Sesterterpenoid and Steroid Metabolites from a Deep-Water Alaska Sponge Inhibit Wnt/β-Catenin Signaling in Colon Cancer Cells



Abstract
:1. Introduction
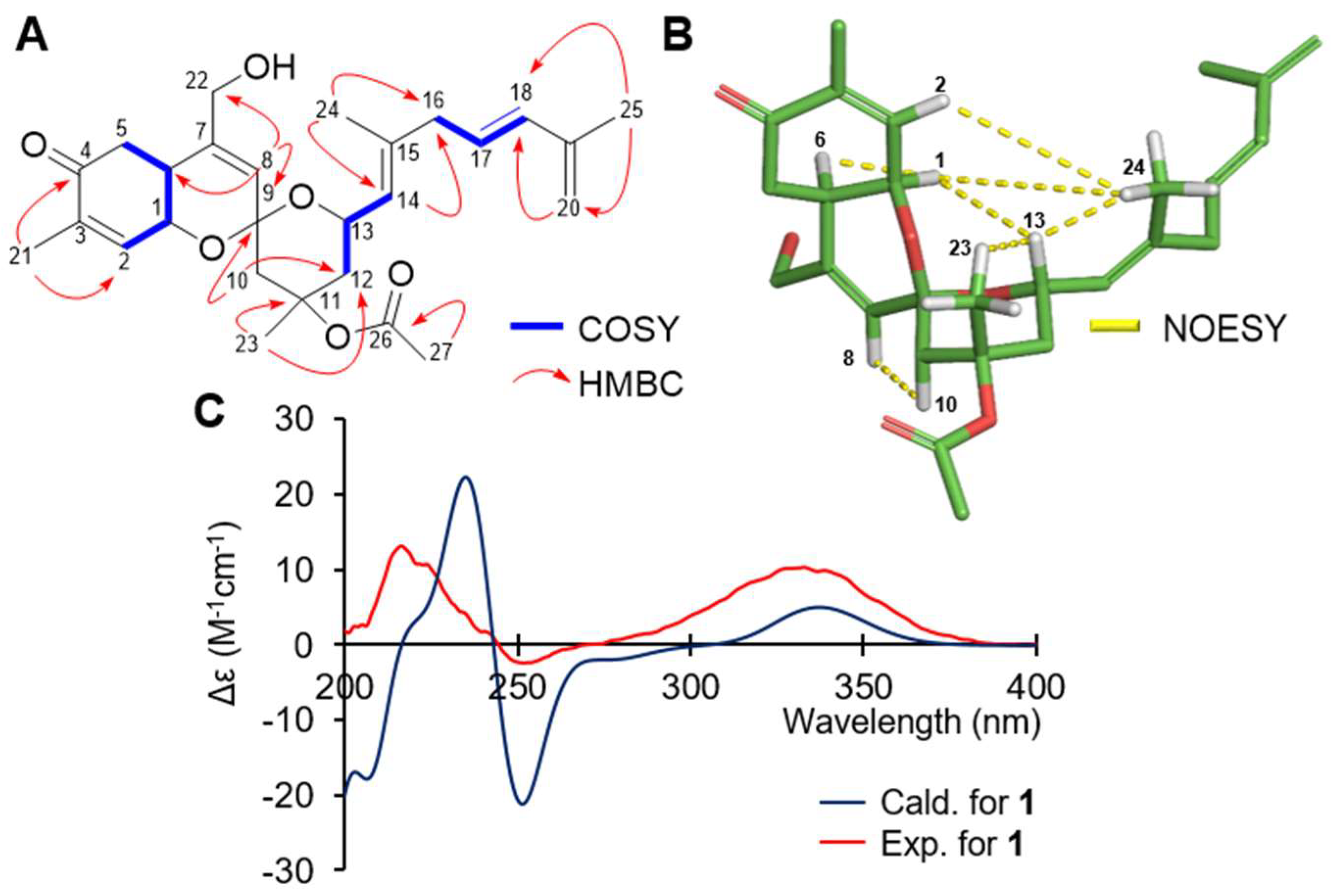
