
Anticancer Activity of Sea Cucumber Triterpene Glycosides

 ,
,
Abstract
:1. Introduction
2. Anticancer Activity
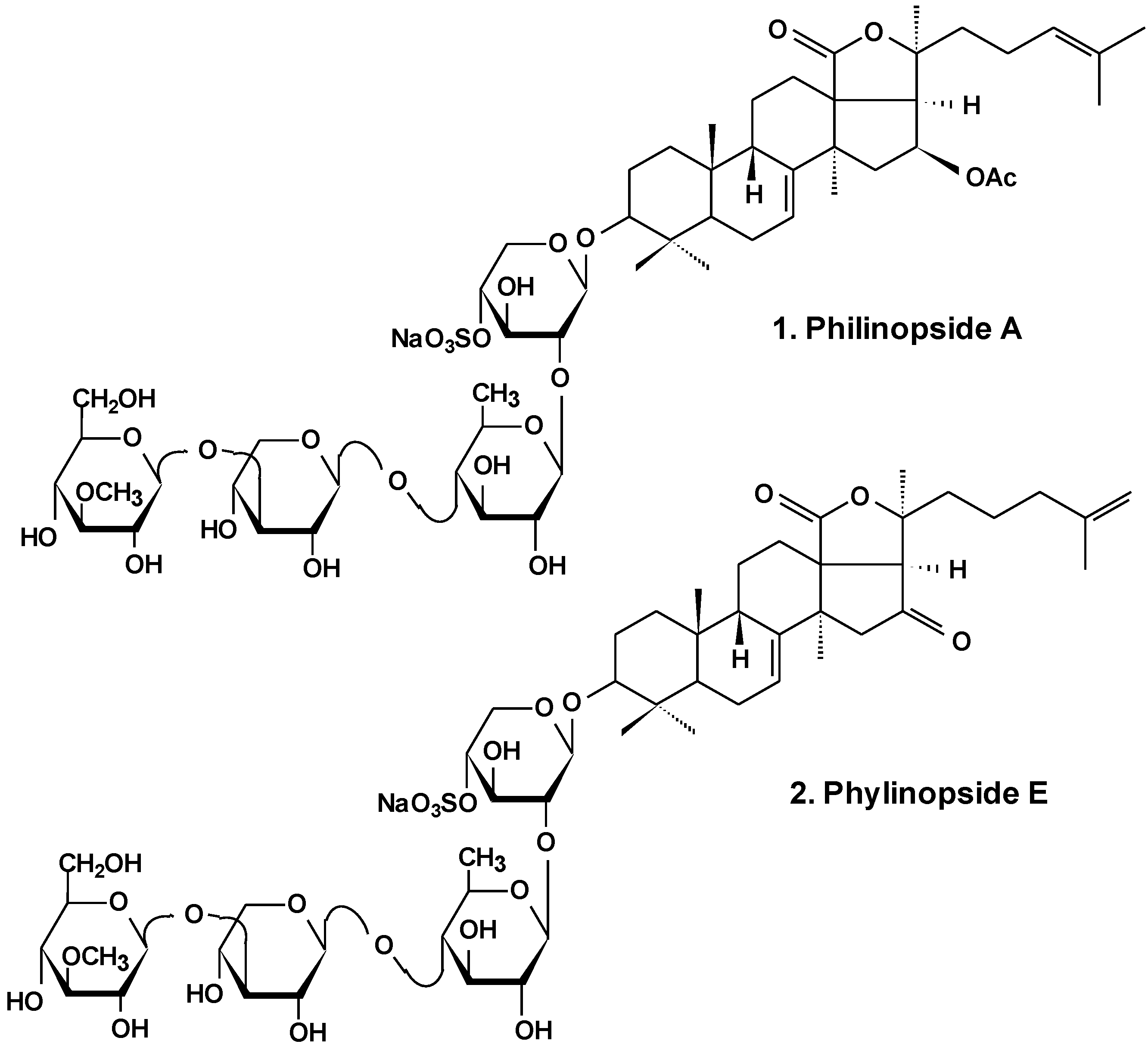

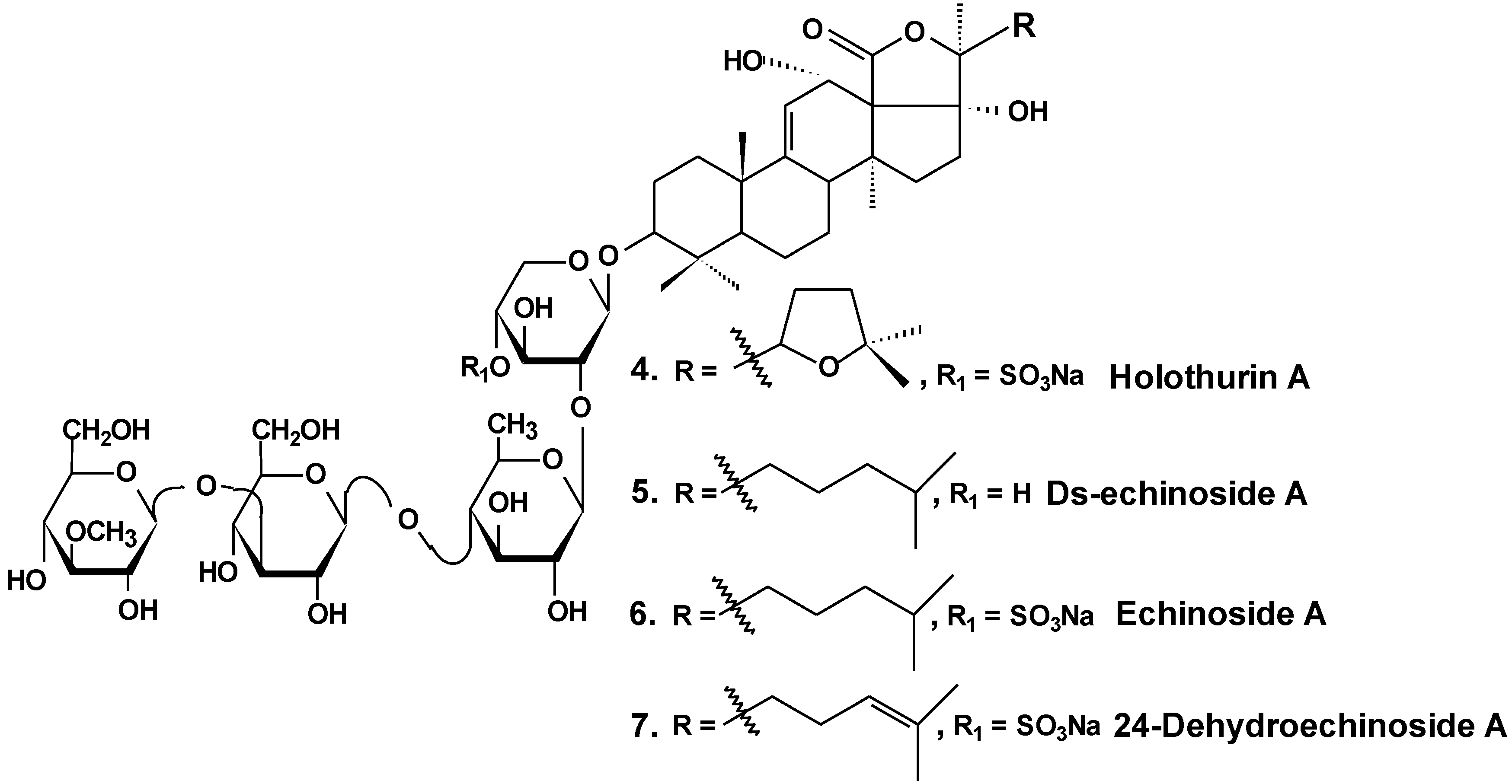
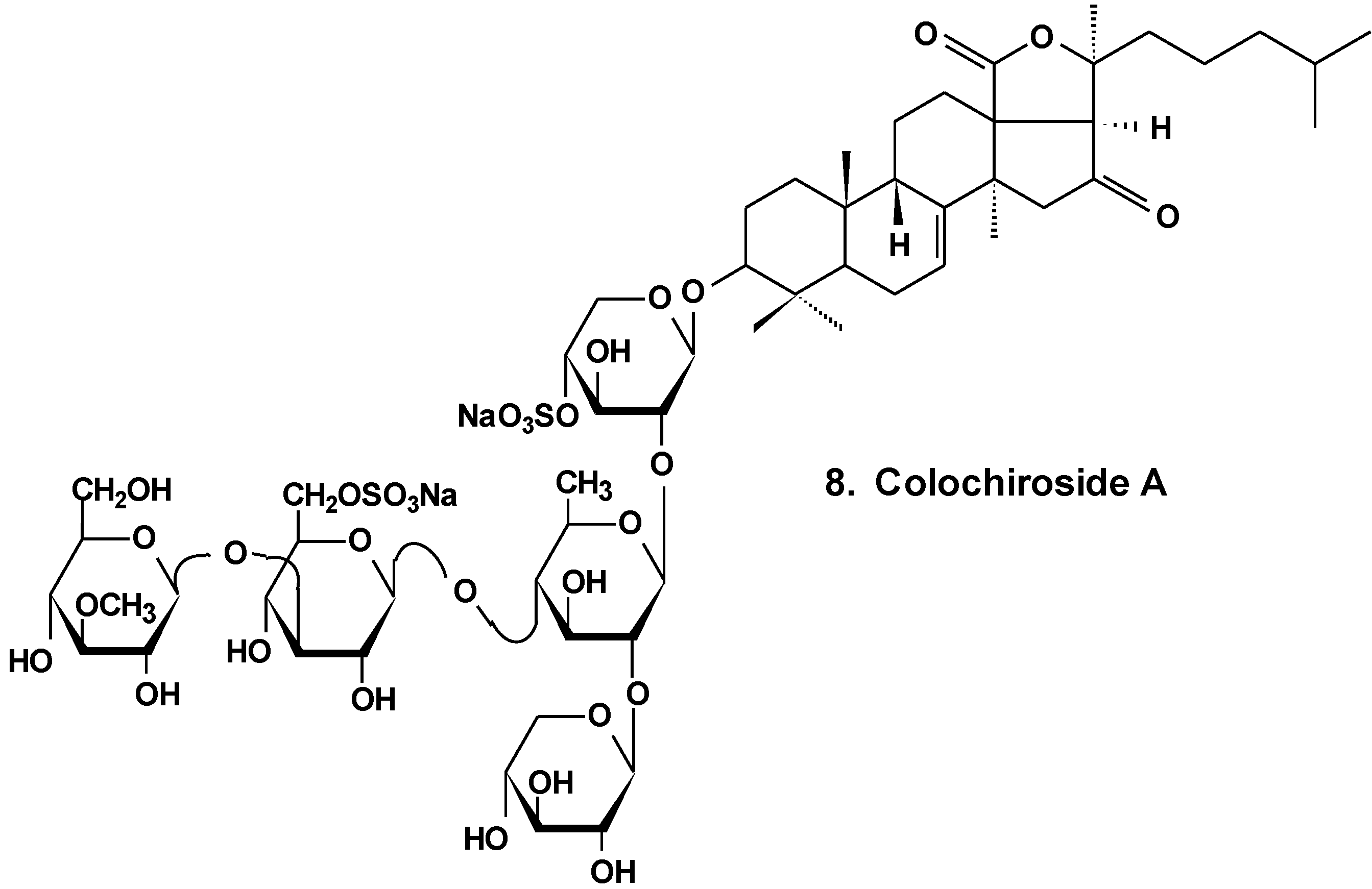
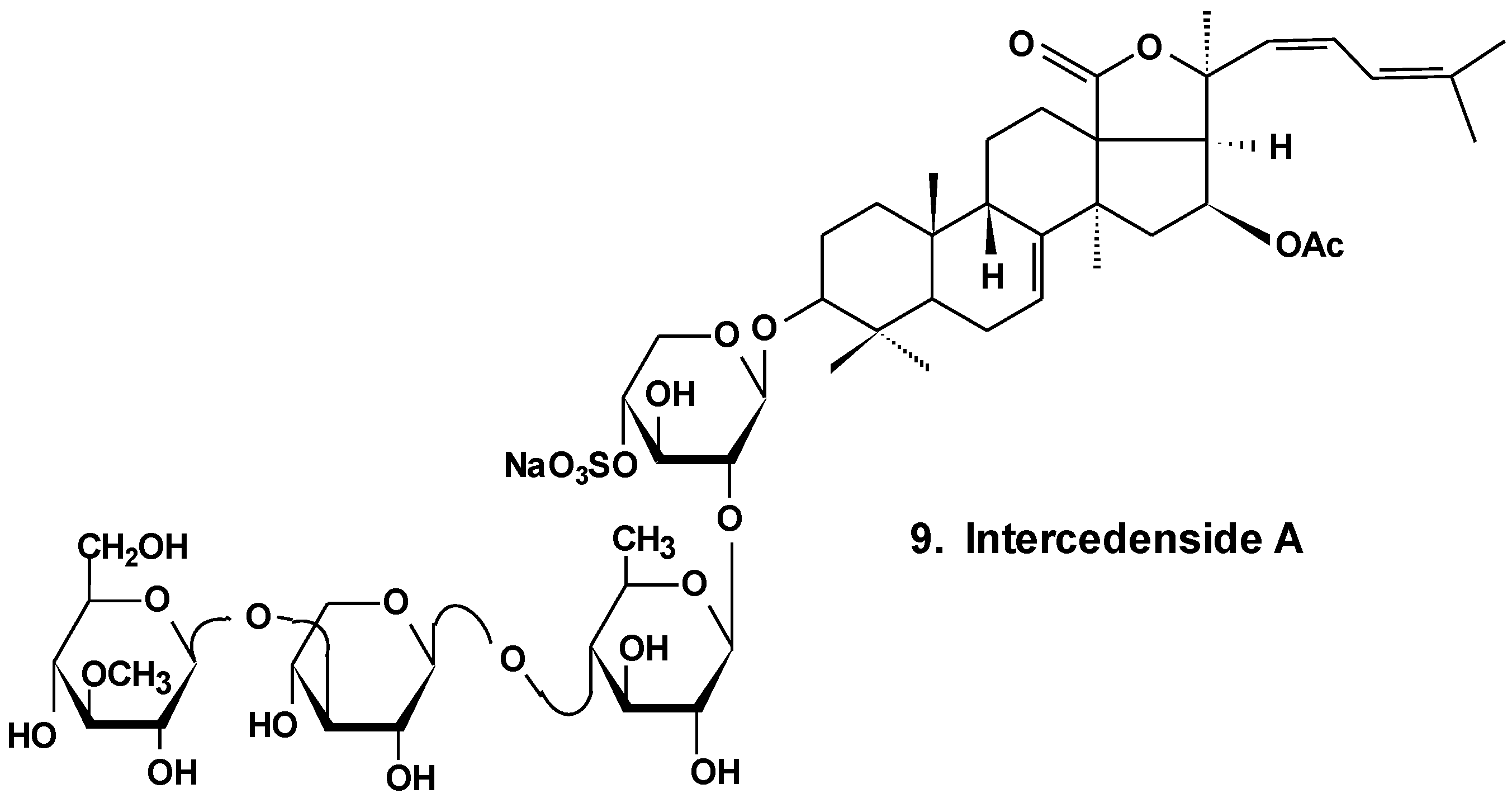


3. Conclusions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
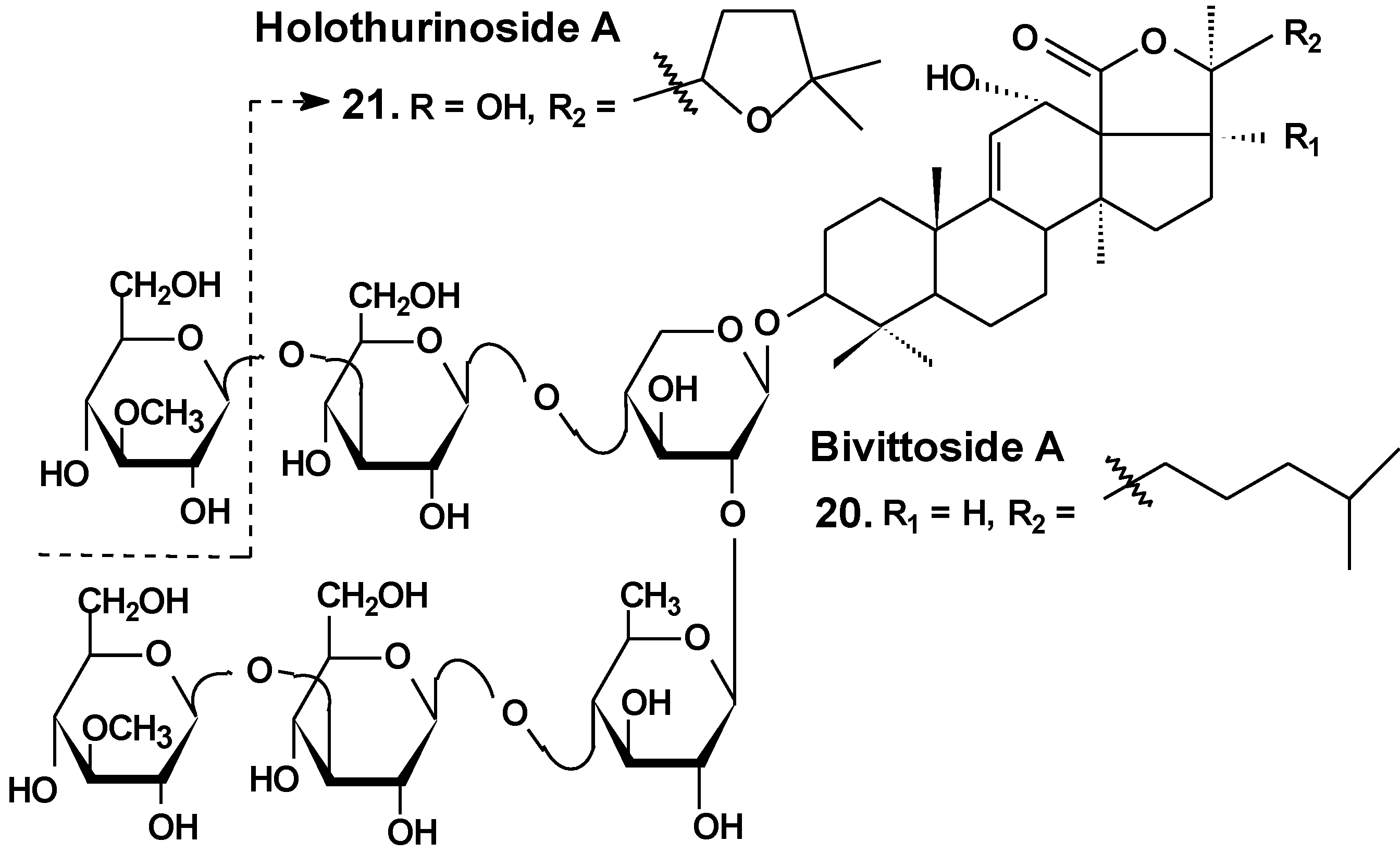
| # | Glycoside | Sea Cucumber sp. | Type of Activity |
|---|---|---|---|
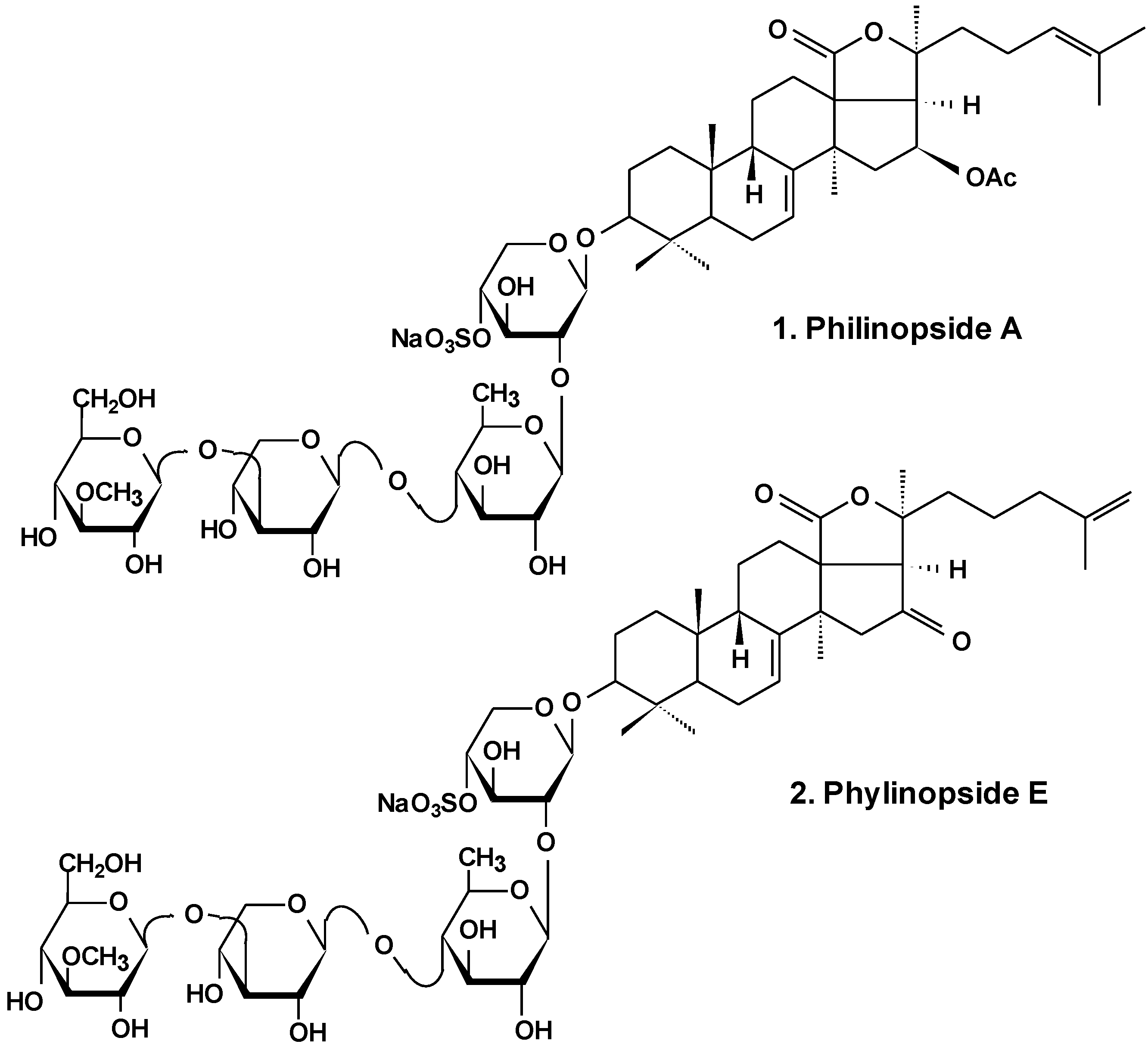
| 1 | Philinopside A | Pentacta quadrangularis | Reduction of cell viability [13,25], induction of apoptosis, inhibition of angiogenesis and tumor growth in vitro and in vivo [26,27,28] |
| 2 | Philinopside E | ||
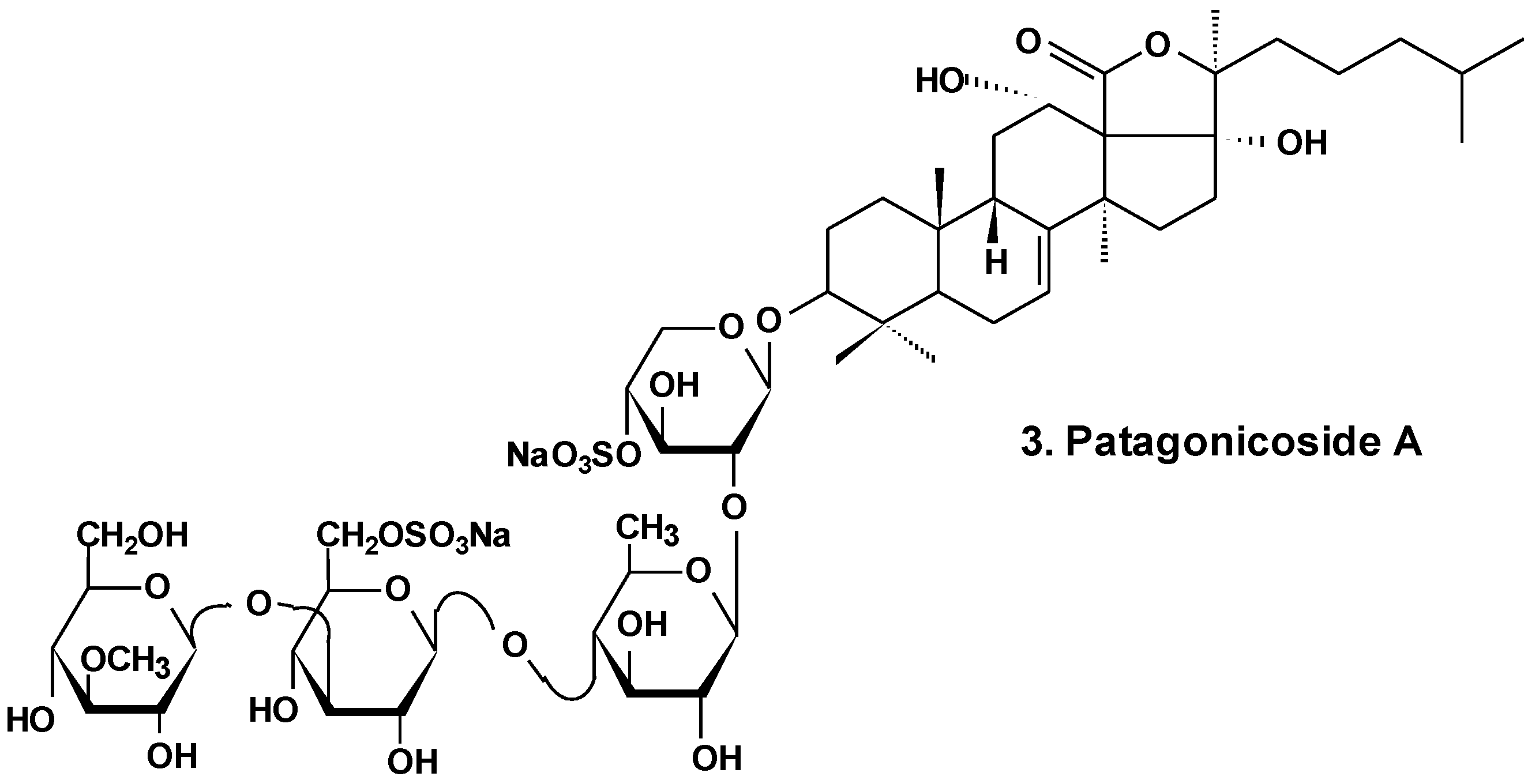
| 3 | Patagonicoside A | Psolus patagonicus | Suppression of cell proliferation [29] |
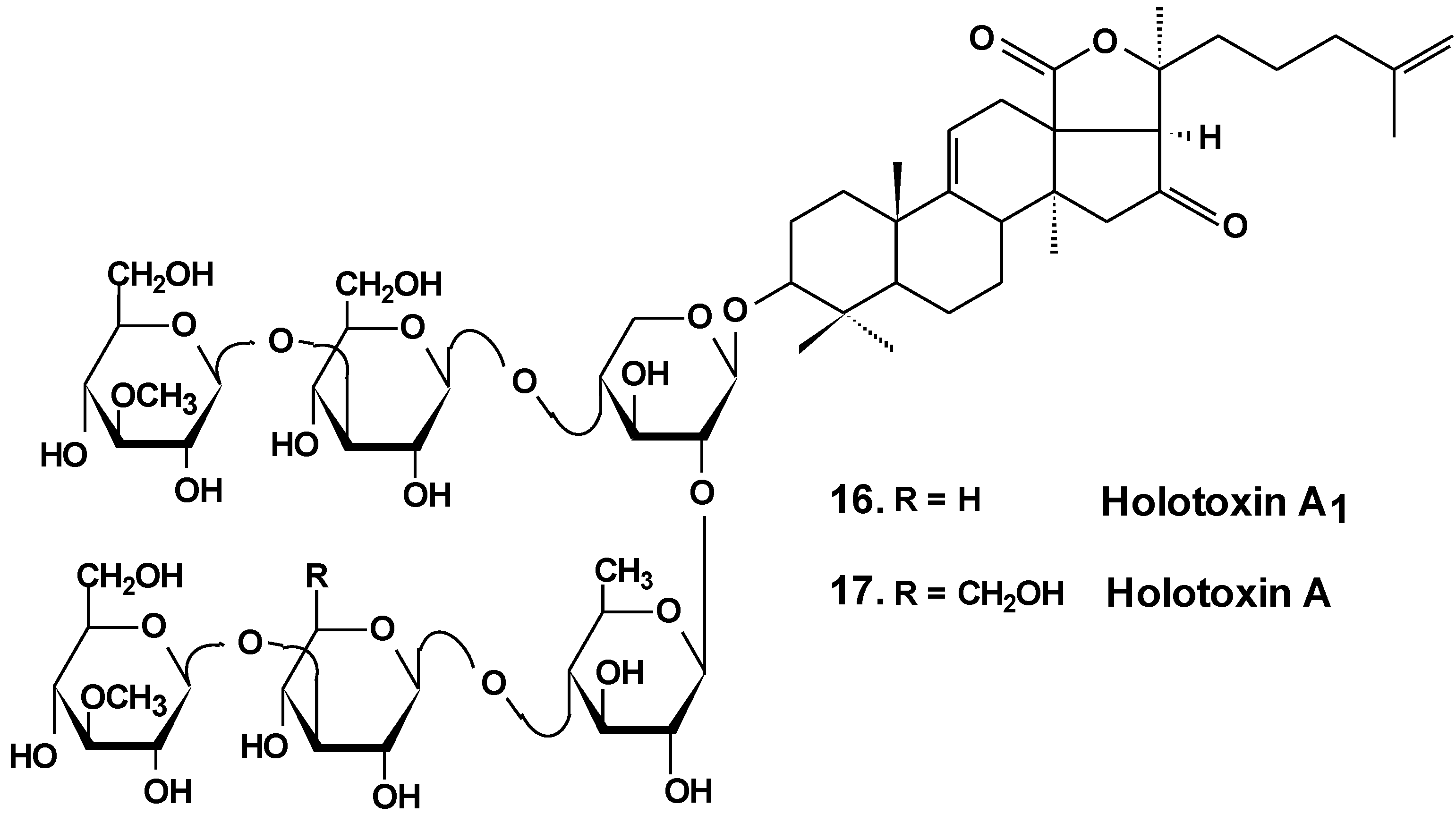
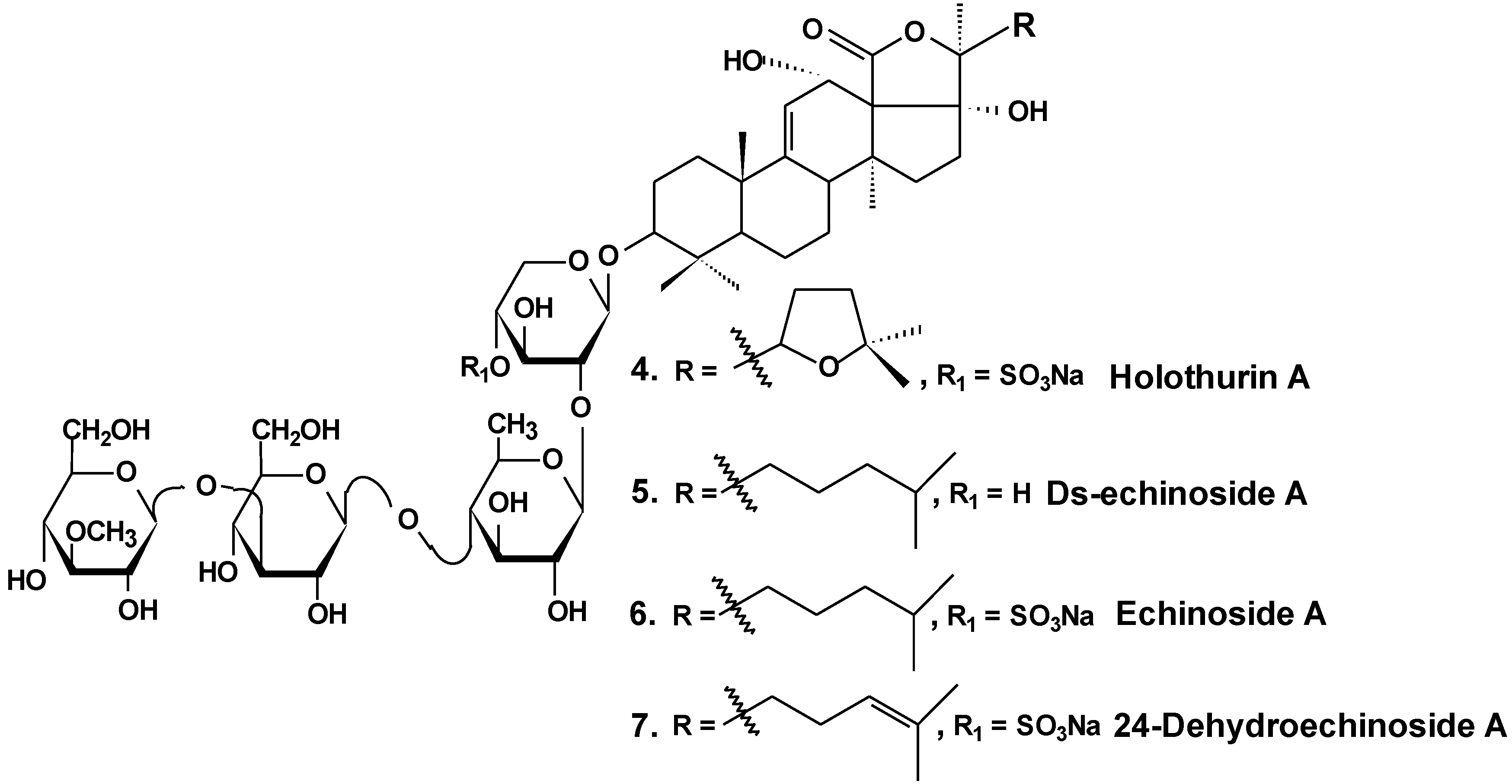
| 4 | Holothurin A | Pearsonothuria graeffei | Reduction of cell viability [30]; inhibition of cell adhesion, migration, metastasis and invasion [30,33]; induction of apoptosis [31]; cell cycle arrest, reduction of tumor growth in vivo [31,32] |
| 5 | Ds-echinoside A | ||
| 6 | Echinoside A | ||
| 7 | 24-dehydro echinoside A | ||
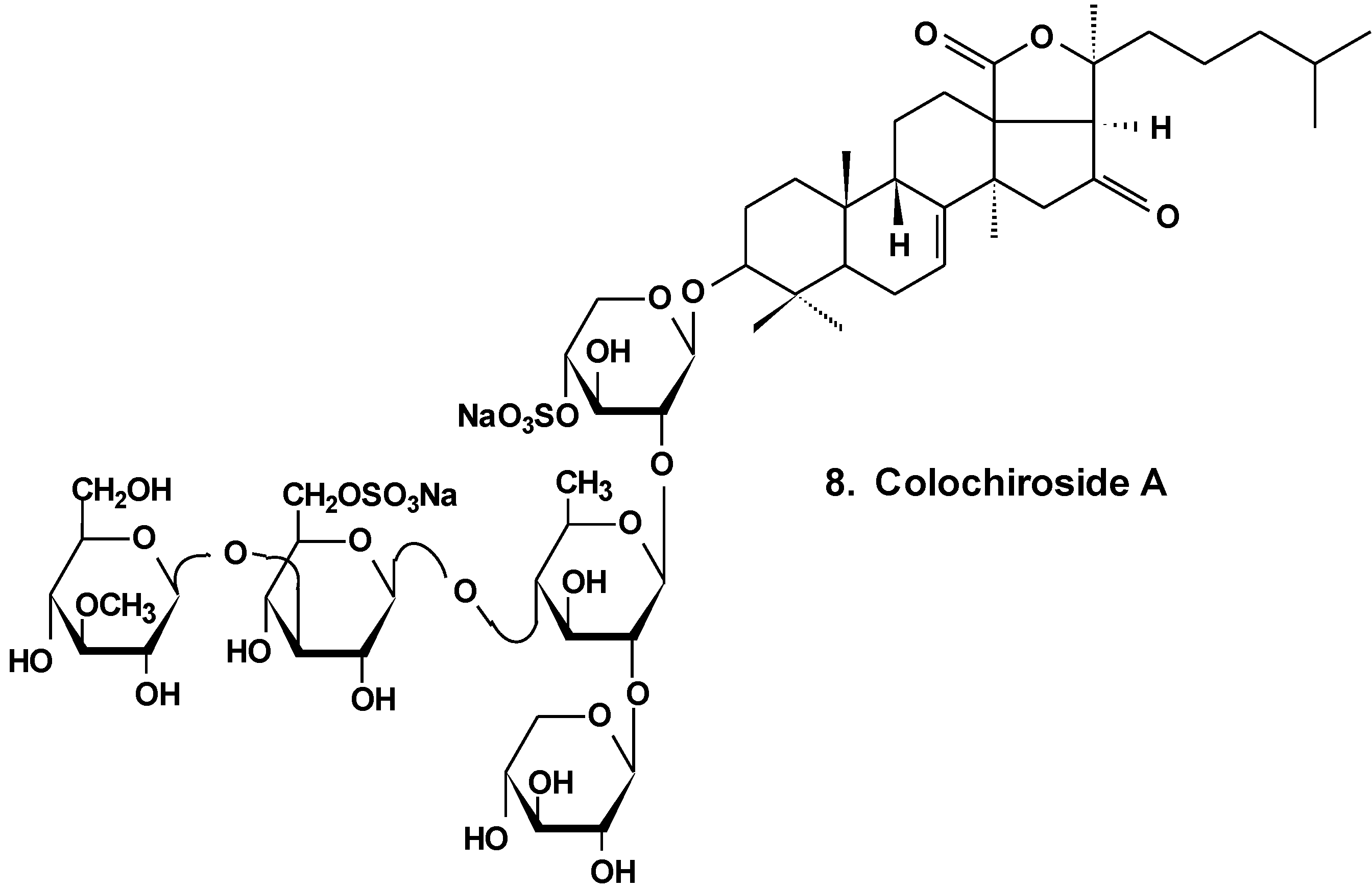
| 8 | Colochiroside A | Colochirus anceps | Reduction of cell viability, reduction of tumor growth in vivo [34] |
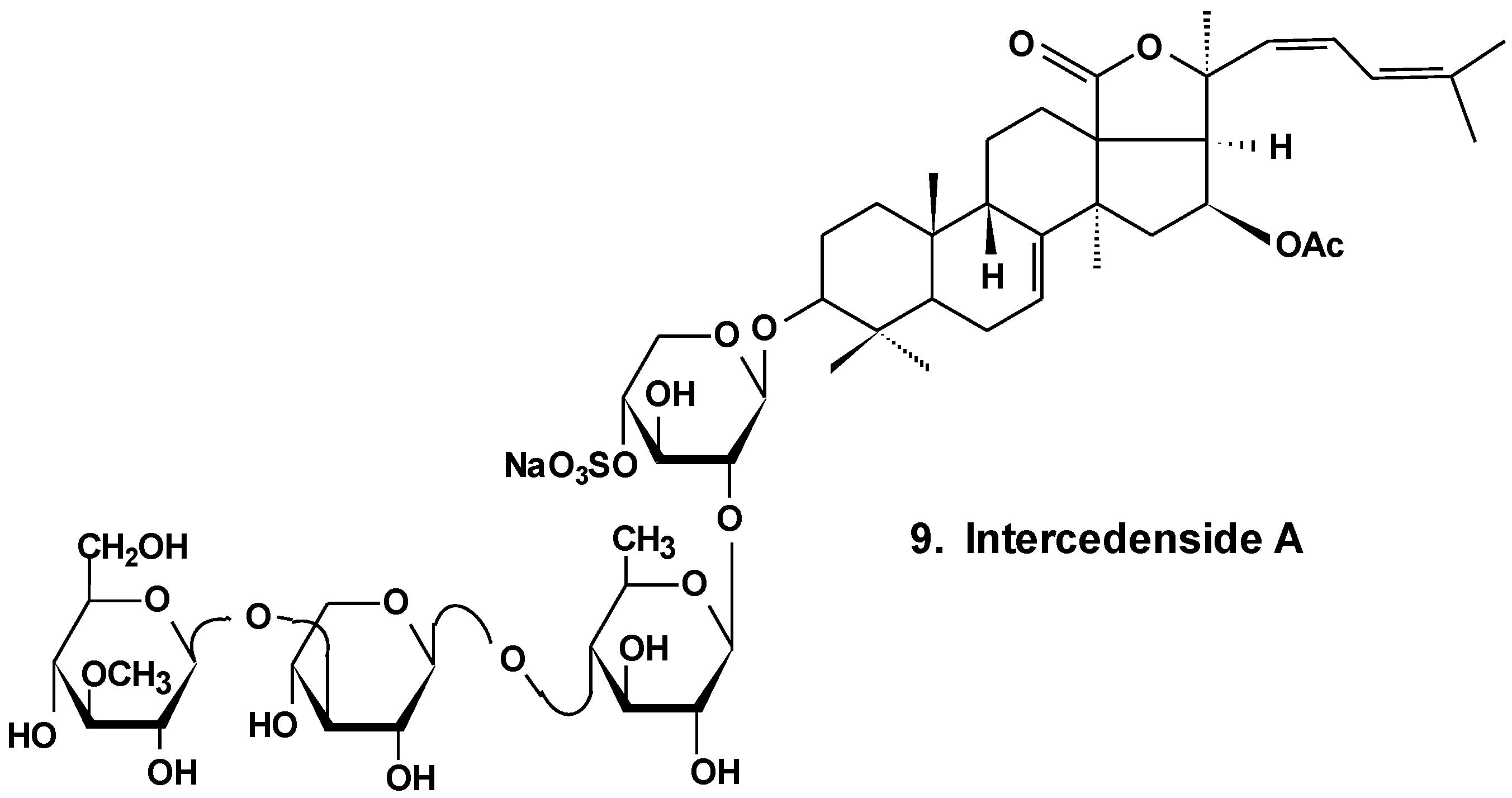
| 9 | Intercedenside A | Mensamaria intercedens | Reduction of cell viability, reduction of tumor growth in vivo [35] |
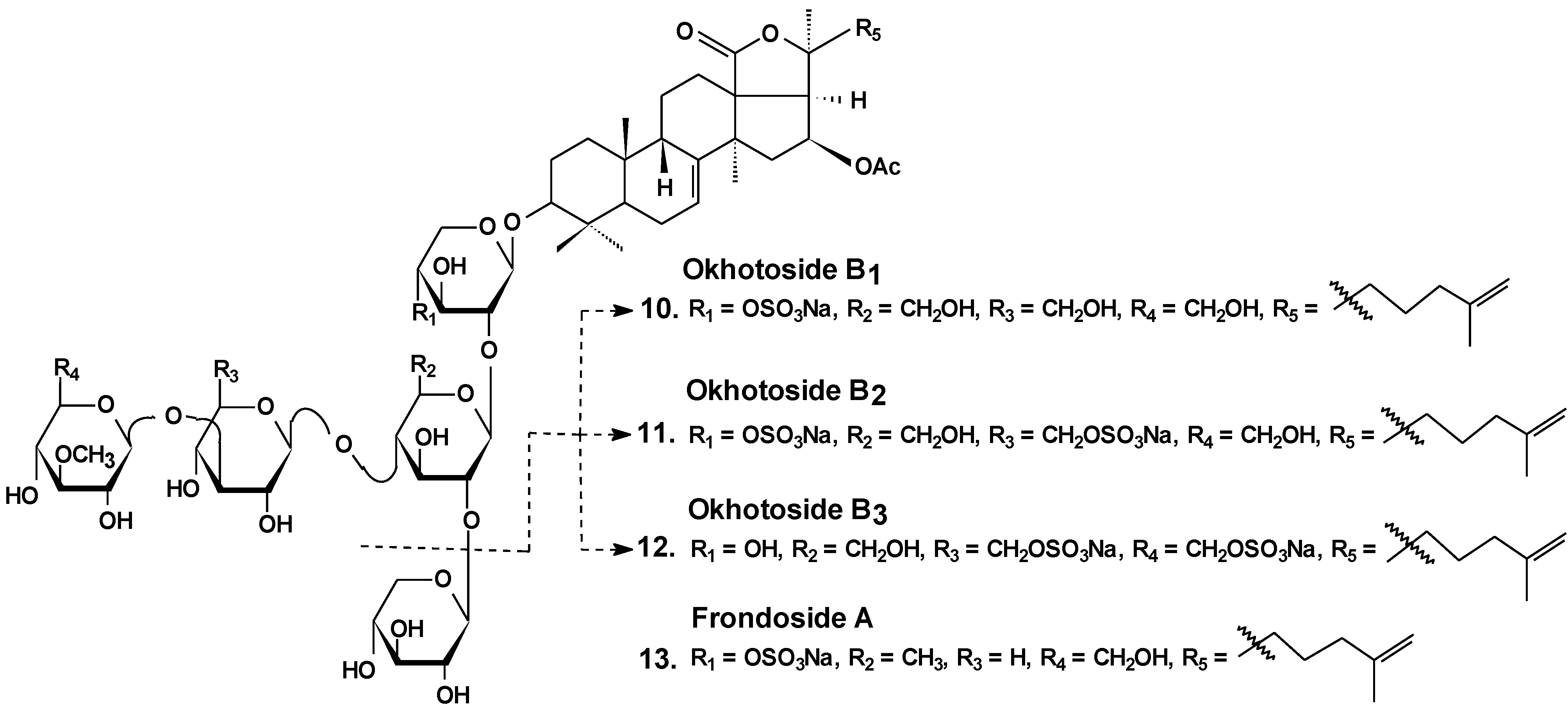
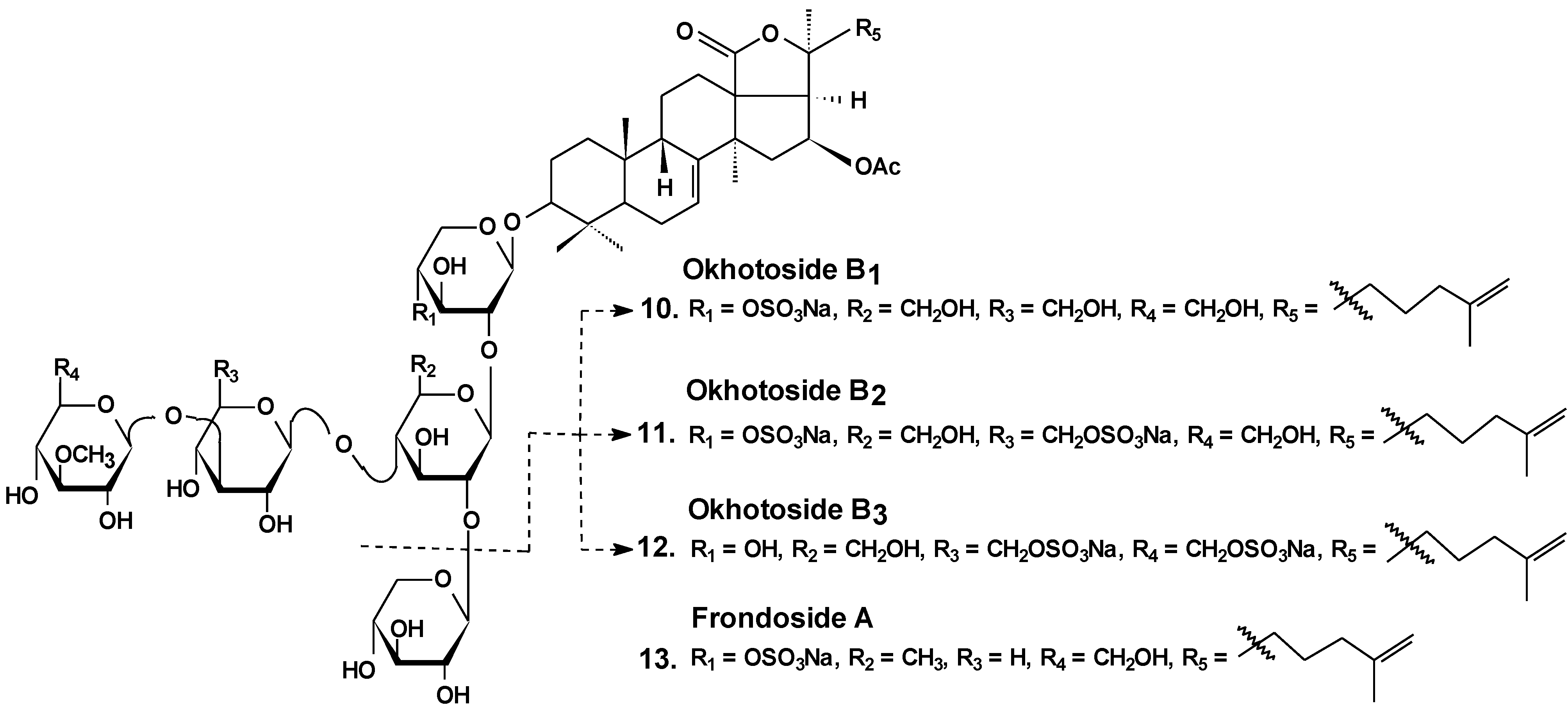
| 10 | Okhotoside B1 | Cucumaria okhotensis | Reduction of cell viability [36] |
| 11 | Okhotoside B2 | ||
| 12 | Okhotoside B3 | ||
| 13 | Frondoside A | Cucumaria frondosa Cucumaria okhotensis | Reduction of cell viability [36]; inhibition of colony formation [36,39,45] and cell proliferation [41,42], cell cycle arrest [39,40,41], induction of apoptosis [40,41,42,58]; inhibition of cell migration [43,44,45] and invasion [44,45]; inhibition of metastasis [43,44,58], angiogenesis [44]; inhibition of MDR [51,52,53]; reduction of tumor growth in vivo [41,44]; enhancement of tumor growth inhibition by cytostatics [42,44,47] |
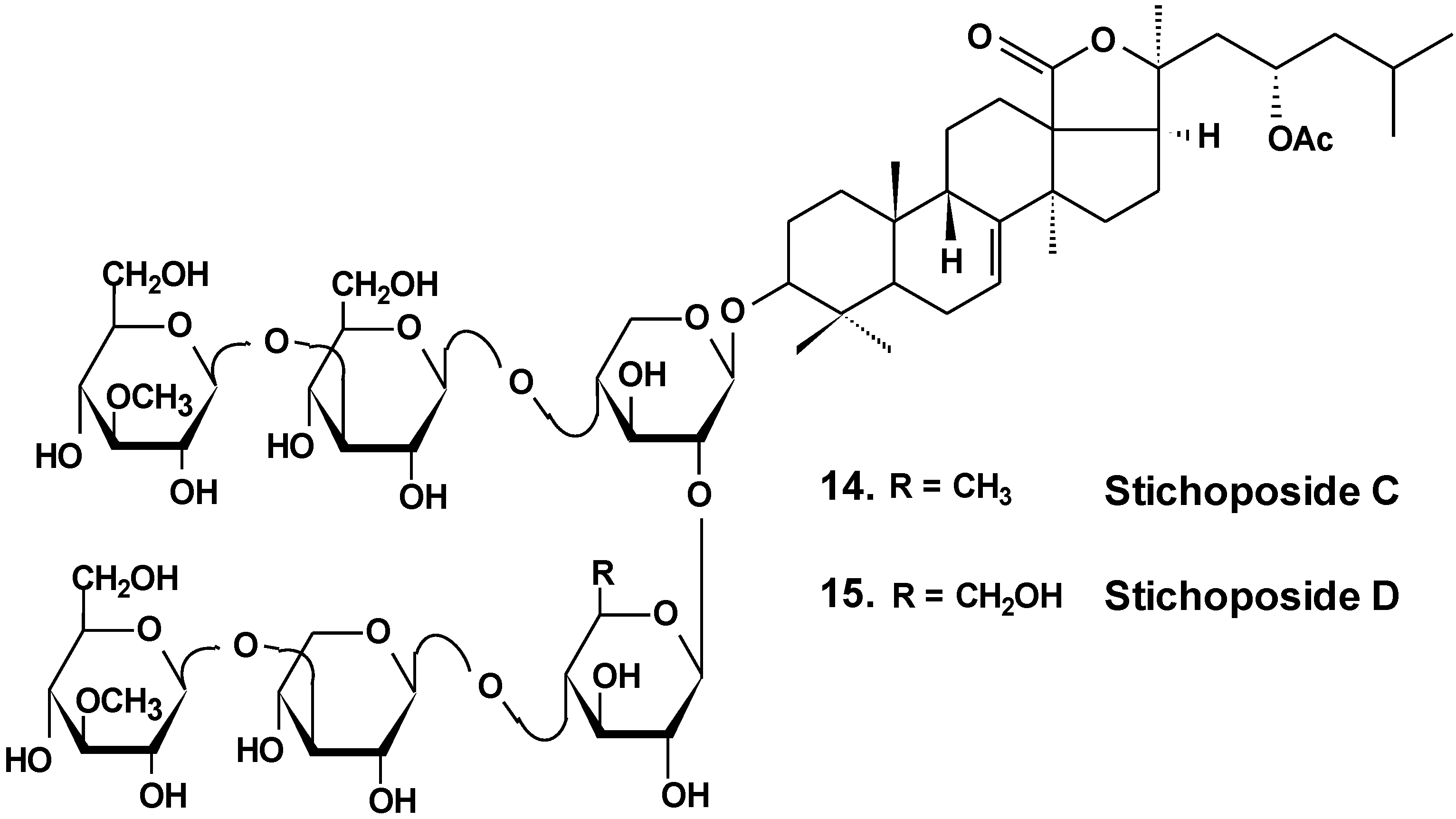
