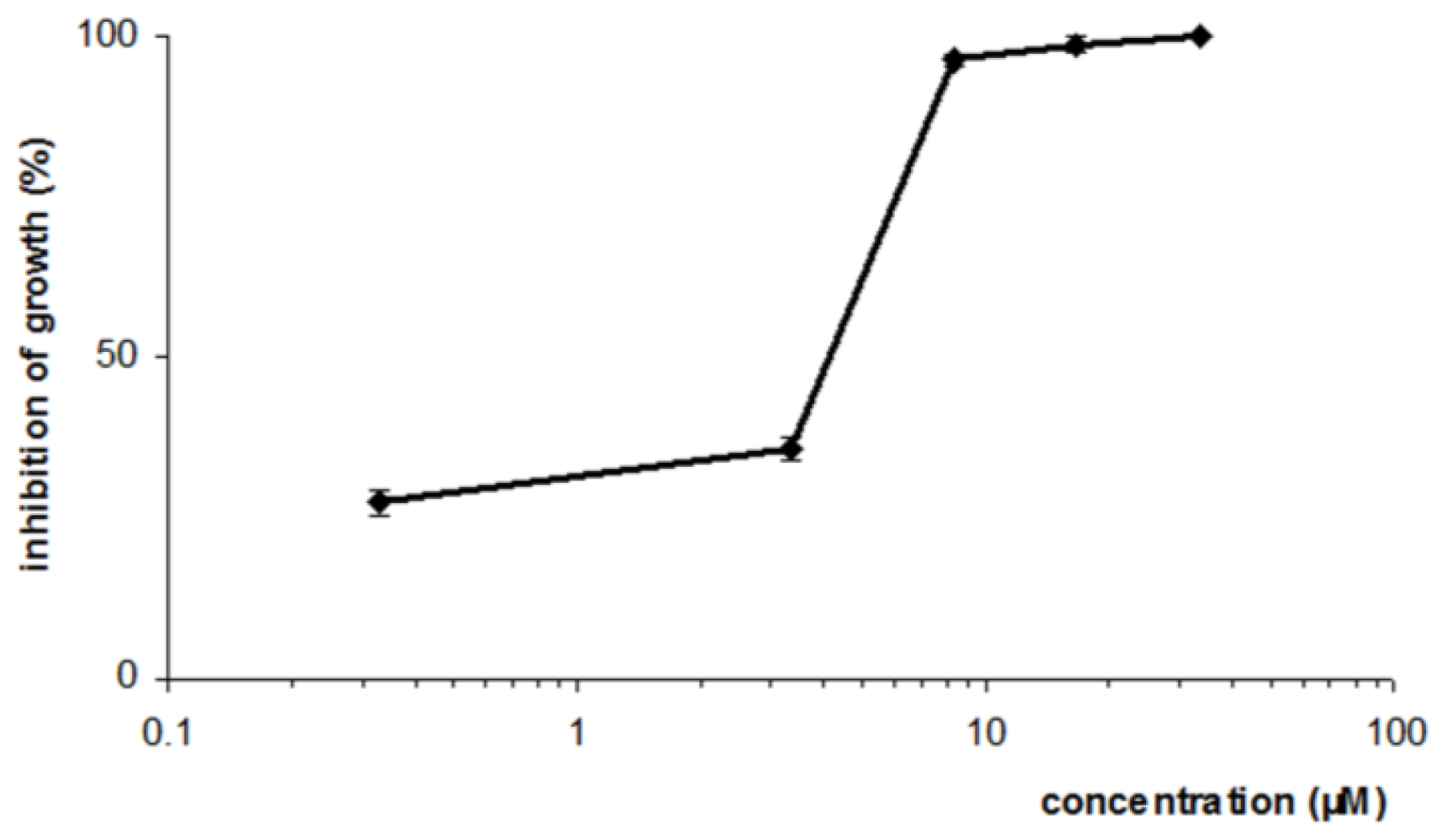
3.5. Antileishmanial activity
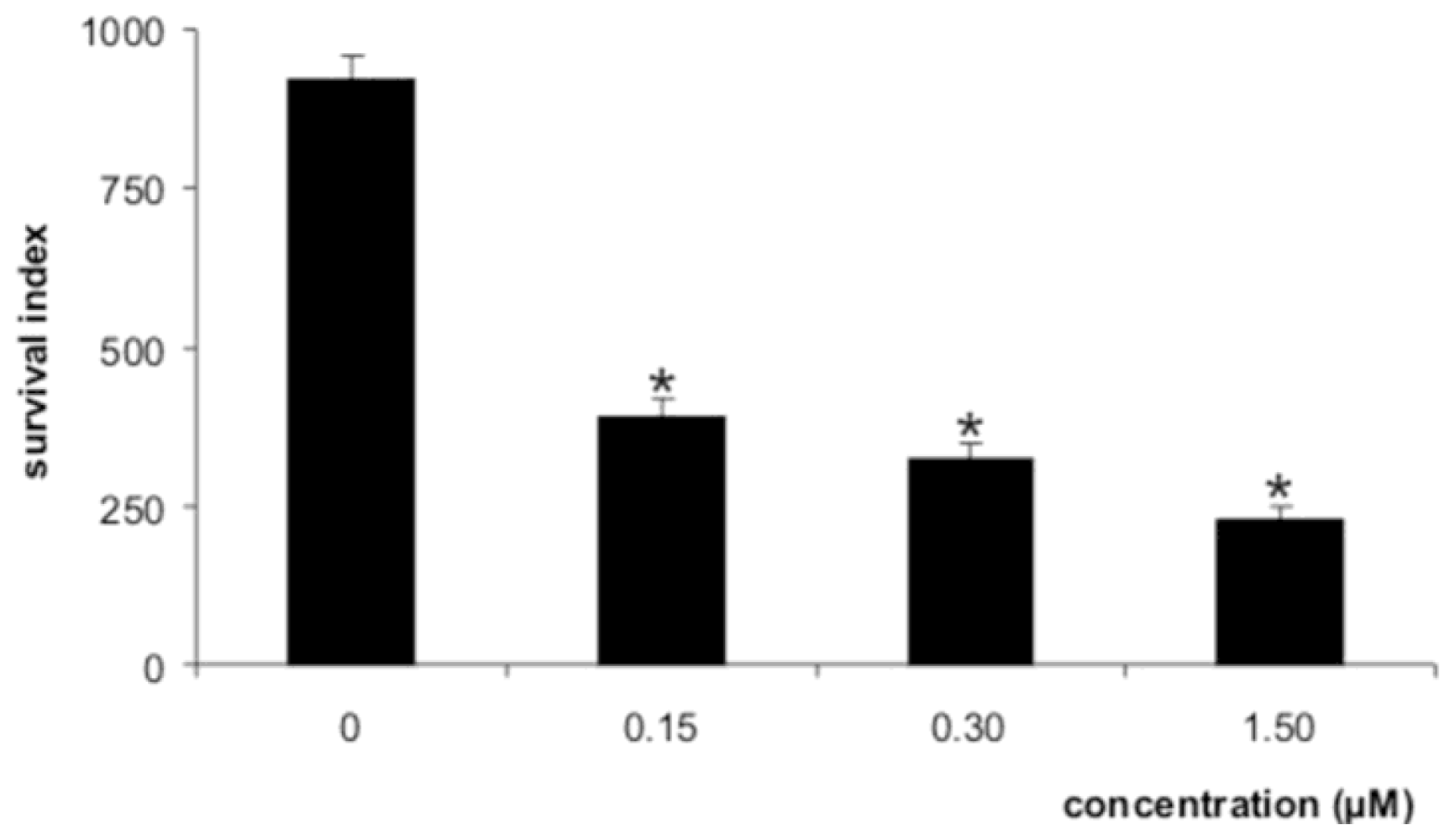
3.6. Activity against intracellular amastigotes
3.7. Cytotoxicity assay
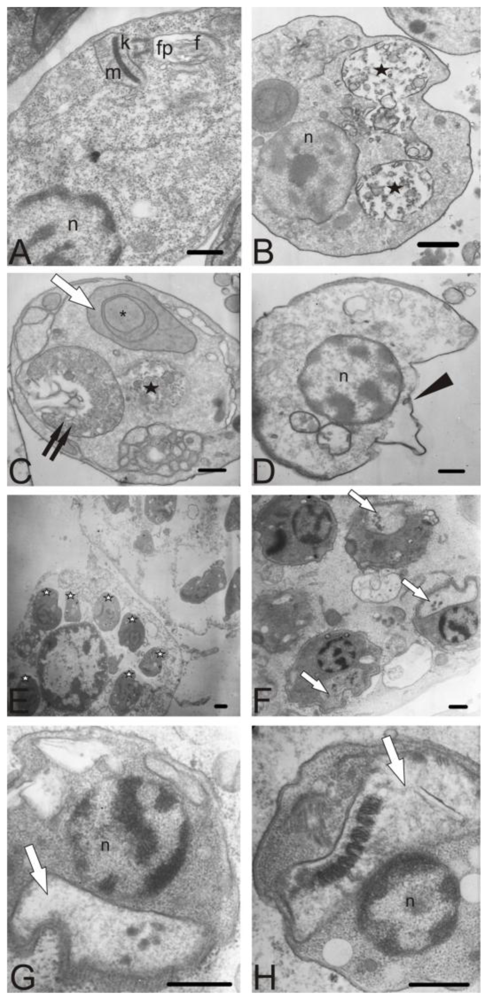
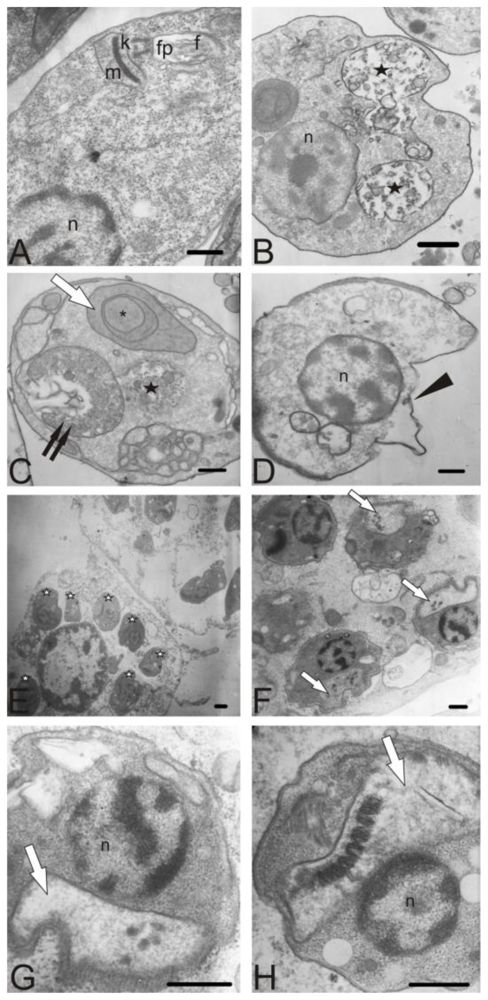
3.8. Electron microscopy
3.9. Statistical analysis
4. Conclusions
Acknowledgements
References
- Murray, HW; Berman, JD; Davies, CR; Saravia, NG. Advances in leishmaniasis. Lancet 2005, 366, 1561–1577. [Google Scholar]
- Bailey, MS; Lockwood, DNJ. Cutaneous leishmaniasis. Clin Dermatol 2007, 25, 203–211. [Google Scholar]
- Desjeux, P. Global control and leishmania HIV co-infection. Clin Dermatol 1999, 17, 317–325. [Google Scholar]
- Blum, J; Desjeux, P; Schwartz, E; Beck, B; Hatz, C. Treatment of cutaneous leishmaniasis among travellers. J Antimicrob Chemother 2004, 53, 158–166. [Google Scholar]
- World Health Organization, Leishmaniasis: Disease burden and epidemiological trends. In Special Programme for Research and Training in Tropical Diseases; WHO: Geneva, Switzerland, 2002; Available online: http://www.who.int/tdr/diseases/leish/files/direction.pdf accessed on 10 March 2010.
