Neurotoxins from Marine Dinoflagellates: A Brief Review


State Key Lab of Marine Environmental Science / Environmental Science Research Center, Xiamen University, Xiamen 361005, P.R. China
Mar. Drugs 2008, 6(2), 349-371; https://doi.org/10.3390/md6020349
Submission received: 9 March 2008
/
Revised: 14 May 2008
/
Accepted: 14 May 2008
/
Published: 11 June 2008
(This article belongs to the Special Issue Marine Toxins)
Abstract
:Dinoflagellates are not only important marine primary producers and grazers, but also the major causative agents of harmful algal blooms. It has been reported that many dinoflagellate species can produce various natural toxins. These toxins can be extremely toxic and many of them are effective at far lower dosages than conventional chemical agents. Consumption of seafood contaminated by algal toxins results in various seafood poisoning syndromes: paralytic shellfish poisoning (PSP), neurotoxic shellfish poisoning (NSP), amnesic shellfish poisoning (ASP), diarrheic shellfish poisoning (DSP), ciguatera fish poisoning (CFP) and azaspiracid shellfish poisoning (ASP). Most of these poisonings are caused by neurotoxins which present themselves with highly specific effects on the nervous system of animals, including humans, by interfering with nerve impulse transmission. Neurotoxins are a varied group of compounds, both chemically and pharmacologically. They vary in both chemical structure and mechanism of action, and produce very distinct biological effects, which provides a potential application of these toxins in pharmacology and toxicology. This review summarizes the origin, structure and clinical symptoms of PSP, NSP, CFP, AZP, yessotoxin and palytoxin produced by marine dinoflagellates, as well as their molecular mechanisms of action on voltage-gated ion channels.
1. Introduction
Over the past few decades, the occurrence of harmful algal blooms (HABs) has increased both in frequency and in geographic distribution in many regions of the world. This has resulted in adverse impacts on public health and the economy, and has become a global concern [1–3]. It is known that certain HAB species can produce potent toxins that impact human health through the consumption of contaminated shellfish, coral reef fish and finfish, or through water or aerosol exposure [4]. In many cases, toxic species are normally present in low concentrations with no environmental or human health impacts. However, when they are present at high cell density and are ingested by filter-feeding shellfish, zooplankton, and herbivorous fishes, toxins are accumulated in these organisms and transferred to higher trophic levels through the food chain, which results in various adverse effects. It is reported that algal toxins result in more than 50,000–500,000 intoxication incidents per year, with an overall mortality rate of 1.5% on a global basis [5]. In addition to their adverse effects on human health, algal toxins are responsible for the death of fish and shellfish and have caused episodic mortalities of marine mammals, birds, and other animals depending on the marine food web [6–9].
Of those causative organisms, dinoflagellates, a very large and diverse group of eukaryotic algae in the marine ecosystem, are the major group producing toxins that impact humans [4, 10]. Dinoflagellate toxins are structurally and functionally diverse, and many present unique biological activities. In the past few decades, extensive studies have been devoted to the toxicology and pharmacology of dinoflagellate toxins [11], and five major seafood poisoning syndromes caused by toxins have been identified from the dinoflagellates (Table 1): paralytic shellfish poisoning (PSP), neurotoxic shellfish poisoning (NSP), amnesic shellfish poisoning (ASP), diarrheic shellfish poisoning (DSP) and ciguatera fish poisoning (CFP). Besides these well-known poisonings, several new poisoning syndromes resulting from newly appearing dinoflagellate toxins, such as azaspiracid toxins, yessotoxin and palytoxin have been reported and characterized recently (Table 1), and this has increased global public concerns regarding dinoflagellate associated toxins. Dinoflagellate toxins can be functionally categorized as neurotoxins and hepatotoxins, according to their clinical symptoms. The neurotoxicity of dinoflagellate toxins is mediated by diverse, highly specific interactions with ion channels involved in neurotransmission (Figure 1). This paper provides a brief overview of the origin, structure and clinical symptoms of PSP, NSP, CFP, AZP, yessotoxin and palytoxin produced by dinoflagellates as well as their molecular mechanisms of action on voltage-gated ion channels.
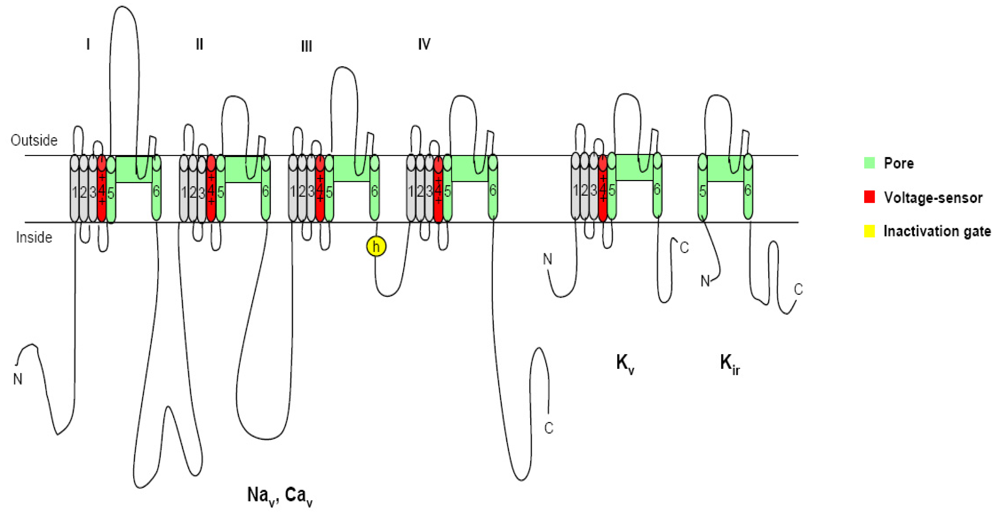
2. Voltage-gated ion channels and neurotoxins
It is known that most dinoflagellate toxins are neurotoxins, which interact with the specific receptors associated with neurotransmitter receptors, or voltage-sensitive ion channels (Figure 1), resulting in the observed neurotoxicity [12]. In organisms including humans, voltage-gated ion channels, such as sodium, calcium, and potassium channels, are electrical signal generators which control contraction of muscle, secretion of hormones, sensing of the environment, processing of information in the brain, and output from the brain to peripheral tissues [13]. These channels share a common structural motif containing six transmembrane segments (S1–S6) and a pore loop (Figure 1). The voltage sensor domain consists of the S1–S4 segments with positively charged residues in the S4 segment as gating charges, while the pore is composed of the S5/S6 segments and the pore loop between them, which are gated by bending of the S6 segment at a hinge glycine or praline residue. In all of these contexts, electrical signals are conducted by members of the ion channel protein super family, a set of more than 140 structurally related pore-forming proteins [14]. Pharmacological studies have disclosed that the functions of the voltage-gated ion channel proteins can be classified into three complementary aspects: ion conductance, pore gating and regulation. These channel proteins are the molecular targets for a broad range of potent neurotoxins, which strongly alter channel functions by binding to specific receptor sites. At present, six different neurotoxin receptor sites on the channel protein have been identified on voltage-gated ion channels using various neurotoxins [15–19]. Hydrophilic low molecular mass toxins and large polypeptide toxins block the channel pore physically and prevent ion conductance. Alkaloid toxins and related lipid soluble toxins alter voltage-dependent gating through binding to intramembranous receptor sites. On the contrary, poplypeptide toxins alter channel gating through binding to extracellular sites [20, 13]. In the section below we describe the molecular mechanisms of action of seven different neurotoxins produced by dinoflagellates.
3. Paralytic shellfish poisoning (PSP)
PSP is a worldwide marine toxin disease with both neurologic and gastrointestinal symptoms, which is caused by the consumption of shellfish contaminated by toxic dinoflagellates [21]. The first PSP event was reported in 1927 near San Francisco, USA, and was caused by a dinoflagellate, A. catenella, which resulted in 102 people being ill and six deaths [22]. Since then, members of three dinoflagellate genera have been reported to be the major sources of PSP toxins: Alexandrium, Gymnodinium, and Pyrodinium [23]. Paralytic shellfish toxins (PSTs) are produced in varying proportions by different dinoflagellate species and even by different isolates within a species. PSP toxins are heat-stable and water-soluble nonproteinaceous toxins. The basic structures of PSP toxins are 3,4-propinoperhydropurine tricyclic systems. Saxitoxin and its analogues can be divided into three categories: the carbamate compounds, which include saxitoxin, neosaxitoxin and gonyautoxins 1–4; the N-sulfocarbamoyl compounds, which include the C and B toxins; and finally the decarbamoyl compounds with respect to the presence or absence of 1-N-hydroxyl, 11-hydroxysulfate, and 21-N-sulfocarbamoyl substitutions as well as epimerization at the C-11 position (Figure 2). In the past few decades at least 24 structurally related imidazoline guanidinium PSP derivatives have been identified and characterized from dinoflagellate species [21, 24].
Saxitoxin is the most toxic and also the most well studied among the PSP associated toxins. In mice, its LD50 peritoneal is 3–10 μg/kg body weight and orally is 263 μg/kg body weight. The lethal oral dose in humans is 1 to 4 mg (5,000 to 20,000 mouse units), depending on the gender and physiological condition of the patient. It is rapidly absorbed through the gastrointestinal tract and excreted in the urine. The symptoms of PSP include a tickling sensation of the lips, mouth and tongue, numbness of the extremities, gastrointestinal problems, difficulty in breathing, and a sense of dissociation followed by complete paralysis [25]. In the case of serious intoxication, PSP leads to a variety of neurological symptoms culminating in respiratory arrest and cardiovascular shock or death [26]. Saxitoxin and its analogues are very dangerous compounds, with possible military potential and have been listed by the Organization for the Prohibition of Chemical Weapons (OPCW) as a Schedule 1 chemical intoxicant, the manufacture, use, transfer and reuse of which are now strictly regulated by the OPCW (Chemical Weapons Convention, September 1998, The Hague, Netherlands).
PSP toxins are the most well known potent neurotoxins that specifically and selectively bind the sodium channels on excitable cells [27]. In 1975, Hill postulated a plugging model for the binding of the sodium channel with saxitoxin [28]. In this model, the toxin molecular penetrates rather deeply inside the channel and plugs it, having formed an ion pair with an anionic site thought to be located near the bottom of the channel. However, this model could not explain the lack of anticipated steric interactions with other structurally unfolded toxins, the gonyautoxins. Later, Kao and Walker proposed a model which placed the toxin molecules on the outside edge of the channel with the guanidinium group on the top of the channel entrance [29]. Meanwhile Shimizu also suggested a three-point binding model involving two hydrogen bonds with the ketal OHs, and ion pairing of the guanidinium group with an anionic site on the outside surface of the membrane [30]. With the success of cloning of the sodium channel [31], more precise information regarding the toxin-binding mode has arisen from the molecular biological studies of the sodium channel. PSP toxins are now regarded as blocking agents that reduce the number of conducting Na+ channels by occupying some site near the outer opening in a 1:1 high affinity specific receptor binding. The extracellular loop sections of S1–S2 (P-loops) of the Na+ channel are considered to be the PSP toxins-binding site. They bind to the site on the voltage-dependent sodium channel with high affinity (Kd~2nM), which inhibits the temporary permeability of Na+ ions by binding tightly to receptor site 1 on the outside surface of the membrane very close to the external orifice of the sodium channel, preventing sodium ions from passing through the membranes of the nerve cells, and thus interfering with the transmission of signals along the nerves. The resulting widespread blockade prevents impulse-generation in peripheral nerves and skeletal muscles. Saxitoxin also affects skeletal muscle directly by blocking the muscle action potential without depolarizing cells, which abolishes peripheral nerve conduction but with no curare-like action at the neuromuscular junction. Surprisingly, selective pressure from the presence of STX in the natural environment can select for mutations in the ion selectivity filter that cause resistance to these toxins in the softshell clam Mya arenaria [32].
