Non-Traditional Vectors for Paralytic Shellfish Poisoning

Abstract
:1. Paralytic Shellfish Toxins and Sources
2. Traditional Vectors of Saxitoxins to Human Consumers
3. Non-Traditional Vectors of Saxitoxins to Human Consumers
3.1 Gastropods
3.1.1 Case Study 1: STXs in Abalone
Spain
South Africa
3.1.2 Case Study 2: STXs in Whelks and Moon Snails
Japan
New England, USA
3.2 Crustaceans
3.2.1 Case Study 3: STXs in Crabs
East Timor
Japan
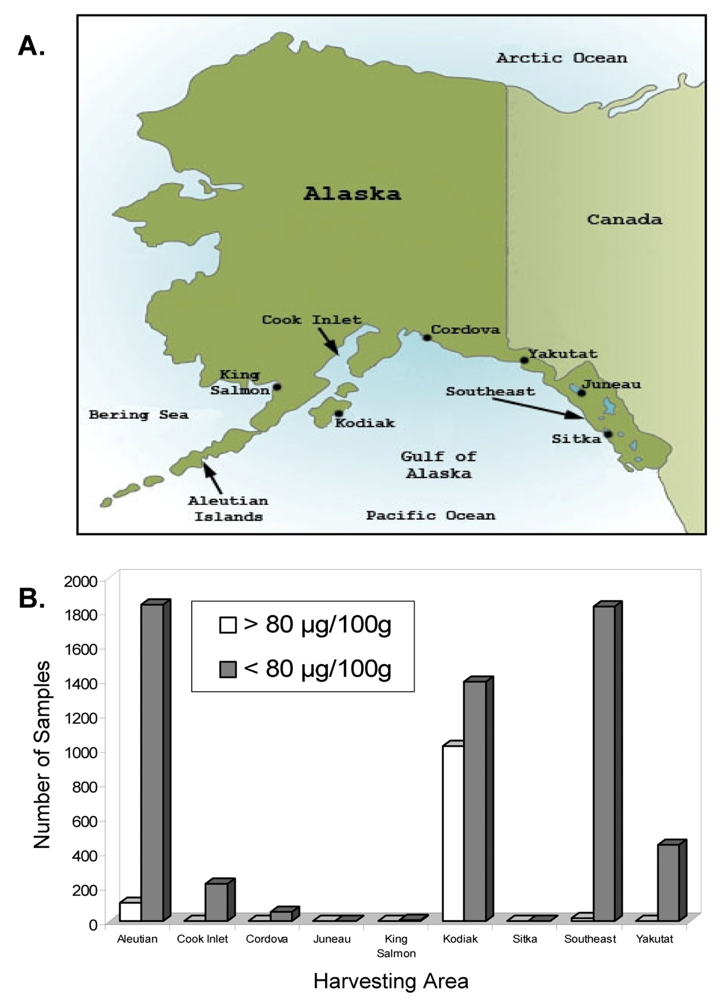
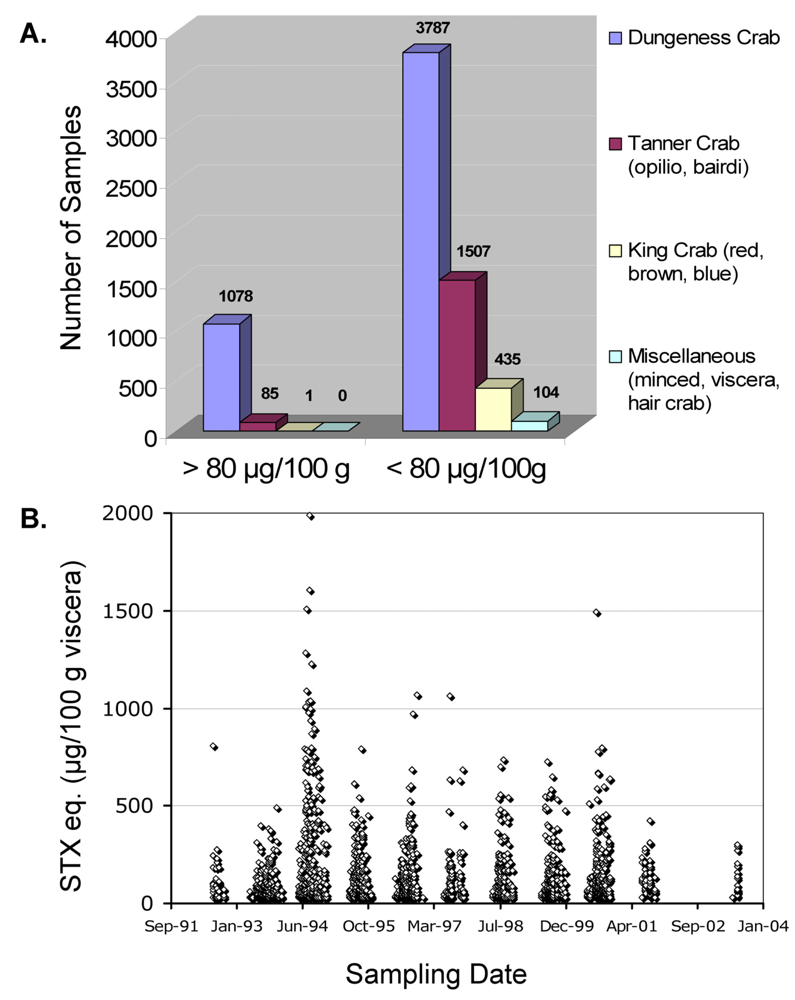
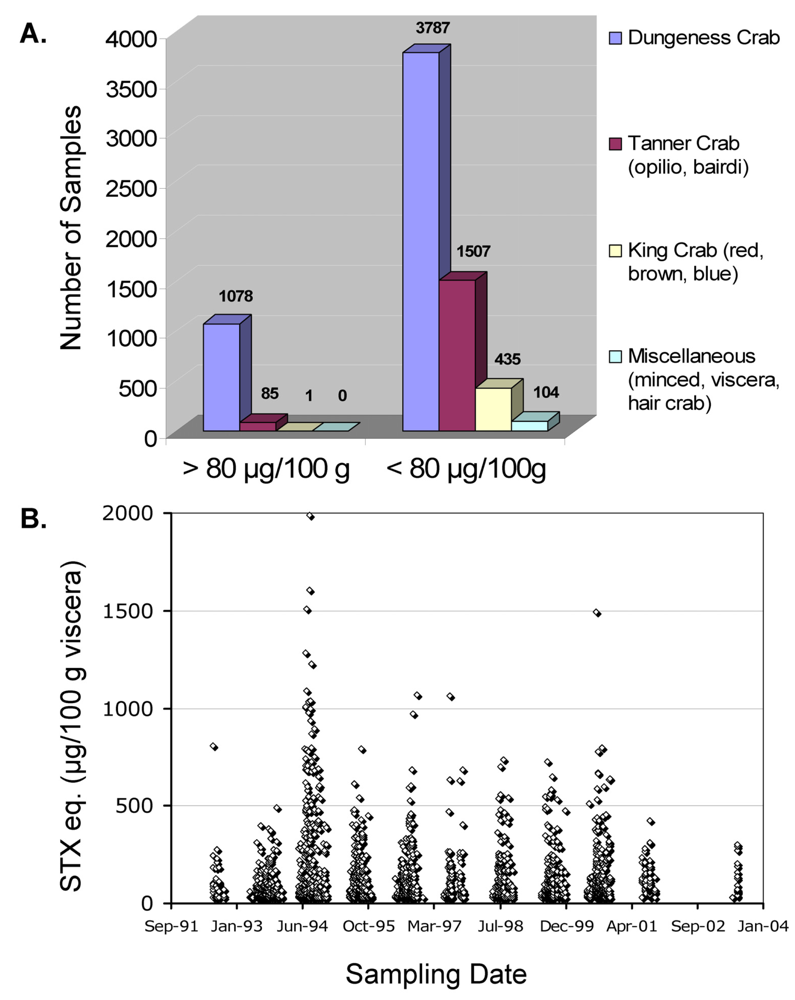
Alaska, USA
3.3 Other invertebrates
3.4 Fish
3.4.1 Case study 4: STXs in planktivorous fish
Far East
3.4.2 Case study 5: STXs in puffer fish
Florida, USA
4. Conclusion
- 1Steidinger et al. (1980) [49] distinguished P. bahamense var. compressa from P. bahamense var. bahamense based on morphological, dimensional, and toxicological characteristics. P. bahamense var. compressa was reported to be the toxic variety responsible for Indo-Pacific PSP events while P. bahamense var. bahamense was reported to be non-toxic. Landsberg et al. (2006) [58] reported for the first time STX(s) production from P. bahamense in Florida, USA putatively responsible for several cases of SPFP. Badylak et al. (2004) [57] confirmed that the P. bahamense variety occurring in Florida waters still conformed to Steidinger et al. (1980) [49] description of P. bahamense var. bahamense except that it now appeared to be toxic. The description of P. bahamense var. compressa vs. P. bahamense var. bahamense is currently being re-evaluated (K. Steidinger, personal communication), therefore throughout the text P. bahamense without varietal designation is used.
- Samples Availability: Available from the authors.
