
Functional Recombinants Designed from a Fetuin/Asialofetuin-Specific Marine Algal Lectin, Rhodobindin

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Cloning of Recombinant Lectins
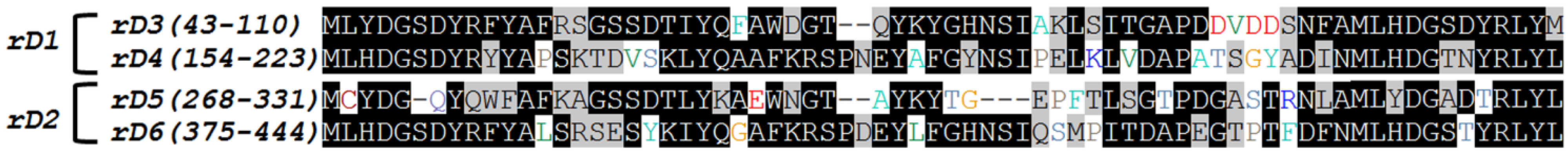

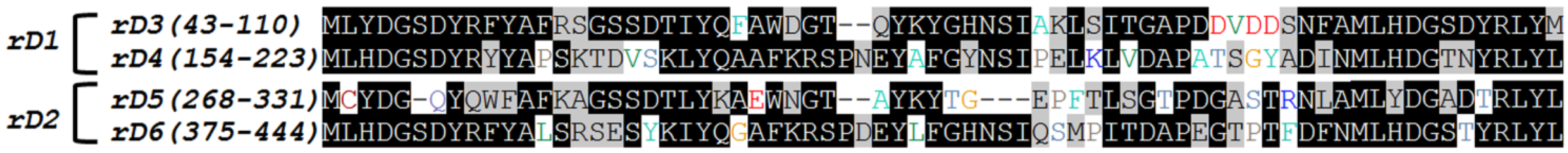{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence(5′-3′) | Sequences Amplified |
|---|---|---|
| Rhodo(1)-EcoRI-F1 | GGATCCGAATTCATGTCTCGCTCATTCAAC | Whole, rD1, rD3 (forward) |
| Rhodo(228)-XhoI-R1 | TCGGCGTCGCGCTGCCCCTCGAGCACCACC | rD1, rD4 (reverse) |
| Rhodo(229)-EcoRI-F1 | GGATCCGAATTCACGCTGCGCCAGTTCGGC | rD2, rD5 (forward) |
| Rhodo(443)-XhoI-R1 | CGATCCAGGCCGTTGCCCTCGAGCACCACC | Whole, rD2, rD6 (reverse) |
| Rhodo(116)-XhoI-R1 | GCAAGCGCGCGACGGACCTCGAGCACCACC | rD3 (reverse) |
| Rhodo(113)-EcoRI-F1 | GGATCCGAATTCCGCGCGACGGACGCGCTC | rD4 (forward) |
| Rhodo(337)-XhoI-R1 | TGTCGCGCACGGGCGCGCTCGAGCACCACC | rD5 (reverse) |
| Rhodo(334)-EcoRI-F1 | GGATCCGAATTCCGCACGGGCGCGTCGCTG | rD6 (forward) |
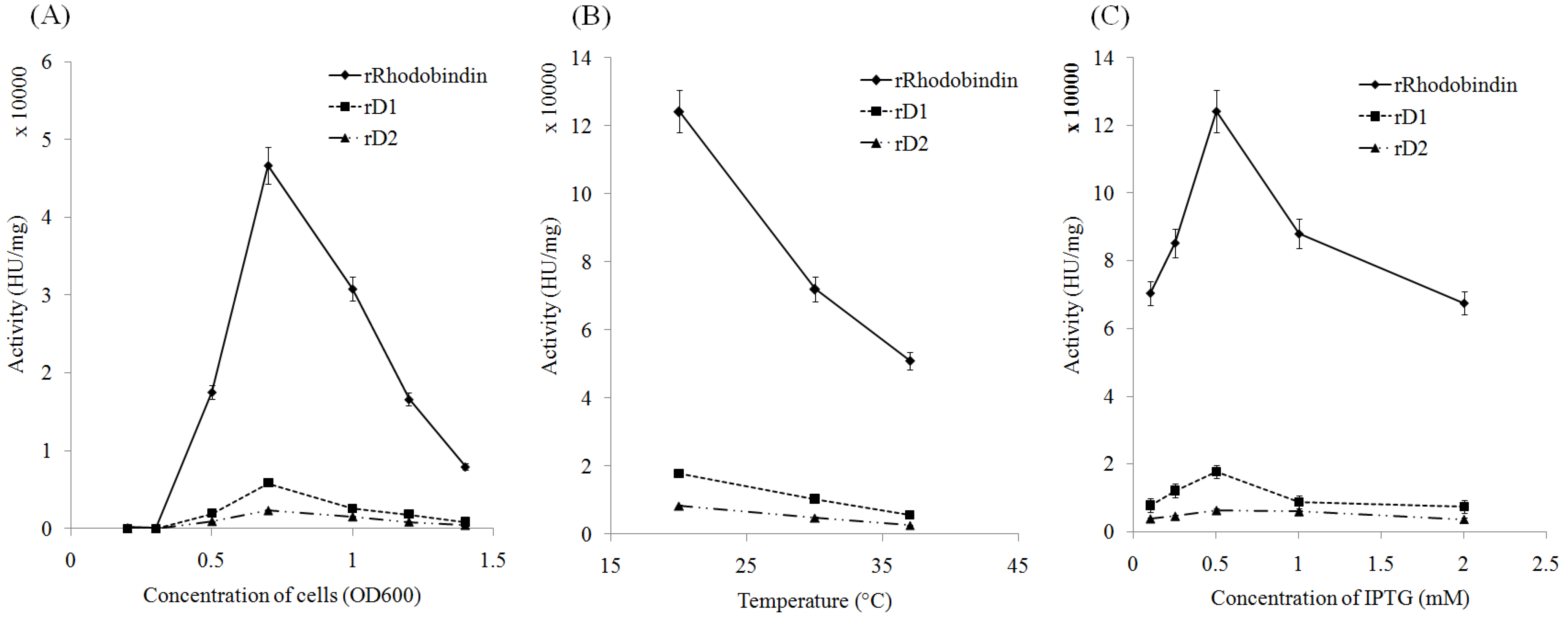
2.2. Expression of Recombinant Rhodobindin
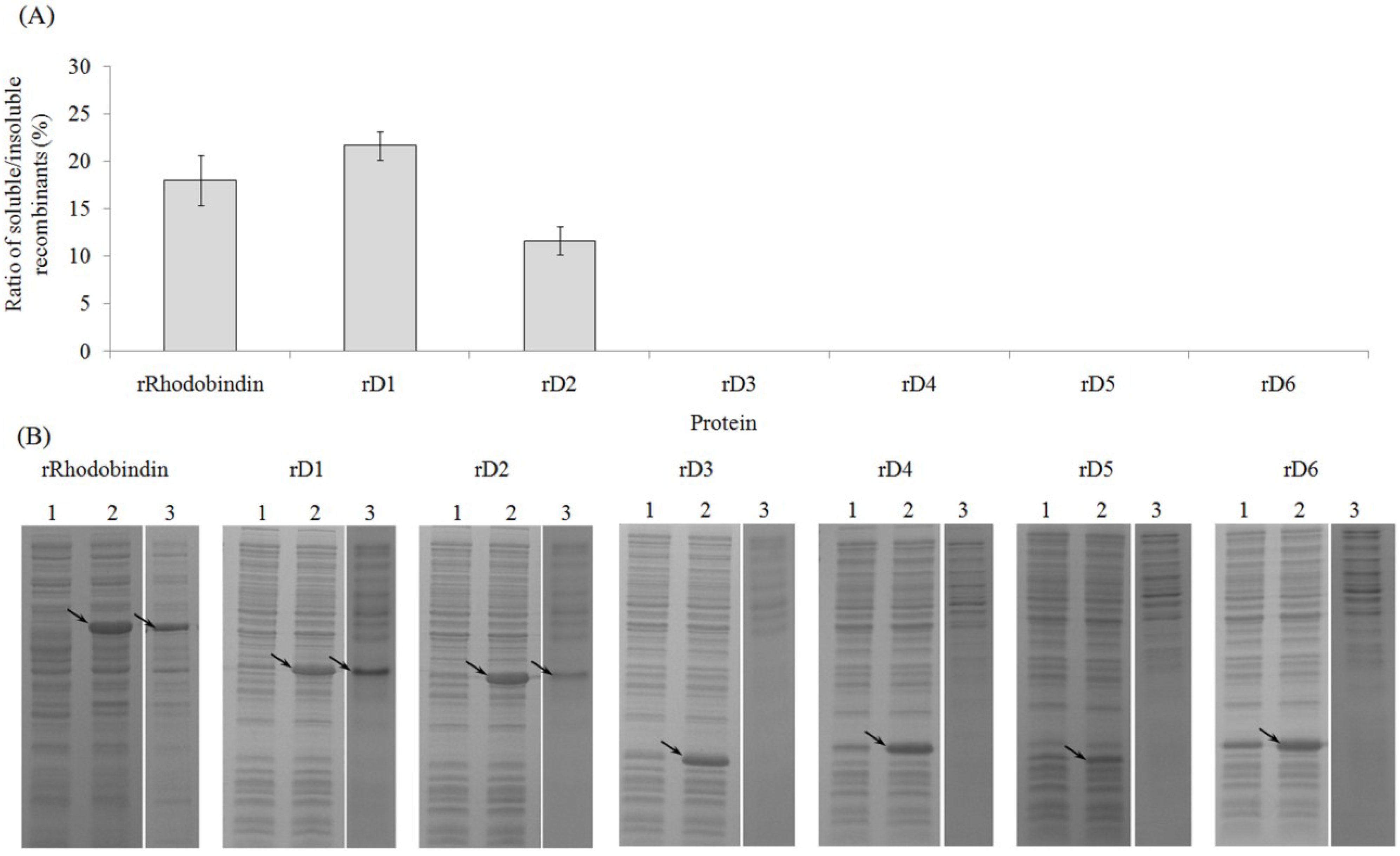
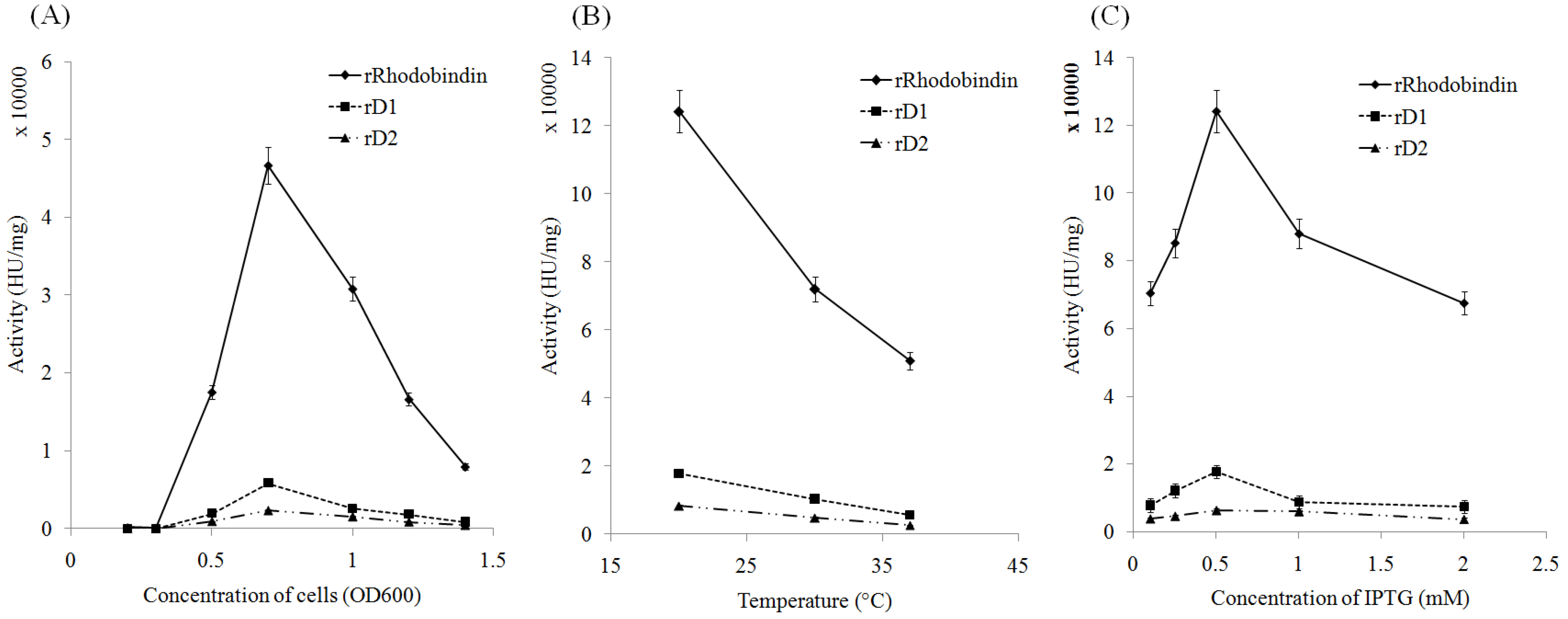
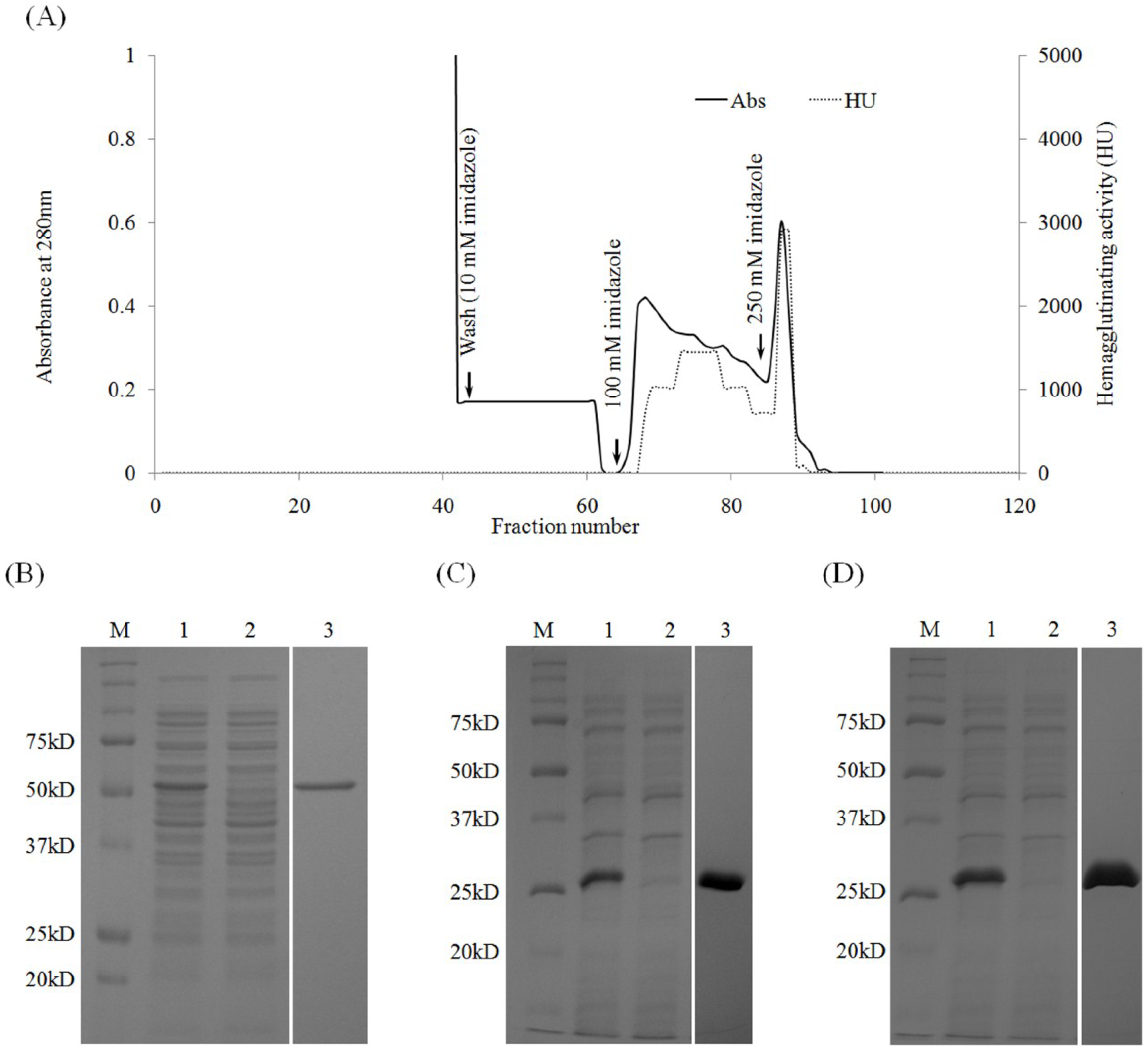
2.3. Purification of Recombinant Lectins