4. Neurotoxic Shellfish Poisoning (NSP)
NSP is caused by the ingestion of shellfish exposed to blooms of the dinoflagellate Kerenia brevis (formerly Gymnodinium breve) [33, 34]. This dinoflagellate species produces two types of lipid soluble toxins: hemolytic and neurotoxic [35], causing massive fish kills, bird deaths, and marine mammal mortalities [36, 37]. The neurotoxic toxins are known as brevetoxins, which are a suite of ladder-like polycyclic ether toxins. Brevetoxin congeners are of two types based on backbone structure: brevetoxin B backbone (type 1; PbTx-2, 3, 5, 6, 8, 9) and brevetoxin A backbone (type 2; PbTx-1, 7, 10) (Figure 3). Among them, PbTx-2T is the major brevetoxin produced by K. brevis [38]. Massive fishes are killed due to neurotoxin exposure, with the possible contribution of the hemolytic fraction. Recently neurotoxins were also found in other fish-killing flagellate species, Chatonella marina, C. antiqua, Fibrocapsa japonica, and Heterosigma akashiwo [39–41].
As with many of the known marine toxins, the brevetoxins are tasteless, odorless, and heat and acid stable (they survive heat up to 300°C). The mouse LD50 is 170 μg/kg body weight (0.15–0.27) intraperitoneally, 94 μg/kg body weight intravenously and 520 μg/kg body weight orally [42]. Pathogenic dose for humans is in the order of 42-72 mouse units. NSP presents itself as a milder gastroenteritis with neurologic symptoms compared with PSP. The symptoms of NSP include nausea, tingling and numbness of the perioral area, loss of motor control, and severe muscular pain [43, 44].
The mechanism of action of brevetoxins has been extensively studied, and brevetoxins are regarded as depolarizing substances that open voltage gated sodium ion channels in cell walls, leading to uncontrolled Na+ influx into the cell [45]. Experiments utilizing neuroblastoma cells and rat synaptosomes have shown that brevetoxins act on neurotoxin binding site 5 on the α-subunit of the voltage-dependent sodium channel in a 1:1 stoichiometry [46]. This action differs from that of PSP toxins which block the sodium channel and prevent sodium ions from passing through the membranes of nerve cells. This enhances the inward flow of Na+ ions into the cell by altering the membrane properties of excitable cell types, resulting in inappropriate opening of the channel under conditions in which it is normally closed, and it also inhibits channel inactivation [36, 45, 47-49]. The toxin appears to produce its sensory symptoms by transforming fast sodium channels into slower ones, which results in persistent activation and repetitive firing [50]. It was reported that brevetoxin could combine with a separate site on the gates of the sodium channel, causing the release of neurotransmitters from autonomic nerve endings. In particular, this can release acetylcholine, leading to smooth tracheal contraction, as well as massive mast cell degranulation [51]. Recently, LePage et al. demonstrated that brevetoxins also triggered Ca influx in rat cerebellar granule neurons. Derivatives PbTx-1, PbTx-2 and PbTx-3 produced a rapid and concentration-dependent increase in cytosolic [Ca2+], indicating that brevetoxin analogues display a range of efficacies to neurotoxin site 2 ligands and are activators of neurotoxin site 5, with PbTx-1 being a full agonist and other derivatives acting as partial agonists [52]. An early investigation also reported that conformational variation of brevetoxins induces a significant change in the gross shape of the molecule, which results in the loss of binding affinity and toxicity of the brevetoxins [46]
5. Ciguatera Fish Poisoning (CFP)
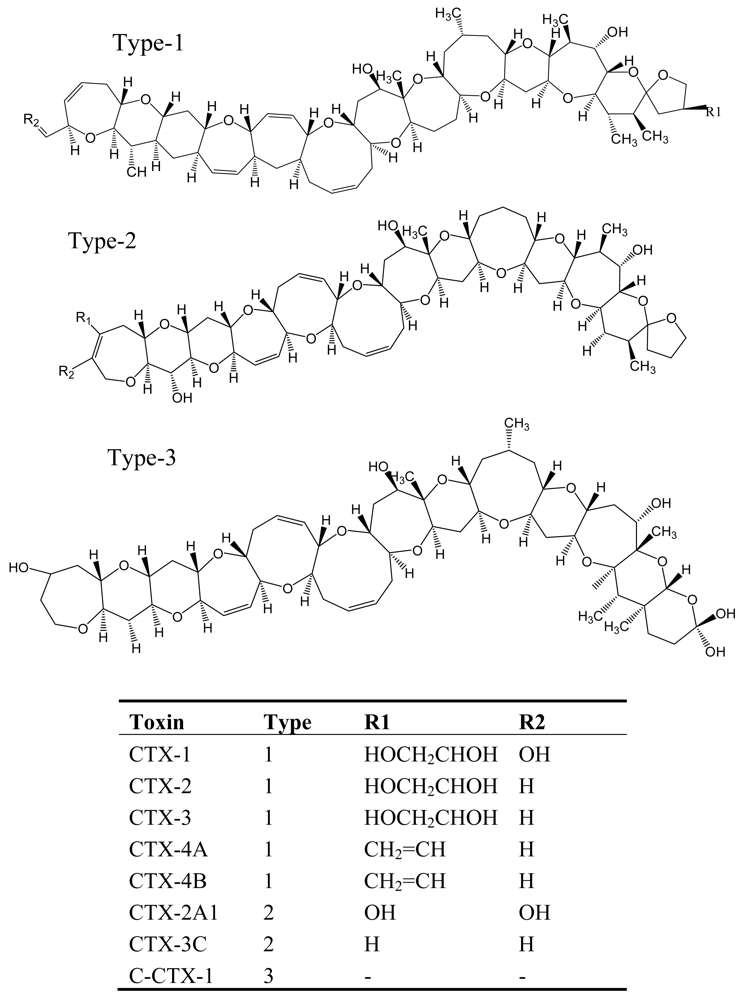
CFP, which is the most commonly reported marine toxin disease in the world, is caused by consumption of contaminated coral reef fishes such as barracuda, grouper, and snapper [53, 54]. It is estimated that approximately 25,000 people are affected annually by ciguatoxins and CFP is regarded as a world health problem [54]. The origin of ciguatera toxins has been identified in a dinoflagellate species, Gambierdiscus toxicus, which originally produces maitotoxins (MTXs), the lipophilic precursors of ciguatoxin [55]. These precursors are biotransformed to ciguatoxins by herbivorous fishes and invertebrates grazing on G. toxicus and then accumulated in higher trophic levels [56]. The ciguatoxins are a family of heat-stable, lipid-soluble, highly oxygenated, cyclic polyether molecules with a structural framework reminiscent of the brevetoxins [57–60], and more than 20 toxins may be involved in CFP (Figure 4) [53].
The biological activities of ciguatoxins have been studied extensively and they are regarded as the most potent activators of sodium and/or calcium fluxes in the cytoplasm in various cells. They produce more than 175 ciguateric symptoms, classified into four categories: gastrointestinal, neurological, cardiovascular and general symptoms [54, 61]. It should be emphasized that the symptoms of ciguatera vary in different oceans: in the Pacific Ocean neurological symptoms predominate, while in the Caribbean Sea the gastrointestinal symptoms dominate due to the difference in toxin composition. Ciguatoxin and maitotoxin are the two most common toxins associated with CFP, and they are the most lethal natural substances known. Pharmacological studies have revealed that CTXs activate the voltage-sensitive sodium channel at nM to pM concentrations [61]. In mice, ciguatoxin is lethal at 0.45 μg/kg ip, and maitotoxin at a dose of 0.15 μg/kg ip. Oral intake of as little as 0.1 μg ciguatoxin can cause illness in the human adult.
Ciguatoxins exert the same action mode as brevetoxins, which selectively target the common binding site 5 on the α-subunit of neuronal sodium channels. However, the affinity of ciguatoxins is higher than that of brevetoxins and, thus, the affinity of CTX-1 for voltage dependent sodium channels is around 30 times higher than that of brevetoxin. Ciguatoxins open sodium channels along the peripheral nerves, particularly at the nodes of Ranvier [62, 63], which results in an influx of Na+ ions, cell depolarization and the appearance of spontaneous action potentials in excitable cells. Consequently, the plasma membrane is unable to maintain either the internal environment of the cells or volume control due to the increased Na+ permeability, which results in alteration of bioenergetic mechanisms, cell and mitochondrial swelling and bleb formation on cell surfaces. With neurophysiological testing, significant slowing of sensory and motor nerve conduction velocities, and F wave latencies has been demonstrated [62, 64, 65]. This observation may be related to nodal swelling and internodal length and volume increase, all of which have been confirmed with in vitro CTX exposure [63, 66].
Studies on cardiovascular effects of ciguatoxins reveal that ciguatoxin affects voltage-dependent Na+ channels causing Na+ to move intracellularly, and normal cellular mechanisms begin to extrude sodium and take up calcium. Calcium is the intracellular trigger for muscle contraction. Although much of the increased calcium is buffered by the sarcoplasmic reticulum, it is likely that locally increased calcium concentrations increase the force of cardiac muscle contraction as is observed in ciguatoxin poisoning. A similar mechanism of ciguatoxin-induced intracellular transport of calcium occurs in intestinal epithelial cells. The increased concentration of intracellular calcium induced by ciguatoxin acts as a second messenger in the cell, which disrupts important ion-exchange systems, resulting in fluid secretion and symptoms of diarrhea [67].
Maitotoxin, another important neurotoxin involved in CFP, is a water soluble, ladder-shaped polycyclic molecule with numerous hydroxyl groups and sulfate groups (Figure 5). Three forms of MTX, MTX-1, MTX-2 and MTX-3 have been identified from G. toxicus [68]. MTX has been proved to be the most potent toxin identified on a weight basis: the LD50 of MTX in mice is less than 0.2 μg/Kg (intraperitoneally) and it is at least 5-fold more toxic than tetrodotoxin. Pharmacological studies demonstrate that MTX is a potent activator of voltage-gated calcium channels which stimulates the movement of Ca2+ ions across biomembranes in a wide variety of organisms. As a consequence of Ca2+ influx, maitotoxins can produce several effects: hormone and neurotransmitter secretion, phosphoinositides breakdown, and activation of voltage gated Ca2+ channels due to membrane depolarization. However, the primary target of MTX still remains undefined and the molecular mechanism of action is not clear. It is postulated that MTX might cause a shift in voltage-dependence of gating that favors opening of voltage-gated calcium channels at resting membrane potentials. However, MTX activates voltage-gated calcium channels indirectly via membrane depolarization as a consequence of activating a nonselective cation current [69]. Recently, Kakizaki et al. reported that maitotoxin induced a profound increase in the Ca2+ influx into cultured brainstem cells after a brief lag period, indicating Ca2+ permeability by acting on the calcium channel in an open state and preventing its closing [70].