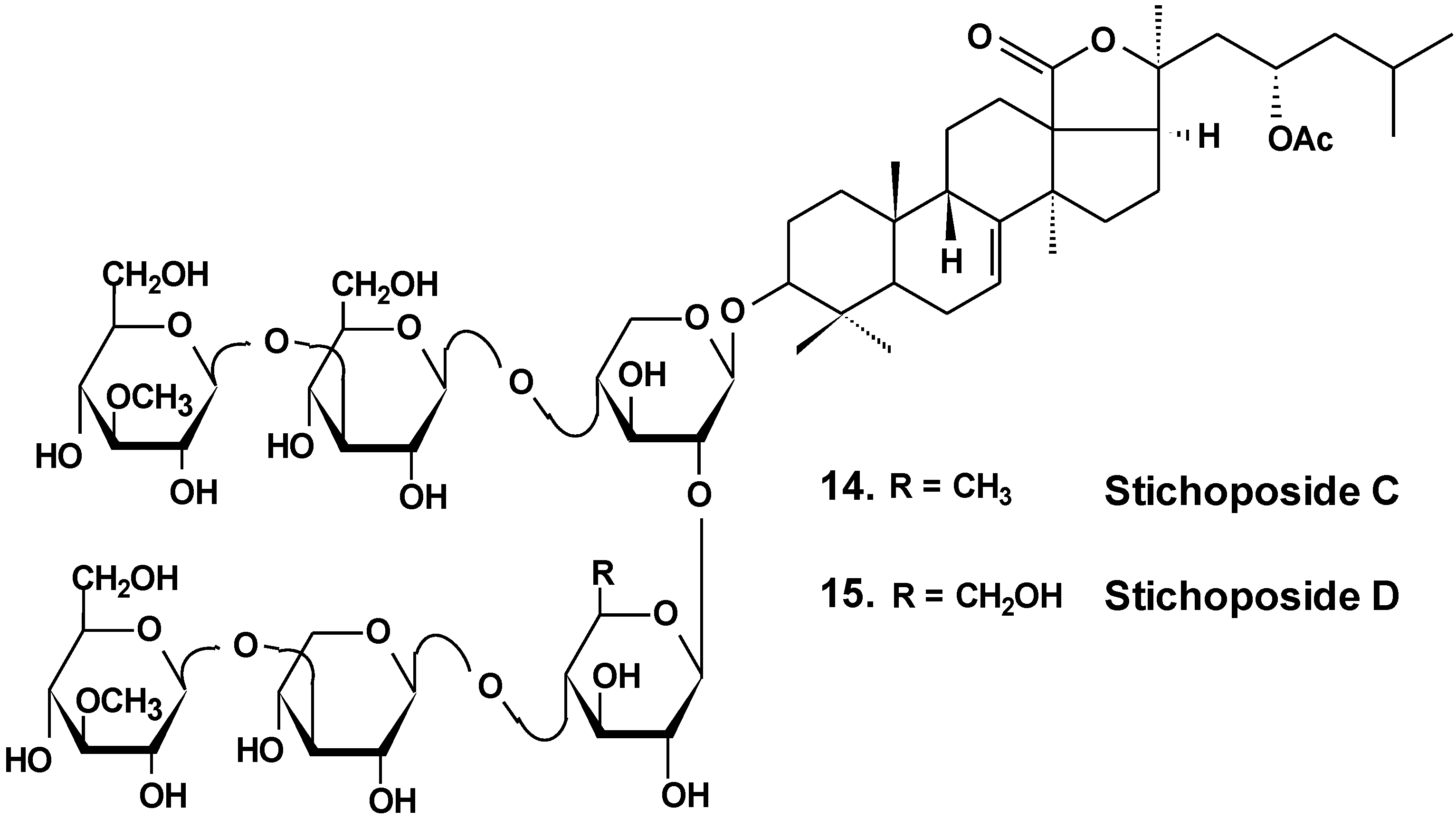
| 14 | Stichoposide C | Thelenota anax | Induction of apoptosis; reduction of tumor growth in vivo [38] |
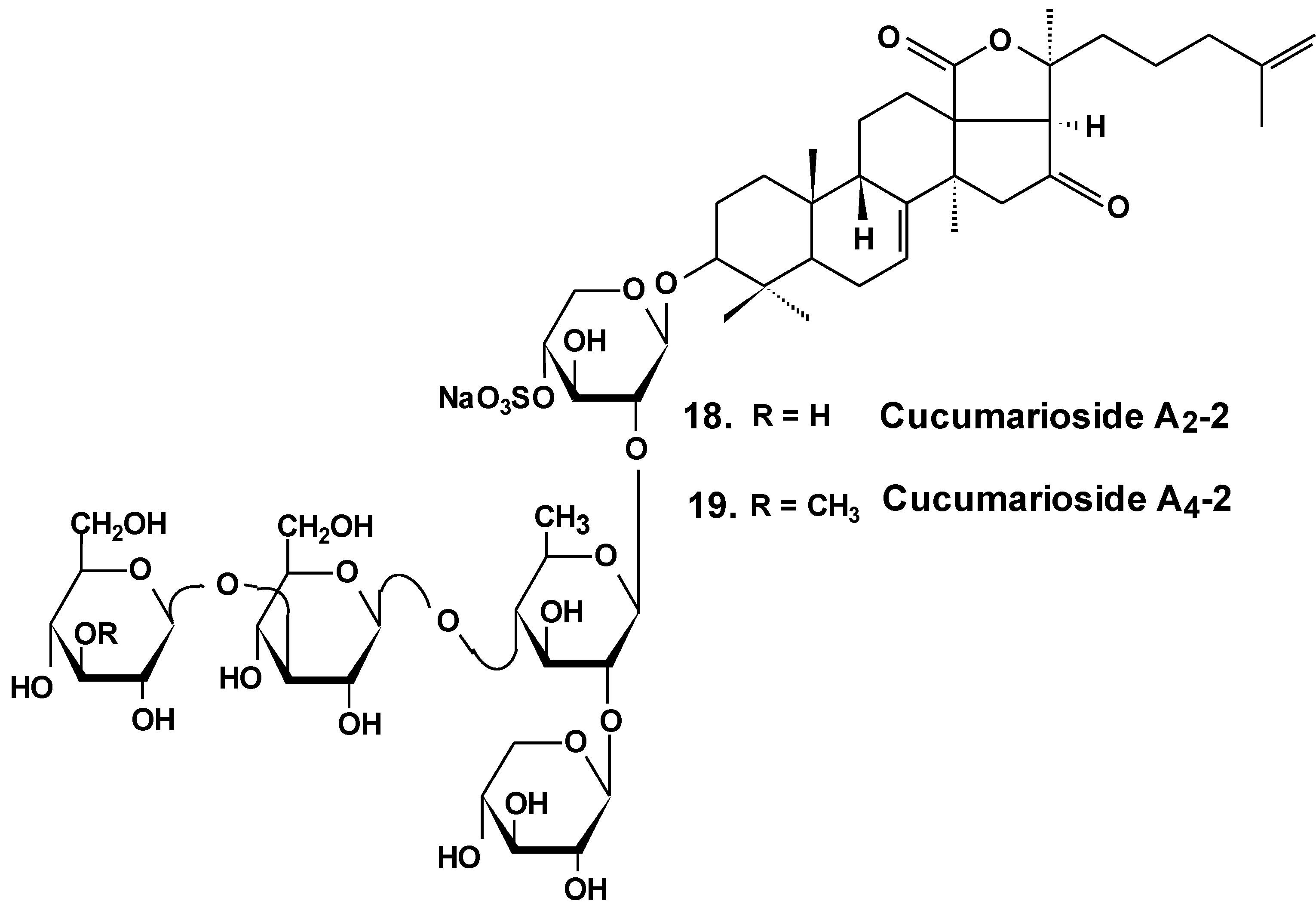
| 18 | Cucumariosides A2-2 | Cucumaria japonica | Inhibition of cell proliferation, cell cycle arrest and induction of apoptosis [48,49,50]; inhibition of metastasis and invasion [58]; reduction of tumor growth in vivo [48,49]; enhancement of tumor growth inhibition by cytostatics [55] inhibition of MDR [51,52,53] |
| 19 | Cucumarioside A4-2 |
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Brusca, R.C.; Brusca, G.J. Invertebrates, 2nd ed.; Sinauer Associates, Inc.: Sunderland, MA, USA, 2003; p. 936. [Google Scholar]
- Bordbar, S.; Anwar, F.; Saari, N. High-value components and bioactives from sea cucumbers for functional foods. A review. Mar. Drugs 2011, 9, 1761–1805. [Google Scholar] [CrossRef] [PubMed]
- Bakus, G.J. Defensive mechanisms and ecology of some tropical holothurians. Mar. Biol. 1968, 2, 23–32. [Google Scholar] [CrossRef]
- Rifkin, J.F. Venomous and Poisonous Marine Animals: A Medical and Biological Handbook; Burnett, J.W., Fenner, P.J., Eds.; UNSW Press: Kensington, NSW, Australia, 1996. [Google Scholar]
- Flammang, P.; Ribesse, J.; Jangoux, M. Biomechanics of adhesion in sea cucumber Cuvierian tubules (Echinodermata, Holothuroidea). Integr. Comp. Biol. 2002, 42, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Dyck, S.V.; Gerbaux, P.; Flammang, P. Qualitative and quantitative saponin contents in five sea cucumbers from the Indian Ocean. Mar. Drugs 2010, 8, 173–189. [Google Scholar] [CrossRef] [PubMed]
- Dyck, S.V.; Caulier, G.; Todesco, M.; Gerbaux, P.; Fournier, I.; Wisztorski, M.; Flammang, P. The triterpene glycosides of Holothuria forskali: Usefulness and efficiency as a chemical defense mechanism against predatory fish. J. Exp. Biol. 2011, 214, 1347–1356. [Google Scholar] [CrossRef] [PubMed]
- Frey, D.G. The use of sea cucumber in poisoning fishes. Copeia 1951, 2, 175–176. [Google Scholar] [CrossRef]
- Podolak, I.; Galanty, A.; Sobolewska, D. Saponins as cytotoxic agents: A review. Phytochem. Rev. 2010, 9, 425–474. [Google Scholar] [CrossRef] [PubMed]