| Total Protein (mg) | Protein Concentration (mg·mL−1) | Total Activity (titer) | Specific Activity (titer·mg−1) | Purification Fold | Percentage of Recovery | |
|---|---|---|---|---|---|---|
| Crude extract | 26 | 0.65 | 3,276,800 | 126,030 | 1 | 100 |
| Affinity chromatography | 0.66 | 0.041 | 2,621,440 | 3,971,878 | 31 | 80 |
| Name | Molecular Weight (Kda) § | Yield (mg per 1L Culture) | Minimum Concentration (µg/mL) ¶ | Minimum Mole Concentration |
|---|---|---|---|---|
| Full-rRhodobindin | 50 | 16.5 | 0.04 | 0.8 nM |
| rD1 | 25 | 12.1 | 0.31 | 12.4 nM |
| rD2 | 25 | 13.3 | 0.63 | 25.2 nM |
| rD1 + rD2 (mixed) £ | - | - | 0.44 | - |
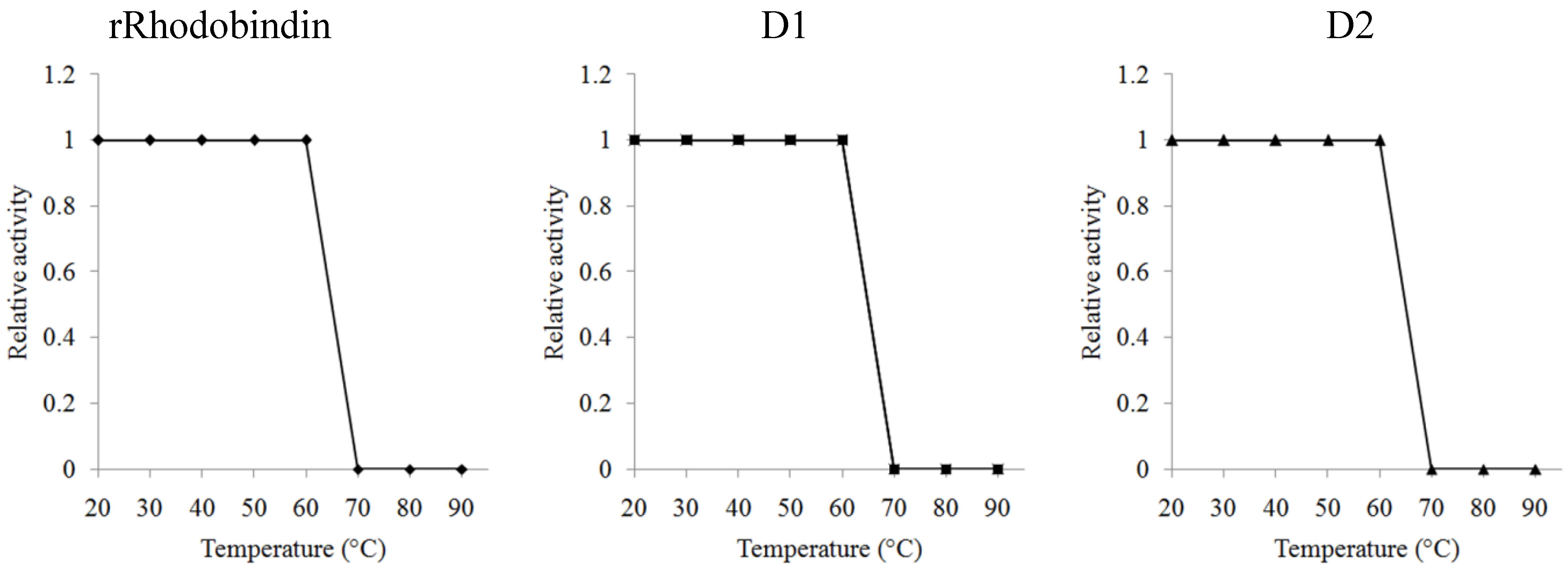
2.4. Carbohydrate Specificity and Heat-Stability of Recombinant Lectins
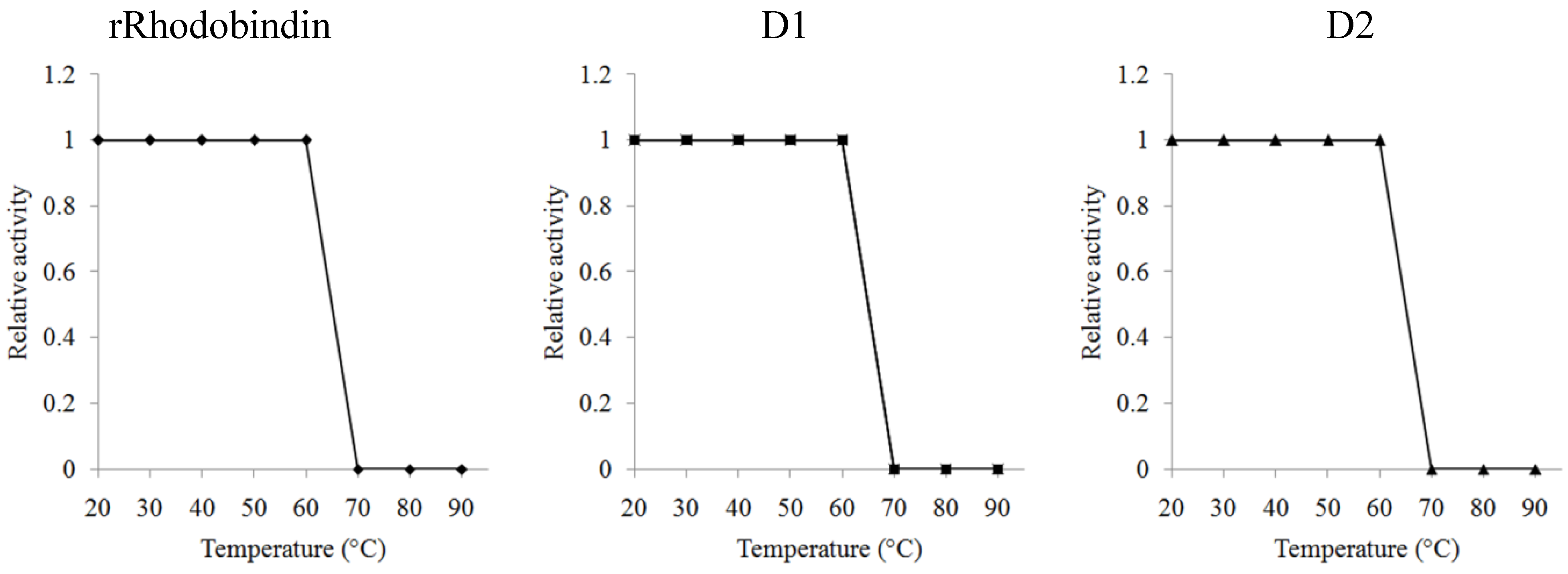
3. Discussion
4. Experimental Section
4.1. Plant and Culture Conditions
4.2. Cloning and Design of Recombinant Lectins
4.3. Optimization of rRhodobindin Expression
4.4. Purification of Recombinant Lectins
4.5. Hemagglutinating Activity Assay and Determination of Carbohydrate Specificity
4.6. Effect of Temperature and Divalent Cations on the Agglutinating Activity of rRhodobindin
4.7. Mass Spectrometry
5. Conclusions
Supplementary Files