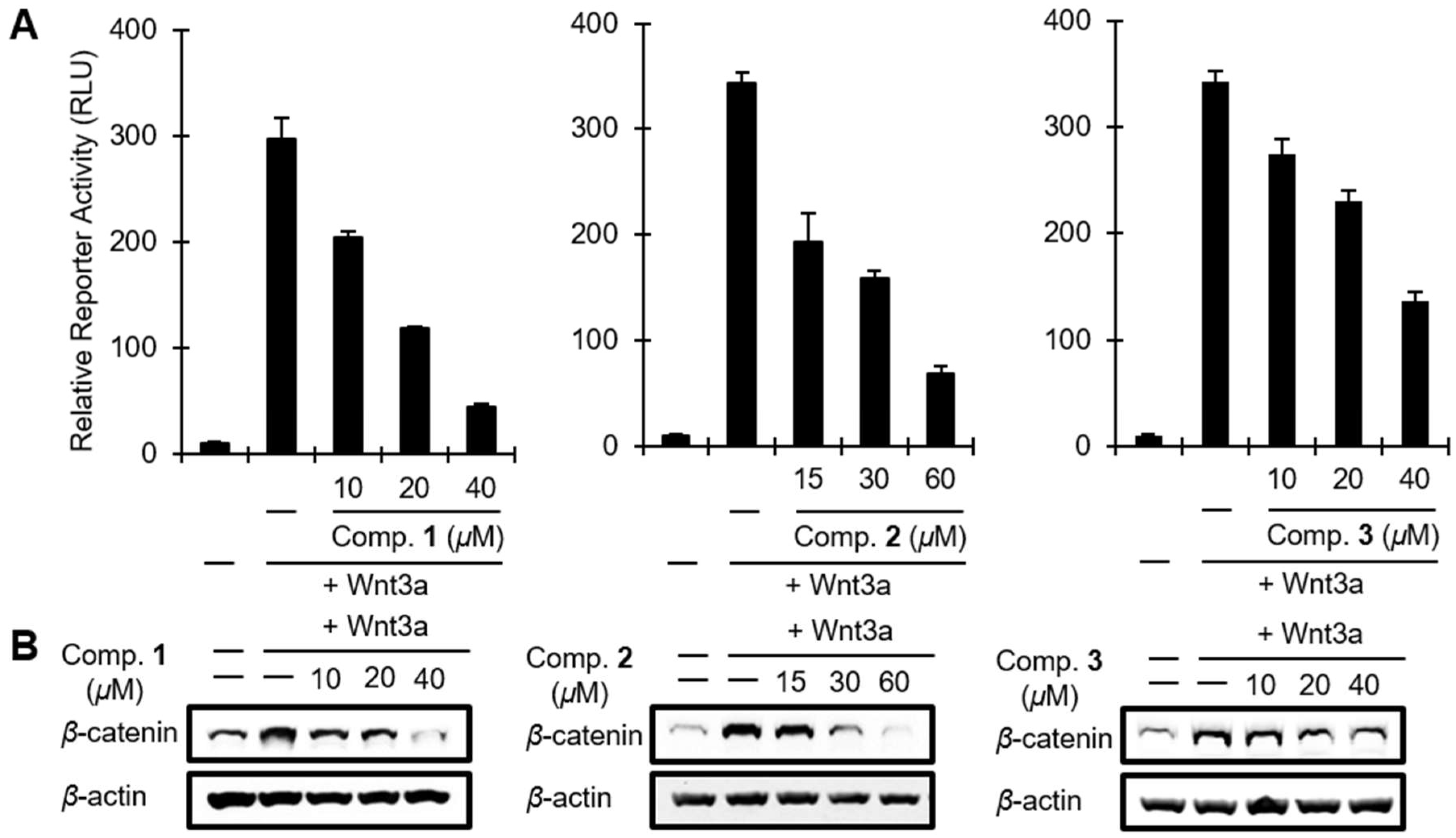
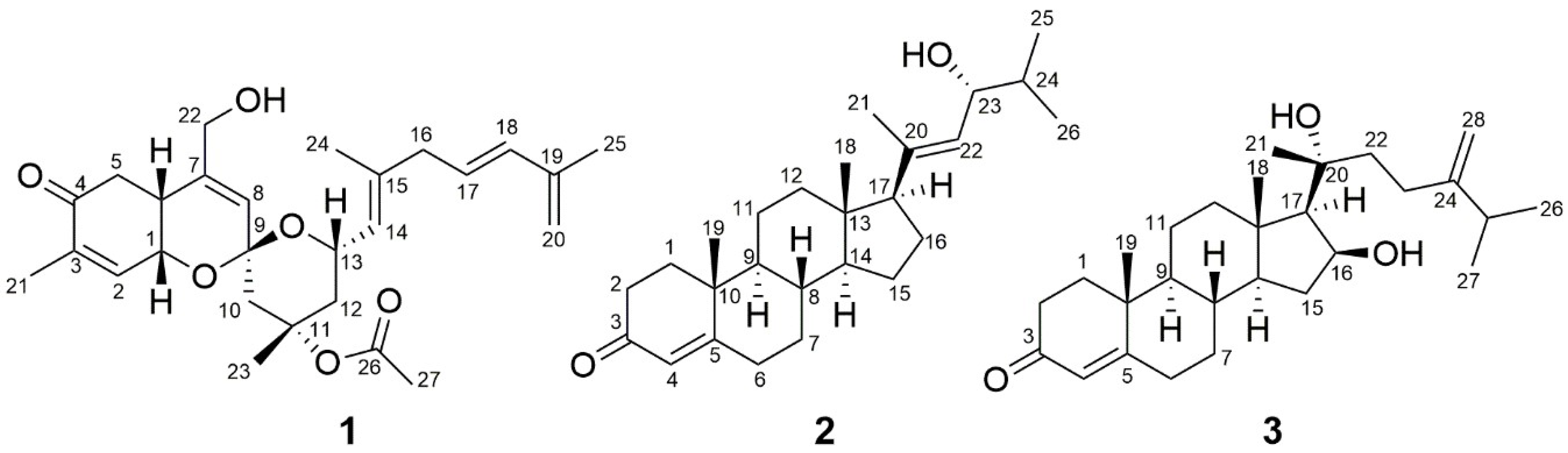
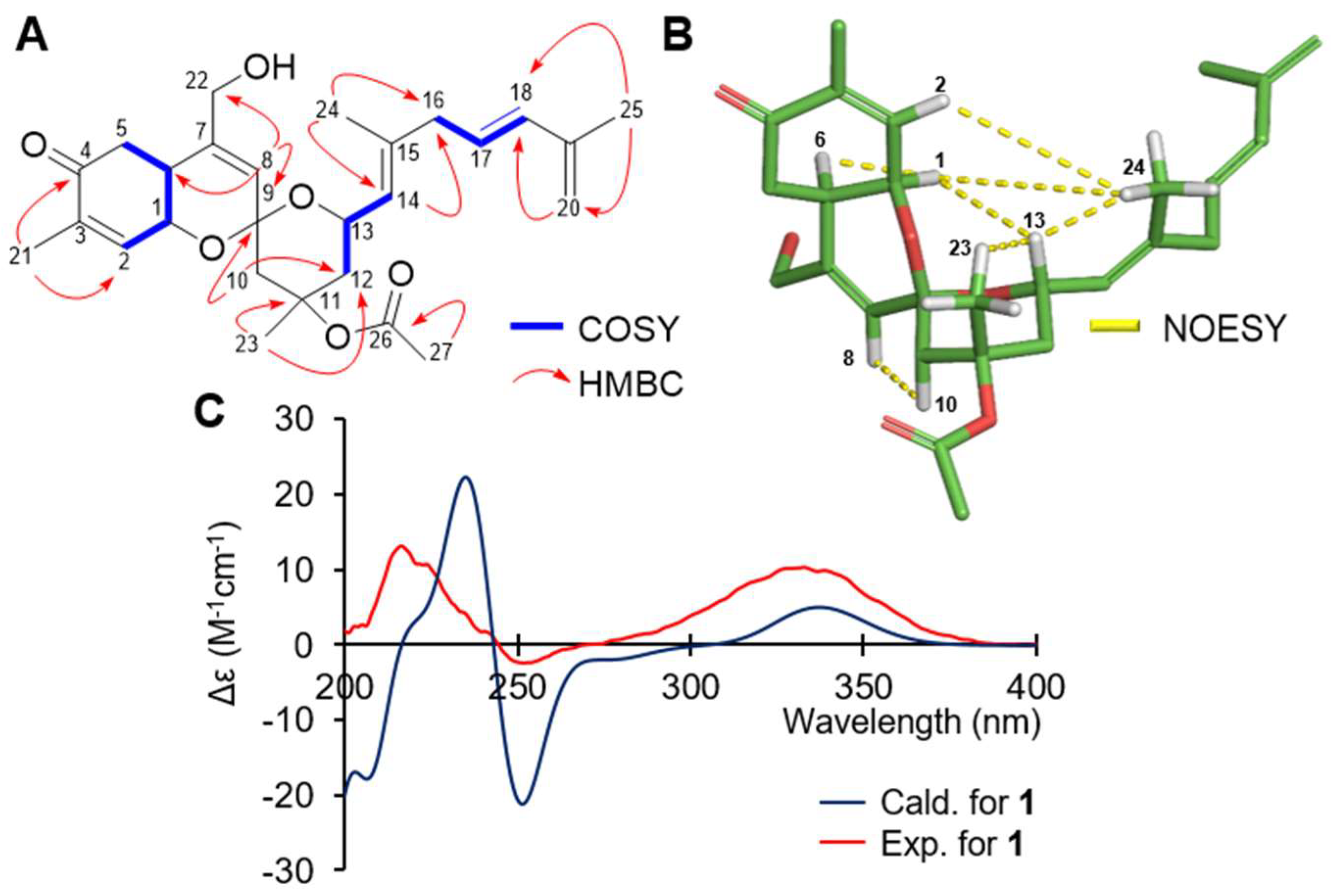
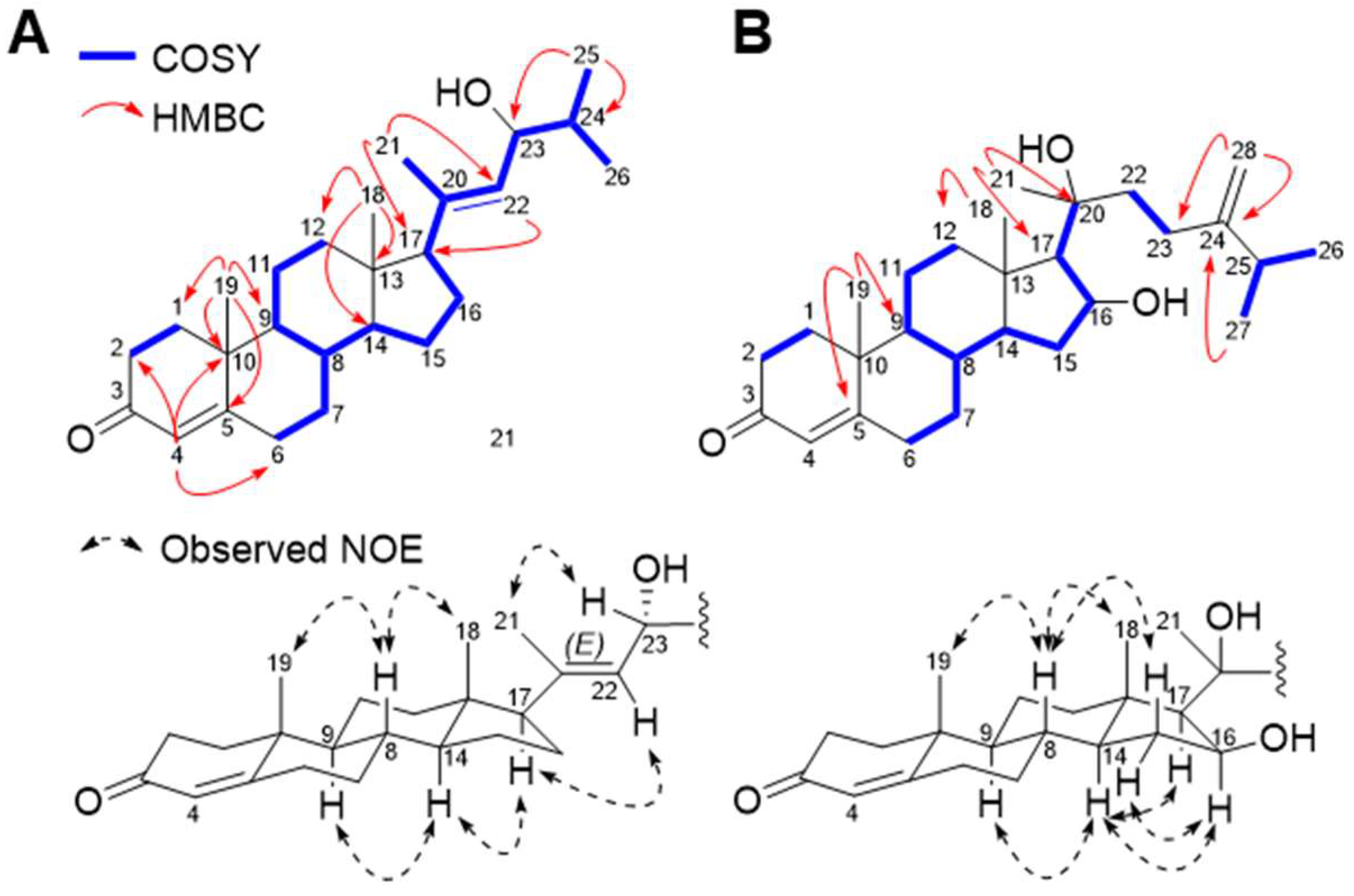
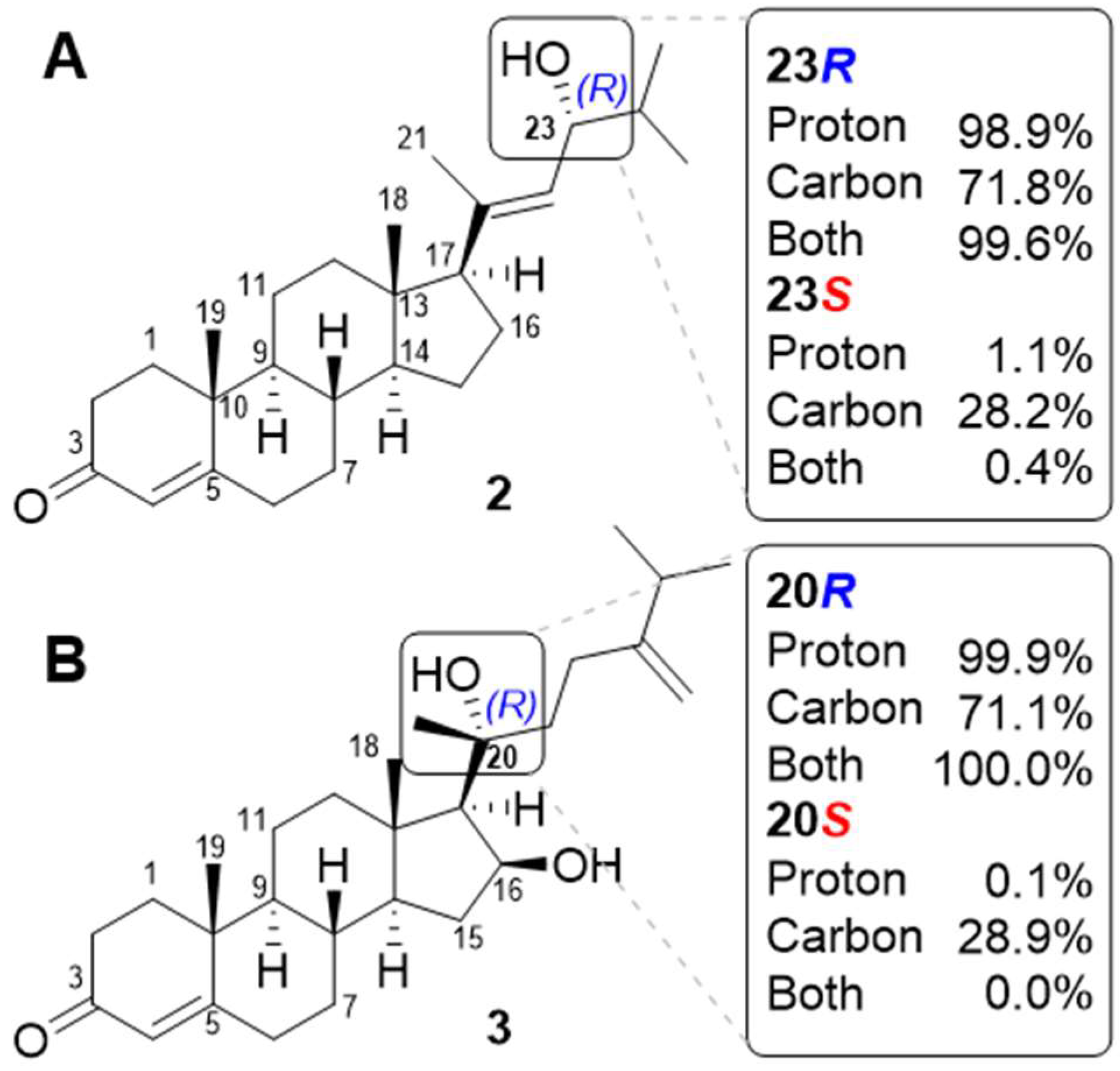