References
- Oshima, Y. Lassus, P, Arzul, G, Erard-Le-Denn, E, Gentien, P, Marcaillou-Le-Baut, C, Eds.; Chemical and enzymatic transformation of paralytic shellfish toxins in marine organisms. In Harmful Marine Algal Blooms; Technique et Documentation, Lavoisier: Paris, 1995; pp. 475–480. [Google Scholar]
- Cembella, AD; Shumway, SE; Lewis, NI. Anatomical distribution and spatio-temporal variation in paralytic shellfish toxin composition in two bivalve species from the Gulf of Maine. J Shellfish Res 1993, 12, 389–403. [Google Scholar]
- Onodera, HY; Oshima, Y; Watanabe, MF; Watanabe, M; Bolch, CJ; Blackburn, S; Yasumoto, T. Yasumoto, T, Oshima, Y, Fukuyo, Y, Eds.; Screening of paralytic shellfish toxins in freshwater cyanobacteria and chemical confirmation of the toxins in cultured Anabaena circinalis from Australia. In Harmful and Toxin Algal Blooms; Intergovernmental Oceanographic Commission of UNESCO: Paris, 1996; pp. 563–566. [Google Scholar]
- Sullivan, JJ. Falconer, IR, Ed.; Methods of analysis for algal toxins: dinoflagellate and diatom toxins. In Algal Toxins in Seafood and Drinking Water; Academic Press: London, 1993; pp. 29–48. [Google Scholar]
- Onoue, Y; Nozawa, K. Natori, S, Hashimoto, K, Ueno, Y, Eds.; Zinc-bound PSP toxins separated from Cochlodinium red tides. In Mycotoxins and Phycotoxins ’88; Elsevier: Amsterdam, 1989; pp. 359–366. [Google Scholar]
- Reyes-Vasquez, G; Ferraz-Reyes, E; Vasquez, E. Taylor, DL, Seliger, HH, Eds.; Toxic dinoflagellate blooms in northeastern Venezuela during 1977. In Toxic Dinoflagellate Blooms; Elsevier: New York, 1979; pp. 191–194. [Google Scholar]
- Prakash, A; Taylor, FJR. A red water bloom of Gonyaulax acatenella in the Straight of Georgia and its relation to paralytic shellfish toxicity. J Fish Res Bd Can 1966, 23, 1625–1270. [Google Scholar]
- Schmidt, RJ; Loeblich, AR, III. Distribution of paralytic shellfish poisoning among Pyrrhophyta. J Mar Biol Assoc UK 1979, 59, 479–487. [Google Scholar]
- Cembella, AD; Sullivan, JJ; Boyer, GL; Taylor, FJR; Anderson, RJ. Variation in paralytic shellfish toxin composition within the Protogonyaulax tamarensis/catenella species complex; red tide dinoflagellates. Biochem Syst Ecol 1987, 15, 171–186. [Google Scholar]
- Ciminiello, PE; Fatturoso, E; Fornio, M; Montresor, M. A new PSP-like toxin in Alexandrium andersoni (Dinophyceae). Harmful Algae News 1999, 18, 1–3. [Google Scholar]
- Ciminiello, PE; Fatturoso, E; Fornio, M; Montresor, M. Saxitoxin and neosaxitoxin as toxic principles of Alexandrium andersoni (Dinophyceae) from the Gulf of Naples, Italy. Toxicon 2000, 38, 1871–1877. [Google Scholar]
- Taylor, FJR; Fukuyo, Y; Larsen, J. Hallegraeff, GM, Anderson, DM, Cembella, AD, Eds.; Taxonomy of harmful dinoflagellates. In Manual on Harmful Marine Microalgae; UNESCO: Paris, 1995; pp. 283–317. [Google Scholar]
- Schantz, EJ; Lynch, JM; Vayvada, G; Matsumoto, K; Rapoport, H. The purification and characterization of the poison produced by Gonaulax catenella in axenic culture. Biochemistry 1966, 5, 1191–1195. [Google Scholar]
- Proctor, NH; Chan, SL; Trevor, AJ. Production of saxitoxin by cultures of Gonyaulax catenella. Toxicon 1975, 13, 1–9. [Google Scholar]
- Onone, Y; Noguchi, T; Hashimoto, K. Studies on paralytic shellfish poison from the oyster cultured in Senzaki Bay, Yamaguchi Prefecture. Nippon Suis Gakk 1980, 46, 1031–1034. [Google Scholar]
- Onone, Y; Noguchi, T; Maruyama, J; Hashimoto, K; Ikeda, T. New toxins separated from oysters and Protogonyaulax catenella from Senzaki Bay, Yamaguchi Prefecture. Nippon Suis Gakk 1981, 47, 1643. [Google Scholar]
- Onone, Y; Noguchi, T; Maruyama, J; Uneda, Y; Hashimoto, K; Ikeda, T. Comparison of PSP compositions between toxic oysters and Protogonyaulax catenella from Senzaki Bat, Yamaguchi Prefecture. Nippon Suis Gakk 1981, 47, 1347–1350. [Google Scholar]
- Boyer, GL; Sullivan, JJ; Anderson, RJ; Harrison, PJ; Taylor, FRJ. Anderson, DM, White, AW, Baden, DG, Eds.; Toxin production in three isolates of Protogonyaulax sp. In Toxic Dinoflagellates; Elsevier: New York, 1985; pp. 281–286. [Google Scholar]
- Ogata, T; Kodama, M. Ichthyotoxicity found in cultured media of Protogonyaulax spp. Mar Biol 1986, 92, 31–34. [Google Scholar]
- Hallegraeff, GM; Bolch, CJ; Blackburn, SI; Oshima, Y. Species of the toxigenic dinoflagellate genus Alexandrium in southeastern Australian waters. Bot Mar 1991, 34, 575–587. [Google Scholar]
- Kim, C-H; Sako, Y; Ishida, Y. Comparison of toxin composition between populations of Alexandrium spp. from geographically distinct areas. Nippon Suisan Gakkaishi 1993, 59, 641–646. [Google Scholar]
- Kodama, M; Ogata, T; Fukuyo, Y; Ishimaru, T; Pholpunthin, P; Wisessang, S; Saitanu, K; Panickyakarn, V; Piyankarnchana, T. Protogonyaulax cohorticula, a toxic dinoflagellate found in the Gulf of Thailand. Toxicon 1988, 26, 709–712. [Google Scholar]
- Fukuyo, Y; Yoshida, K; Ogata, T; Ishimaru, T; Kodama, M; Pholpunthin, P; Wisessang, S; Phanichyakarn, V; Piyakarnchana, T. Okaichi, T, Anderson, DM, Nemoto, T, Eds.; Suspected causative dinoflagellates of paralytic shellfish poisoning in the Gulf of Thailand. In Red Tides, Biology, Environmental Science, and Toxicology; Elsevier: New York, 1989; pp. 403–406. [Google Scholar]
- White, AW; Maranda, L. Paralytic shellfish toxins in the dinoflagellate Gonyaulax excavate and in shellfish. J Fish Res Bd Can 1978, 35, 397–402. [Google Scholar]
- Anderson, DM; Kulis, DM; Sullivan, JJ; Hall, S. Toxin composition in one isolate of the dinoflagellate Alexandrium fundyense. Toxicon 1990, 28, 885–893. [Google Scholar]
- Hallegreaff, GM; Steffensen, DA; Wetherbee, R. Three estuarine Australian dinoflagellates that can produce paralytic shellfish toxins. J Plankton Res 1988, 10, 533–541. [Google Scholar]
- Oshima, Y; Hirota, M; Yasumoto, T; Hallegraeff, GM; Blackburn, SI; Steffensen, DA. Production of paralytic shellfish toxins by the dinoflagellate Alexandrium minutum Halim from Australia. Bull Jap Soc Sci Fish 1989, 55, 925. [Google Scholar]
- Franco, JM; Fernandez, P; Reguera, B. Toxin profiles of natural populations and cultures of Alexandrium minutum Halim and Galician (Spain) coastal waters. J Appl Phycol 1994, 6, 275–279. [Google Scholar]
- Mascarenhas, I; Alvito, P; Franca, S; Sousa, I; Martinez, AG; Rodriguez Vasquez, JA. Lassus, P, Arzul, G, Erard-Le-Dean, E, Gentein, P, Marcaillou-Le-Baut, C, Eds.; The dinoflagellate Alexandrium lusitanicum isolated from the coast of Portugal: observations on toxicity and ultrastructure during growth phases. In Harmful Marine Algal Blooms; Lavoisier: Paris, 1995; pp. 71–76. [Google Scholar]
- Lush, GJ; Halegraeff, GM. Yasumoto, T, Oshima, Y, Fukuyo, Y, Eds.; High toxicity of the red tide dinoflagellate Alexandrium minutum to the brine shrimp Artemia salina. In Harmful and Toxic Algal Blooms; International Intergovernmental Oceanographic Commission of UNESCO: Paris, 1996; pp. 389–392. [Google Scholar]
- Hansen, PJ; Cembella, AD; Moestrup, Ø. The marine dinoflagellate Alexandrium ostenfeldii: paralytic shellfish toxin concentration, composition, and toxicity to a tintinnid ciliate. J Phycol 1992, 28, 597–603. [Google Scholar]
- Mackenzie, L; White, D; Oshima, Y; Kapa, J. The resting cyst and toxicity of Alexandrium ostenfeldii. Phycologia 1996, 35, 1348–155. [Google Scholar]
- Cembella, AD; Quilliam, MA; Lewis, NI; Bauder, AG; Wright, JLC. Reguera, B, Blanco, J, Fernández, MI, Wyatt, T, Eds.; Identifying the planktonic origin and distribution of spirolodes in coastal Nova Scotian waters. In Harmful Algae; Xunta de Galicia and IOC: Galicia and Paris, 1998; pp. 481–484. [Google Scholar]
- Cembella, AD; Lewis, NI; Quilliam, MA. The marine dinoflagellate Alexandrium ostenfeldii (Dinophyceae) as the causative organism of spirolide shellfish toxins. Phycologia 2000, 39, 67–74. [Google Scholar]
- Prakash, A. Growth and toxicity of a marine dinoflagellate, Gonyaulax tamarensis. J Fish Res Bd Can 1967, 24, 1589–1606. [Google Scholar]
- Oshima, Y; Yasumoto, T. Taylor, DL, Seliger, E, Eds.; Analysis of toxins in cultured Gonyaulax excavata. In Toxic Dinoflagellate Blooms; Elsevier: New York, 1979; pp. 377–380. [Google Scholar]
- Schantz, EJ. Ragelis, EP, Ed.; Historical perspective on paralytical shellfish poisoning. In Seafood Toxins; American Chemical Society Symposium Series: Washington D.C., 1984; pp. 99–111. [Google Scholar]
- Ogata, T; Kodama, M; Ishimaru, T. Toxin production in the dinoflagellate Protogonyaulax tamarensis. Toxicon 1987, 25, 923–928. [Google Scholar]
- Cembella, AD; Therriault, J-C; Beland, P. Toxicity of cultured isolates and natural populations of Protogonyaulax tamarensis from the St. Lawrence Estuary. J Shellfish Res 1988, 7, 611–621. [Google Scholar]
- Lee, JS; Jeon, J-K; Han, M-S; Oshima, Y; Yasumoto, T. Paralytic shellfish toxins in the mussel Mytilus edulis and dinoflagellate Alexandrium tamarense from Jinhae Bay, Korea. Bull Korean Fish Soc 1992, 25, 144–150. [Google Scholar]
- Oshima, Y; Sugino, K; Itakura, H; Hirota, M; Yasumoto, T. Graneli, E, Sundstroem, B, Edler, L, Anderson, DM, Eds.; Comparative studies on paralytic shellfish toxin profile of dinoflagellates and bivalves. In Toxic Marine Phytoplankton; Elsevier Press: New York, 1990; pp. 391–396. [Google Scholar]
- Wisessang, S; Ogata, T; Kodama, M; Fukuyo, Y; Ishimaru, T; Saitanu, K; Yongvanich, T. Piyakarnchana. Accumulation of paralytic shellfish toxins by green mussel Perna viridis by feeding on cultured cells of Alexandrium cohorticula isolated from the Gulf of Thailand. Nippon Suis Gakk 1991, 57, 127–131. [Google Scholar]
- Oshima, Y; Yasumoto, T; Hallegraeff, G; Blackburn, S. Gopalakrishnakone, P, Tans, CK, Eds.; Paralytic shellfish toxins and causative organisms in the tropical Pacific and Tasmanian waters. In Progress in Venom and Toxin Research; National University of Singapore: Singapore, 1987; pp. 423–428. [Google Scholar]
- Oshima, Y; Blackburn, SI; Hallegraeff, GM. Comparative study on paralytic shellfish toxin profiles of the dinoflagellate Gymnodinium catenatum from three different countries. Mar Biol 1993, 116, 471–476. [Google Scholar]
- Oshima, Y; Itakura, H; Lee, K-C; Yasumoto, T; Blackburn, S; Hallegraeff, G. Smadya, TJ, Shimizu, Y, Eds.; Toxin production by the dinoflagellate Gymnodinium catenatum. In Toxic Phytoplankton Blooms in the Sea; Elsevier: Amsterdam, 1993; pp. 907–912. [Google Scholar]
- Anderson, DM; Sullivan, JJ; Reguera, B. Paralytic shellfish poisoning in north-west Spain: the toxicity of the dinoflagellate Gymnodinium catanatum. Toxicon 1989, 27, 665–674. [Google Scholar]
- Maclean, JL. Red tide and paralytic shellfish poisoning in Papua New Guinea. Papua New Guinea Agric J 1973, 24, 131–138. [Google Scholar]
- Maclean, JL. Taylor, DL, Seliger, SS, Eds.; Indo-Pacific red tides. In Toxic Dinoflagellate Blooms; Elsevier: New York, 1979; pp. 173–178. [Google Scholar]
- Steidinger, KA; Tester, LS; Taylor, FJR. A redescription of Pyrodinium bahamense var. compressa (Böhm) stat. nov. from Pacific red tides. Phycologia 1980, 19, 329–334. [Google Scholar]
- Harada, T; Oshima, Y; Kamiya, H; Yasumoto, T. Confirmation of paralytic shellfish toxins in the dinoflagellate Pyrodinium bahamense var. compressa and bivalves in Palau. Nippon Suis Gakk 1982, 48, 821–825. [Google Scholar]
- Harada, T; Oshima, Y; Yasumoto, T. Structures of two paralytic shellfish toxins, gonyautoxins V and VI, isolated from a tropical dinoflagellate, Pyrodinium bahamense var. compressa. Agric Biol Chem 1983, 46, 1861–1864. [Google Scholar]
- Oshima, Y; Kotaki, Y; Harada, T; Yasumoto, T. Ragelis, EP, Ed.; Paralytic shellfish toxins in tropical waters. In Seafood Toxins; American Chemical Society Symposium Series: Washington D.C., 1984; pp. 161–170. [Google Scholar]
- Rosales-Loessener, F; de Porras, E; Dix, MW. Okaichi, T, Anderson, DM, Nemoto, T, Eds.; Toxic shellfish poisoning in Guatemala. In Red Tides, Biology, Environmental Science and Toxicology; Elsevier: New York, 1989; pp. 113–116. [Google Scholar]
- Usup, G; Kulis, DM; Anderson, DM. Growth and toxin production of the toxic dinoflagellate Pyrodinium bahamense var. compressum in laboratory cultures. Nat Toxins 1994, 2, 35–43. [Google Scholar]
- Usup, G; Kulis, DM; Anderson, DM. Lassus, P, Arzul, G, Erard-Le-Denn, E, Gentein, P, Marciallou-Le-Baut, C, Eds.; Toxin production in a Malaysian isolate of the toxic dinoflagellate Pyrodinium bahamense var. compressum. In Harmful Marine Algal Blooms; Lavoisier: Paris, 1995; pp. 519–524. [Google Scholar]
- Orellana-Capeda, E; Martinez-Romero, E; Munoz-Cabrera, L; Lopez-Ramirez, P; Cabrera-Mancilla, E; Ramirez-Camarena, C. Reguera, B, Blanco, J, Fernandez, ML, Wyatt, T, Eds.; 1998; Toxicity associated with blooms of Pyrodinium bahamense var. compressum in southwestern Mexico. In Harmful Algae; Xunta de Galicia and IOC: Galicia and Paris, 1988; p. 60.