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zheng, T.; Peelen, D.; Smith, L.M. Lectin arrays for profiling cell surface carbohydrate expression. J. Am. Chem. Soc. 2005, 127, 9982–9983. [Google Scholar] [CrossRef] [PubMed]
- Pytela, R.; Pierschbacher, M.D.; Ruoslahti, E. Identification and isolation of a 140 kd cell surface glycoprotein with properties expected of a fibronectin receptor. Cell 1985, 40, 191–198. [Google Scholar] [CrossRef] [PubMed]
- Bies, C.; Lehr, C.; Woodley, J.F. Lectin-mediated drug targeting: History and applications. Adv. Drug Deliv. Rev. 2004, 56, 402–435. [Google Scholar] [CrossRef]
- Sancho, D.; Mourão-Sá, D.; Joffre, O.P.; Schulz, O.; Rogers, N.C.; Pennington, D.J.; Carlyle, J.R.; Sousa, C.R. Tumor therapy in mice via antigen targeting to a novel, DC-restricted C-type lectin. J. Clin. Investig. 2008, 118, 2098–2110. [Google Scholar] [CrossRef] [PubMed]
- Silva, L.C.N.; Correia, M.T.S. Plant lectins and Toll-like receptors: Implications for therapy of microbial infections. Front. Microbiol. 2014, 5, 1–3. [Google Scholar] [PubMed]
- Liu, B.; Bian, H.J.; Bao, J.K. Plant lectins: Potential antineoplastic drugs from bench to clinic. Cancer Lett. 2010, 287, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Fu, L.L.; Zhou, C.C.; Yao, S.; Yu, J.Y.; Liu, B.; Bao, J.K. Plant lectins: Targeting programmed cell death pathways as antitumor agents. Int. J. Biochem. Cell Biol. 2011, 43, 1442–1449. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Teixeira, J.A.; Domingues, L. Recombinant production of plant lectins in microbial systems for biomedical application—The frutalin case study. Front. Plant Sci. 2014, 5, 390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, S.K.; Ng, T.B. Lectins: Production and practical applications. Appl. Microbiol. Biotechnol. 2011, 89, 45–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.H.; Fritz, L. Ultrastructure and cytochemistry of early spermatangial development in Antithamnion nipponicum (Ceramiaceae, Rhodophyta). J. Phycol. 1993, 29, 797–805. [Google Scholar] [CrossRef]
- Kim, G.H.; Kim, S.H. The role of F-actin during fertilization in the red alga Aglaothamnion oosumiense (Rhodophyta). J. Phycol. 1999, 35, 806–814. [Google Scholar] [CrossRef]
- Han, J.W.; Klochkova, T.A.; Shim, J.B.; Yoon, K.; Kim, G.H. Isolation and characterization of a sex-specific lectin in a marine red alga, Aglaothamniono osumiense Itono. Appl. Environ. Microbiol. 2012, 78, 7283–7289. [Google Scholar] [CrossRef] [PubMed]
- Shim, E.; Shim, J.; Klochkova, T.A.; Han, J.W.; Kim, G.H. Purification of a sex-specific lectin involved in gamete binding of Aglaothamnion callophyllidicola (Rhodophyta). J. Phycol. 2012, 48, 916–924. [Google Scholar] [CrossRef]
- Hara, T.; Aramaki, Y.; Takada, S.; Koike, S.; Tsuchiya, S. Receptor-mediated transfer of pSV2CAT DNA to mouse liver cells using asialofetuin-labeled liposomes. Gene Ther. 1995, 2, 784–788. [Google Scholar] [PubMed]
- Pal, D.; Dasgupta, S.; Kundu, R.; Maitra, S.; Das, G.; Mukhopadhyay, S.; Ray, S.; Majumdar, S.S.; Bhattacharya, S. Fetuin-A acts as an endogenous ligand of TLR4 to promote lipid-induced insulin resistance. Nat. Med. 2012, 18, 1279–1285. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Jose, A.; Teixeira, J.A.; Domingues, L. Recombinant lectins: An array of tailor-made glycan-interaction biosynthetic tools. Crit. Rev. Biotechnol. 2013, 33, 66–80. [Google Scholar] [CrossRef] [PubMed]
- Francis, D.M.; Page, R. Strategies to optimize protein expression in E. coli. Curr. Protoc. Protein Sci. 2010, 61, 1–29. [Google Scholar]
- Vallejo, L.F.; Rinas, L. Strategies for the recovery of active proteins through refolding of bacterial inclusion body proteins. Microb. Cell Fact. 2004, 3, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Berlec, A.; Strukelj, B. Current state and recent advances in biopharmaceutical production in Escherichia coli, yeasts and mammalian cells. J. Ind. Microbiol. Biotechnol. 