- Osbourn, A.; Goss, R.J.; Field, R.A. The saponins-polar isoprenoids with important and diverse biological activities. Nat. Prod. Rep. 2011, 28, 1261–1268. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.K.; Himaya, S.W. Triterpene glycosides from sea cucumbers and their biological activities. Adv. Food. Nutr. Res. 2012, 65, 297–319. [Google Scholar] [PubMed]
- Kalinin, V.I.; Aminin, D.L.; Avilov, S.A.; Silchenko, A.S.; Stonik, V.A. Triterpene glycosides from sea cucumbers (Holothurioidae, Echinodermata), biological activities and functions. In Studies in Natural Product Chemistry (Bioactive Natural Products); Atta-ur-Rahman, Ed.; Elsevier Science Publisher: Amsterdam, The Netherlands, 2008; Volume 35, pp. 135–196. [Google Scholar]
- Han, H.; Xu, Q.Z.; Tang, H.F.; Yi, Y.H.; Gong, W. Cytotoxic holostane-type triterpene glycosides from the sea cucumber Pentacta quadrangularis. Planta Med. 2010, 76, 1900–1904. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.S.; Yi, Y.H.; Li, L.; Sun, P.; Han, H.; Sun, G.Q.; Wang, X.H.; Wang, Z.L. Argusides D and E, two new cytotoxic triterpene glycosides from the sea cucumber Bohadschia argus Jaeger. Chem. Biodivers. 2008, 5, 1425–1433. [Google Scholar] [CrossRef] [PubMed]
- Avilov, S.A.; Silchenko, A.S.; Antonov, A.S.; Kalinin, V.I.; Kalinovsky, A.I.; Smirnov, A.V.; Dmitrenok, P.S.; Evtushenko, E.V.; Fedorov, S.N.; Savina, A.S.; et al. Synaptosides A and A1, triterpene glycosides from the sea cucumber Synapta maculata containing 3-O-methylglucuronic acid and their cytotoxic activity against tumor cells. J. Nat. Prod. 2008, 71, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Althunibat, O.Y.; Ridzwan, B.H.; Taher, M.; Jamaludin, M.D.; Ikeda, M.A.; Zali, B.I. In vitro antioxidant and antiproliferative activities of three Malaysian sea cucumber species. Eur. J. Sci. Res. 2009, 37, 376–387. [Google Scholar]
- Wu, J.; Yi, Y.H.; Tang, H.F.; Wu, H.M.; Zhou, Z.R. Hillasides A and B, two new cytotoxic triterpen glycosides from the sea cucumber Holoturia hilla. J. Asian. Nat. Prod. Res. 2007, 9, 609–615. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.Y.; Tang, H.F.; Yi, Y.H. Cytotoxic triterpene glycosides from the sea cucumber Pseudocolochirus violaceus. Fitoterapia 2007, 78, 283–287. [Google Scholar] [CrossRef] [PubMed]