6. Azaspiracid Shellfish Poisoning (AZP)
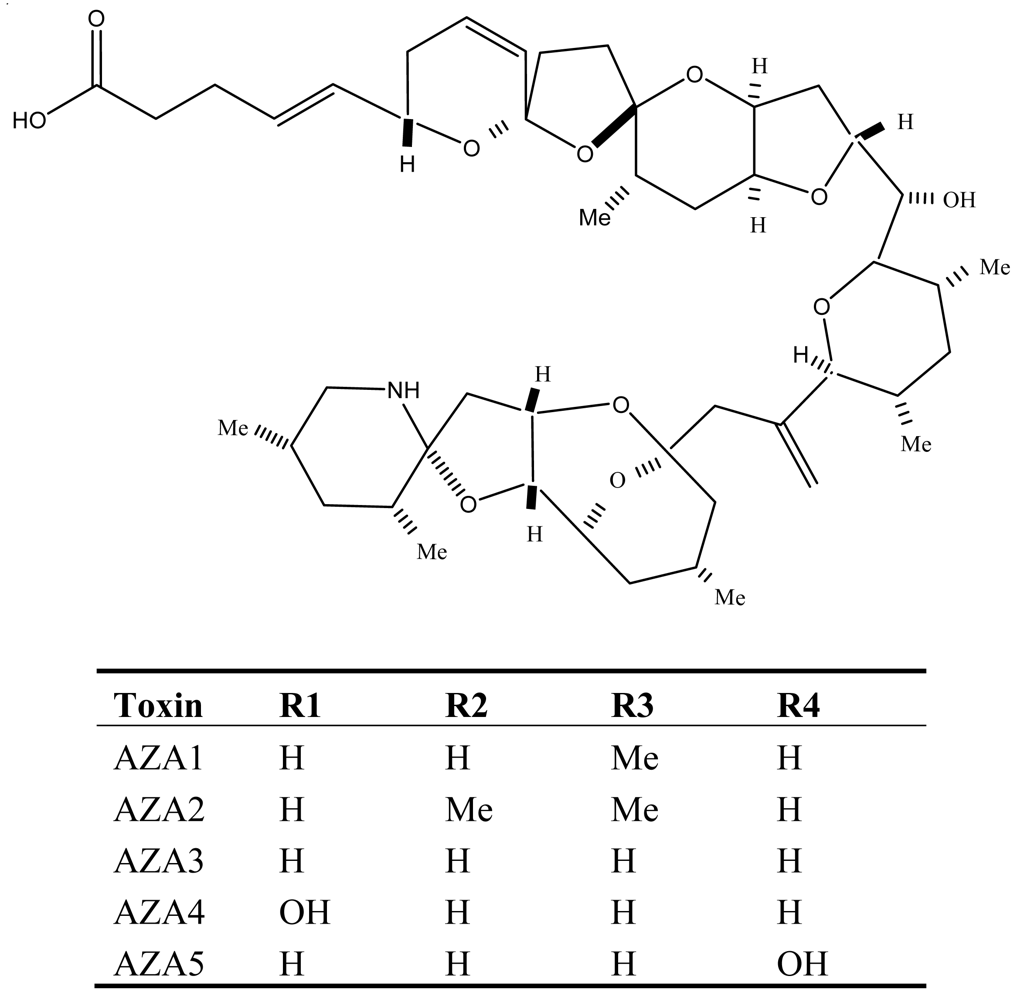
Azaspiracid poisoning (AZP), first reported from the Netherlands but later becoming a continuing problem in Europe [71], is a newly identified marine toxin disease. It is caused by consumption of contaminated shellfish associated with the dinoflagellate Protoperidinium crassipes, which can produce high intracellular concentrations of azaspiracid (AZA1), a lipophilic, polyether toxin. Nowadays about one dozen derivatives (AZA2 to 11) of azaspiracid (AZA1) have been identified and characterized from P. crassipes and contaminated shellfish [72–74]. AZAs differ significantly from other dinoflagellate toxins, in that they have unique structural features characterized by a tri-spiro assembly, an azazpiro ring fused with a 2,9-dinoxabicyclo[3.3.1] nonane and a terminal carboxylic acid group (Figure 6).
The symptoms of AZP include nausea, vomiting, severe diarrhea and stomach cramps. Neurotoxic symptoms were also observed [72, 75, 76]. However, the extremely limited availability of the pure toxins has impeded the necessary investigations of AZP. Some experiments carried out with mice showed that AZP, unlike okadaic acid (OA) and its analog, dinophysistoxin-1, which need an initiator [77], can cause lung tumor formation during repeated administration or after withdrawal of AZP without the combined use of any initiator [78]. Also the toxin can cause necrosis in the lamina propria of the small intestine and in lymphoid tissues such as the thymus, spleen and Peyer's patches [78]. The action mechanism of AZAs is unknown at present. Some studies indicate that AZAs might have different targets, since AZA1 and AZA2 increase [Ca2+]i by activation of Ca2+ -release from internal stores and Ca2+-influx, while AZA3 induces only Ca2+-influx. AZA5 does not modify intracellular Ca2+ homeostasis. Recent investigation of the effect of AZA4 on cytosolic calcium concentration [Ca2+]i in fresh human lymphocytes demonstrated that AZA4 inhibits store-operated Ca2+ channels (SOC channels) and Ca2+ influx and that this process is reversible [76]. It was postulated that AZA4 inhibits SOC channels by direct interaction with the channel pore, with another region of channel protein or with a closely associated regulatory protein and it was also found that AZA4 acts through another type of Ca2+ channel, probably some non selective cation channel usually activated by MTX [76]. AZA groups are novel inhibitors of Ca2+ channels, SOC and non-SOC channels. Further study is needed to determine the primary target and the molecular mechanisms of action of AZAs on Ca2+ channels.
7. Yessotoxin (YTX)
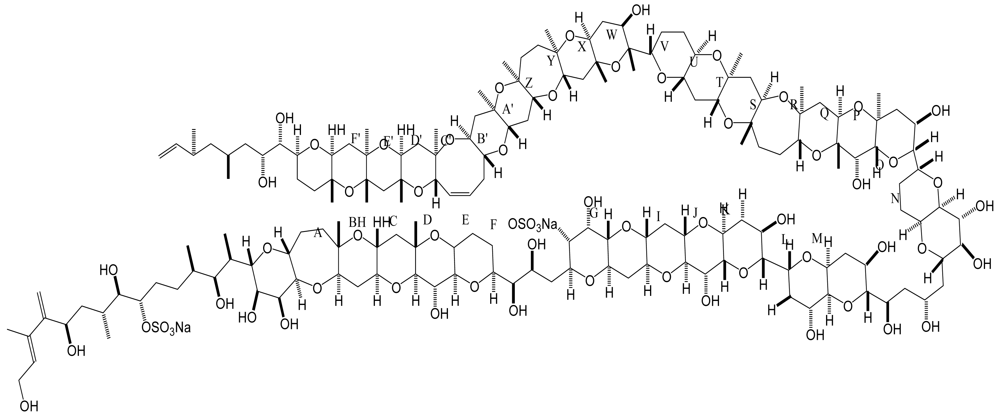
YTX and it analogues, which are disulphated polyether compounds of increasing occurrence in seafood, were originally isolated from the scallop Patinopecten yessoensis, collected at Mutsu Bay, Japan [79]. Since then, YTXs have been found in Europe, South America and New Zealand, and become a worldwide concern due to its potential risk to human health. YTXs were produced by three dinoflagellate species, Protoceratium reticulatum, Lingulodinium polyedrum and Gonyaulax spinifera [80–83].
YTX and its derivatives, 45-hydroxy YTX (45-OH-YTX), 45,46,47-trinor YTX, homo YTX, and 45-hydroxyhomo YTX [84, 85] are disulfated polyether lipophilic toxins originally isolated from Japanese scallops (Figure 7) [80]. Recently several new YTX analogues: carboxyyessotoxin (with a COOH group on the C44 of YTX instead of a double bond); carboxyhomoyessotoxin (with a COOH group on the C44 of homoYTX instead of a double bond); 42,43,44,45,46,47,55-heptanor-41-oxo YTX and 42,43,44,45,46,47,55-heptanor-41-oxohomo YTX in Adriatic mussels (M. galloprovincialis) have been identified in dinoflagellates [86, 87].
Originally, YTXs were classified among the toxins responsible for DSP, mainly because they appear and are extracted together with the DSP toxins, OA and the dinophysistoxins (DTXs) [80]. However, YTXs are proved to be not diarrheogenic compared to OA and its derivatives, the DTXs, which cause intestinal fluid accumulation or inhibition of protein phosphatase 2A. Terao et al. demonstrated that the heart is the main target organ of YTXs in mice [88]. Toxicological studies indicated that acute oral administration at doses up to 10 mg/kg YTX or repeated (seven days) oral exposure to high (2 mg/kg/day) doses of the toxin caused no mortality nor strong signs of toxicity in mice [89–91]. YTX caused motor discoordination in the mouse before death due to cerebellar cortical alterations [90, 92, 93]. Histopathological study revealed that YTX provoked alterations in the Purkinje cells of the cerebellum, including cytological damage to the neuronal cell body and change in the neurotubule and neurofilament immunoreactivity [93].
Recently it was demonstrated that YTX is a potent neurotoxin to neuronal cells. However, the action site and the mechanism are unknown [94]. YTX was observed to induce a two-fold increase in cytosolic calcium in cerebellar neurons that was prevented by the voltage-sensitive calcium channel antagonists nifedipine and verapamil. These results suggest YTX might interact with calcium channels and/or sodium channels directly. Previous studies also showed that YTX activated nifedipine-sensitive calcium channels in human lymphocytes [95], and YTX was postulated to activate non-capacitative calcium entry and inhibit capacitive calcium entry by emptying of internal calcium stores.
8. Palytoxin (PTX)
PTX is a polyhydroxylated compound that shows remarkable biological activity at an extremely low concentration [96]. This toxin was first isolated from the soft coral Palythoa toxica and subsequently from many other organisms such as seaweeds and shellfish. Recently, palytoxin was also found in a benthic dinoflagellate, Ostrepsis siamensis, which caused blooms along the coast of Europe [97–102], extensive death of edible mollusks and echinoderms [99, 100] and human illnesses [98, 99]. Cases of death resulting from PTX have been reported to be due to consumption of contaminated crabs in the Philippines [103], sea urchins in Brazil [104] and fish in Japan [105–107]. PTX has become of worldwide concern due to its potential impact on animals including humans.