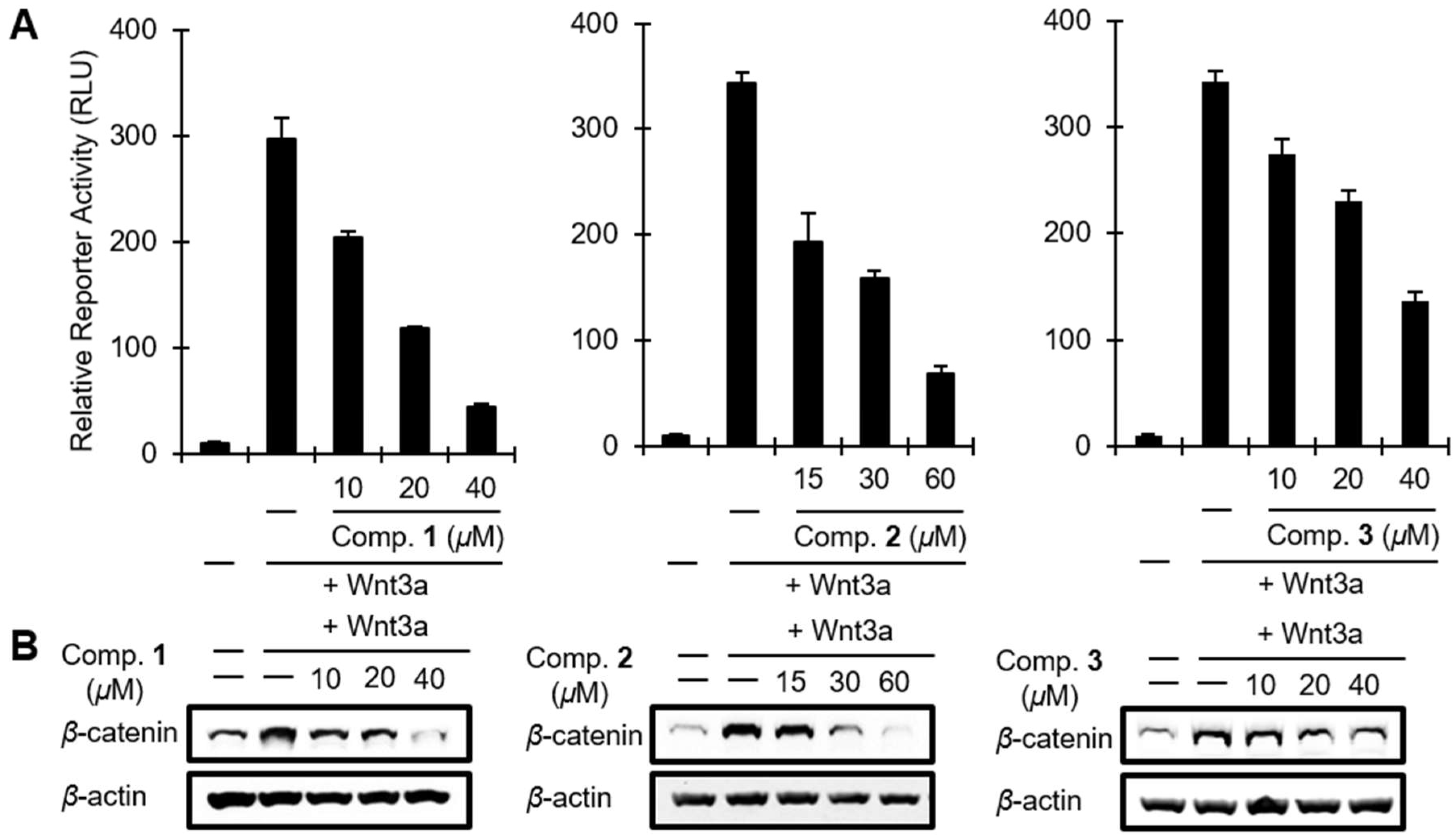
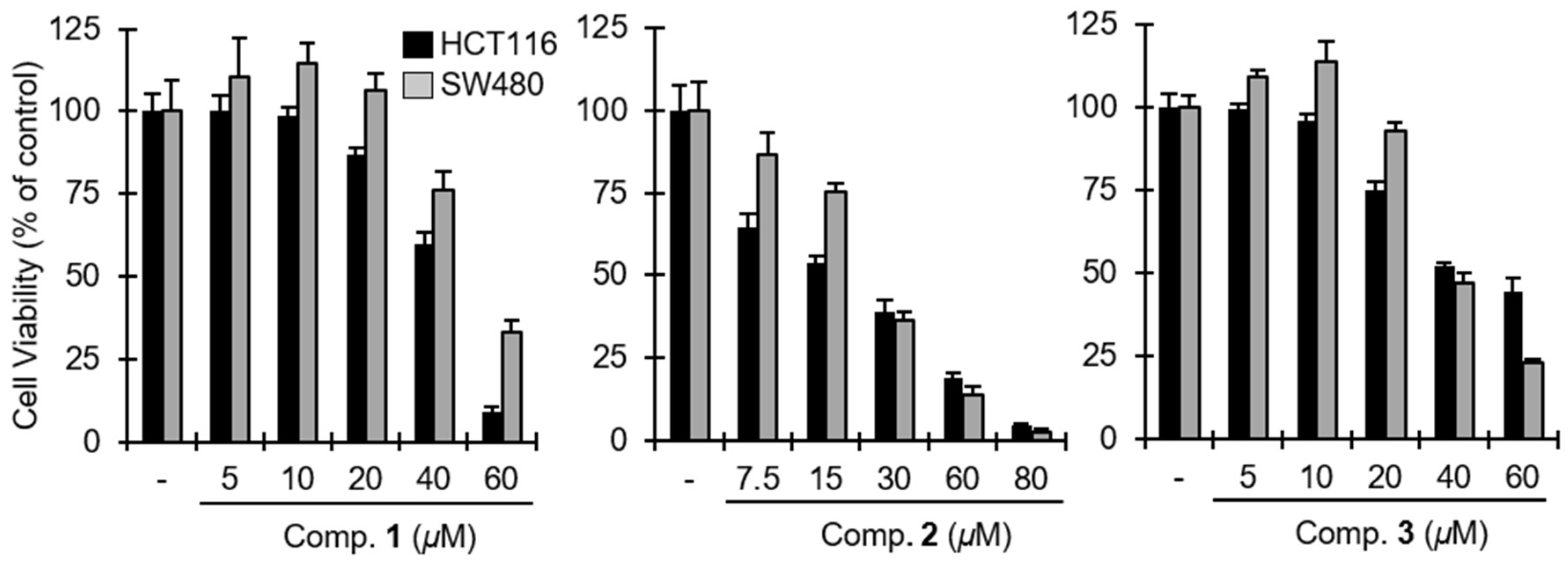
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Sponge Collection
3.3. Taxonomic Notes
3.4. Ecological Observations
3.5. Organic Extraction and Compound Isolation
3.6. Computational Details for ECD Simulation
3.7. Computational NMR Chemical Shifts Calculations for DP4 Analysis
3.8. Cell Culture and Reporter Assays
3.9. Western Blotting
3.10. Cell Viability Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- National Cancer Institute. Available online: http://seer.cancer.gov (accessed on 10 February 2018).