- Badylak, S; Kelley, K; Phlips, EJ. A description of Pyrodinium bahamense (Dinophyceae) from the Indian River Lagoon, Florida, USA. Phycologia 2004, 43, 653–657. [Google Scholar]
- Landsberg, JH; Hall, S; Johannessen, JN; White, KD; Conrad, SM; Abbott, JP; Flewelling, LJ; Richardson, RW; Dickey, RW; Jester, ELE; Etheridge, SM; Deeds, JR; Van Dolah, FM; Leighfield, TA; Zou, Y; Beaudry, CG; Benner, RA; Rogers, PL; Scott, PS; Kawabata, K; Wolny, JL; Steidinger, KA. Saxitoxin puffer fish poisoning in the United States, with the first report of Pyrodinium bahamense as the putative toxin source. Environ Health Perspect 2006, 114, 1502–1507. [Google Scholar]
- May, V; McBarron, EJ. Occurrence of the blue-green alga, Anabaena circinalis Rabenh, in New South Wales and toxicity to mice and honey bees. J Aust Inst Agric Sci 1973, 39, 264–266. [Google Scholar]
- McBarron, EJ; Walker, RI; Gardner, I; Walker, KH. Toxicity of livestock to the blue-green alga Anabaena circinalis. Aust Vet J 1975, 51, 587–588. [Google Scholar]
- Sivonen, K; Himberg, K; Luukkainen, R; Niemela, SI; Poon, GK; Codd, GA. Preliminary characterization of neurotoxic cyanobacterial blooms and strains from Finland. Tox Assess Int J 1989, 4, 339–352. [Google Scholar]
- Kiviranta, J; Sivonen, K; Niemala, SI; Huovinen, K. Detection of toxicity of cyanobacteria by Artenia salina bioassay. Environ Toxicol Water Qual 1991, 6, 423–436. [Google Scholar]
- Humpage, AR; Rositano, J; Breitag, AH; Brown, R; Baler, PD; Nicholson, BC; Steffensen, DA. Paralytic shellfish poisons from Australian cyanobacterial blooms. Aust J Mar Freshwat Res 1994, 45, 476–471. [Google Scholar]
- Negri, AP; Jones, GJ. Bioaccumulation of paralytic shellfish poisoning (PSP) toxins from the cyanobacterium Anabaena circinalis by the freshwater mussel Alathyria condola. Toxicon 1995, 33, 667–668. [Google Scholar]
- Negri, AP; Jones, GJ; Blackburn, S. Reguera, B, Blanco, J, Fernandez, ML, Wyatt, T, Eds.; Paralytic shellfish toxins in Australian strains of the freshwater cyanobacteria Anabaena circinalis. In Harmful Algae; Xunta de Galicia and IOC: Galicia and Paris, 1998; pp. 345–348. [Google Scholar]
- Negri, AP; Jones, GJ; Hindmarch, M. Sheep mortality associated with paralytic shellfish poisoning toxins from the cyanobacterial Anabaena circinalis. Toxicon 1995, 33, 1321–1329. [Google Scholar]
- Rapala, J; Robertson, A; Negri, AP; Berg, KA; Tuomi, P; Lyra, C; Erkomaa, K; Lahti, K; Hoppu, K; Lepistö, L. First report of saxitoxin in Finnish lakes and possible associated effects on human health. Environ Toxicol 2005, 20, 331–340. [Google Scholar]
- Pereira, P; Li, RH; Carmichael, WW; Dias, E; Franca, S. Taxonomy and production of paralytic shellfish toxins by the freshwater cyanobacterium Aphanizomenon gracile LMECYA40. Eur J Phycol 2004, 39, 361–368. [Google Scholar]
- Jackim, E; Gentile, J. Toxins of a blue-green alga: similarity to saxitoxin. Science 1968, 162, 915–916. [Google Scholar]
- Sawyer, PJ; Gentile, JH; Sasner, JJ, Jr. Demonstration of a toxin from Aphanizomenon flosaquae (L.). Ralfs. Can J Microbiol 1968, 14, 1199–1204. [Google Scholar]
- Gentile, JH; Maloney, TE. Toxicity and environmental requirements of a strain of Aphanizomenon flosaquae (L.) Ralfs. Can J Microbiol 1969, 15, 165–173. [Google Scholar]
- Alam, M; Shimidzu, Y; Iwaka, M; Sasner, JJ. Reinvestigation of the toxins from the blue-green algae, Aphanizomenon flosaquae, by a high performance chromatographic method. J Environ Sci Hlth A 1978, 13, 493–499. [Google Scholar]
- Ikawa, M; Wegener, K; Foxall, TL; Sasner, JJ, Jr. Comparison of the toxins of the blue-green alga Aphanizomenon flosaquae with the Gonyaulax toxins. Toxicon 1982, 20, 747–752. [Google Scholar]
- Ikawa, M; Auger, K; Mosley, S; Sasner, J; Noguchi, T; Hashimoto, K. Anderson, DM, White, AW, Baden, DG, Eds.; Toxin profiles of the blue-green alga Aphanizomenon flosaquae. In Toxic Dinoflagellates; Elsevier: New York, 1985; pp. 299–304. [Google Scholar]
- Mahmood, NA; Carmichael, WW. Paralytic shellfish poisons produced by the freshwater cyanobacterium Aphanizomenon flosaquae NH-5. Toxicon 1986, 24, 175–186. [Google Scholar]
- Pereira, P; Onodera, H; Andrinolo, D; Franca, S; Araújo, F; Lagos, N; Oshima, Y. Paralytic shellfish toxins in the freshwater cyanobacterium Aphanizomenon flosaquae, isolated from Montargil Reservoir, Portugal. Toxicon 2000, 38, 1689–1702. [Google Scholar]
- Li, RH; Carmichael, WW; Pereira, P. Morphological and 16S rRNA gene evidence for reclassification of the paralytic shellfish toxin producing Aphanizomenon flosaquae LMECYA 31 as Aphanizomenon issatschenkoi (Cyanophyceae). J Phycol 2003, 39, 814–818. [Google Scholar]
- Lagos, N; Onodera, H; Zagatto, PA; Azevedo, SM; Oshima, Y. The first evidence of paralytic shellfish toxins in the freshwater cyanobacterium Cylindrospermopsis raciborskii, isolated from Brazil. Toxicon 1999, 37, 1359–1373. [Google Scholar]
- Molica, R; Onodera, H; Garcia, C; Rivas, M; Andrinolo, D; Nascimento, S; Meguro, H; Oshima, Y; Azevedo, S; Lagos, N. Toxins in the freshwater cyanobacterium Cylindrospermopsis raciborskii (Cyanophyceae) isolated from Tabocas reservoir in Caruaru, Brazil, including demonstration of a new saxitoxin analogue. Phycologia 2002, 41, 606–611. [Google Scholar]
- Carmichael, WW; Evans, WR; Yin, QQ; Bell, P; Moczydlowski, E. Evidence for paralytic shellfish poisons in the freshwater cyanobacterium Lyngbya wollei (Farlow ex Gomont) comb. Nov. Appl Environ Microbiol 1997, 63, 3104–3110. [Google Scholar]
- Onodera, H; Satake, M; Oshima, Y; Yasumoto, T; Carmichael, WW. New saxitoxin analogues from the freshwater filamentous cyanobacterium Lyngbya wollei. Nat Tox 1997, 5, 146–151. [Google Scholar]
- Pomati, F; Sacchi, S; Rossetti, C; Giovannardi, S; Onodera, H; Oshima, Y; Neilan, BA. The freshwater cyanobacterium Planktothrix sp. FP1: molecular identification and detection of paralytic shellfish poisoning toxins. J Phycol 2000, 36, 553–562. [Google Scholar]
- Bravo, I; Cacho, E; Franco, JM; Miguez, A; Reyero, M I; Martinez, A. Yasumoto, T, Oshima, Y, Fukuyo, Y, Eds.; Study of PSP toxicity in Haliotis tuberculata from the Galician coast. In Harmful and Toxic Algal Blooms; Intergovernmental Oceanographic Commission of UNESCO: Paris, 1996; pp. 421–424. [Google Scholar]
- Shumway, SE. A review of the effects of algal blooms on shellfish and aquaculture. J World Aquacult Soc 1990, 21, 65–104. [Google Scholar]
- Shumway, SE. Phycotoxin-related shellfish poisoning: bivalve molluscs are not the only vectors. Rev Fish Sci 1995, 3, 1–31. [Google Scholar]
- Pereira, P; Dias, E; Franca, S; Pereira, E; Carolino, M; Vasconcelos, V. Accumulation and depuration of cyanobacterial paralytic shellfish toxins by the freshwater mussel Anodonata cygnea. Aquatic Toxicol 2004, 68, 339–350. [Google Scholar]
- Anderson, DM. Graneli, E, Sundstroem, B, Edler, L, Anderson, DM, Eds.; Toxin variability in Alexandrium species. In Toxic marine phytoplankton; Elsevier Press: New York, 1990; pp. 41–51. [Google Scholar]
- Anderson, DM; Kulis, DM. Biogeography of toxic dinoflagellates in the genus Alexandrium from the northeastern United States and Canada. Mar Biol 1994, 120, 467–478. [Google Scholar]
- Landsberg, JH. Neoplasia and biotoxins in bivalves: is there a connection? J Shellfish Res 1996, 15, 205–233. [Google Scholar]
- Landsberg, JH. The effects of harmful algal blooms on aquatic organisms. Rev Fish Sci 2002, 10, 113–390. [Google Scholar]
- Sakamoto, S; Ogata, T; Sato, S; Kodama, M; Takeuchi, T. Causative organism of paralytic shellfish toxins other than toxic dinoflagellates. Mar Ecol Prog Ser 1992, 89, 229–235. [Google Scholar]
- Doucette, GJ; Kodama, M; Franca, S; Gallacher, S. Anderson, DM, Cembella, AD, Hallegraeff, GM, Eds.; Bacterial interactions with harmful algal bloom species: bloom ecology, toxigenesis, and cytology. In Physiological Ecology of Harmful Algal Blooms; Springer-Verlag: Heidelberg, 1998; pp. 619–647. [Google Scholar]
- Kodama, M; Ogata, T; Sakamoto, S; Sato, S; Honda, T; Miwatani, T. Production of paralytic shellfish toxins by a bacterium Moraxella sp. isolated from Protogonyaulax tamarensis. Toxicon 1990, 28, 707–714. [Google Scholar]
- Kodama, M; Ogata, T; Sato, S; Sakamoto, S. Possible association of marine bacteria with paralytic shellfish toxicity in bivalves. Mar Ecol Prog Ser 1990, 61, 203–206. [Google Scholar]
- Kodama, M; Doucette, GJ; Green, DH. Relationships between bacteria and harmful algae. In Ecology of Harmful Algae; Springer-Verlag: Berlin, 2006; pp. 243–255. [Google Scholar]
- Ogata, T; Pholpunthin, P; Kukuyo, Y; Kodama, M. Occurrence of Alexandrium cohorticula in Japanese coastal water. J Appl Phycol 1990, 2, 351–356. [Google Scholar]
- Silva, ES. Intracellular bacteria: the origin of dinoflagellate toxicity. J Environ Pathol Toxicol Oncol 1990, 10, 124–128. [Google Scholar]
- Catterall, WA. Anderson, DM, White, AW, Baden, DG, Eds.; The voltage-sensitive sodium channel: a receptor for multiple toxins. In Toxic Dinoflagellates; Elsevier: New York, 1985; pp. 329–342. [Google Scholar]
- Kao, CY. Falconer, IR, Ed.; Paralytic shellfish poisoning. In Algal Toxins in Seafood and Drinking Water; Academic Press: London, 1993; pp. 75–86. [Google Scholar]
- Adnan, Q. Distribution of dinoflagellates at Jakarta Bay, Taman Jaya, Banten, and Benoa Bay, Bali: A report of an incident of fish poisoning at eastern Nusa Tenggara. Consult. Meet. on Toxic Red Tides and Shellfish Toxicity in Southeast Asia, Singapore, 11–14 September, 1984; White, AW, Anraku, M, Hooi, K-K, Eds.; pp. 25–27.