- Sun, P.; Liu, B.S.; Yi, Y.H.; Li, L.; Gui, M.; Tang, H.F.; Zhang, D.Z.; Zhang, S.L. New cytotoxic lanostane-type triterpene glycoside from the sea cucumber Holothuria impatiens. Chem. Biodivers. 2007, 4, 450–457. [Google Scholar] [CrossRef] [PubMed]
- Nigrelli, R.F. The effects of holothurin on fish and mice with sarcoma 180. Zoologica (NY) 1952, 37, 89–90. [Google Scholar]
- Sullivan, T.D.; Ladue, K.T.; Nigrelli, R.F. The effects of holothurin, a steroid saponin of animal origin, on Krebs-2 ascites tumors in Swiss mice. Zoologica 1955, 40, 49–52. [Google Scholar]
- Sullivan, T.D;; Nigrelli, R.F. The antitumorous action of biologics of marine origin I. Survival of Swiss mice inoculated with Krebs-2 ascites tumor and treated with holothurin, a steroid saponin from the sea cucumber, Actinopyga agassizi. Proc. Am. Assoc. Cancer Res. 1956, 2, 151–155. [Google Scholar]
- Friess, S.L.; Standaert, F.G.; Whitcomb, E.R.; Nigrelli, R.F.; Chanley, J.D.; Sobotka, H. Some pharmacologic properties of holothurin A, a glycosidic mixture from the sea cucumber. Ann. N. Y. Acad. Sci. 1960, 90, 893–901. [Google Scholar] [CrossRef] [PubMed]
- Nigrelli, R.F.; Stempien, M.F.J.; Ruggieri, G.D.; Liguori, V.R.; Cecil, J.T. Substances of potential biomedical importance from marine organisms. Fed. Proc. 1967, 26, 1197–1205. [Google Scholar] [PubMed]
- Zhang, S.L.; Li, L.; Yi, Y.H.; Sun, P. Philinopsides E and F, two new sulfated triterpene glycosides from the sea cucumber Pentacta quadrangularis. Nat Prod Res. 2006, 20, 399–407. [Google Scholar] [CrossRef] [PubMed]
- Tong, Y.; Zhang, X.; Tian, F.; Yi., Y.; Xu, Q.; Li, L.; Tong, L.; Lin, L.; Ding, J. Philinopside A, a novel marine-derived compound possesing dual anti-angiogenetic and anti-tumor effects. Int. J. Cancer 2005, 114, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Tian, F.; Zhang, X.; Tong, Y.; Yi, Y.; Zhang, S.; Li, L.; Sun, P.; Lin, L.; Ding, J. PE, a new sulfated saponin from sea cucumber, exhibits anti-angiogenic and anti-tumor activities in vitro and in vivo. Cancer Biol. Ther. 2005, 48, 874–882. [Google Scholar] [CrossRef]