- Clevers, H.; Nusse, R. Wnt/β-catenin signaling and disease. Cell 2012, 149, 1192–1205. [Google Scholar] [CrossRef] [PubMed]
- Zhan, T.; Rindtorff, N.; Boutros, M. Wnt signaling in cancer. Oncogene 2017, 36, 1461–1473. [Google Scholar] [CrossRef] [PubMed]
- Fearnhead, N.S.; Britton, M.P.; Bodmer, W.F. The ABC of APC. Hum. Mol. Genet. 2001, 10, 721–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, P.J. β-catenin signaling and cancer. Bioessays 1999, 21, 1021–1030. [Google Scholar] [CrossRef]
- Takahashi, M.; Tsunoda, T.; Seiki, M.; Nakamura, Y.; Furukawa, Y. Identification of membrane-type matrix metalloproteinase-1 as a target of the β-catenin/Tcf4 complex in human colorectal cancers. Oncogene 2002, 21, 5861–5867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, T.C.; Sparks, A.B.; Rago, C.; Hermeking, H.; Zawel, L.; da Costa, L.T.; Morin, P.J.; Vogelstein, B.; Kinzler, K.W. Identification of c-MYC as a target of the APC pathway. Science 1998, 281, 1509–1512. [Google Scholar] [CrossRef] [PubMed]
- Tetsu, O.; McCormick, F. β-Catenin regulates expression of cyclin D1 in colon carcinoma cells. Nature 1999, 398, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Novellasdemunt, L.; Antas, P.; Li, V.S. Targeting Wnt signaling in colorectal cancer. A review in the theme: Cell signaling: Proteins, pathways and mechanisms. Am. J. Physiol. Cell Physiol. 2015, 309, C511–C521. [Google Scholar] [CrossRef] [PubMed]
- Thakur, R.; Mishra, D.P. Pharmacological modulation of β-catenin and its applications in cancer therapy. J. Cell. Mol. Med. 2013, 17, 449–456. [Google Scholar] [CrossRef] [PubMed]
- Gwak, J.; Oh, J.; Cho, M.; Bae, S.K.; Song, I.S.; Liu, K.H.; Jeong, Y.; Kim, D.E.; Chung, Y.H.; Oh, S. Galangin suppresses the proliferation of β-catenin response transcription-positive cancer cells by promoting adenomatous polyposis coli/Axin/glycogen synthase kinase-3β-independent β-catenin degradation. Mol. Pharmacol. 2011, 79, 1014–1022. [Google Scholar] [CrossRef] [PubMed]
- Hwang, I.H.; Oh, J.; Zhou, W.; Park, S.; Kim, J.H.; Chittiboyina, A.G.; Ferreira, D.; Song, G.Y.; Oh, S.; Na, M.; et al. Cytotoxic activity of rearranged drimane meroterpenoids against colon cancer cells via down-regulation of β-catenin expression. J. Nat. Prod. 2015, 78, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Abbas, S.; Kelly, M.; Bowling, J.; Sims, J.; Waters, A.; Hamann, M. Advancement into the arctic region for bioactive sponge secondary metabolites. Mar. Drugs 2011, 9, 2423–2437. [Google Scholar] [CrossRef] [PubMed]
- Molinski, T.F.; Dalisay, D.S.; Lievens, S.L.; Saludes, J.P. Drug development from marine natural products. Nat. Rev. Drug Discov. 2009, 8, 69–85. [Google Scholar] [CrossRef] [PubMed]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.G.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2017, 34, 235–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehbub, M.F.; Perkins, M.V.; Zhang, W.; Franco, C.M. New marine natural products from sponges (Porifera) of the order Dictyoceratida (2001 to 2012); a promising source for drug discovery, exploration and future prospects. Biotechnol. Adv. 2016, 34, 473–491. [Google Scholar] [CrossRef] [PubMed]
- Newman, D.J.; Cragg, G.M. Natural products as sources of new drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [PubMed]
- Bensemhoun, J.; Bombarda, I.; Aknin, M.; Vacelet, J.; Gaydou, E.M. Ptilomycalin D, a polycyclic guanidine alkaloid from the marine sponge Monanchora dianchora. J. Nat. Prod. 2007, 70, 2033–2035. [Google Scholar] [CrossRef] [PubMed]
- Guzii, A.G.; Makarieva, T.N.; Denisenko, V.A.; Dmitrenok, P.S.; Kuzmich, A.S.; Dyshlovoy, S.A.; Krasokhin, V.B.; Stonik, V.A. Monanchocidin: A new apoptosis-inducing polycyclic guanidine alkaloid from the marine sponge Monanchora pulchra. Org. Lett. 2010, 12, 4292–4295. [Google Scholar] [CrossRef] [PubMed]
- Hua, H.M.; Peng, J.; Fronczek, F.R.; Kelly, M.; Hamann, M.T. Crystallographic and NMR studies of antiinfective tricyclic guanidine alkaloids from the sponge Monanchora unguifera. Bioorg. Med. Chem. 2004, 12, 6461–6464. [Google Scholar] [CrossRef] [PubMed]
- Hua, H.M.; Peng, J.; Dunbar, D.C.; Schinazi, R.F.; de Castro Andrews, A.G.; Cuevas, C.; Garcia-Fernandez, L.F.; Kelly, M.; Hamann, M.T. Batzelladine alkaloids from the caribbean sponge Monanchora unguifera and the significant activities against HIV-1 and AIDS opportunistic infectious pathogens. Tetrahedron 2007, 63, 11179–11188. [Google Scholar] [CrossRef]
- Makarieva, T.N.; Ogurtsova, E.K.; Denisenko, V.A.; Dmitrenok, P.S.; Tabakmakher, K.M.; Guzii, A.G.; Pislyagin, E.A.; Es’kov, A.A.; Kozhemyako, V.B.; Aminin, D.L.; et al. Urupocidin A: a new, inducing iNOS expression bicyclic guanidine alkaloid from the marine sponge Monanchora pulchra. Org. Lett. 2014, 16, 4292–4295. [Google Scholar] [CrossRef] [PubMed]