- Shumway, SE; Barter, J; Sherman-Caswell, S. Auditing the impact of toxic algal blooms on oysters. Environ Audit 1990, 2, 41–56. [Google Scholar]
- Shumway, SE; Cembella, AD. The impact of toxic algae on scallop culture and fisheries. Rev Fish Sci 1993, 1, 121–150. [Google Scholar]
- Bricelj, VM; Shumway, SE. An overview of the occurrence and transfer kinetics of paralytic shellfish toxins in bivalve molluscs. In Harmful and Toxic Algal Blooms, Proceedings of the Seventh International Conference on Toxic Phytoplankton, Sendai, Japan, 12–16 July, 1995; Yasumoto, T, Oshima, Y, Fukuyo, Y, Eds.; IOC of UNESCO: Paris, 1998; pp. 431–436. [Google Scholar]
- Bricelj, VM; Shumway, SE. Paralytic shellfish toxins in bivalve molluscs: occurrence, transfer kinetics, and biotransformation. Rev Fish Sci 1998, 6, 315–383. [Google Scholar]
- Schantz, EJ; Mold, JD; Stanger, DW; Shavel, J; Riel, J; Bowden, JP; Lynch, RS; Whyler, B; Reigel, B; Sommer, H. Paralytic shellfish poison. VI. A procedure for the isolation and purification of the poison from toxic clams and mussel tissues. J Am Chem Soc 1957, 78, 5230–5235. [Google Scholar]
- Schantz, EJ. Biochemical studies of paralytic shellfish poisons. Ann NY Acad Sci 1960, 89, 843–855. [Google Scholar]
- Twarog, BM; Yamaguchi, H. LoCicero, VR, Ed.; Resistance to paralytic shellfish toxins in bivalve molluscs. In Proc First Int Conf Toxic Dinoflagellate Blooms; Massachusetts Science and Technology Foundation: Wakefield, MA, 1975; pp. 381–393. [Google Scholar]
- Hartwell, AD. LoCicero, VR, Ed.; Hydrographic factors affecting the distribution and movement of toxic dinoflagellates in the western Gulf of Maine. In Proc First Int Conf Toxic Dinoflagellate Blooms; Massachusetts Science and Technology Foundation: Wakefield, MA, 1975; pp. 47–68. [Google Scholar]
- Shumway, SE; Sherman, SA; Cembella, AD; Selvin, R. Accumulation of paralytic shellfish toxins by surfclams, Spisula solidissima (Dillwyn, 1897) in the Gulf of Maine: seasonal changes, distribution between tissues, and notes on feeding habits. Nat Toxins 1994, 2, 236–251. [Google Scholar]
- Bricelj, VM; Cembella, AD. Lassus, P, Arzul, G, Erard-Le-Denn, E, Gentien, P, Marcaillou-Le-Baut, C, Eds.; Fate of gonyautoxins in surfclams, Spisula solidissima, grazing upon toxigenic Alexandrium. In Harmful Marine Algal Blooms; Technique et Documentation, Lavoisier: Paris, 1995; pp. 413–418. [Google Scholar]
- Cembella, AD; Shumway, SE. Lassus, P, Arzul, G, Erard-Le-Denn, E, Gentien, P, Marcaillou-Le-Baut, C, Eds.; Anatomical and spatio-temporal variation in PSP toxin composition in natural populations of the surf clam Spisula solidissima in the Gulf of Maine. In Harmful Marine Algal Blooms; Technique et Documentation, Lavoisier: Paris, 1995; pp. 421–426. [Google Scholar]
- Lassus, P; Wildish, DJ; Bardouil, M; Martin, JL; Bohec, M; Bougrier, S. Yasumoto, T, Oshima, Y, Fukuyo, Y, Eds.; Ecophysiological study of toxic Alexandrium spp. effects on the oyster Crassostrea gigas. In Harmful and Toxic Algal Blooms; Proceedings of the Seventh International Conference on Toxic Phytoplankton, Sendai, Japan, 12–16 July, 1995, IOC of UNESCO: Paris, 1996; pp. 409–412. [Google Scholar]
- Curtis, KM; Trainer, VL; Shumway, SE. Paralytic shellfish toxins in geoduck clams (Panope abrupta): variability, anatomical distribution, and comparison of two toxin detection methods. J Shellfish Res 2000, 19, 313–319. [Google Scholar]
- Smith, EA; Grant, F; Ferguson, CM; Gallacher, S. Biotransformations of paralytic shellfish toxins by bacteria isolated from bivalve mollusks. Appl Environ Microbiol 2001, 67, 2345–2353. [Google Scholar]
- Li, AMY; Yu, PKN; Hsieh, DPH; Wang, W-X; Wu, RSS; Lam, PKS. Uptake and depuration of paralytic shellfish toxins in the greenlipped mussel, Perna viridis: a dynamic model. Environ Toxicol Chem 2005, 24, 129–135. [Google Scholar]
- Twarog, B; Hidaka, T; Yamaguchi, H. Resistance to tetrodotoxin and saxitoxin in nerves of bivalve mollusks. Toxicon 1972, 10, 273–278. [Google Scholar]
- Bricelj, VM; Cembella, AD; Laby, D; Shumway, SE; Cucci, CL. Comparative physiological and behavioral responses to PSP toxins in two bivalve molluscs, the softshell clam, Mya arenaria, and surfclam, Spisula solidissima. Harmful and Toxic Algal Blooms 1996, 405–404. [Google Scholar]
- Bricelj, VM; Greene, M; Cembella, AD. Smayda, TJ, Shimuzu, Y, Eds.; Growth of the blue mussel Mytilus edulis on toxic Alexandrium fundyense and effects of gut passage on dinoflagellate cells. In Toxic Phytoplankton Blooms in the Sea; Elsevier Press: Amsterdam, 1993; pp. 371–376. [Google Scholar]
- Bricelj, VM; Connell, L; Konoki, K; Macquarrie, SP; Scheuer, T; Catterall, WA; Trainer, VL. Sodium channel mutation leading to saxitoxin resistance in clams increases risk of PSP. Nature 2005, 434(7034), 763–767. [Google Scholar]
- Beitler, MK; Liston, J. Graneli, E, Sundstrom, B, Edler, L, Anderson, DM, Eds.; Uptake and distribution of PSP toxins in butter clams. In Toxic Marine Phytoplankton; Elsevier Press: New York, 1990; pp. 257–262. [Google Scholar]
- Martin, JL; White, AW; Sullivan, JJ. Graneli, E, Sundstroem, B, Edler, L, Anderson, DM, Eds.; Anatomical distribution of paralytic shellfish toxins in softshell clams. In Toxic Marine Phytoplankton; Elsevier Press: New York, 1990; pp. 379–384. [Google Scholar]
- Cembella, AD; Shumway, SE; Larocque, R. Sequestering and putative biotransformation of paralytic shellfish toxins by the sea scallop Placopecten magellanicus: Seasonal and spatial scales in natural populations. J Exp Mar Biol Ecol 1994, 180, 1–22. [Google Scholar]
- Jamieson, GS; Chandler, RA. Paralytic shellfish poison in sea scallops (Placopecten magellanicus) in the west Atlantic. Can J Fish Aquat Sci 1983, 40, 313–318. [Google Scholar]
- Sullivan, JJ; Iwaoka, WT; Liston, J. Enzymatic transformation of PSP toxins in the littleneck clam (Protothaca staminaea). Biochem Biophys Res Commun 1983, 114, 465–472. [Google Scholar]
- Robertson, A; Stirling, D; Robillot, C; Llewellyn, L; Negri, A. First report of saxitoxin in octopi. Toxicon 2004, 44, 765–771. [Google Scholar]
- Daigo, K; Noguchi, T; Miwa, A; Kawai, N; Hashimoto, K. Resistance of nerves from certain toxic crabs to paralytic shellfish poison and tetrodotoxin. Toxicon 1988, 26, 485–490. [Google Scholar]
- Nagashima, Y; Ohgoe, H; Yamamoto, K; Shimakura, K; Shomi, K. Reguera, B, Blanco, J, Fernandez, ML, Wyatt, T, Eds.; Resistance of non-toxic crabs to paralytic shellfish poisoning toxins. In Harmful Algae; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Grafisant, Santiago de Compostela, Spain, 1998; pp. 604–606. [Google Scholar]
- Choi, M-C; Yu, PKN; Hsieh, DPH; Lam, PKS. Trophic transfer of paralytic shellfish toxins from clams (Ruditapes philippinarum) to gastropods (Nassarius festivus). Chemosphere 2006, 64, 1642–1649. [Google Scholar]
- Quayle, DB. Paralytic shellfish poisoning in eastern Canada. In Bull Fish Res Bd Can; Volume 168, Fisheries Research Board of Canada: Ottawa, Canada, 1971. [Google Scholar]
- Tufts, NR. Taylor, DL, Seliger, HH, Eds.; Molluscan transvectors of paralytic shellfish poisoning. In Toxic Dinoflagellate Blooms; Elsevier: New York, 1979; pp. 403–408. [Google Scholar]
- Prakash, A; Medcof, JC; Tennant, AD. Paralytic shellfish poisoning in Eastern Canada. Fish Res Bd Can 1971, Bull. No. 177. 87. [Google Scholar]
- White, AW; Nassif, J; Shumway, SE; Whitaker, DK. Recent occurrence of paralytic shellfish toxins in offshore shellfish in the northeastern United States. Toxic Phytoplankton Blooms in the Sea 1993, 435–440. [Google Scholar]
- Worms, J; Bouchard, N; Cormier, R; Pauley, KE; Smith, JC. Smayda, TJ, Shimizu, Y, Eds.; New occurrences of paralytic shellfish poisoning toxins in the southern Gulf of St. Lawrence, Canada. In Toxic Phytoplankton Blooms in the Sea; Elsevier: Amsterdam, 1993; pp. 353–358. [Google Scholar]
- Carreto, JI; Elbusto, C; Sancho, H; Carignan, M; Yasumoto, T; Oshima, Y. Comparative studies on paralytic shellfish toxin profiles of marine snails, mussels and an Alexandrium tamarense isolate from the Mar del Plata coast (Argentina). Rev Invest, Des Pesq 1996, 10, 101–107. [Google Scholar]
- Ito, K; Asakawa, M; Beppu, R; Takayama, H; Miyazawa, K. PSP-toxicification of the carnivorous gastropod Rapana venosa inhabiting the estuary of Nikoh River, Hiroshima Bay, Hiroshima Prefecture, Japan. Mar Poll Bull 2004, 48, 1116–1121. [Google Scholar]
- Sang, JWT; Ming, TT. Red tide and paralytic shellfish poisoning in Sabah, Malaysia. Consult. Meet. on Toxic Red Tides and Shellfish Toxicity in Southeast Asia, Singapore, 11–14 September, 1984; White, AW, Anraku, M, Hooi, K-K, Eds.; pp. 35–42.