- Tian, F.; Zhu, C.H.; Zhang, X.W.; Xie, X.; Xin, X.L.; Yi, Y.H.; Lin, L.P.; Geng, M.Y.; Ding, J. Philinopside E, a new sulfated saponin from sea cucumber, blocks the interaction between kinase insert domain-containing receptor (KDR) and alphavbeta3 integrin via binding to the extracellular domain of KDR. Mol. Pharmacol. 2007, 72, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Careaga, V.P.; Bueno, C.; Muniain, C.; Alche, L.; Maier, M.S. Antiproliferative, cytotoxic and hemolytic activities of a triterpene glycoside from Psolus patagonicus and its desulfated analog. Chemotherapy 2009, 55, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Liu, Z.; Xue, Y.; Wang, J.; Li, H.; Tang, Q.; Wang, Y.; Dong, P.; Xue, C. Ds-echinoside A, a new triterpene glycoside derived from sea cucumber, exhibits antimetastatic activity via the inhibition of NF-κB-dependent MMP-9 and VEGF expressions. J. Zhejiang Univ. Sci. B 2011, 12, 534–544. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Xue, Y.; Wang, J.F.; Li, H.; Long, T.T.; Li, Z.; Wang, Y.M.; Dong, P.; Xue, C. In vitro and in vivo anti-tumor activities of echinoside A and ds-echinoside A from Pearsonothuria graeffei. J. Sci. Food Agric. 2012, 92, 965–974. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Miao, Z.H.; Chen, Z.; Chen., Q.; Gui, M.; Lin, L.P.; Sun, P.; Yi, Y.H.; Ding, J. Echinoside A, a new marine-derived anticancer saponin, targets topoisomerase2α by unique interference with its DNA binding and catalytic cycle. Ann. Oncol. 2010, 21, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Xue, Y.; Liu, Z.; Li, H.; Wang, J.; Li, Z.; Wang, Y.; Dong, P.; Xue, C. Differential effects of sulfated triterpene glycosides, Holothurin A1, and 24-Dehydroechinoside A, on antimetastasic activity via regulation of the MMP-9 signal pathway. J. Food Sci. 2010, 75, 280–288. [Google Scholar] [CrossRef]
- Zhang, Y.; Yi, Y. Studies on antitumor activities of triterpene glycoside colochiroside A from sea cucumber Colochirus anceps. Zhongguo Zhong Yao Za Zhi 2011, 36, 504–507. [Google Scholar] [PubMed]
- Zou, Z.; Yi, Y.; Wu, H.; Wu, J.; Liaw, C.; Lee, K. Intercedensides A–C, three new cytotoxic triterpene glycosides from the sea cucumber Mensamaria intercedens Lampert. J. Nat. Prod. 2003, 66, 1055–1060. [Google Scholar] [CrossRef] [PubMed]
- Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I.; Kalinovsky, A.I.; Dmitrenok, P.S.; Fedorov, S.N.; Stepanov, V.G.; Dong, Z.; Stonik, V.A. Constituents of the Sea Cucumber Cucumaria okhotensis. Structures of okhotosides B1–B3 and cytotoxic activities of some glycosides from this species. J. Nat. Prod. 2008, 71, 351–356. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.J.; Yuan, W.P.; Cong, R.S.; Yang, X.X.; Wang, W.W.; Jing, Z. Studies on the purification of water-soluble holothurian glycosides from Apostichopus japonicus and their tumor suppressing activity. Yao Xue Xue Bao 2009, 44, 25–31. [Google Scholar] [PubMed]
- Yun, S.H.; Park, E.S.; Shin, S.W.; Na, Y.W.; Han, J.Y.; Jeong, J.S.; Shastina, V.V.; Stonik, V.A.; Park, J.I.; Kwak, J.Y. Stichoposide C induces apoptosis through the generation of ceramide in leukemia and colorectal cancer cells and shows in vivo antitumor activity. Clin. Cancer Res. 2012, 18, 5934–5948. [Google Scholar] [CrossRef] [PubMed]
- Janakiram, N.B.; Mohammed, A.; Zhang, Y.; Choi, C.; Woodward, C.; Collin, P.; Steele, V.E.; Rao, C.V. Chemopreveintive effects of Frondanol A5, a Cucumaria frondosa extract, against rat colon carcinogenesis and inhibition of human colon cancer cell growth. Cancer Prev. Res. 2010, 3, 82–91. [Google Scholar] [CrossRef]
- Roginsky, A.B.; Ding, X.; Woodward, C.; Ujiki, M.B.; Singh, B.; Bell, R.H.; Collin, P.; Adrian, T.E. Woodward C. Anti-pancreatic cancer effects of a polar extract from the edible sea cucumber, Cucumaria frondosa. Pancreas 2010, 39, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Roginsky, A.B.; Ding, X.; Woodward, C.; Collin, P.; Newman, R.A.; Bell, R.H.; Adrian, T.E. Review of the apoptosis pathways in pancreatic cancer and the apoptotic effects of the novel sea cucumber compound, frondoside A. Ann. N.Y. Acad. Sci. 2008, 1138, 181–198. [Google Scholar] [CrossRef] [PubMed]
- Al Marzouqi, N.; Iratni, R.; Nemmar, A.; Arafat, K.; Al Sultan, M.A.H.; Collin, P. Frondoside A inhibits human breast cancer cell survival, migration, invasion and the growth of breast tumor xenografts. Eur. J. Pharmacol. 2011, 651, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.; Kundu, N.; Collin, P.D.; Goloubeva, O.; Fulton, A.M. Frondoside A inhibits breast cancer metastasis and antagonizes prostaglandin E receptors EP4 and EP2. Breast Cancer Res. Treat. 2012, 132, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Attoub, S.; Arafat, K.; Gelaude, A.; al Sultan, M.A.; Bracke, M.; Collin, P.; Takahashi, T.; Adrian, T.E.; de Wever, O. Frondoside A suppressive effects on lung cancer survival, tumor growth, angiogenesis, invasion, and metastasis. PLoS One 2013, 8, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Park, S.Y.; Kim, Y.H.; Kim, Y.; Lee, S.J. Frondoside A has an anti-invasive effect by inhibiting TPA-induced MMP-9 activation via NF-κB and AP-1 signaling in human breast cancer cells. Int. J. Oncol. 2012, 41, 933–940. [Google Scholar] [PubMed]