- Fernández-Guerrero, ML; Robles, P; Rivas, P; Mójer, F; Muniz, G; Górgolas, M. Visceral leishmaniasis in immunocompromised patients with and without AIDS: A comparison of clinical features and prognosis. Acta Trop 2004, 90, 11–16. [Google Scholar]
- Carnaúba, D, Jr; Konishi, CT; Petri, V; Martinez, ICP; Shimizu, L; Pereira-Chioccola, VL. Atypical disseminated leishmaniasis similar to post-kala-azar dermal leishmaniasis in a Brazilian AIDS patient infected with Leishmania (Leishmania) infantum chagasi: A case report. Int J Infect Dis 2009, 13, 504–507. [Google Scholar]
- Herwaldt, BL. Miltefosine—the long-awaited therapy for visceral leishmaniasis. N Engl J Med 1999, 341, 1840–1842. [Google Scholar]
- Carrión, J; Folgueira, C; Alonso, C. Immunization strategies against visceral leishmaniosis with the nucleosomal histones of Leishmania infantum encoded in DNA vaccine or pulsed in dendritic cells. Vaccine 2008, 26, 2537–2544. [Google Scholar]
- Momeni, AZ; Reiszadae, MR; Aminjavaheri, M. Treatment of cutaneous leishmaniasis with a combination of allopurinol and low-dose meglumine antimoniate. Int J Dermatol 2002, 41, 441–443. [Google Scholar]
- Natera, S; Machuca, C; Padrón-Nieves, M; Romero, A; Díaz, E; Ponte-Sucre, A. Leishmania spp.: Proficiency of drug-resistant parasites. Int J Antimicrob Agents 2007, 29, 637–642. [Google Scholar]
- Palumbo, E. Current treatment for cutaneous leishmaniasis: A review. Am J Ther 2009, 16, 178–182. [Google Scholar]
- Sims, JJ; Lin, GHY; Wing, RM. Marine natural products: Elatol, a halogenated sesquiterpene alcohol from the red alga Laurencia elata. Tetrahedron Lett. 1974, 39, 3487–3490. [Google Scholar]
- König, GM; Wright, AD. Sesquiterpene content of the antibacterial dichlormethane extract of the red alga Laurencia obtusa. Planta Med. 1997, 63, 186–187. [Google Scholar]
- Juagdan, EG; Kalidindi, R; Scheuer, P. Two new chamigranes from an hawaiian red alga, Luurenciu cartikzginea. Tetrahedron 1997, 2, 521–528. [Google Scholar]
- Vairappan, CS. Potent antibacterial activity of halogenated metabolites from Malaysian red algae, Laurencia majuscule (Rhodomelaceae, Ceramiales). Biomol Eng 2003, 20, 255–259. [Google Scholar]
- Lhullier, C; Donnangelo, A; Caro, M; Palermo, JA; Horta, PA; Falkenberg, M; Eloir, P; Schenkel, EP. Isolation of elatol from Laurencia microcladia and its palatability to the sea urchin Echinometra lucunter. Biochem. Syst. Ecol. 2009, 37, 254–259. [Google Scholar]
- Vairappan, CS; Anangdan, SP; Tan, KL; Matsunaga, S. Role of secondary metabolites as defense chemicals against ice-ice disease bacteria in biofouler at carrageenophyte farms. J Appl Phycol 2009, 22, 305–311. [Google Scholar]
- Veiga-Santos, P; Pelizzaro-Rocha, KJ; Santos, AO; Ueda-Nakamura, T; Dias-Filho, BP; Silva, SO; Sudatti, DB; Bianco, EM; Pereira, RC; Nakamura, CV. In vitro anti-trypanosomal activity of elatol isolated from red seaweed Laurencia dendroidea. Parasitology 2010, 14, 1–10. [Google Scholar]
- White, DE; Stewart, IC; Grubbs, RH; Stoltz, BM. The catalytic asymmetric total synthesis of elatol. J Am Chem Soc 2008, 23, 810–811. [Google Scholar]