2013, 40, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liao, X.; Chen, G.; Yap, Y.; Zhang, X. Cloning, expression and purification of Microcystis viridis lectin in Escherichia coli. Mol. Biotechnol. 2011, 47, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, C.; Costa, S.; Teixeira, J.A.; Domingues, L. cDNA cloning and functional expression of the α-d-Galactose-Binding lectin frutalin in Escherichia coli. Mol. Biotechnol. 2009, 43, 212–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Zhang, X. Recombinant Microcystis viridis lectin as a potential anticancer agent. Pharmazie 2010, 65, 922–923. [Google Scholar] [PubMed]
- Kwon, M.; Jeong, S.; Lee, K.H.; Park, Y.K.; Yu, J. Mimicry of tandem repeat peptides against cell surface carbohydrate. J. Am. Chem. Soc. 2002, 124, 13996–13997. [Google Scholar] [CrossRef] [PubMed]
- Sharon, N.; Lis, H. Lectins: From hemagglutinins to biological recognition molecules. A historical overview. Glycobiology 2004, 14, 53R–62R. [Google Scholar] [CrossRef] [PubMed]
- Arata, Y.; Hirabayashi, J.; Kasai, K. The two lectin domains of the tandem-repeat 32-kDa Galectin of the nematode Caenorhabditis elegans have different binding properties. Studies with recombinant protein. J. Biochem. 1997, 121, 1002–1009. [Google Scholar] [CrossRef] [PubMed]