- Kovalchuk, S.N.; Kozhemyako, V.B.; Atopkina, L.N.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I.; Rasskazov, V.A.; Aminin, D.L. Estrogenic activity of triterpene glycosides in yeast two-hybrid assay. J. Steroid Biochem. Mol. Biol. 2006, 101, 226–231. [Google Scholar] [CrossRef] [PubMed]
- Al Shemaili, J.; Mensah-Brown, E.; Parekh, K.; Thomas, S.A.; Attoub, S.; Hellman, B.; Nyberg, F.; Adem, A.; Collin, P.; Adrian, T.E. Frondoside A enhances the antiproliferative effects of gemcitabine in pancreatic cancer. Eur. J. Cancer 2014, 50, 1391–1398. [Google Scholar] [CrossRef] [PubMed]
- Fedorov, S.N.; Shubina, L.K.; Kapustina, I.I.; Avilov, S.A.; Kwak, J.Y.; Park, J.I.; Jin, J.O.; Kwon, J.X; Shastina, V.V.; Stonik, V.A. Agent stimulating apoptosis of human leukaemia cells. Patent of the Russian Federation 2009 2360692, 21 December 2007. [Google Scholar]
- Jin, J.O.; Shastina, V.V.; Shin, S.W.; Xu, Q.; Park, J.I.; Rasskazov, V.A.; Avilov, S.A.; Fedorov, S.N.; Stonik, V.A.; Kwak, J.Y. Differential effects of triterpene glycosides, frondoside A and cucumarioside A2–2 isolated from sea cucumbers on caspase activation and apoptosis of human leukemia cells. FEBS Lett. 2009, 583, 697–702. [Google Scholar] [CrossRef] [PubMed]
- Menchinskaya, E.S.; Pislyagin, E.A.; Kovalchyk, S.N.; Davydova, V.N.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I.; Aminin, D.L. Antitumor activity of cucumarioside A2–2. Chemotherapy 2013, 59, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Aminin, D.L.; Menchinskaya, E.S.; Pislyagin, E.A.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I.; Stonik, V.A. New anticancer compounds from sea cucumbers. Molecular mechanisms of action. FEBS J. 2014, 281, 488. [Google Scholar]
- Menchinskaya, E.S.; Aminin, D.L.; Avilov, S.A.; Silchenko, A.S.; Andryjashchenko, P.V.; Kalinin, V.I.; Stonik, V.A. Agent inhibiting multiple drug resistance of tumor cells. Patent of the Russian Federation 2013 2494742, 10 August 2012. [Google Scholar]