PTX is a large, very complex molecule with both lipophilic and hydrophilic regions, and has the longest chain of continuous carbon atoms in any known natural product (Figure 8). Recently several analogues, ostreocin-D (42-hydroxy-3, 26-didemethyl-9,44-dideoxypalytoxin) and mascarenotoxins were identified in O. siamensis. PTX is regarded as one of the most potent toxins so far known [108], the LD50s 24 h after intravenous injection vary from 0.025 μg/kg in rabbits and about the same in dogs to 0.45 μg/kg in mice, with monkeys, rats and guinea pigs around 0.9 μg/kg. Toxic symptoms include fever inaction, ataxia, drowsiness, and weakness of limbs followed by death.
Over the past few decades much effort has been devoted to define the action mechanisms of PTXs, however these have not been identified. Pharmacological and electrophysiological studies have demonstrated that PTXs act as a haemolysin and alter the function of excitable cells. PTX selectively binds to the Na+, K+-ATPase with a Kd of 20 pM [109] and transforms the pump into a channel permeable to monovalent cations with a single-channel conductance of 10 pS [110–113]. Presently, three primary sites of action of PTXs have been postulated: PTX first opens a small conductance, non-selective cationic channel which results in membrane depolarization, K+ efflux and Na+ influx. Subsequently, the membrane depolarization may open voltage dependent Ca2+ channels in synaptic nerve terminals, cardiac cells and smooth muscle cells, while Na+ influx may load cells with Na+ and favor Ca2+ uptake by the Na+/Ca2+ exchanger in synaptic terminals, cardiac cells and vascular smooth muscle cells. Then the increase of [Ca2+li stimulates the release of neurotransmitters by nerve terminals, of histamine by mast cells and of vasoactive factors by vascular endothelial cells as a signal. It also induces contractions of striated and smooth muscle cells. Additional effects of a rise in [Ca2+]i may be activation of phospholipase C [114] and phospholipase A2 [115]. There are reports that PTX opens an H+ conductive pathway which results in activation of the Na+/H+ exchanger [116, 117]. Other investigators suggest that PTX raises [Ca2+]i independently of the activity of voltage dependent Ca2+ channels and Na+/Ca’+ exchange [118]. The last two actions might act as the opening of H+ specific and Ca2+ specific channels. Overall, PTX might posses more than one site of action in excitable cells and act as an agonist for low conductance channels conducting Na+/K+, Ca2+ and H+ ions.
9. Summary
This paper briefly outlines the origin, structure, symptoms and molecular action mechanisms of neurotoxins produced by marine dinoflagellates. These toxins vary in chemical structure and mechanism of action, and produce very distinct biological effects, which provides a potential application of these toxins in pharmacology and toxicology. However, some of them have not been well studied due to the limited supply of pure toxins and their molecular action mechanisms are unknown. Moreover, novel species of neurotoxins produced by dinoflagellates have been found and identified, which provide a challenge for the characterization of their toxin mechanisms and their effects on marine organisms and humans. Further work using the cell-based approach is needed to determine the precise mode of action of these novel neurotoxins from marine dinoflagellates.
Acknowledgements
The authors thank Prof. John Hodgkiss for helping to revise the manuscript. This work was partially supported by research grants from the Ministry of Science and Technology of the People’s Republic of China (Project No. 2005DFA20430), the National Natural Science Foundation of China (40376032 and 40476053), Fujian Provincial Department or Science and Technology, the Excellent Group and the Program for New Century Excellent Talents in Xiamen University to Prof. D.-Z. Wang.
References and Notes
- Anderson, DM. Okaichi, T, Anderson, DM, Nemoto, T, Eds.; Toxic algal blooms and red tides: a global perspective. In Red Tides: Biology, Environmental Science and Toxicology; Elsevier: New York, 1989; pp. 11–16. [Google Scholar]
- Smayda, TJ. Graneli, E, Sundstrom, B, Edler, L, Anderson, DM, Eds.; Novel and nuisance phytoplankton blooms in the sea: evidence for a global epidemic. In Toxic Marine Phytoplankton; Elsevier: New York, 1990; pp. 29–40. [Google Scholar]
- Hallegraeff, GM. Hallegraeff, GM, Anderson, DM, Cembella, AD, Eds.; Harmful algal blooms: a Global review. In Manual on Harmful Marine Microalgae; UNESCO: Landais, France, 2005; pp. 25–49. [Google Scholar]
- Van Dolah, FM. Botana, L, Ed.; Diversity of Marine and Freshwater Algal Toxins. In Seafood Toxicology: Pharmacology, Physiology and Detection; Marcel Dekker: New York, 2000; pp. 19–43. [Google Scholar]
- Quod, JP; Turquet, J. Ciguatera fish poisoning in Reunion island (SW Indian Ocean): epidemiology and clinical patterns. Toxicon 1996, 34, 779–785. [Google Scholar]
- Geraci, JR; Anderson, DM; Timperi, RJ; St Aubin, DJ; Early, GA; Prescott, JH; Mayo, CA. Humpback whales (Megaoetera novaeangliae) fatally poisoned by dinoflagellate toxin. Can J Fish Aq Sc 1989, 46, 1895–1898. [Google Scholar]
- Landsberg, JH; Steidinger, K. Reguera, B, Blanco, J, Fernandez, ML, Wyatt, T, Eds.; A historical review of red tide events caused by Gymnodinium breve as related to mass mortalities of the endangered manatee (Trichechus manatus latirostris) in Florida, USA. In Harmful Microalgae; IOC of UNESCO and Xunta de Galicia: Spain, 1998; pp. 97–100. [Google Scholar]
- Scholin, CA; Gulland, F; Doucette, GJ; Benson, S; Busman, M; Chavez, FP; Cordaro, J; DeLong, R; De Vogelaere, A; Harvey, J; Haulena, M; Lefebvre, K; Lipscomb, T; Loscutoff, S; Lowenstine, LJ; Marin, R, III; Miller, PE; McLellan, WA; Moeller, PDR; Powell, CL; Rowles, T; Silvagni, P; Silver, M; Spraker, T; Trainer, V; Van Dolah, FM. Mortality of sea lions along the central California coast linked to a toxic diatom bloom. Nature 2000, 430, 80–84. [Google Scholar]
- Flewelling, LJ; Naar, JP; Abbott, JP; Baden, DG; Barros, NB; Bossart, GD; Bottein, MY; Hammond, DG; Haubold, EM; Heil, CA; Henry, MS; Jacocks, HM; Leighfield, TA; Pierce, RH; Pitchford, TD; Rommel, SA; Scott, PS; Steidinger, KA; Truby, EW; Van Dolah, FM; Landsberg, JH. Brevetoxicosis: red tides and marine mammal mortalities. Nature 2005, 435, 755–756. [Google Scholar]
- Trainer, VL; Baden, DG. High affinity binding of red tide neurotoxins to marine mammal brain. Aquat Toxicol 1999, 46, 139–128. [Google Scholar]
- Botana, LM. Seafood and Freshwater toxins: pharmacology, physiology, and detection; Marcel Dekker: New York, 2000. [Google Scholar]
- Gessner, BD. Botana, LM, Ed.; Neurotoxic toxins. In Seafood and Freshwater toxins: Pharmacology, Physiology and Detection; Marcel Dekker: New York, 2000; pp. 65–90. [Google Scholar]
- Catterall, WA; Cestele, S; Yarov-Yarovoy, V; Yu, FH; Konoki, K; Scheuer, T. Voltage-gated channels and gating modifier toxins. Toxicon 2007, 49, 124–141. [Google Scholar]
- Yu, FH; Catterall, WA. The VGL-chanome: a protein superfamily specialized for electrical signaling and ionic homeostasis. Science STKE 2004, re 15. [Google Scholar]
- Catterall, WA. Neurotoxins that act on voltage-sensitive sodium channels in excitable membranes. Annu Rev Pharmacol Toxicol 1980, 20, 15–43. [Google Scholar]
- Martin-Eauclaire, MF; Couraud, F. Chang, LW, Dyer, RS, Eds.; Scorpion neurotoxins: effects and mechanisms. In Handbook of Neurotoxicology; Marcel-Dekker: New York, USA, 1992; pp. 683–716. [Google Scholar]
- Catterall, WA; Risk, M. Toxin T46 from Ptychodiscus brevis (formerly Gymnodinium breve) enhances activation of voltage-sensitive sodium channels by veratridine. Mol Pharmacol 1981, 19, 345–348. [Google Scholar]
- Poli, MA; Mende, TJ; Baden, DG. Brevetoxins, unique activators of voltage-sensitive sodium channels, bind to specific sites in rat brain synaptosones. Mol Pharmacol 1986, 30, 129–135. [Google Scholar]
- Fainzillber, M; Kofman, O; Zoltkin, E; Gordon, D. A new neurotoxin receptor site on sodium channels is identified by a conotoxin that affects sodium channel inactivation in mollusks and acts as an antagonist in rat brain. J Biol Chem 1994, 269, 2574–2580. [Google Scholar]
- Cestele, S; Catterall, WA. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar]
- Kodama, M. Botana, L, Ed.; Ecology, classification, and origin. In Seafood and freshwater toxins: Pharmacology, Physiology and Detection; Marcel Dekker: New York, 2000; pp. 125–150. [Google Scholar]
- Sommer, H; Meyer, KF. Paralytic shellfish poisoning. Arch Pathol 1937, 24, 560–598. [Google Scholar]
- Shumway, SE. A review of the effects of algal blooms on shellfish and aquaculture. J World Aquac Soc 1990, 21, 65–104. [Google Scholar]
- Shimizu, Y. Microalgal metabolites: a respective. Ann Rev Microbiol 1996, 50, 431–465. [Google Scholar]
- Halsetead, BW. Poisonous and venomous marine animals of the world; Princeton: Darwin, 1978. [Google Scholar]
- Lagos, N; Andrinolo, D. Botana, LM, Ed.; Paralytic shellfish poisoning (PSP): toxicology and kinetics. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection; Marcel Dekker: New York, USA, 2000; pp. 203–215. [Google Scholar]
- Kao, CY. Tetrodotoxin, saxitoxin and their significance in the study of excitation phenomena. Pharmacol Rev 1966, 18, 997–1049. [Google Scholar]
- Hill, B. The receptor for tetrodotoxin and saxitoxin: a structural hypothesis. Biophys J 1975, 15, 615–619. [Google Scholar]