- Ming, TT; Wong, T-SJ. Hallegraeff, GM, MacLean, JL, Eds.; Summary of red tide and paralytic shellfish poisoning in Sabah. In Biology, Epidemiology and Management of Pyrodinium red tides; Bandar Seri Begawan: Darussalam, Brunei, 1989; pp. 19–26. [Google Scholar]
- Kan, SK; Singh, N; Chan, MK. Oliva vidua fulminans, a marine mollusc, responsible for five fatal cases of neurotoxic food poisoning in Sabah, Malaysia. Trans R Soc Trop Med Hyg 1986, 80, 64–65. [Google Scholar]
- Jaafar, MH; Subramaniam, S. White, AW, Anraku, M, Hooi, K-K, Eds.; Occurrences of red tide in Brunei Darussalam and methods of monitoring and surveillance. In Toxic Red Tides and Shellfish Toxicity in Southeast Asia; Southeast Asian Fisheries Development Center and the International Development Research Center, Singapore, 1984; pp. 17–24. [Google Scholar]
- Jaafar, MH; de Silva, MWRN; Sharifuddin, PHY. Hallegraeff, GM, MacLean, JL, Eds.; Pyrodinium red tide occurrences in Brunei Darussalam. In Biology, Epidemiology and Management of Pyrodinium Red Tides; Bandar Seri Begawan, Brunei: Darussalam, 1989; pp. 9–17. [Google Scholar]
- Li, Y; Jia, X; Yang, M; Zhong, Y; Quan, G; Fan, P. Paralytic shellfish poison in contaminated shellfish along coast of China. Trop Oceanol 1999, 19, 90–96. [Google Scholar]
- Li, Y; Chen, S. Report of 8 incidents of intoxication due to Nassarius (Zeuxis) succinctus. Chin J Surg 1981, 20, 231–232. [Google Scholar]
- Nagashima, Y; Arakawa, O; Shiomi, K; Noguchi, T. Yasumoto, T, Oshima, Y, Fukuyo, Y, Eds.; Paralytic shellfish toxins in a trumpet shell, Charonia lampas, from Spain. In Harmful and Toxic Algal Blooms; Intergovernmental Oceanic Commission of UNESCO, Japan: Sendai, 1996; pp. 425–427. [Google Scholar]
- Liao, HJ; Hwang, DF. Survey of paralytic toxins in shellfish in southern Taiwan between 1995 and 1997. J Nat Toxins 2000, 9, 63–71. [Google Scholar]
- Hwang, DF; Cheng, CA; Jeng, SS. Gonyautoxin-3 as a minor toxin in the gastropod Niotha clathrata in Taiwan. Toxicon 1994, 32, 1573–1579. [Google Scholar]
- Sato, S; Ogata, T; Kodama, M. Smayda, TJ, Shimizu, Y, Eds.; Wide distribution of toxins with sodium channel blocking activity similar to tetrodotoxin and paralytic shellfish toxins in marine animals. In Toxic Phytoplankton Blooms in the Sea; Elsevier: Amsterdam, 1993; pp. 429–434. [Google Scholar]
- Negri, A; Llewellyn, L. Comparative analyses by HPLC and the sodium channel and saxiphilin 3H-saxitoxin receptor assays for paralytic shellfish toxins in crustaceans and molluscs from tropical North West Australia. Toxicon 1998, 36, 283–298. [Google Scholar]
- Martinez, A; Franco, JM; Bravo, I; Mazoy, M; Cacho, E. Smayda, TJ, Shimizu, Y, Eds.; PSP toxicity in Haliotis tuberculata from NW Spain. In Toxic Phytoplankton Blooms in the Sea; Elsevier: Amsterdam, 1993; pp. 419–423. [Google Scholar]
- Bravo, I; Reyero, MI; Cacho, E; Franco, JM. Paralytic shellfish poisoning in Haliotis tuberculata from the Galician coast: geographical distribution, toxicity by lengths and parts of the mollusc. Aquat Toxicol 1999, 46, 79–85. [Google Scholar]
- Pitcher, GC; Franco, JM; Doucette, GJ; Powell, CL; Mouton, A. Paralytic shellfish poisoning in the abalone Haliotis midae on the west coast of South Africa. J Shellfish Res 2001, 20, 895–904. [Google Scholar]
- Bravo, I; Franco, JM; Alonso, A; Dietrich, R; Molist, P. Cytological study and immunohistochemical location of PSP toxins in foot skin of the ormer, Haliotis tuberculata, from the Galician coast (NW Spain). Mar Biol 2001, 138, 709–715. [Google Scholar]
- Huchette, SMH; Clavier, J. Status of the ormer (Haliotis tuberculata L.) industry in Europe. J Shellfish Res 2004, 23(4), 951–955. [Google Scholar]
- Ruiz Sebastián, C; Etheridge, SM; Cook, PA; O’Ryan, C; Pitcher, GC. Phylogenetic analysis of toxic Alexandrium (Dinophyceae) isolates from South Africa: Implications for the global phylogeography of the Alexandrium tamarense species complex. Phycologia 2005, 44, 49–60. [Google Scholar]
- Etheridge, SM; Pitcher, GC; Roesler, CS. Steidinger, KA, Landsberg, JH, Thomas, CR, Vargo, GA, Eds.; Depuration and transformation of PSP toxins in the South African abalone Haliotis midae. In Harmful Algae 2002; Florida Fish and Wildlife Conservation Commission, Florida Institute of Oceanography, and Intergovernmental Oceanographic Commission of UNESCO, 2004; pp. 175–177. [Google Scholar]
- Anderson, DM; Kaefer, BA; McGillicuddy, DJ, Jr; Mickelson, MJ; Keay, KE; Libby, PS; Manning, JP; Mayo, CA; Wittaker, DK; Hickey, JM; He, R; Lynch, DR; Smith, KW. Initial observations of the 2005 Alexandrium fundyense bloom in southern New England: General patterns and mechanisms. Deep Sea Research Part II: Topical Studies on Oceanography 2005, 52, 2856–2879. [Google Scholar]
- Yasumoto, T; Oshima, Y; Konta, T. Analysis of paralytic shellfish toxins of xanthid crabs in Okinawa. Bull Jap Soc Sci Fish 1981, 47, 957–959. [Google Scholar]
- Arakawa, O; Noguchi, T; Shida, Y; Onoue, Y. Occurrence of carbamoyl-N-hydroxy derivatives of saxitoxin and neosaxitoxin in a xanthid crab Zosimus aeneus. Toxicon 1994, 32, 175–183. [Google Scholar]
- Arakawa, O; Nishio, S; Noguchi, T; Shida, Y; Onoue, Y. A new saxitoxin analogue from a Xanthid crab Atergatis floridus. Toxicon 1995, 33, 1577–1584. [Google Scholar]
- Arakawa, O; Noguchi, T; Onoue, Y. Paralytic shellfish toxin profiles of xanthid crabs Zosimus aeneus and Atergatis floridus collected on reefs of Ishigaki Island. Fish Sci 1995, 61, 659–662. [Google Scholar]
- Kotaki, Y; Tajiri, Y; Oshima, Y; Yasumoto, T. Identification of a calcareous red alga as the primary source of paralytic shellfish toxins in coral reef crabs and gastropods. Bull Jap Soc Sci Fish 1983, 49, 283–286. [Google Scholar]
- Jonas-Davies, J; Liston, J. Anderson, DM, White, AW, Baden, DG, Eds.; The occurrence of PSP toxins in intertidal organisms. In Toxic dinoflagellates; Elsevier: New York, 1985; pp. 467–472. [Google Scholar]
- Desbiens, M; Cembella, AD. Lassus, P, Arzul, G, Erard-Le-Denn, E, Gentien, P, Marcaillou-Le-Baut, C, Eds.; Occurrence and elimination kinetics of PSP toxins in the American lobster (Homarus americanus). In Harmful Marine Algal Blooms; Technique et Documentation, Lavoisier: Paris, 1995; pp. 433–438. [Google Scholar]
- Oikawa, H; Fujita, T; Satomi, M; Suzuki, T; Kotana, Y; Yano, Y. Accumulation of paralytic shellfish poisoning toxins in the edible shore crab Telmessus acutidens. Toxicon 2002, 40, 1593–1599. [Google Scholar]
- Oikawa, H; Fujita, T; Saito, K; Satomi, M; Yano, Y. Difference in the level of paralytic shellfish poisoning toxin accumulation between the crabs Telmessus acutidens and Charydbis japonica collected in Onahama, Fukushima Prefecture. Fish Sci 2007, 73, 395–403. [Google Scholar]
- Raj, U; Haq, U; Oshima, T; Yasumoto, T. The occurrence of paralytic shellfish toxins in two species of xanthid crab from Suva Barrier Reef, Fiji Islands. Toxicon 1983, 21, 547–551. [Google Scholar]
- Tsai, YH; Hwang, DF; Chai, TJ; Jeng, SS. Toxicity and toxic components of two xanthid crabs, Atergatis floridus and Demania reynaudi. Toxicon 1997, 35, 1327–1335. [Google Scholar]
- Tsai, YH; Hwang, DF; Chai, TJ; Jeng, SS. Occurrence of paralytic toxin in Taiwanese crab Atergatopsis germaini. Toxicon 1996, 34, 467–474. [Google Scholar]
- Llewellyn, LE; Endean, R. Toxins extracted from Australian specimens of the crab, Eriphia sebana (Xanthidae). Toxicon 1989, 27, 579–586. [Google Scholar]
- Llewellyn, LE; Endean, R. Toxicity and paralytic shellfish toxin profiles of the xanthid crabs, Lophozozymus pictor and Zosimus aeneus, collected from some Australian coral reefs. Toxicon 1989, 27, 596–600. [Google Scholar]
- Tsai, YH; Hwang, DF; Chai, TJ; Jeng, SS. Occurrence of tetrodotoxin and paralytic shellfish poison in the Taiwanese crab Lophozozymus pictor. Toxicon 1995, 33, 1669–1673. [Google Scholar]
- Ho, PH; Tsai, Y-H; Hwang, C-C; Hwang, P-A; Hwang, J-H; Hwang, D-F. Paralytic toxins in four species of coral reef crabs from Kenting National Park in southern Taiwan. Food Control 2006, 17, 439–445. [Google Scholar]
- Llewellyn, LE; Dodd, MJ; Robertson, A; Ericson, G; de Koning, C; Negri, AP. Postmortem analysis of samples from a human victim of a fatal poisoning caused by the xanthid crab, Zosimus aenus. Toxicon 2002, 40, 1463–1469. [Google Scholar]
- Yasumura, D; Oshima, Y; Yasumoto, T; Alcala, AC; Alcala, LC. Tetrodotoxin and paralytic shellfish toxins in Philippine crabs. Agric Biol Chem 1986, 50, 593–598. [Google Scholar]
- Fusetani, N; Endo, H; Hashimoto, K; Takahashi, K. Occurrence of potent toxins in the horseshoe crab Carcinoscorpius rotundicauda. Toxicon 1982, 20, 662–664. [Google Scholar]