- Menchinskaya, E.S.; Aminin, D.L.; Avilov, S.A.; Silchenko, A.S.; Andryjashchenko, P.V.; Kalinin, V.I.; Stonik, V.A. Inhibition of tumor cells multidrug resistance by cucumarioside A2–2, frondoside А and their complexes with а cholesterol. Nat. Prod. Commun. 2013, 8, 1377–1380. [Google Scholar] [PubMed]
- Stonik, V.A.; Aminin, D.L.; Boguslavski, V.M.; Avilov, S.A.; Agafonova, I.G.; Silchenko, A.S.; Ponomarenko, L.P.; Prokofieva, N.G.; Chaikina, E.L. Immunomodulating agent and pharmaceutical composition based on the same. Patent of the Russian Federation 2005 2271820, 2 July 2004. [Google Scholar]
- Aminin, D.L.; Chaykina, E.L.; Agafonova, I.G.; Avilov, S.A.; Kalinin, V.I.; Stonik, V.A. Antitumor activity of the immunomodulatory lead Cumaside. Int. Immunopharmacol. 2010, 10, 648–654. [Google Scholar] [CrossRef] [PubMed]
- Patil, T.D.; Thakare, S.V. In silico evaluation of selected triterpene glycosides as a human dna topoisomerase II alpha (α) inhibitor. Int. J. Pharm. Pharm. Sci. 2012, 4, 201–204. [Google Scholar]
- Park, J.-I.; Bae, H.-R.; Kim, C.G.; Stonik, V.A.; Kwak, J.-Y. Relationships between chemical structures and functions of triterpene glycosides isolated from sea cucumbers. Front. Chem. 2014, 2, 1–14. [Google Scholar]
- Kim, C.G.; Kwak, J.-Y. Anticancer effect of triterpene glycosides, frondoside A and cucumarioside A2-2 isolated from sea cucumbers. In Handbook of Anticancer Drugs from Marine Origin; Kim, S.-K., Ed.; Springer International Publishing: Cham, Switzerland, 2015; pp. 673–682. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aminin, D.L.; Menchinskaya, E.S.; Pisliagin, E.A.; Silchenko, A.S.; Avilov, S.A.; Kalinin, V.I. Anticancer Activity of Sea Cucumber Triterpene Glycosides. Mar. Drugs 2015, 13, 1202-1223. https://doi.org/10.3390/md13031202
Aminin DL, Menchinskaya ES, Pisliagin EA, Silchenko AS, Avilov SA, Kalinin VI. Anticancer Activity of Sea Cucumber Triterpene Glycosides. Marine Drugs. 2015; 13(3):1202-1223. https://doi.org/10.3390/md13031202
Chicago/Turabian StyleAminin, Dmitry L., Ekaterina S. Menchinskaya, Evgeny A. Pisliagin, Alexandra S. Silchenko, Sergey A. Avilov, and Vladimir I. Kalinin. 2015. "Anticancer Activity of Sea Cucumber Triterpene Glycosides" Marine Drugs 13, no. 3: 1202-1223. https://doi.org/10.3390/md13031202