- Takishima, S.; Ishiyama, A.; Iwatsuki, M.; Otoguro, K.; Yamada, H.; Omura, S.; Kobayashi, H.; van Soest, R.W.; Matsunaga, S. Merobatzelladines A and B, anti-infective tricyclic guanidines from a marine sponge Monanchora sp. Org. Lett. 2009, 11, 2655–2658. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Lee, T.G.; Patil, R.S.; Mun, B.; Yang, I.; Kim, H.; Hahn, D.; Won, D.H.; Lee, J.; Lee, Y.; et al. Monanchosterols A and B, bioactive bicyclo[4.3.1]steroids from a Korean sponge Monanchora sp. J. Nat. Prod. 2015, 78, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Mun, B.; Lee, Y.; Venkat Reddy, M.; Park, Y.; Lee, J.; Kim, H.; Hahn, D.; Chin, J.; Ekins, M.; et al. Bioactive sesterterpenoids from a Korean sponge Monanchora sp. J. Nat. Prod. 2013, 76, 170–177. [Google Scholar] [CrossRef] [PubMed]
- Kapustina, I.I.; Tabakmakher, K.M.; Makar’eva, T.N. Sterols from the toxin-containing far-east sponge Monanchora pulchra. Chem. Nat. Compd. 2012, 47, 1025–1027. [Google Scholar] [CrossRef]
- Santalova, E.A.; Makarieva, T.N.; Ponomarenko, L.P.; Denisenko, V.A.; Krasokhin, V.B.; Mollo, E.; Cimino, G.; Stonik, V.A. Sterols and related metabolites from five species of sponges. Biochem. Syst. Ecol. 2007, 35, 439–446. [Google Scholar] [CrossRef]
- Lambe, L.M. Sponges from the western coast of North America. Trans. R. Soc. Can. 1894, 12, 113–138. [Google Scholar]
- Smith, S.G.; Goodman, J.M. Assigning stereochemistry to single diastereoisomers by GIAO NMR calculation: the DP4 probability. J. Am. Chem. Soc. 2010, 132, 12946–12959. [Google Scholar] [CrossRef] [PubMed]
- Forestieri, R.; Merchant, C.E.; de Voogd, N.J.; Matainaho, T.; Kieffer, T.J.; Andersen, R.J. Alotaketals A and B, sesterterpenoids from the marine sponge Hamigera species that activate the cAMP cell signaling pathway. Org. Lett. 2009, 11, 5166–5169. [Google Scholar] [CrossRef] [PubMed]
- Rho, J.R.; Hwang, B.S.; Sim, C.J.; Joung, S.; Lee, H.Y.; Kim, H.J. Phorbaketals A, B, and C, sesterterpenoids with a spiroketal of hydrobenzopyran moiety isolated from the marine sponge Phorbas sp. Org. Lett. 2009, 11, 5590–5593. [Google Scholar] [CrossRef] [PubMed]
- Daoust, J.; Chen, M.; Wang, M.; Williams, D.E.; Garcia Chavez, M.A.; Wang, Y.A.; Merchant, C.E.; Fontana, A.; Kieffer, T.J.; Andersen, R.J. Sesterterpenoids isolated from a northeastern Pacific Phorbas sp. J. Org. Chem. 2013, 78, 8267–8273. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.; Rho, J.-R. A new sesterterpenoid showing anti-inflammatory effect from the marine sponge Haliclona species. J. Korean Magn. Resonance Soc. 2015, 19, 23–28. [Google Scholar] [CrossRef]
- Wang, M.; Tietjen, I.; Chen, M.; Williams, D.E.; Daoust, J.; Brockman, M.A.; Andersen, R.J. Sesterterpenoids isolated from the sponge Phorbas sp. Activate latent HIV-1 provirus expression. J. Org. Chem. 2016, 81, 11324–11334. [Google Scholar] [CrossRef] [PubMed]
- Woo, J.K.; Kim, C.K.; Kim, S.H.; Kim, H.; Oh, D.C.; Oh, K.B.; Shin, J. Gombaspiroketals A-C, sesterterpenes from the sponge Clathria gombawuiensis. Org. Lett. 2014, 16, 2826–2829. [Google Scholar] [CrossRef] [PubMed]
- Byun, M.R.; Lee, C.H.; Hwang, J.H.; Kim, A.R.; Moon, S.A.; Sung, M.K.; Roh, J.R.; Hwang, E.S.; Hong, J.H. Phorbaketal A inhibits adipogenic differentiation through the suppression of PPARγ-mediated gene transcription by TAZ. Eur. J. Pharmacol. 2013, 718, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Seo, Y.J.; Lee, K.T.; Rho, J.R.; Choi, J.H. Phorbaketal A, isolated from the marine sponge Phorbas sp., exerts its anti-inflammatory effects via NF-κB inhibition and heme oxygenase-1 activation in lipopolysaccharide-stimulated macrophages. Mar. Drugs 2015, 13, 7005–7019. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; Gwak, J.; Cho, M.; Song, T.; Won, J.; Kim, D.E.; Shin, J.G.; Oh, S. Hexachlorophene inhibits Wnt/β-catenin pathway by promoting Siah-mediated beta-catenin degradation. Mol. Pharmacol. 2006, 70, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Gwak, J.; Lee, J.H.; Chung, Y.H.; Song, G.Y.; Oh, S. Small molecule-based promotion of PKCα-mediated β-catenin degradation suppresses the proliferation of CRT-positive cancer cells. PLoS ONE 2012, 7, e46697. [Google Scholar] [CrossRef] [PubMed]
- Van Soest, R.W.M.; Boury-Esnault, N.; Hooper, J.N.A.; Rützler, K.; de Voogd, N.J.; Alvarez, B.; Hajdu, E.; Pisera, A.B.; Manconi, R.; Schönberg, C.; et al. World porifera database. Monanchora pulchra (lambe, 1895). Available online: http://www.marinespecies.org/porifera/porifera.php?p=taxdetails&id=169022 (accessed on 9 April 2018).