- White, DE; Stewart, IC; Seashore-Ludlow, BA; Grubbs, RH; Stoltz, BM. A general enantioselective route to the chamigrene natural product family. Tetrahedron 2010, 26, 4668–4686. [Google Scholar]
- Wang, B; Zhang, W; Duan, X; Li, X. In vitro antioxidative activities of extract and semi-purified fractions of the marine red alga, Rhodomela confervoides (Rhodomelaceae). Food Chem 2009, 113, 1101–1105. [Google Scholar]
- Iliopoulou, D; Roussis, V; Pannecouque, C; De Clercq, E; Vagias, C. Halogenated sesquiterpenes from the red alga Laurencia obtusa. Tetrahedron 2002, 58, 6749–6755. [Google Scholar]
- Sudatti, DB; Rodrigues, SV; Pereira, RC. Quantitative GC-ECD analysis of halogenated metabolites: Determination of surface and within-thallus elatol of Laurencia obtusa. J. Chem. Ecol. 2006, 32, 835–843. [Google Scholar]
- Kladi, M; Vagias, C; Stavri, M; Rahman, MM; Gibbons, S; Roussis, V. C15 acetogenins with antistaphylococcal activity from the red alga Laurencia glandulifera. Phytochem. Lett. 2008, 1, 31–36. [Google Scholar]
- Sudatti, DB; Rodrigues, SV; Coutinho, R; Gama, BAP; Salgado, LT; Amado Filho, GM; Pereira, RC. Transport and defensive role of elatol at the surface of the red seaweed Laurencia obtusa (Ceramiales, Rhodophyta). J Phycol 2008, 44, 584–591. [Google Scholar]
- Salgado, LT; Viana, NB; Andrade, LR; Leal, RN; Gama, BAP; Attias, M; Pereira, RC; Amado Filho, GM. Intra-cellular storage, transport and exocytosis of halogenated compounds in marine red alga Laurencia obtusa. J. Struct. Biol. 2008, 162, 345–355. [Google Scholar]
- Vairappan, CS; Suzuki, M; Abe, T; Masuda, M. Antibacterial halogenated metabolites from the Malaysian Laurencia species. Phytochemistry 2001, 58, 291–297. [Google Scholar]
- Schaeffer, DJ; Krylov, VS. Anti-HIV activity of extracts and compounds from algae and cyanobacteria. Ecotoxicol Environ Saf 2000, 45, 208–227. [Google Scholar]
- Matsuhiro, B; Conte, AF; Damonte, EB; Kolender, AA; Matulewicz, MC; Mejías, EG; Pujol, CA; Zúniga, EA. Structural analysis and antiviral activity of a sulfated galactan from the red seaweed Schizymenia binderi (Gigartinales, Rhodophyta). Carbohydr Res 2005, 340, 2392–2402. [Google Scholar]
- Freile-Pelegrin, Y; Robledo, D; Chan-Bacab, MJ; Ortega-Morales, BO. Antileishmanial properties of tropical marine algae extracts. Fitoterapia 2008, 79, 374–377. [Google Scholar]
- Kang, JY; Khan, MNA; Park, NH; Cho, JY; Lee, MC; Fujii, H; Hong, YK. Antipyretic, analgesic, and anti-inflammatory activities of the seaweed Sargassum fulvellum and Sargassum thunbergii in mice. J Ethnopharmacol 2008, 116, 187–190. [Google Scholar]
- Moo-Puc, R; Robledo, D; Freile-Pelegrin, Y. Evaluation of selected tropical seaweeds for in vitro anti-trichomonal activity. J Ethnopharmacol 2008, 120, 92–97. [Google Scholar]
- Mayer, AMS; Rodríguez, AD; Berlinck, RGS; Hamann, MT. Marine pharmacology in 2005–2006: Marine compounds with anthelmintic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiprotozoal, antituberculosis, and antiviral activities, affecting the cardiovascular, immune and nervous systems, and other miscellaneous mechanisms of action. Biochim Biophys Acta 2009, 1790, 283–308. [Google Scholar]