- Arata, Y.; Hirabayashi, J.; Kasai, K. Sugar binding properties of the two lectin domains of the tandem repeat-type Galectin LEC-1 (N32) of Caenorhabditis elegans. J. Biol. Chem. 2001, 276, 3068–3077. [Google Scholar] [CrossRef] [PubMed]
- Lehr, C.M. Lectin-mediated drug delivery: The second generation of bioadhesives. J. Control. Release 2000, 65, 19–29. [Google Scholar] [CrossRef] [PubMed]
- Sharon, N. Lectins: Carbohydrate-specific reagents and biological recognition molecules. J. Biol. Chem. 2007, 282, 2753–2764. [Google Scholar] [CrossRef] [PubMed]
- Woodley, J.F.; Naisbett, F. The potential of lectins for delaying the intestinal transit of drugs. Proc. Int. Symp. Control. Release Bioact. Mater. 1988, 15, 125–126. [Google Scholar]
- Gabor, F.; Bogner, E.; Weissenboeck, A.; Wirthet, M. The lectin–cell interaction and its implications to intestinal lectin-mediated drug delivery. Adv. Drug Deliv. Rev. 2004, 56, 459–480. [Google Scholar] [CrossRef] [PubMed]
- Gavrovic-Jankulovic, M.; Prodanovic, R. Drug delivery: Plant lectins as bioadhesive drug delivery systems. J. Biomater. Nanobiotechnol. 2011, 2, 614–621. [Google Scholar] [CrossRef]