- Kao, CY; Walkwe, SE. Active groups of saxitoxin and tetrodotoxin as deduced from action of saxitoxin analogs on frog muscle and squid axon. J Physiol 1982, 323, 619–637. [Google Scholar]
- Shimizu, Y. Recent progress in marine toxin research. Pure Appl Chem 1980, 54, 1973–1980. [Google Scholar]
- Numa, S; Noda, M. Molecular structure of sodium channels. Ann NY Acad Sci 1986, 479, 338–355. [Google Scholar]
- Bricelj, VM; Connel, L; Konoki, K; Macquarrie, SP; Scheuer, T; Catterall, WA; Trainer, VL. Sodium channel mutation leading to saxitoxin resistance in clams increase risk of PSP. Nature 2005, 434, 763–767. [Google Scholar]
- Steidinger, KA. Phytoplankton ecology: A conceptual review based on eastern. Gulf of Mexico research. CRC Crit Rev Microbiol 1973, 3, 49–67. [Google Scholar]
- Baden, DG. Marine food-borne dinoflagellate toxins. Int Rev Cytol 1983, 82, 99–150. [Google Scholar]
- Baden, DG; Mende, TJ. Toxicity of two toxins from the Florida red tide marine dinoflagellate Gymnodinium breve. Toxicon 1982, 20, 457–461. [Google Scholar]
- Poli, M; Mende, TJ; Baden, DG. Brevetoxins, unique activators of voltage-sensitive sodium channels bind to specific sites in rat brain synaptosomes. Mol Pharmacol 1986, 30, 129–135. [Google Scholar]
- Baden, D; Fleming, LE; Bean, JA. deWolf, FA, Ed.; Chapter: Marine Toxins. In Handbook of Clinical Neurology: Intoxications of the Nervous System Part H. Natural Toxins and Drugs; Elsevier: Amsterdam, 1995; pp. 141–175. [Google Scholar]
- Baden, DG; Bourdelais, AJ; Jacocks, H; Michelliza, S; Naar, J. Natural and Derivative Brevetoxins: Historical Background, Multiplicity, and Effects. Environ Health Persp 2005, 113. [Google Scholar]
- Sagir Ahmed, MD; Arakawa, O; Onoue, Y. Lassus, P, Arzul, G, Erhard, E, Gentien, P, Marcaillou, C, Eds.; Toxicity of cultured Chatonella marina. In Harmful Marine Algal Blooms; Lavoisier: Paris, 1995; pp. 499–504. [Google Scholar]
- Khan, S; Arakawa, O; Onoue, Y. Neurotoxins in a toxic red tide of Heterosigma akashiwo (Raphidophyceae) in Kagoshima Bay, Japan. Aquacul Res 1997, 28, 9–14. [Google Scholar]
- Hallegraeff, GM; Munday, BL; Baden, DG; Whitney, PL. Reguera, B, Blanco, J, Ferandz, ML, Wyatt, TEd, Eds.; Chatonnella maria raphidophyte bloom associated with mortality of cultured bluefin tuna (Thunnus maccoyii) in south Australia. In Harmful algae; Xunta de Galacia and IOC: Santiago de Compostela, Spain, 1998; pp. 93–96. [Google Scholar]
- Kirkpatrick, B; Fleming, LE; Squicciarini, D; Backer, LC; Clark, R; Abraham, W; Benson, J; Chenge, YS; Johnson, D; Pierce, R; Zaias, J; Bossart, GD; Baden, DG. Literature review of Florida red tide: Implications for human health effects. Harmful algae 2004, 3, 99–115. [Google Scholar]
- Morris, PD; Campbell, DS; Taylor, TJ; Freeman, JI. Clinical and epidemiological features of neurotoxic shellfish poisoning in North Carolina. Am J Public Health 1991, 81(4), 471–474. [Google Scholar]
- Baden, DG; Adams, DJ. Botana, LM, Ed.; Brevetoxins: Chemistry, mechanism of action, and methods of detection. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection; 2000; Marcel Dekker: New York; pp. 505–532. [Google Scholar]
- Baden, DG. Marine food-born dinoflagellate toxins. Int Rev Cytol 1983, 82, 99–150. [Google Scholar]
- Rein, KS; Baden, DG; Gawley, RE. Conformational analysis of the sodium channel modulator, brevetoxin A, comparison with brevetoxin B conformations, and a hypothesis about the common pharmacophore of the “site” toxins. J Org Chem 1994, 59, 2101–2106. [Google Scholar]
- Gallagher, P; Shinnick-Gallagher, P. Effect of brevetoxin in the rat phrenic nerve diaphragm preparation. Brit J Pharmacol 1980, 69, 367–372. [Google Scholar]
- Halstead, BW. Poisonous and Venomous Marine Animals of the World; Darwin Press: Princeton, 1988. [Google Scholar]
- Trainer, VL; Thomsen, WJ; Catterall, WA; Baden, DG. Photoaffinity labeling of the brevetoxin receptor on sodium channels in rat brain synaptosomes. Mol Pharmacol 1991, 40, 988–994. [Google Scholar]
- Watters, MR. Organic neurotoxins in seafoods. Clin Neurol Neurosurg 1995, 97, 119–124. [Google Scholar]
- Fleming, LE; Baden, DG. Neurotoxic Shellfish Poisoning: Public Health and Human Health Effects. White Paper for the Proceedings of the Texas Conference on Neurotoxic Shellfish Poisoning, Proceedings of the Texas NSP Conference, Corpus Christi (Texas), April, 1998; pp. 27–34.
- LePage, KT; Baden, DG; Murray, TF. Brevetoxin derivatives act as partial agonists at neurotoxin site 5 on the voltage-gated Na+ channel. Brain Res 2003, 959, 120–127. [Google Scholar]
- Guzman-Perez, SE; Park, DL. Botana, L, Ed.; Ciguatera toxins: Chemistry and diction. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection; Marcel Dekker: New York, 2000; pp. 401–418. [Google Scholar]
- Terao, K. Botana, L, Ed.; Ciguatera toxins: toxicology. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection; Marcel Dekker: New York, 2000; pp. 449–472. [Google Scholar]
- Yasumoto, T; Nakajima, I; Bagnis, R; Adachi, R. Finding of a dinoflagellate as a likely culprit of ciguatera. Jpn Soc Sci Fish 1977, 43, 1021–1026. [Google Scholar]
- Legrand, AM. Reguera, B, Blanco, J, Fernandez, M, Wyatt, T, Eds.; 1998; Ciguatera toxins: origin, transfer through the food chain and toxicity to humans. In Harmful Algae, Proceedings of the VIII International Conference on Harmful Algae; Xunta de Galicia and IOC of UNESCO: Vigo, Spain, 1999; pp. 39–43.
- Scheuer, PJ; Takahashi, W; Tsutsumi, J; Yoshida, T. Ciguatoxin: isolation and chemical nature. Science 1967, 155, 1267–1268. [Google Scholar]
- Tachibana, K; Nukina, M; John, YD; Scheuer, PJ. Recent developments in the molecular structure of ciguatoxin. Biol Bull 1987, 172, 122–127. [Google Scholar]
- Murata, M; Legrand, AM; Ishibashi, Y; Yasumoto, T. Structures and configurations of ciguatoxin from the moray eel Gymnothorax javanicus and its likely precursor from the dinoflagellate Gambierdiscus toxicus. J Am Chem Soc 1990, 112, 4380–4386. [Google Scholar]
- Lewis, RJ; Vernoux, J-P; Brereton, IM. Structure of Caribbean ciguatoxin isolated from Caranx latus. J Am Chem Soc 1998, 120, 5914–5920. [Google Scholar]
- Lewis, RJ; Molgo, J; Adams, DJ. Botana, L, Ed.; Pharmacology of toxins involved in ciguatera and related fish poisonings. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection; Marcel Dekker, Inc.: New York, 2000; pp. 419–447. [Google Scholar]
- Cameron, J. Effects of ciguatoxin on nerve excitability in rats (part I). J Neurol Sci 1991, 101, 87–92. [Google Scholar]
- Mattei, C; Dechraoui, MY; Molgó, J; Meunier, FA; Legrand, AM; Benoit, E. Neurotoxins targetting receptor site 5 of voltage-dependent sodium channels increase the nodal volume of myelinated axons. J Neurosci Res 1999, 55, 666–673. [Google Scholar]
- Allsop, JL; Martini, L; Lebris, H; Pollard, J; Walsh, J; Hodgkinson, S. Neurologic manifestations of ciguatera. 3 cases with a neurophysiologic study and examination of one nerve biopsy. Rev Neurol 1986, 142, 590–597. [Google Scholar]
- Cameron, J. Electrophysiological studies on ciguatera poisoning in man (part II). J Neurol Sci 1991, 101, 93–97. [Google Scholar]
- Benoit, E. Nodal swelling produced by ciguatoxin-induced selective activation of sodium channels in myelinated nerve fibers. Neuroscience 1996, 71, 1121–1131. [Google Scholar]
- Lehane, L; Lewis, RJ. Ciguatera: recent advances but the risk remains. Int J Food Microbiol 2000, 61, 91–125. [Google Scholar]
- Holmes, MJ; Lewis, RJ. Purification characterization of large and small maitotoxins from cultured Gambierdiscus toxicus. Nat Toxins 1994, 2, 64–72. [Google Scholar]
- Estacion, M. Botana, L, Ed.; Ciguatera toxins: Mechanism of action and pharmacology of maitotoxin. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection; Marcel Dekker, Inc.: New York, 2000; pp. 473–504. [Google Scholar]
- Kakizaki, A; Takahashi, M; Akagi, H; Tachikawa, E; Yamamoto, T; Taira, E; Yamakuni, T; Ohizumi, Y. Ca2+ channel activating action of maitotoxin in cultured brainstem neurons. Eur J Pharmacol 2006, 536, 223–231. [Google Scholar]
- Statake, M; Ofuji, K; Naoki, H; James, KJ; Furey, A; McMahon, T. Azaspiracid, a new marine toxin having unique spiro ring assembles, isolated from Irish mussels, Mytilus edulis. J Am Chem Soc 1998, 120, 9967–9968. [Google Scholar]
- Oufji, K; Statake, M; McMahon, T; Silker, J; James, KJ; Naoki, H. Two analogs of azaspiracid isolated from mussels, Mytilus edulis, involved in human intoxication in Ireland. Nat Toxins 1999, 7, 99–102. [Google Scholar]
- James, K; Lehane, M; Moroney, C; Fernandez-Puente, P; Statake, M; Yasumoto, T. Azaspiracid shellfish poisoning: unusual toxin dynamics in shellfish and the increased risk of acute human intoxications. Food Addit Contam 2002, 19, 555–561. [Google Scholar]
- James, K; Sierra, MD; Lehane, M; Brana Magdalena, A; Furey, A. Detection of five newly hydroxyl analogues of azaspiracids in shellfish using multiple tandem mass spectrometry. Toxicon 2003, 41, 277–283. [Google Scholar]
- Ito, E; Statake, M; Ofuji, K; Kurita, N; McMahon, T; James, K. Multiple organ damage caused by a new toxin azaspiracid, isolated from mussels produced in Ireland. Toxicon 2000, 38, 917–930. [Google Scholar]
- Alfonso, A; Roman, Y; Vieytes, MR. Azaspiracid-4 inhibits Ca2+ entry by stored operated channels in human T lymphocytes. Biochem Pharmacol 2005, 69, 1627–1636. [Google Scholar]
- Suganuma, M; Fujiki, H; Suguri, H; Yoshizawa, S; Hirota, M; Nakayasu, M; Ojika, M; Wakamatsu, K; Yamada, K; Sugimura, T. Specific binding of okadaic acid, a new tumor promoter. Proc Natl Acad Sci USA 1988, 85, 1768–1771. [Google Scholar]