- Stone, R.P.; Lehnert, H.; Reiswig, H. A guide to the deep-water sponges of the Aleutian Island Archipelago. NOAA Professional Paper NMFS 2011, 12, 1–187. [Google Scholar]
- Dignam, J.D.; Lebovitz, R.M.; Roeder, R.G. Accurate transcription initiation by RNA polymerase II in a soluble extract from isolated mammalian nuclei. Nucleic Acids Res. 1983, 11, 1475–1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, E.; Frenkel, B. Glucocorticoids inhibit the transcriptional activity of LEF/TCF in differentiating osteoblasts in a glycogen synthase kinase-3β-dependent and -independent manner. J. Biol. Chem. 2005, 280, 2388–2394. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | δC, type | δH (J in Hz) | ||
|---|---|---|---|---|
| 1 | 63.0 | CH | 4.47 | dd, 4.7, 3.0 |
| 2 | 139.4 | CH | 6.62 | dd, 5.9, 1.6 |
| 3 | 139.5 | C | ||
| 4 | 198.8 | C | ||
| 5 | 38.0 | CH2 | 2.59 | dd, 13.7, 2.6 |
| 2.49 | m | |||
| 6 | 33.3 | CH | 2.54 | m |
| 7 | 142.5 | C | ||
| 8 | 125.1 | CH | 5.66 | s |
| 9 | 96.8 | C | ||
| 10 | 44.5 | CH2 | 2.32 | dd, 13.4, 1.5 |
| 1.87 | m a | |||
| 11 | 80.1 | C | ||
| 12 | 42.5 | CH2 | 2.15 | brd, 13.2 |
| 1.76 | m a | |||
| 13 | 65.1 | CH | 4.65 | m |
| 14 | 125.2 | CH | 5.25 | d, 7.6 |
| 15 | 139.0 | C | ||
| 16 | 42.9 | CH2 | 2.80 | m |
| 17 | 127.5 | CH | 5.61 | m |
| 18 | 135.2 | CH | 6.15 | d, 15.6 |
| 19 | 142.1 | C | ||
| 20 | 115.3 | CH2 | 4.90 | d, 5.2 |
| 21 | 16.1 | CH3 | 1.86 | s |
| 22 | 63.8 | CH2 | 4.12 | m |
| 23 | 24.6 | CH3 | 1.78 | s |
| 24 | 17.0 | CH3 | 1.74 | s |
| 25 | 18.9 | CH3 | 1.84 | s |
| 26 | 170.3 | C | ||
| 27 | 22.6 | CH3 | 1.96 | s |
| No. | 2 | 3 | ||||||
|---|---|---|---|---|---|---|---|---|
| δC, type | δH (J in Hz) | δC, type | δH (J in Hz) | |||||
| 1 | 35.8 | CH2 | 2.02 | ddd, 13.4, 5.1, 3.2 | 35.8 | CH2 | 2.01 | m |
| 1.67 | m a | 1.68 | m | |||||
| 2 | 33.1 | CH2 | 2.40 | dd, 14.6, 5.1 | 34.1 | CH2 | 2.41 | m |
| 2.28 | ddd, 14.6, 4.3, 2.5 | 2.34 | m | |||||
| 3 | 199.8 | C | 199.7 | C | ||||
| 4 | 124.0 | CH | 5.73 | s | 124.1 | CH | 5.74 | s |
| 5 | 171.6 | C | 171.1 | C | ||||
| 6 | 34.1 | CH2 | 2.43 | m a | 32.9 | CH2 | 2.40 | m |
| 2.36 | dt, 17.0, 3.8 | 2.27 | m | |||||
| 7 | 32.1 | CH2 | 1.84 | m a | 32.0 | CH2 | 1.83 | m |
| 1.03 | dd, 12.7, 3.3 | 1.00 | m | |||||
| 8 | 35.9 | CH | 1.52 | m a | 34.7 | CH | 1.65 | m |
| 9 | 54.2 | CH | 0.94 | m a | 54.0 | CH | 0.93 | m |
| 10 | 38.8 | C | 38.7 | C | ||||
| 11 | 21.1 | CH2 | 1.58 | m a | 20.9 | CH2 | 1.52 | m |
| 1.42 | dd, 13.0, 4.0 | |||||||
| 12 | 38.5 | CH2 | 1.84 | m a | 40.4 | CH2 | 2.17 | m |
| 1.17 | m a | 1.17 | m | |||||
| 13 | 44.0 | C | 43.1 | C | ||||
| 14 | 55.5 | CH | 1.08 | m a | 53.9 | CH | 0.87 | m |
| 15 | 24.4 | CH2 | 1.73 | m a | 37.5 | CH2 | 2.27 | m |
| 1.23 | m a | 1.34 | m | |||||
| 16 | 24.8 | CH2 | 1.84 | m a | 74.2 | CH | 4.65 | m |
| 17 | 59.1 | CH | 2.08 | t, 9.5 | 60.6 | CH | 1.25 | m |
| 18 | 13.4 | CH3 | 0.63 | s | 15.1 | CH3 | 1.21 | s |
| 19 | 17.6 | CH3 | 1.18 | s | 17.5 | CH3 | 1.21 | s |
| 20 | 138.5 | C | 76.5 | C | ||||
| 21 | 17.8 | CH3 | 1.70 | s | 26.7 | CH3 | 1.32 | s |
| 22 | 128.1 | CH | 5.24 | d, 8.8 | 42.6 | CH2 | 1.68 | m |
| 23 | 73.7 | CH | 4.15 | dd, 8.8, 6.7 | 28.7 | CH2 | 2.03 | m |
| 24 | 34.7 | CH | 1.70 | m a | 156.3 | C | ||
| 25 | 18.5 | CH3 | 0.96 | d, 6.7 | 33.9 | CH | 2.23 | m |
| 26 | 18.3 | CH3 | 0.88 | d, 6.7 | 22.1 | CH3 | 1.03 | d, 7.0 |
| 27 | 22.1 | CH3 | 1.03 | d, 7.0 | ||||
| 28 | 106.5 | CH2 | 4.75 | brs | ||||
| 4.68 | s | |||||||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, H.B.; Tuan, N.Q.; Oh, J.; Son, Y.; Hamann, M.T.; Stone, R.; Kelly, M.; Oh, S.; Na, M. Sesterterpenoid and Steroid Metabolites from a Deep-Water Alaska Sponge Inhibit Wnt/β-Catenin Signaling in Colon Cancer Cells. Mar. Drugs 2018, 16, 297. https://doi.org/10.3390/md16090297
Park HB, Tuan NQ, Oh J, Son Y, Hamann MT, Stone R, Kelly M, Oh S, Na M. Sesterterpenoid and Steroid Metabolites from a Deep-Water Alaska Sponge Inhibit Wnt/β-Catenin Signaling in Colon Cancer Cells. Marine Drugs. 2018; 16(9):297. https://doi.org/10.3390/md16090297
Chicago/Turabian StylePark, Hyun Bong, Nguyen Quoc Tuan, Joonseok Oh, Younglim Son, Mark T. Hamann, Robert Stone, Michelle Kelly, Sangtaek Oh, and MinKyun Na. 2018. "Sesterterpenoid and Steroid Metabolites from a Deep-Water Alaska Sponge Inhibit Wnt/β-Catenin Signaling in Colon Cancer Cells" Marine Drugs 16, no. 9: 297. https://doi.org/10.3390/md16090297