- Han, J.W.; Jung, M.G.; Kim, M.J.; Yoon, K.S.; Lee, K.P.; Kim, G.H. Purification and characterization of a d-mannose specific lectin from the green marine alga, Bryopsis Plumosa. Phycol. Res. 2010, 58, 143–150. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, J.W.; Jung, M.G.; Shim, E.Y.; Shim, J.B.; Kim, Y.M.; Kim, G.H. Functional Recombinants Designed from a Fetuin/Asialofetuin-Specific Marine Algal Lectin, Rhodobindin. Mar. Drugs 2015, 13, 2183-2195. https://doi.org/10.3390/md13042183
Han JW, Jung MG, Shim EY, Shim JB, Kim YM, Kim GH. Functional Recombinants Designed from a Fetuin/Asialofetuin-Specific Marine Algal Lectin, Rhodobindin. Marine Drugs. 2015; 13(4):2183-2195. https://doi.org/10.3390/md13042183
Chicago/Turabian StyleHan, Jong Won, Min Gui Jung, Eun Young Shim, Jun Bo Shim, Young Min Kim, and Gwang Hoon Kim. 2015. "Functional Recombinants Designed from a Fetuin/Asialofetuin-Specific Marine Algal Lectin, Rhodobindin" Marine Drugs 13, no. 4: 2183-2195. https://doi.org/10.3390/md13042183
APA StyleHan, J. W., Jung, M. G., Shim, E. Y., Shim, J. B., Kim, Y. M., & Kim, G. H. (2015). Functional Recombinants Designed from a Fetuin/Asialofetuin-Specific Marine Algal Lectin, Rhodobindin. Marine Drugs, 13(4), 2183-2195. https://doi.org/10.3390/md13042183