- Ito, E; Statake, M; Ofuji, K; Higashi, M; Harigaya, K; McMahon, T. Chronic effects in mice caused by oral administration of sublethal doses of azaspiracid, a new marine toxin isolated from mussels. Toxicon 2002, 40, 193–202. [Google Scholar]
- Murata, M; Kumagai, M; Lee, JS; Yasumoto, T. Isolation and structure of yessotoxin, a novel polyether compound implicated in diarrheic shellfish poisoning. Tetrahedron Lett 1987, 28, 5869–5872. [Google Scholar]
- Draisci, R; Ferretti, E; Palleschi, L; Marchiafava, C; Poletti, R; Milandri, A; Ceredi, A; Pompei, M. High levels of yessotoxin in mussels and presence of yessotoxin and homoyessotoxin in dinoflagellates of the Adriatic Sea. Toxicon 1999, 37, 1187–1193. [Google Scholar]
- Paz, B; Riobó, P; Luisa Fernández, M; Fraga, S; Franco, JM. Production and release of yessotoxins by the dinoflagellates Protoceratium reticulatum and Lingulodinium polyedrum in culture. Toxicon 2004, 44, 251–258. [Google Scholar]
- Satake, M; Ichimura, T; Sekiguchi, K; Yoshimatsu, S; Oshima, Y. Confirmation of yessotoxin and 45, 46, 47-trinoryessotoxin production by Protoceratium reticulatum collected in Japan. Nat Toxins 1999, 7, 147–150. [Google Scholar]
- Rhodes, L; McNabb, P; de Salas, M; Briggs, L; Beuzenberg, V; Gladstone, M. Yessotoxin production by Gonyaulax spinifera. Harmful Algae 2006, 5, 148–155. [Google Scholar]
- Satake, M; Terasawa, K; Kadowaki, Y; Yasumoto, T. Relative configuration of yessotoxin and isolation of two new analogs from toxic scallops. Tetrahedron Lett 1996, 37, 5955–5958. [Google Scholar]
- Satake, M; Viviani, R; Yasumoto, T. Yessotoxin in mussels of the northern Adriatic Sea. Toxicon 1997, 35, 177–183. [Google Scholar]
- Ciminiello, P; Fattorusso, E; Forino, M; Poletti, R. 42,43,44,45,46,47,55- Heptanor-41-oxohomoyessotoxin, a new biotoxin from mussels of the northern Adriatic sea. Chem Res Toxicol 2001, 14, 596–599. [Google Scholar]
- Ciminiello, P; Dell’Aversano, C; Fattorusso, E; Forino, M; Magno, S; Poletti, R. Direct detection of yessotoxin and its analogues by liquid chromatography coupled with electrospray ion trap mass spectrometry. J Chromatogr A 2002, 968, 61–69. [Google Scholar]
- Tarao, K; Ito, E; Oarada, M; Murata, M; Yasumoto, T. Histopathiological studies on experimental marine toxin poisoning-5. The effects in mice of yessotoxin isolated from Patinopecten yessoessis and a desulfated derivative. Toxicon 1990, 28, 1095–1104. [Google Scholar]
- Aune, T; Sørby, R; Yasumoto, T; Ramstad, H; Landsverk, T. Comparison of oral and intraperitoneal toxicity of yessotoxin towards mice. Toxicon 2002, 40, 77–82. [Google Scholar]
- Tubaro, A; Sosa, S; Carbonatto, M; Altinier, G; Vita, F; Melato, M; Satake, M; Yasumoto, T. Oral and intraperitoneal acute toxicity studies of yessotoxin and homoyessotoxins in mice. Toxicon 2003, 41, 783–792. [Google Scholar]
- Tubaro, A; Sosa, S; Altinier, G; Soranzo, MR; Satake, M; Della Loggia, R; Yasumoto, T. Short-term toxicity of homoyessotoxins, yessotoxin and okadaic acid in mice. Toxicon 2004, 43, 439–445. [Google Scholar]
- Wolf, LW; Laregina, MC; Tolbert, DL. A behavioural study of the development of hereditary cerebellar ataxia in the shaker rat mutant. Behav Brain Res 1996, 75, 67–81. [Google Scholar]
- Franchini, A; Marchesini, E; Poletti, R; Ottaviani, E. Acute toxic effect of the algal yessotoxin on Purkinje cells from the cerebellum of Swiss CDq mice. Toxicon 2004, 43, 347–352. [Google Scholar]
- Perez-Gomez, A; Ferrero-Gutierrez, A; Novelli, A; Franco, JM; Paz, B; Fernandez-Sanchze, MT. Potent neurotoxic action of the shellfish biotoxin yessotoxin on cultured cerebellar neurons. Toxicol Sci 2006, 90, 168–177. [Google Scholar]
- De la Rosa, LA; Alfonso, A; Vieytes, MR; Botana, LM. Modulation of cytosolic calcium levels of human lymphocytes by yessotoxin, a novel marine phycotoxin. Biochem Pharmacol 2001, 61, 827–833. [Google Scholar]
- Moore, RE; Scheuer, PJ. Palytoxin: a new marine toxin from a coelenterate. Science 1971, 172, 495–498. [Google Scholar]
- Penna, A; Vila, M; Fraga, S; Giacobbe, MG; Andreoni, F; Riobó, P; Veronesi, C. Characterization of Ostreopsis and Coolia (Dinophyceae) isolates in the western Mediterranean Sea based on morphology, toxicity, and internal transcribed spacer 5.8S rDNA sequences. J Phycol 2005, 41, 212–225. [Google Scholar]
- Gallitelli, M; Ungaro, N; Addante, LM; Gentiloni, N; Sabbà, C. Respiratory illness as a reaction to tropical algal blooms occurring in a temperate climate. JAMA 2005, 293, 2599–2600. [Google Scholar]
- Sansoni, G; Borghini, B; Camici, G; Casotti, M; Righini, P; Rustighi, C. Fioriture algali di Ostreopsis Ovata (Gonyaulacales: Dinophyceae): Unproblema emergente. Biol Ambientale 2003, 17, 17–23. [Google Scholar]
- Ciminiello, P; Dell’Aversano, C; Fattorusso, E; Forino, M; Magno, GS; Tartaglione, L; Grillo, C; Melchiorre, N. The Genoa 2005 outbreak: Determination of putative palytoxin in Mediterranean Ostreopsis ovata by a new liquid chromatography tandem mass spectrometry method. Anal Chem 2006, 78, 6153–6159. [Google Scholar]
- Riobó, P; Paz, B; Franco, JM. Analysis of palytox-in- like in Ostreopsis cultures by liquid chromatography with precolumn derivatization and fluorescence detection. Anal Chim Acta 2006, 566, 217–223. [Google Scholar]
- Monti, M; Minocci, M; Beran, A; Ivena, L. First record of Ostreopsis cfr. Ovata on macroalgae in the northern Adriatic. Sea, Mar Pol Bull 2007, 54, 598–601. [Google Scholar]
- Alcala, AC; Alcala, LC; Garth, JS; Yasumura, D; Yasumoto, T. Human fatality due to ingestion of the crab Demania reynaudii contained a palytoxin-like toxin. Toxicon 1998, 26, 105–107. [Google Scholar]
- Granéli, E; Ferreira, CEL; Yasumoto, T; Rodrigues, E; Neves, MHB. Sea urchins poisoning by the benthic dinoflagellate Ostreopsis ovata on the Brazilian coast. In Book of Abstracts of Xth International Conference on Harmful Algae; Florida, 2002. [Google Scholar]
- Fukui, M; Murata, M; Inoue, A; Gawel, M; Yasumoto, T. Occurrence of palytoxin in the Trigger fish Melichtys vidua. Toxicon 1987, 25, 1121–1124. [Google Scholar]
- Onuma, Y; Satake, M; Ukena, T; Roux, J; Chanteau, S; Rasolofonirina, N; Ratsimaloto, N; Naoki, H; Yasumoto, T. Identification of putative palytoxin as the cause of clupeotoxism. Toxicon 1999, 37, 55–65. [Google Scholar]
- Taniyama, S; Arakawa, O; Terada, M; Nishio, S; Takatani, T; Mahmud, Y; Noguchi, T. Ostreopsis sp., a possible origin of palytoxin (PTX) in parrotfish Scarus ovifrons. Toxicon 2003, 42, 29–33. [Google Scholar]
- Moore, RE; Bartolini, G; Barchi, J; Bothmer-By, AA; Dadok, J; Ford, J. Absolute stereochemistry of palytoxin. J Am Chem Soc 1982, 104, 3776–3779. [Google Scholar]
- Bottinger, H; Beress, L; Habermann, E. Involvement of (Na+, K+-ATPase) in binding and actions of palytoxin on human erythrocytes. Biochim Biophys Acta 1986, 861, 164–176. [Google Scholar]
- Habermann, E. Palytoxin acts through the Na+, K+-ATPase. Toxicon 1989, 27, 1171–1187. [Google Scholar]
- Kim, SY; Marx, KA; Wu, CH. Involvement of the Na+, K+-ATPase in the introduction of ion channels by palytoxin. Naunyn-Schmiedeberg’s Arch Pharmacol 1995, 351, 542– 554. [Google Scholar]
- Hirsh, JK; Wu, CH. Palytoxin-induced single-channel currents from the sodium pump synthesized by in vitro expression. Toxicon 1997, 35, 169–176. [Google Scholar]
- Scheiner-Bobis, G; Meyer zu Heringdorf, D; Christ, M; Habermann, E. Palytoxin induces K+ efflux from yeast cells expressing the mammalian sodium pump. Mol Pharmacol 1994, 45, 1132– 1136. [Google Scholar]
- Habermann, E; Laux, M. Depolarization increases inositol phosphate production in a particulate preparation from rat brain. Naunyn-Schmiederberg’s Arch Pharmac 1986, 334, l–15. [Google Scholar]
- Levine, L; Fujiki, H. Stimulation of arachidonic acid metabolism by different types of tumor promoters. Carcinogenesis 1985, 6, 1631–1635. [Google Scholar]
- Frelin, C; Vigne, P; Breittmayer, JP. Mechanism of the cardiotoxic action of palytoxin. Mol Pharmacol 1991, 38, 904–909. [Google Scholar]
- Yoshizumi, M; Houchi, H; Ishimura, Y; Masuda, Y; Morita, K; Oka, M. Mechanism of palytoxin induced Na+ influx into cultured bovine adrenal chromaffin cells: possible involvement of Na+/H+ exchange system. Neurosci Left 1991, 130, 103–l 06. [Google Scholar]
- Satoh, E; Nakazato, Y. Mode of action of palytoxin on the release of acetylcholine from rat cerebrocortical synaptosomes. J Neurochem 1991, 57, 1276–1280. [Google Scholar]
- Tsumuraya, T; Fujii, I; Inoue, M; Tatami, A; Miyazaki, K; Hirama, M. Production of monoclonal antibodies for sandwich immunoassay detection of ciguatoxin 51-hydroxy CTX3C. Toxicon 2006, 48, 287–294. [Google Scholar]
- Ishikawa, Y; Nishiyama, S. Synthesis of the BCD ring system of azaspiracid: construction of the trispiro ring structure by the thioether approach. Tetrahedron Lett 2004, 45, 351–354. [Google Scholar]
- Miles, CO; Samdal, IA; Aasen, JAG; Jensen, DJ; Quilliam, MA; Petersen, D; Briggs, LM; Wilkins, AL; Rise, F; Cooney, JM; MacKenzie, AL. Evidence for numerous analogs of yessotoxin in Protoceratium reticulatum. Harmful Algae 2005, 6, 1075–1091. [Google Scholar]
- Cha, JK; Christ, WJ; Finan, JM; Fujiko, H; Kishi, Y; Klein, LL; Ko, SS; Leder, J; McWhorter, WW, Jr; Pfaff, KP; Yonaga, M; Uemura, D; Hirata, Y. Stereochemistry of palytoxin. Complete structure. J Am Chem Soc 1982, 104, 7369–7371. [Google Scholar]
Figure 1.