- Jiang, TJ; Niu, T; Xu, YX. Transfer and metabolism of paralytic shellfish poisoning from scallop (Chlamys nobilis) to spiny lobster (Panulirus stimpsoni). Toxicon 2006, 48, 988–994. [Google Scholar]
- Koyama, K; Noguchi, T; Ueda, Y; Hashimoto, K. Occurrence of neosaxitoxin and other paralytic shellfish poisons in toxic crabs belonging to the family Xanthidae. Nippon Suis Gakk 1981, 47, 965. [Google Scholar]
- Llewellyn, LE. Haemolymph protein in xanthid crabs: its selective binding of saxitoxin and possible role in toxin bioaccumulation. Mar Biol 1997, 128, 599–606. [Google Scholar]
- Koyama, K; Noguchi, T; Uzu, A; Hashimoto;, K. Individual, local, and size-dependent variations in toxicity of the xanthid crab Zosimus aeneus. Nippon Suis Gakk 1983, 49, 1273–1279. [Google Scholar]
- Arakawa, O; Noguchi, T; Onoue, Y. Transformation of gonyautoxins in the xanthid crab Atergatis floridus. Fish Sci 1998, 64, 334–337. [Google Scholar]
- Alacala, A; Alacala, L; Garth, J; Yasumura, D; Yasumoto, T. Human fatality due to ingestion of the crab Demania reynaudii that contained a palytoxin-like toxin. Toxicon 1988, 26, 105–107. [Google Scholar]
- Lau, C; Tan, C; Li, Q; Ng, F; Tuen, R; Khoo, H. Bioactivity and mechanism of action of Lophozozymus pictor toxin. Toxicon 1995, 33, 901–908. [Google Scholar]
- Nocuchi, T; Uzu, A; Koyama, K; Maruyama, J; Nagashima, Y; Hashimyo, K. Occurrence of tetrodotoxin as the major toxin in a xanthid crab Atergatis floridus. Nippon Suis Gakk 1983, 49, 1887–1892. [Google Scholar]
- Oikawa, H; Fujita, T; Saito, K; Watabe, S; Yano, Y. Comparison of paralytic shellfish toxin between carnivorous crabs (Telmessus acutidens and Charybdis japonica) and their prey mussel (Mytilus galloprovincialis) in an inshore food chain. Toxicon 2004, 43, 713–719. [Google Scholar]
- Oikawa, H; Satomi, M; Watabe, S; Yano, Y. Accumulation and depuration rates of paralytic shellfish poisoning toxins in the shore crab Telmessus acutidens by feeding toxic mussels under laboratory controlled conditions. Toxicon 2005, 45, 163–169. [Google Scholar]
- Gessner, BD; Middaugh, JP. Paralytic shellfish poisoning in Alaska: A 20-year retrospective analysis. Am J Epidem 1995, 141, 766–770. [Google Scholar]
- Asakawa, M; Nishimura, F; Miyazawa, K; Noguchi, T. Occurrence of paralytic shellfish poisons in the starfish, Asterias amurensis in Kure Bay, Hiroshima Prefecture, Japan. Toxicon 1997, 35, 1081–1087. [Google Scholar]
- Lin, S-J; Tsai, Y-H; Lin, HP; Hwang, D-F. Paralytic toxins in Taiwanese starfish Astropecten scoparius. Toxicon 1998, 36, 799–803. [Google Scholar]
- White, AW. Taylor, DL, Seliger, HH, Eds.; Dinoflagellate toxins in phytoplankton and zooplankton fractions during a bloom of Gonyaulax excavata. In Toxic Dinoflagellate Blooms; Elsevier: New York, 1979; pp. 381–384. [Google Scholar]
- White, AW. Marine zooplankton can accumulate and retain dinoflagellate toxins and cause fish kills. Limnol Oceanogr 1981, 26, 103–109. [Google Scholar]
- Turiff, N; Runge, JA; Cembella, AD. Toxin accumulation and feeding behavior of the planktonic copepod Calanus finmarchicus exposed to the red-tide dinoflagellate Alexandrium excavatum. Mar Biol 1995, 123, 55–64. [Google Scholar]
- Teegarden, GJ; Cembella, AD. Grazing of toxic dinoflagellates, Alexandrium spp., by adult copepods of coastal Maine: implications for the fate of paralytic shellfish toxins in marine food webs. J Exp Mar Biol Ecol 1996, 196, 145–176. [Google Scholar]
- Teegarden, GJ; Cembella, AD. Yasumoto, T, Oshima, Y, Fukuyo, Y, Eds.; Grazing of toxic dinoflagellates, (Alexandrium spp.) by estuarine copepods: particle selection of PSP toxins in marine food webs. In Harmful and Toxic Algal Blooms; Proceedings of the Seventh International Conference on Toxic Phytoplankton, Sendai, Japan, 12–16 July, 1995, IOC of UNESCO: Paris, 1996; pp. 393–396. [Google Scholar]
- Turner, JY; Doucette, GJ; Powell, CL; Kulis, DM; Keafer, BA; Anderson, DM. Accumulation of red tide toxins in larger size fractions of zooplankton assemblages from Massachusetts Bay, USA. Mar Ecol Prog Ser 2000, 203, 95–107. [Google Scholar]
- Haya, K; Martin, JL; Waiwood, BA; Burridge, LE; Hungerford, JM; Zitko, V. Graneli, E, Sundstrom, B, Edler, L, Anderson, DM, Eds.; Identification of paralytic shellfish toxins in mackerel from southwest Bay of Fundy, Canada. In Toxic Marine Phytoplankton; Elsevier Press: New York, 1990; pp. 350–355. [Google Scholar]
- Castonguay, M; Levasseur, M; Beaulieu, J-L; Gregoire, F; Michaud, S; Bonneau, E; Bates, SS. Accumulation of PSP toxins in Atlantic mackerel: seasonal and ontogenetic variations. J Fish Biol 1997, 50, 1203–1213. [Google Scholar]
- Montoya, NG; Akselman, R; Pajaro, M; Perrotta, RG; Franco, J; Carreto, JI. Mortandad de caballa (Scomber japonicus) en la plataforma bonarense (Mar Argentino) asociada a un florecimento del dinoflagelado toxico Alexandrium tamarense. Rev Invest Des Pesq 1997, 11, 145–152. [Google Scholar]
- Nakashima, K; Arakawa, O; Taniyama, S; Nonaka, M; Takatani, T; Yamamori, K; Fuchi, Y; Noguchi, T. Occurrence of saxitoxins as a major toxin in the ovary of a marine puffer Arothron firmamentum. Toxicon 2004, 43, 207–212. [Google Scholar]
- Sato, S; Ogata, T; Borja, V; Gonzales, C; Fukuyo, Y; Kodama, M. Frequent occurrence of paralytic shellfish poisoning toxins as dominant toxins in marine puffer from tropical water. Toxicon 2000, 38, 1101–1109. [Google Scholar]
- Zaman, L; Arakawa, O; Shimosu, A; Onoue, Y. Occurrence of paralytic shellfish poison in Bangladeshi freshwater puffers. Toxicon 1997, 235, 423–431. [Google Scholar]
- Oliveira, JS; Fernandes, SCR; Schwartz, JAT; Pires, OR, Jr; de Freitas, JC. Toxicity and toxin identification in Colomesus asellus, an Amazonian (Brazil) freshwater puffer fish. Toxicon 2006, 48, 55–63. [Google Scholar]
- Kodama, M; Ogata, T; Kawamukai, K; Oshima, Y; Yasumoto, T. Occurrence of saxitoxin and other toxins in the liver of pufferfish Takifugu pardalis. Toxicon 1983, 21, 897–900. [Google Scholar]
- Nakamura, M; Oshima, Y; Yasumoto, T. Occurrence of saxitoxin in puffer fish. Toxicon 1984, 22, 381–385. [Google Scholar]
- Ahmed, MS; Jaime, E; Reichelt, M; Luckas, B. Hallegraeff, GM, Blackburn, SI, Bolch, CJ, Lewis, RJ, Eds.; Paralytic shellfish poisoning in freshwater puffer fish (Tetraodon cutcutia) from the River Burigonga, Bangladesh. In Harmful Algal Blooms 2000; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 2001; pp. 19–21. [Google Scholar]
- Sato, S; Kodama, M; Ogata, T; Saitanu, K; Furuya, M; Hirayama, K; Kakinuma, K. Saxitoxin as a toxic principle of a freshwater puffer, Tetraodon fangi, in Thailand. Toxicon 1997, 35, 137–140. [Google Scholar]
- Kungsuwan, A; Arakawa, O; Promset, M; Onoue, Y. Occurrence of paralytic shellfish poisons in Thai freshwater puffers. Toxicon 1997, 35, 1341–1346. [Google Scholar]
- Ngy, L; Tada, K; Yu, C-F; Takatani, T; Arakawa, O. Occurrence of paralytic shellfish toxins in Cambodian Mekong pufferfish Tetraodon turgidus: Selective toxin accumulation in the skin. Toxicon 2008, 51, 280–288. [Google Scholar]
- White, AW. Recurrence of kills of Atlantic herring (Clupea harengus harengus) caused by dinoflagellate toxins transferred through herbivorous zooplankton. Can J Fish Aquat Sci 1980, 37, 2262–2265. [Google Scholar]
- White, AW. Ragelis, EP, Ed.; Paralytic shellfish toxins and finfish. In Seafood Toxins; ACS Symposium Series 262, 1984; pp. 171–180. [Google Scholar]
- Kwong, RWM; Wang, WX; Lam, PKS; Yu, PKN. The uptake, distribution and elimination of paralytic shellfish toxins in mussels and fish exposed to toxic dinoflagellates. Aquat Toxicol 2006, 80, 82–91. [Google Scholar]
- Jiang, T-J; Wang, D-Z; Niu, T; Xu, YX. Trophic transfer of paralytic shellfish toxins from the cladoceran (Moina mongolica) to larvae of the fish (Sciaenops ocellatus). Toxicon 2007, 50, 639–645. [Google Scholar]
- Maclean, JL; White, AW. Anderson, DM, White, AW, Baden, DG, Eds.; Toxic dinoflagellate blooms in Asia: A growing concern. In Toxic dinoflagellates; Elsevier: New York, 1985; pp. 517–520. [Google Scholar]
- Gonzalez, CL; Ordonez, JA; Maala, AM. Okaichi, T, Anderson, DM, Nemoto, T, Eds.; Red tide: the Philippine experience. In Red Tides, Biology, Environmental Science and Toxicology; Elsevier: New York, 1989; pp. 97–100. [Google Scholar]
- Sibunka, JD; Pacheco, AL. Biological and fisheries data on northern puffer, Sphoeroides maculatus (Bloch and Schneider); National Marine Fisheries Service Technical Series Report No. 26: Highlands, New Jersey, 1981. [Google Scholar]
- CDC (Centers for Disease Control and Prevention). Neurologic illness associated with eating Florida puffer-fish. Morbid Mort Weekly Rep 2002, 51, 321–323.