- Orhan, I; Sener, B; Kaiser, M; Brun, R; Tasdemir, D. Inhibitory activity of marine sponge-derived natural products against parasitic protozoa. Mar Drugs 2010, 15, 47–58. [Google Scholar]
- Scala, F; Fattorusso, E; Menna, M; Taglialatela-Scafati, O; Tierney, M; Kaiser, M; Tasdemir, D. Bromopyrrole alkaloids as lead compounds against protozoan parasites. Mar Drugs 2010, 8, 2162–2174. [Google Scholar]
- Lazardi, K; Urbina, JA; Souza, W. Ultrastructural alterations induced by two ergosterol biosynthesis inhibitors, Ketoconazoleand Terbinafine, on epimastigotas and amastigotes of Trypanosoma (Schizotrypanum) cruzi. Antimicrob. Agents Chemother. 1990, 34, 2097–2105. [Google Scholar]
- Rodrigues, JCF; Attias, M; Rodriguez, C; Urbina, JA; Souza, W. Ultrastructural and biochemical alterations induced by 22,26-Azasterol, a Δ 24(25)-Sterol methyltransferase inhibitor, on promastigote and amastigote forms of Leishmania amazonensis. Antimicrob. Agents Chemother. 2002, 46, 487–499. [Google Scholar]
- Lorente, OS; Rodrigues, JCF; Jiménez, CJ; Joyce-Menekse, M; Rodrigues, C; Croft, SL; Yardley, V; Luca-Fradley, K; Ruiz-Pérez, LM; Urbina, J; Souza, W; Pacanowska, DG; Gilbert, IH. Novel azasterols as potential agents for treatment of leishmaniasis and trypanosomiasis. Antimicrob Agents Chemother 2004, 48, 487–499. [Google Scholar]
- Rodrigues, JCF; Bernardes, CF; Visbal, G; Urbina, JA; Vercesi, AE; Souza, W. Sterol methenyl transferase inhibitors alter the ultrastructure and function of the Leishmania amazonensis mitochondrion leading to potent growth inhibition. Protist 2007, 158, 447–456. [Google Scholar]
- Cassano, V. Taxonomia e filogenia do complexo Laurencia (Ceramiales, Rhodophyta), com ênfase no estado do Rio de Janeiro, Brasil. Ph.D. Thesis, Instituto de Botânica da Secretaria de Estado do Meio Ambiente, São Paulo, Brasil, 2009; p. 328. [Google Scholar]
- König, GM; Wright, AD. Laurencia rigida: Chemical investigations of its antifouling dichloromethane extract. J Nat Prod 1997, 60, 967–970. [Google Scholar]
- Warren, LG. Metabolism of Schizotrypanum cruzi Chagas. I. Effect of culture age and substrate concentration on respiratory rate. J Parasitol 1960, 46, 529–539. [Google Scholar]


© 2010 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Santos, A.O.d.; Veiga-Santos, P.; Ueda-Nakamura, T.; Filho, B.P.D.; Sudatti, D.B.; Bianco, É.M.; Pereira, R.C.; Nakamura, C.V. Effect of Elatol, Isolated from Red Seaweed Laurencia dendroidea, on Leishmania amazonensis. Mar. Drugs 2010, 8, 2733-2743. https://doi.org/10.3390/md8112733
Santos AOd, Veiga-Santos P, Ueda-Nakamura T, Filho BPD, Sudatti DB, Bianco ÉM, Pereira RC, Nakamura CV. Effect of Elatol, Isolated from Red Seaweed Laurencia dendroidea, on Leishmania amazonensis. Marine Drugs. 2010; 8(11):2733-2743. https://doi.org/10.3390/md8112733
Chicago/Turabian StyleSantos, Adriana Oliveira dos, Phercyles Veiga-Santos, Tânia Ueda-Nakamura, Benedito Prado Dias Filho, Daniela Bueno Sudatti, Éverson Miguel Bianco, Renato Crespo Pereira, and Celso Vataru Nakamura. 2010. "Effect of Elatol, Isolated from Red Seaweed Laurencia dendroidea, on Leishmania amazonensis" Marine Drugs 8, no. 11: 2733-2743. https://doi.org/10.3390/md8112733