The voltage-gated channels: The different members of the ion channel family structurally related to the voltage-gated ion channels are illustrated as transmembrane folding diagrams in which cylinders represent probable transmembrane alpha helices. Green, S5–S6 pore forming segments; red, S4 voltage sensor; and gray, S1–S3 tansemembrane segments [13].
Figure 1.
The voltage-gated channels: The different members of the ion channel family structurally related to the voltage-gated ion channels are illustrated as transmembrane folding diagrams in which cylinders represent probable transmembrane alpha helices. Green, S5–S6 pore forming segments; red, S4 voltage sensor; and gray, S1–S3 tansemembrane segments [13].
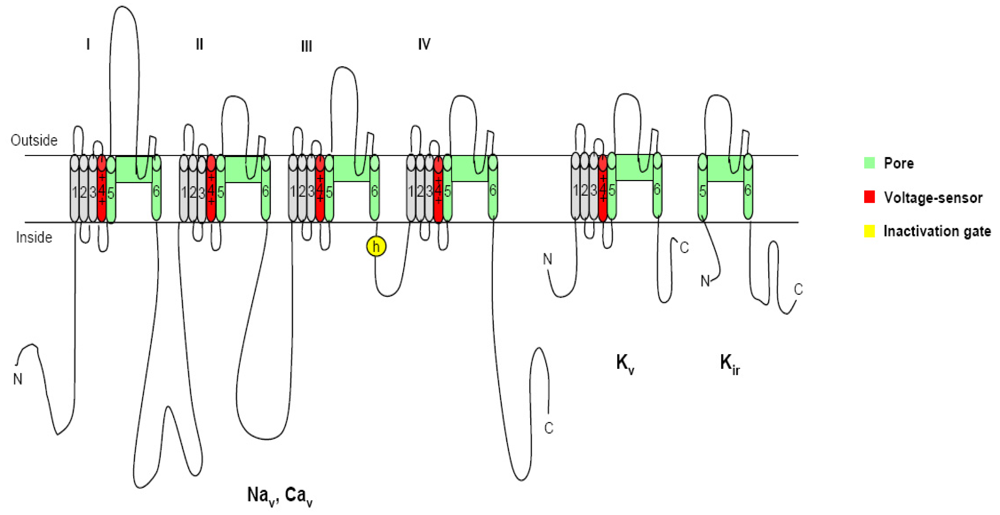
Figure 2.
Structure and species of paralytic shellfish poisoning toxins from marine dinoflagellates [4].
Figure 2.
Structure and species of paralytic shellfish poisoning toxins from marine dinoflagellates [4].
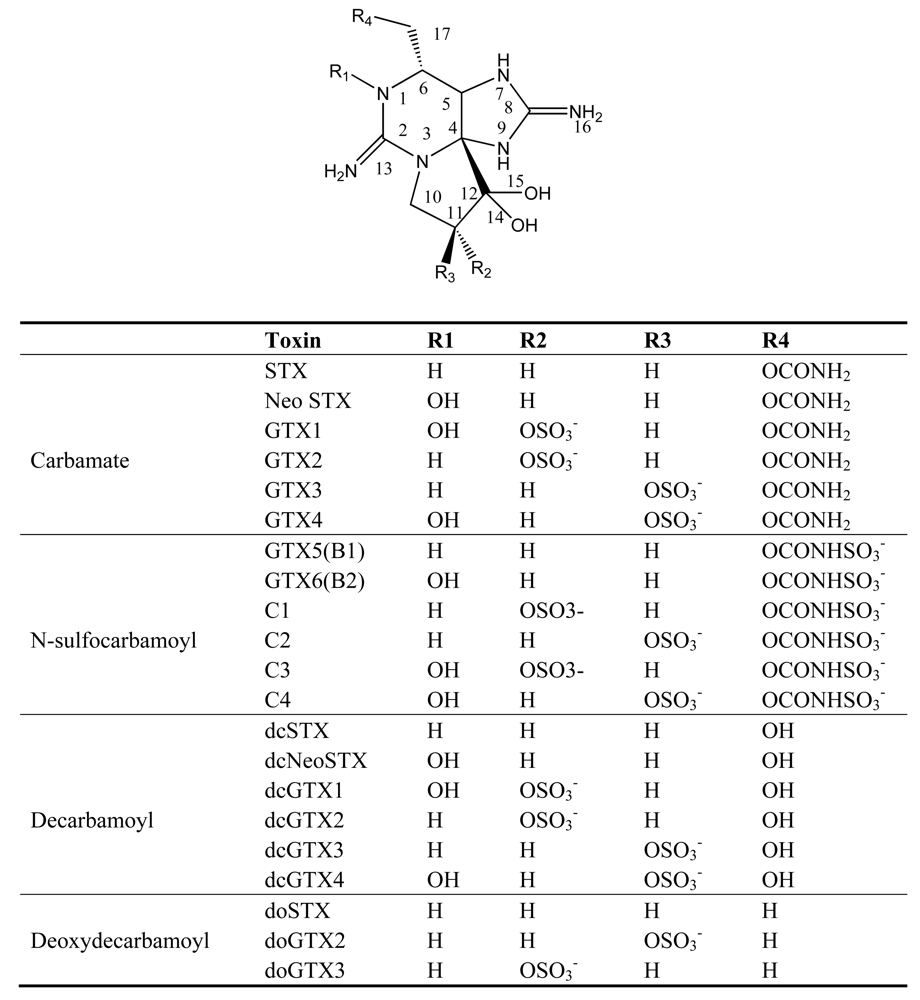
Figure 3.
Structure and species of neurotoxic shellfish poisoning toxins from marine dinoflagellates [4].
Figure 3.
Structure and species of neurotoxic shellfish poisoning toxins from marine dinoflagellates [4].
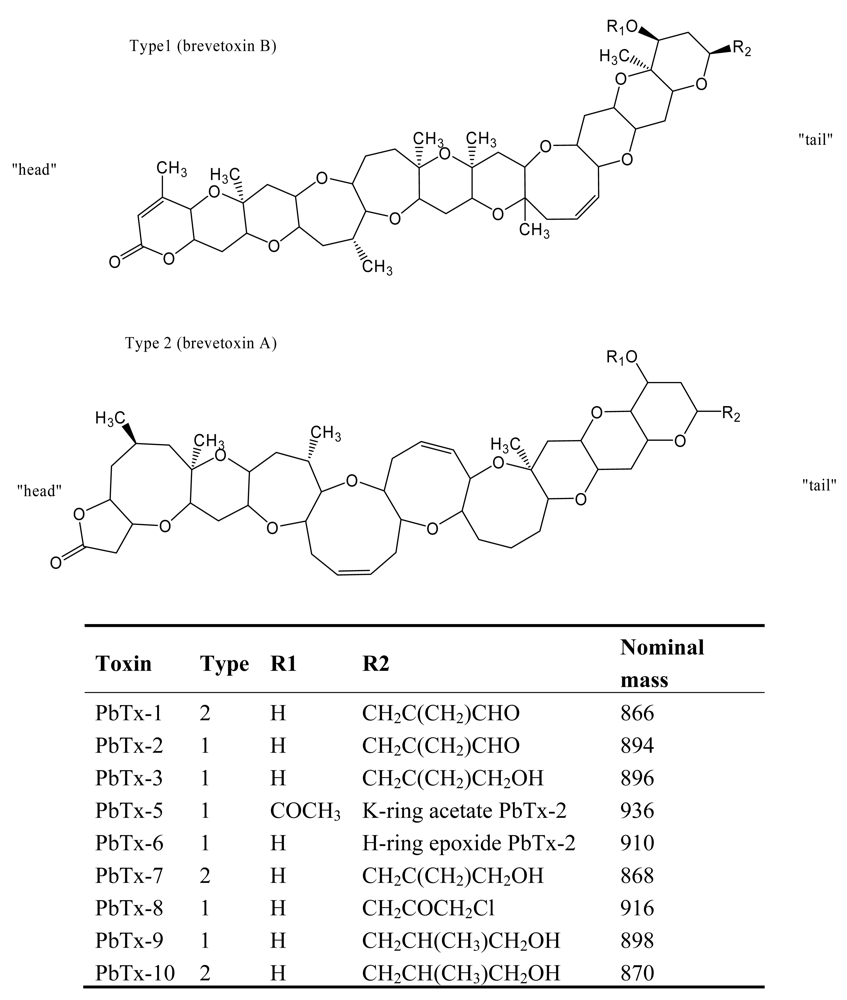
Figure 4.
Structure and species of ciguatoxins from the dinoflagellate G. toxicus [62].
Figure 4.
Structure and species of ciguatoxins from the dinoflagellate G. toxicus [62].
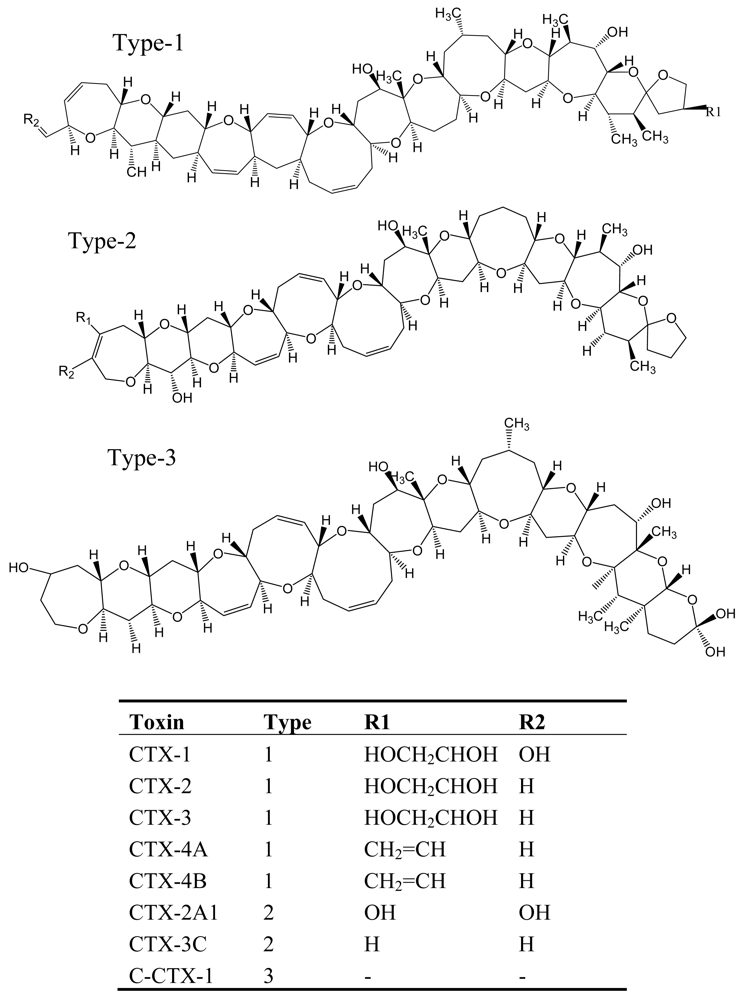
Figure 5.