- Quilliam, M; Wechsler, D; Marcus, S; Ruck, B; Wekell, M; Hawryluk, T. Detection and identification of paralytic shellfish poisoning toxins in Florida pufferfish responsible for incidents of neurologic illness. In Harmful Algae 2002, Proceedings of the Xth International Conference on Harmful Algae; Steidinger, KA, Landsberg, JH, Tomas, CR, Vargo, GA, Eds.; Florida Fish and Wildlife Conservation Commission and Intergovernmental Oceanographic Commission of UNESCO, 2004; pp. 116–118. [Google Scholar]
- Etheridge, S; Deeds, J; Hall, S; White, K; Flewelling, L; Abbott, J; Landsberg, J; Conrad, S; Bodager, D; Jackow, G. Detection methods and their limitations: PSP toxins in Florida puffer fish responsible for human poisoning events in 2004. African J Mar Sci 2006, 28, 383–387. [Google Scholar]
- Abbott, J; Landsberg, JH; Flewelling, LJ; Baird, L; Fisk, S. Transfer of saxitoxins within the Indian River Lagoon, Florida food web. ProcSecond Symposium on Harmful Algae in the U.S, 9–13 December, 2003; Woods Hole Oceanographic Institution: Woods Hole, MA; pp. 1–2. Available online: http://www.whoi.edu/redtide/2ndsymposium/SEGoMx_-abstracts.pdf [accessed 3 March 2008].
- Deeds, JR; White, KD; Etheridge, SM; Landsberg, JH. Concentrations of saxitoxin and tetrodotoxin in three species of puffer fish from the Indian River Lagoon, Florida, USA - the location for multiple cases of saxitoxin puffer fish poisoning from 2002–2004. Trans Am Fish Soc. in press.

{kind=link}
{kind=link}
| Species | Saxitoxin and derivatives | References |
|---|---|---|
| Dinoflagellates | ||
| Alexandrium acatenella | STX | 7–9 |
| Alexandrium andersoni | STX, NEO | 10–11 |
| Alexandrium angustitabulatum | unknown toxin composition | 12 |
| Alexandrium catenella | STX, GTX1–4, NEO,B1–2, C1–4 | 13–21 |
| Alexandrium cohorticula | STX, GTX1–4 | 22–23 |
| Alexandrium fundyense | STX, NEO, GTX1–4, C1–2, B1 | 9, 24–25 |
| Alexandrium minutum (= A. lusitanicum) | GTX1–4 | 20, 26–30 |
| Alexandrium ostenfeldii | GTX2–3, B2, C1–2 | 31–34 |
| Alexandrium tamarense | STX, NEO, GTX1–4, B1, C1, C2, C4 | 9, 21, 35–40 |
| Alexandrium tamiyavanichi | STX, GTX1–4, B1, C1–4 | 41–42 |
| Cochlodinium polykrikoides (= C. heterolobatum, Cochlodinium type’78) | zinc-bound carbamoyl hydroxy NEO | 5 |
| Gymnodinium catenatum | STX, NEO, trace GTX2–3, B1–2, C1–4 | 30, 41, 43–46 |
| Pyrodinium bahamense | STX, NEO, B1–B2 | 41, 47–58 |
| Cyanobacteria | ||
| Anabaena circinalis | STX, GTX1–4, C1–C2, dcGTX2–3 | 3, 59–66 |
| Anabaena lemmermannii | STX | 67 |
| Aphanizomenon gracile | STX, NEO | 68 |
| Aphanizomenon issatschenkoi (as A. flosaquae) | NEO, STX | 69–77 |
| Cylindrospermopsis raciborskii | STX, NEO, GTX2–3 | 78–79 |
| Lyngbya wollei | dcSTX, dcGTX2–3, acetylated STX analogues | 80–81 |
| Planktothrix sp. | STX | 82 |
| Rivularia sp. | GTX2, GTX4 | 83 |
| Gastropod species and presumptive microalgal source | Common name | Maximum STX(s) concentration | Incident | Location | Reference |
|---|---|---|---|---|---|
| Alexandrium acatenella | |||||
| Polinices lewisii | Lewis moon snail | 176–600 μg STX eq./100g tissue | British Columbia, Canada | 129 | |
| Alexandrium catenella | |||||
| Adelomelon ancilla | Volute | toxic | Chile | 85 | |
| Argobuccinum sp. | Whelk | Stomach 5629 μg STX eq./100g tissue; Muscle 92 μg STX eq./100g tissue | |||
| Concholepas concholepas | Barnacle rock shell | toxic | |||
| Trophon sp. | Trophon | toxic | |||
| Nassarius sp. | Nassa mud snail (dog whelk) | 9 μg STX eq./100g tissue | Washington, USA | 85 | |
| Neptunea spp. | 200–250 MU* 100 g−1 whole individuals | Alaska, USA | 85 | ||
| Thais sp. | Oyster drill | 23 μg STX eq./100g tissue (GTX 2 and GTX 3 only) | Washington, USA | 85 | |
| Thais lamellosa | Oyster drill | Whole animal positive | |||
| Thais lima | Oyster drill | Whole animal 180 μg STX eq./100g tissue | |||
| Alexandrium tamarense | |||||
| Littorina sitkana | Sitka periwinkle | Trace whole animal | Washington, USA | 85 | |
| Lunatia heros (as Polinicies heros) | Northern moon snail | 1450 μg STX eq./100g tissue | 2 cases PSP | Massachusetts, USA | 130 |
| Buccinum undatum | Waved whelk | whole body 608 μg STX eq./100g tissue; digestive gland 1600 μg STX eq./100g tissue
3337 μg STX eq./100g tissue | 12 cases PSP, 4 fatalities
Illnesses and deaths | Quebec, Canada
Gulf of Maine, USA | 85, 131 85, 132 |
| Crepidula fornicata | Slipper limpet | 46–58 μg STX eq./100g tissue | |||
| Colus stimpsoni | Stimpson’s colus | toxic | |||
| Lunatia heros (=Euspira heros, Polinices heros) | Northern moon snail | 2922 μg STX eq./100g tissue | |||
| Neptunea decemcostata | Ten-ridged whelk | Raw~3000–4000, steamed 1060 μg STX eq./100g tissue | |||
| Thais lapillus | Purpura | 34 μg STX eq./100g tissue | |||
| Lunatia heros (=Euspira heros, Polinices heros) | Northern moon snail | 247 μg STX eq./100g tissue | Gulf of St. Lawrence, Canada | 133 | |
| Adelomedon brasiliana | Volute | 28 MU g−1 whole | Argentina | 134 | |
| Zidona angulata* | Volute | 210 MU g−1 viscera; 25 MU g−1 foot; 17 MU g−1 mucus | Mild case of PSP | ||
| Busycon spp. | Whelk | 50–500 MU 100 g−1 | Quebec, Canada | 85 | |
| Rapana venosa | Veined rapa whelk | 11.4 MU g−1 viscera | Hiroshima Bay, Japan | 135 | |
| Gymnodinium catenatum | |||||
| Haliotis tuberculata | Abalone | 467 μg STX eq./100g muscle | Spain | 83 | |
| Pyrodinium bahamense | |||||
| Lambis lambis | Spider conch | ND – 175 MU 100 g−1 whole | Several PSP cases | Sabah, Malaysia | 136–137 |
| Oliva vidua fulminans | Olive | 2525 MU 100 g−1 whole | 5 human fatalities; 8 cases of PSP | Malaysia | 136–138 |
| Natica sp.** | “Tekuyong” | 71–876 MU 100 g−1 | Borneo | 139–140 | |
| Unknown origin | |||||
| Nassarius siguijorensis | Nassa | 370 MU 100 g−1 | Daya Bay, Guangdong Province | 141 | |
| Nassarius succinctus | Nassa | 68 cases of PSP, March–Aug 1979; 1 fatality and 7 hospitalized | Zhejiang Povince, China | 128, 142 | |
| Nassarius spp. | Nassa | 50 PSP cases, 3 fatalities, April– May 2002
55 PSP cases, 1 fatality; summer 2004 | Fujian Province, China
Yin Chuan city, China | 128 128 | |
| Nassarius spp. | Nassa | 107,413 MU 100 g−1 | Zhoushan Islands, China | 128 | |
| Charonia lampas | Trumpet shell | 17.5 MU g−1 digestive gland | Galicia, Spain | 143 | |
| Natica lineata | Lined moon shell | PSP toxins | Taiwan | 144 | |
| Natica vitellus | Calf moon shell | ||||
| Niotha clathrata | Basket shell | PSP, GTX-3 | 144–145 | ||
| Neptunea arthritica | Arthritic neptune | GTX 1–4, neoSTX, STX | Sanriku coast, Japan | 146 | |
| Tectus fenestratus | Fenestrate top shell | 18.7 μg STX eq./100g tissue | Northwest Australia | 147 | |
| Tectus nilotica maxima | Top shell | 5.0 MU g−1 whole | Ishigaki Island, Japan | 52 | |
| Tectus pyramis | Top shell | 19 MU g−1 whole | Ishigaki Island, Japan | 52 | |
| Turbo argyrostoma | Turban shell | 20 MU g−1 whole | Ishigaki Island, Japan | 52 | |
| Turbo marmorata | Turban shell | 4.2 MU g−1 whole | Ishigaki Island, Japan | 52 | |
| Sampling Date | Common Name | Scientific Name | Number of Animals | Sampling Coordinates | STX eq. (μg/100g) |
|---|---|---|---|---|---|
| 7-8-06 | Ocean Quahog | Arctica islandica | 8 | 41 00.183N | 7.2 |
| 70 44.543W | |||||
| 7-8-06 | Ocean Quahog | Arctica islandica | 3 | 41 06.476N | 11.6 |
| 70 27.150W | |||||
| 7-9-06 | Whelk | Busycon sp. | 3 | 41 25.057N | 234.3 |
| 70 02.751W | |||||
| 7-9-06 | Atlantic Surfclam | Spisula solidissima | 3 | 41 25.057N | 15.6 |
| 70 02.751W | |||||
| 7-9-06 | Blue Mussels | Mytilus edulus | 12 | 41 23.836N | 19.5 |
| 69 53.954W | |||||
| 7-9-06 | Blue Mussels | Mytilus edulus | 12 | 41 23.836N | 26.3 |
| 69 53.954W | |||||
| 7-9-06 | Northern Moon Snail | Lunatia heros | 3 | 41 26.084N | 265.5 |
| 70 03.000W | |||||
| 7-9-06 | Northern Moon Snail | Lunatia heros | 7 | 41 23.836N | 321.0 |
| 69 53.954W | |||||
| 7-10-06 | Sea Scallops | Placopecten magellanicus | 9 | 42 09.865N | 228.8 |
| 70 18.279W | |||||
| 7-10-06 | Sea Scallop viscera (F) | Placopecten magellanicus | 1 | 42 09.865N | 93.6 |
| 70 18.279W | |||||
| 7-10-06 | Sea Scallop viscera (M) | Placopecten magellanicus | 1 | 42 09.865N | 131.9 |
| 70 18.279W | |||||
| 7-11-06 | Ocean Quahog | Arctica islandica | 11 | 42 12.025N | <LOD |
| 70 22.017W | |||||
| 7-11-06 | Sea Scallop | Placopecten magellanicus | 6 | 42 11.391N | 50.6 |
| 70 19.700W | |||||
| 7-11-06 | Northern Moon Snails | Lunatia heros | 6 | 42 11.391N | 318.9 |
| 70 19.700W | |||||
| 7-11-06 | Ocean Quahogs | Arctica islandica | 12 | 42 12.025N | <LOD |
| 70 22.017W | |||||
| 7-11-06 | Blue Mussels | Mytilus edulus | 9 | 42 12.025N | 5.0 |
| 70 22.017W | |||||
| 7-11-06 | Atlantic Surfclam | Spisula solidissima | 2 | 42 11.391N | 16.1 |
| 70 19.700W | |||||
| 7-11-06 | Ocean Quahog | Arctica islandica | 5 | 42 12.025N | 12.0 |
| 70 22.017W | |||||
| 7-11-06 | Ocean Quahog | Arctica islandica | 4 | 42 11.391N | 0.2 |
| 70 19.700W |
| Crustacean species and presumptive microalgal source | Common name | Maximum STX(s) concentration | Location | Reference |
|---|---|---|---|---|
| Alexandrium catenella | ||||
| Cancer magister | Dungeness crab | 72 μg STX eq./100g viscera | Washington, USA | 85 |
| Cancer productus | Red rock crab | 285 μg STX eq./100g viscera
27 μg STX eq./100g muscle | Washington, USA | 161 |
| Fabia subquadrata | Pea crab | 32 μg STX eq./100g whole crabs | Washington, USA | 85 |
| Hemigrapsus nudus | Purple shore crab | 44 μg STX eq./100g whole body minus legs and carapace | Washington, USA | 161 |
| Hemigrapsus oregonensis | Green shore crab | 31 μg STX eq./100g whole | Washington, USA | 161 |
| Pagurus sp. | Hermit crab | 35 μg STX eq./100g whole crabs | Washington, USA | 85 |
| Pugettia producta | Northern kelp crab | 146 μg STX eq./100g eggs; 1710 μg STX eq./100g viscera; 48 μg STX eq./100g muscle | Washington, USA | 161 |
| Balanus spp. | Barnacles | 84 μg STX eq./100g whole | Washington, USA | 161 |
| Alexandrium tamarense | ||||
| Anonyx sarsi | Gammarid amphipod | 180 μg STX eq./100g (tissue not specified) | St.Lawrence estuary, Canada | 162 |
| Cancer borealis | Jonah crab | 56 μg STX eq./100g (tissue not specified) | Maine, USA | 85 |
| Homarus americanus | American lobster | 1512 μg STX eq./100g hepatopancreas (bioassay); 961 μg STX eq./100g hepatopancreas (HPLC); 69 μg STX eq./100g meat (HPLC) | Bay of Gaspe, Canada | 162 |
| Pyrodinium bahamense | ||||
| ND | Crab | 339 MU* 100 g−1 | Brunei Darussalam | 141 |
| ND | Mangrove crabs | 239 MU 100 g−1 guts; 175 MU 100 g−1 gills | Sabah, Malaysia | 138 |
| Portunus pelagicus | Blue manna crab | 175 MU 100 g−1 whole crab; 288 MU
100 g−1 gills; 328 MU 100 g−1 guts 1.8 μg STX eq./100g whole | Sabah, Malaysia
Northwest Australia | 138 147 |
| Panulirus versicolor | Painted spiny lobster | 175 MU 100 g−1 whole lobster; 175 MU 100 g−1 body only | Sabah, Malaysia | 138 |
| Panulirus longipes | Longlegged spiny lobster | 211 MU 100 g−1 whole lobster; 177 MU 100 g−1 head and legs | Sabah, Malaysia | 138 |
| ND | Penaeid shrimp | 175 MU 100 g−1 frozen tails; 268 MU 100 g−1 body only | Sabah, Malaysia | 138 |
| ND | Penaeid shrimp “Udang” | 190 MU 100 g−1 | Brunei Darussalam | 141 |
| Unknown origin | ||||
| Hemigrapsus sanguineus | Asian shore crab | 0.16 MU g−1 hepatopancreas | Sanriku coast, Japan | 146 |
| Metopograpsus frontalis | Mangrove shore crab | 10.0 μg STX eq./100g whole | Northwest Australia | 147 |
| Pachygrapsus crassipes | Striped shore crab | 0.10 MU g−1 hepatopancreas | Sanriku coast, Japan | 146 |
| Percnon planissimum | Sally lightfoot crab | 7.4 MU g−1 whole | Ishigaki Island, Japan | 52 |
| Pilumnus pulcher | Hairy crab | 80 μg STX eq./100g whole | Northwest Australia | 147 |
| Pilumnus vespertilio | Hairy crab | 120 μg STX eq./100g whole
6.1 MU g−1 whole | Northwest Australia
Ishigaki Island, Japan | 147 52 |
| Schizophrys aspera | Eyelash spider crab | 2.3 MU g−1 whole | Ishigaki Island, Japan | 52 |
| Telmessus acutidens | Edible shore crab | 2723 μg STX eq./100g viscera | Fukushima Prefecture, Japan | 163,164 |
| Actaeodes tomentosus | Xanthid crab | 130 MU g−1 whole | Ishigaki Island, Japan | 52 |
| Atergatis floridus | Xanthid crab | Positive STX, NEO, GTX2
16,611 μg STX eq./100g whole 490 MU g−1 whole Positive GTX 1–4 | Fiji Islands
Northwest Australia Ishigaki Island, Japan | 165 147 52 |
| Atergatopsis germaini | Xanthid crab | Positive GTX 3, NEO, STX | Taiwan | 167 |
| Demania reynaudi | Xanthid crab | Positive GTX 3–4, NEO | Taiwan | 166 |
| Eriphia scabricula | Xanthid crab | 180 MU g−1 whole | Ishigaki Island, Japan | 52 |
| Eriphia sebana | Xanthid crab | Positive STX, NEO, GTX1, GTX2 | Great Barrier Reef, Australia | 168 |
| Euzanthus exsculptus | Xanthid crab | 29 μg STX eq./100g whole | Northwest Australia | 147 |
| Lophozozymus octodentatus | Xanthid crab | 23 μg STX eq./100g whole | Northwest Australia | 147 |
| Lophozozymus pictor | Xanthid crab | 18.9 MU g−1 whole crab
Positive GTX | Australia
Taiwan | 169 170 |
| Neoxanthias impressus | Xanthid crab | 147 μg STX eq./100g whole
10 MU g−1 whole | Northwest Australia
Ishigaki Island, Japan | 147 52 |
| Platypodia granulosa | Xanthid crab | 110 MU g−1 whole | Ishigaki Island, Japan | 52 |
| Platypodia pseudogranulosa | Xanthid crab | 10 μg STX eq./100g whole | Northwest Australia | 147 |
| Xanthias lividus | Xanthid crab | Positive GTX | Taiwan | 171 |
| Zosimus aeneus | Xanthid crab | Positive STX, NEOSTX, GTXI-3
Positive GTX 660 MU g−1 whole 108,000 μg STX eq./100g chelae muscle; 720 μg STX eq./100g cephalothorax muscle 78 μg STX eq./100g whole 259 MU g−1 whole crab | Fiji Islands
Taiwan Ishigaki Island, Japan Japan Northwest Australia Philippines | 165 171 52 172 147 173 |
| Procambarus clarkii | Red swamp crayfish | 0.23 MU g−1 hepatopancreas | Sanriku, Japan | 146 |
| Carcinoscorpius rotundicauda | Mangrove horseshoe crab | STX | Thailand | 174 |
| Fish species and presumptive microalgal source | Common name | Maximum STX(s) concentration | Location | Reference |
|---|---|---|---|---|
| Alexandrium fundyense | ||||
| Scomber scombrus | Atlantic mackerel | 209 μg STX eq./100g liver; 367 μg STX eq./100g liver | Bay of Fundy; Gulf of St. Lawrence | 194–195 |
| Alexandrium tamarense | ||||
| Scomber japonicus | Chub mackerel | 2800 μg STX eq./100g muscle; 500 μg STX eq./100g liver; 72 μg STX eq./100g gills | Argentina | 196 |
| Pyrodinium bahamense | ||||
| Rastrelliger sp. | Short mackerel | 99 MU 100 g−1 tissue | Brunei Darussalam | 141 |
| Sardinella sp. | Sardinella | 99 MU 100 g−1 tissue
572 μg STX eq./100g guts | Brunei Darussalam
Sabah, Malaysia | 141 139 |
| Sphoeroides nephelus | Southern puffer fish | 1,443 μg STX eq./100g liver; 14,571 μg STX eq./100g muscle | USA | 58 |
| Sphoeroides testudineus | Checkered puffer fish | 51.1 μg STX eq./100g liver; 104.3 μg STX eq./100g muscle | USA | 58 |
| Sphoeroides spengleri | Bandtail puffer fish | 364.5 μg STX eq./100g muscle | USA | 58 |
| Unknown origin | ||||
| Cololabis saira | Pacific saury | 0.14 MU g−1 viscera | Iwate, Japan | 146 |
| Gadus macrocephalus | Pacific cod | 0.10 MU g−1 viscera;
0.10 MU g−1 intestine | Iwate, Japan | 146 |
| Lamna ditropis | Salmon shark | 0.17 MU g−1 liver | Iwate, Japan | 146 |
| Oncorhynchus keta | Chum salmon | 1.53 MU g−1 liver;
0.69 MU g−1 viscera | Iwate, Japan | 146 |
| Scarus (= Ypsiscarus) ovifrons | Knobsnout parrotfish | 0.26 MU g−1 liver;
1.58 MU g−1 intestine | Iwate, Japan | 146 |
| Arothron firmamentum | Starry toadfish | 740 MU g−1 ovary | Japan | 197 |
| A. hispidus | White-spotted puffer | Positive STX in liver, muscle, skin, and intestine | Philippines | 198 |
| A. mappa | Map puffer | Positive STX in liver, muscle, skin, and intestine | Philippines | 198 |
| A. manillensis | Narrow-lined puffer | Positive STX in liver, muscle, skin, and intestine | Philippines | 198 |
| A. nigropunctatus | Black spotted puffer | Positive STX in liver, muscle, skin, and intestine | Philippines | 198 |
| A. reticularis | Reticulated puffer | Positive STX in liver, muscle, skin, and intestine | Philippines | 198 |
| A. stellatus | Starry toadfish | Positive STX in liver, muscle, skin, and intestine | Philippines | 198 |
| Chelonodon patoca | Milk-spotted puffer | 22.0 MU g−1 muscle; 40 MU g−1 skin; 12.0 MU g−1 liver; 2.8 MU g−1 ovary (data shown as mean)
Positive STX in liver, muscle, skin, and intestine | Bangladesh
Philippines | 199 198 |
| Colomesus asellus | Amazon puffer | 53.2 MU whole body | Brazil | 200 |
| Takifugu pardalis | Panther puffer | Positive for STX in liver | Japan | 201 |
| T. poecilonotus | Fine patterned puffer | Positive for STX in liver, ovary and digestive tract | Japan | 202 |
| T. radiates | Puffer | Positive for STX in liver | Japan | 202 |
| T. vermicularis | Purple puffer | Positive for STX in liver, ovary and digestive tract | Japan | 202 |
| Tetraodon cutcutia | Ocellated puffer | 7.6 MU g−1 muscle; 20 MU g−1 skin; 6.0 MU g−1 liver;
5.6 MU g−1 ovary (data shown as mean) 182 MU 100 g−1 skin; 238 MU 100 g−1 muscle; 106 MU 100 g−1 liver | Thailand
Bangladesh | 199 203 |
| T. cochinchinensis (as T. fangi) | Puffer | Positive for STX whole body | Thailand | 204 |
| T. suvatii | Arrowhead puffer | 191 MU g−1 muscle; 230 MU g−1 skin; 174 MU g−1 liver; 117 MU g−1 egg | Thailand | 205 |
| T. turgidus | Brown puffer | <2 MU g−1 muscle; 37 MU g−1 skin; <2 MU g−1 liver; 27 MU g−1 ovary | Cambodia | 206 |
Share and Cite
Deeds, J.R.; Landsberg, J.H.; Etheridge, S.M.; Pitcher, G.C.; Longan, S.W. Non-Traditional Vectors for Paralytic Shellfish Poisoning. Mar. Drugs 2008, 6, 308-348. https://doi.org/10.3390/md6020308
Deeds JR, Landsberg JH, Etheridge SM, Pitcher GC, Longan SW. Non-Traditional Vectors for Paralytic Shellfish Poisoning. Marine Drugs. 2008; 6(2):308-348. https://doi.org/10.3390/md6020308
Chicago/Turabian StyleDeeds, Jonathan R., Jan H. Landsberg, Stacey M. Etheridge, Grant C. Pitcher, and Sara Watt Longan. 2008. "Non-Traditional Vectors for Paralytic Shellfish Poisoning" Marine Drugs 6, no. 2: 308-348. https://doi.org/10.3390/md6020308