Structure of maitotoxin from the dinoflagellate G. toxicus [120].
Figure 5.
Structure of maitotoxin from the dinoflagellate G. toxicus [120].
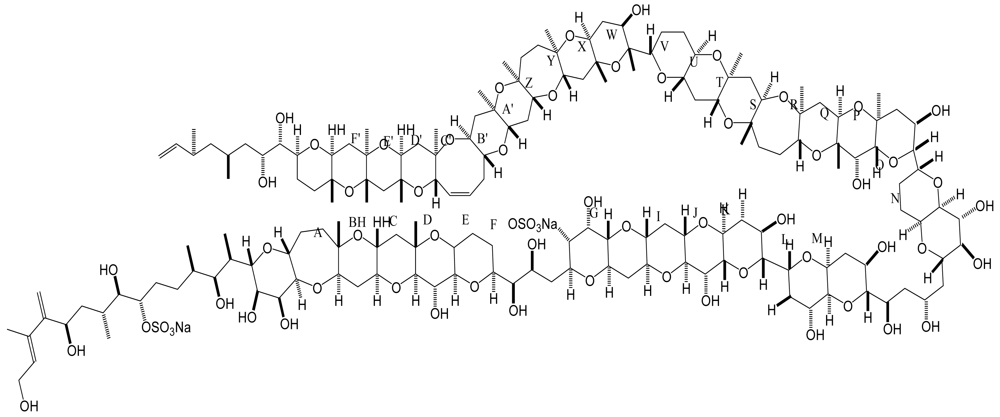
Figure 6.
Structure and species of azaspiracid poisoning toxins from marine dinoflagellates [121].
Figure 6.
Structure and species of azaspiracid poisoning toxins from marine dinoflagellates [121].
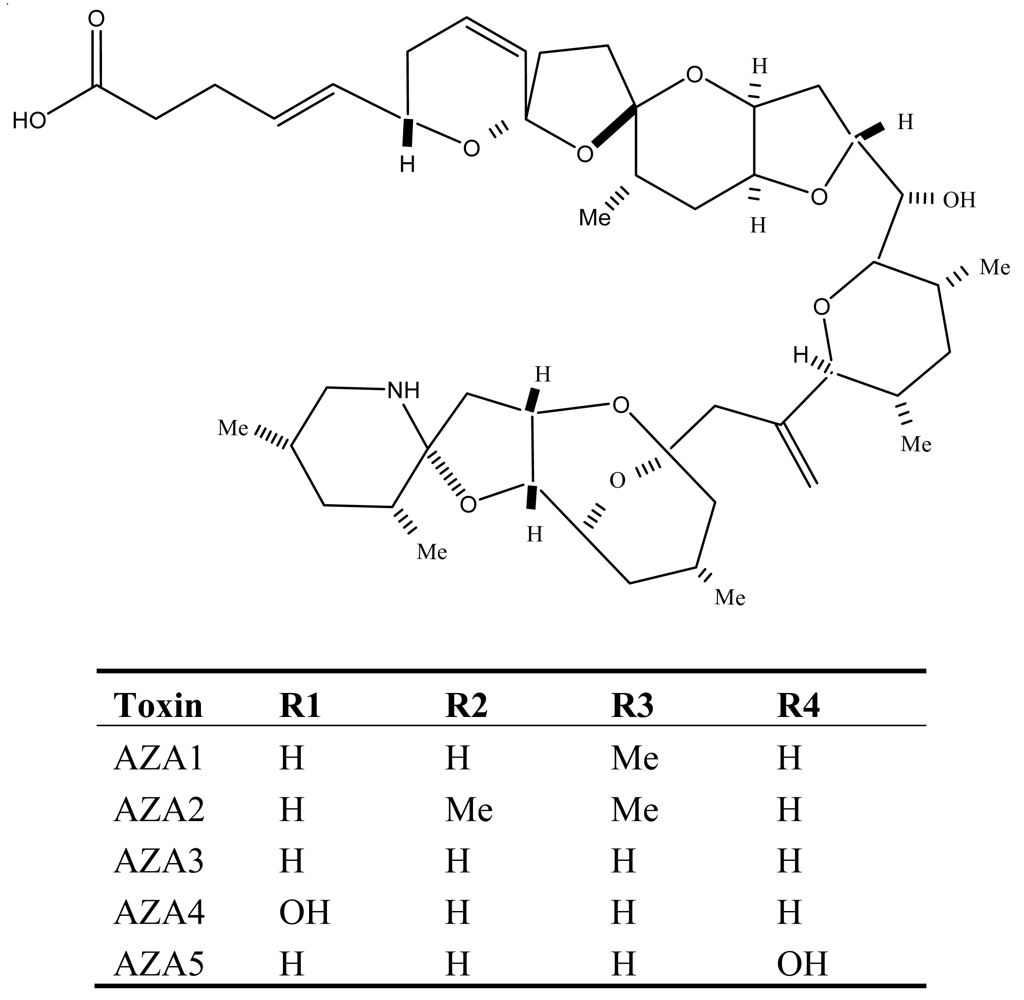
Figure 7.
Structure and species of yessotoxins from marine dinoflagellates [122].
Figure 7.
Structure and species of yessotoxins from marine dinoflagellates [122].
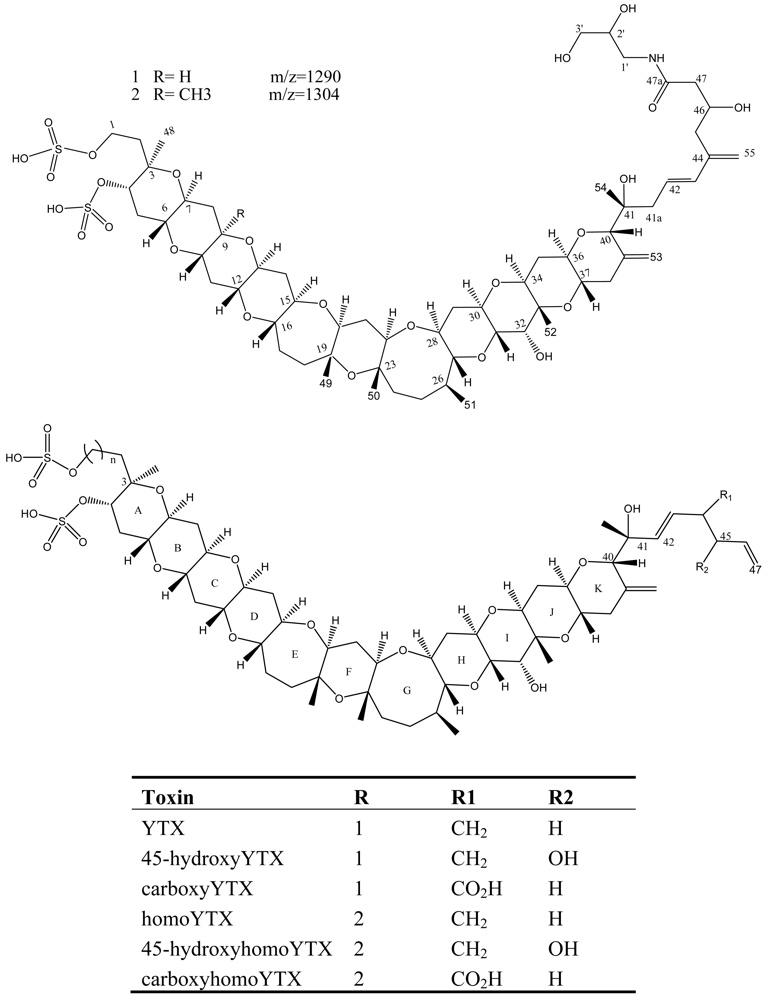
Figure 8.
Structure of palytoxin from marine dinoflagellates [123].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of poisoning | Toxins | Sources of toxins | Primary vector | Action target | Ref. |
|---|---|---|---|---|---|
| PSP | Saxitoxins and gonyautoxins | Alexandrium spp., Gymnodinium spp., Pyrodinium spp. | Shellfish | Voltage-gated sodium channel 1 | 23, 27–29 |
| NSP | Brevetoxins | Kerenia brevis, Chatonella marina, C. antiqua, Fibrocapsa japonica, Heterosigma akashiwo | Shellfish | Voltage-gated sodium channel 5 | 33, 39–41, 46, 52 |
| Yessotoxins | Protoceratium reticulatum, Lingulodinium polyedrum Gonyaulax spinifera | Shellfish | Voltage-gated calcium/sodium channel? | 94–95 | |
| CFP | Ciguatoxins | Gambierdiscus toxicus | Coral reef fish | Voltage-gated sodium channel 5 | 55, 62, 63 |
| CFP | Maitotoxins | Gambierdiscus toxicus | Coral reef fish | Voltage-gated calcium channel | 69, 70 |
| AZP | Azaspiracids | Protoperidinium crassipes | Shellfish | Voltage-gated calcium channel | 72, 76 |
| Palytoxin poisoning | Palytoxins | Ostrepsis siamensis | Shellfish | Na+-K+ ATPase | 97, 109–111 |
Notes: PSP, paralytic shellfish poisoning; NSP, neurotoxic shellfish poisoning; CFP, ciguatera fish poisoning, AZP, azaspiracid poisoning.
Share and Cite
MDPI and ACS Style
Wang, D.-Z. Neurotoxins from Marine Dinoflagellates: A Brief Review. Mar. Drugs 2008, 6, 349-371. https://doi.org/10.3390/md6020349
AMA Style
Wang D-Z. Neurotoxins from Marine Dinoflagellates: A Brief Review. Marine Drugs. 2008; 6(2):349-371. https://doi.org/10.3390/md6020349
Chicago/Turabian StyleWang, Da-Zhi. 2008. "Neurotoxins from Marine Dinoflagellates: A Brief Review" Marine Drugs 6, no. 2: 349-371. https://doi.org/10.3390/md6020349