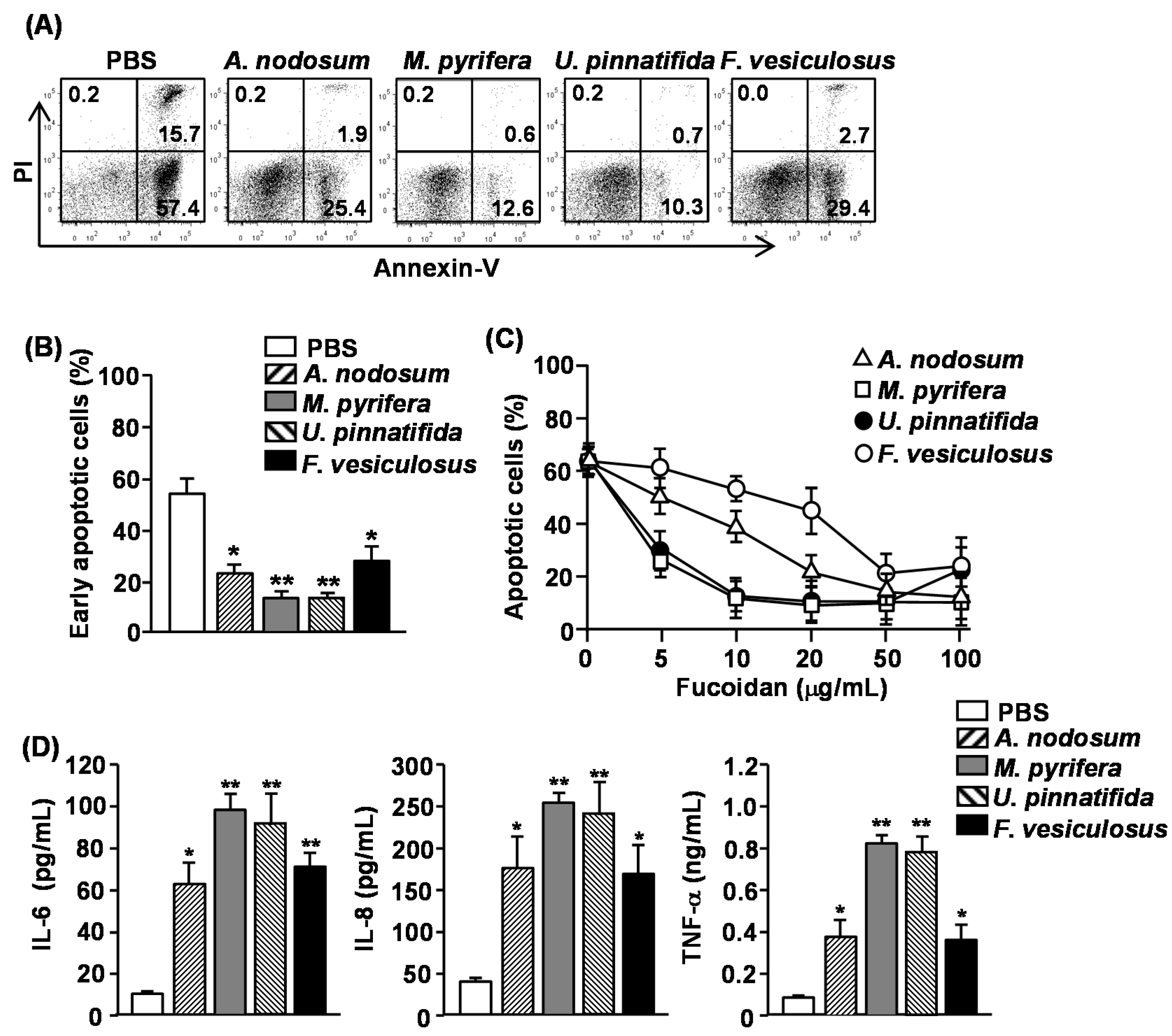
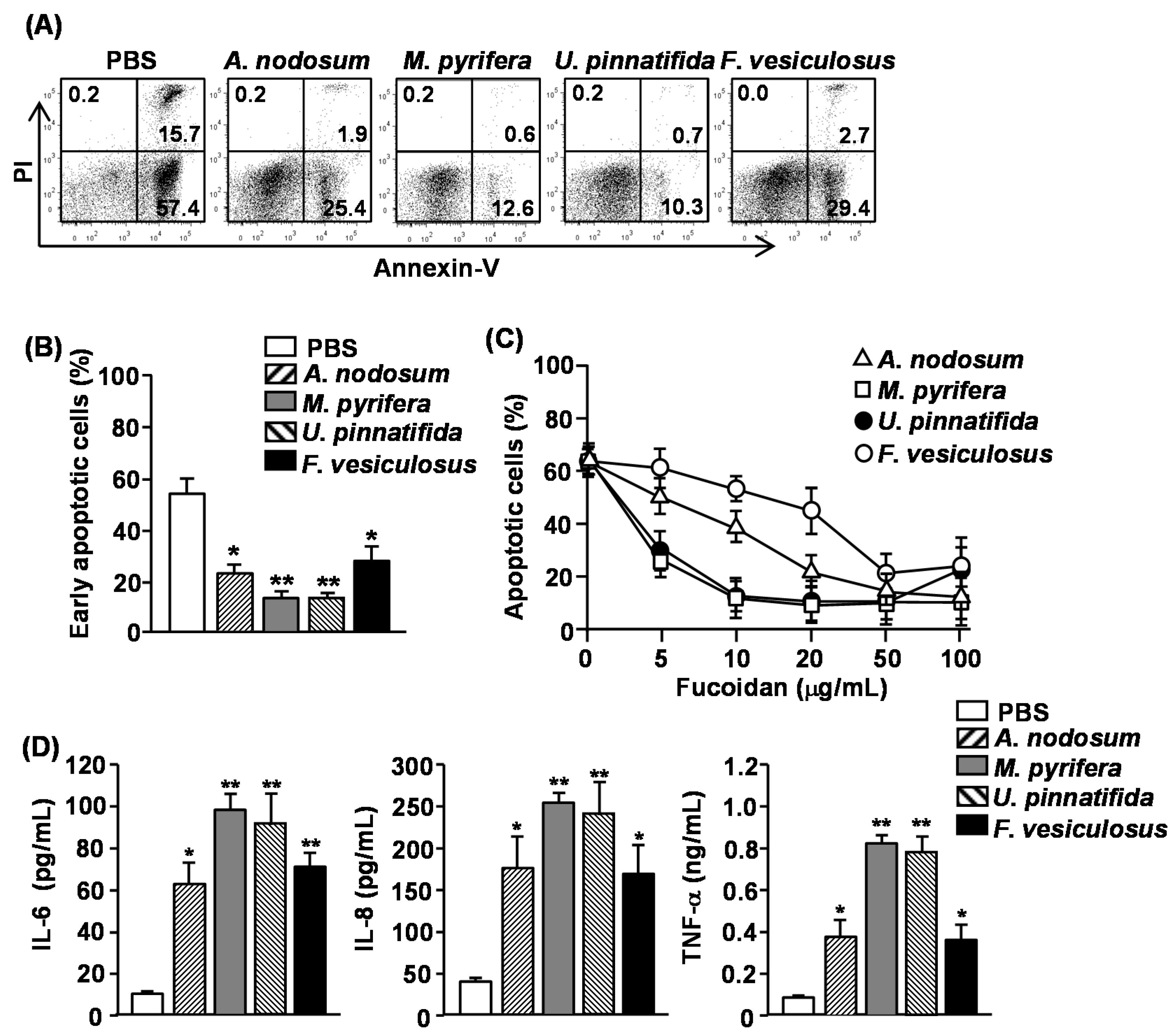
2.2. Effect of Fucoidans on Apoptosis and Pro-Inflammatory Cytokine Production in Human Neutrophils
Numerous studies have shown that neutrophil apoptosis can be delayed or accelerated by cytokines and other inflammatory mediators [
10,
11,
12]. Moreover, our recent study showed that fucoidans from
U. pinnatifida delayed neutrophil apoptosis [
13]. We therefore assessed whether other fucoidans also have similar effect on human neutrophils. Since a common marker of apoptotic cells is phosphatidyl serine (PS) exposure on the outer leaflet of the plasma membrane, we measured PS exposure by Annexin V staining and flow cytometry to identify the apoptotic cells. In addition, cells were simultaneously stained with propidium Iodide (PI) to identify the late-apoptotic or necrotic cells with membrane that is permeable to PI. Purified human neutrophils were incubated in the presence or absence of 50 μg/mL fucoidan and apoptosis was measured. More than 70% of neutrophils showed spontaneous apoptosis as indicated by positive Annexin V staining after 24 h of culture (
Figure 1A). All fucoidans markedly reduced the percentage of Annexin V
+PI
− cells, which were cells in the early stage of apoptosis, indicating that they inhibited apoptotic cell death (
Figure 1A,B). The most potent inhibitors were fucoidan from
M. pyrifera and
U. pinnatifida, which reduced neutrophil apoptosis by more than 80% as compared to controls, whereas fucoidans from
A. nodosum and
F. vesiculosus reduced neutrophil apoptosis by approximately 50%. We next assessed the dose-dependent effect of four different fucoidans on neutrophil apoptosis. Fucoidan from
M. pyrifera or
U. pinnatifida showed a considerable, dose-dependent inhibiting effect on neutrophil apoptosis at concentrations between 5–100 μg/mL, whereas those from
A. nodosum and
F. vesiculosus only prevented neutrophil apoptosis at concentration of 50–100 μg/mL (
Figure 1C).
Activated neutrophils can survive much longer than non-activated cells and can produce a number of inflammatory chemokines and cytokines [
10,
14,
15]. We therefore assessed whether fucoidan stimulation in human neutrophil can induce the production of pro-inflammatory cytokines from human neutrophils. Purified human neutrophils were treated with 50 μg/mL fucoidans. After 24 h of culture, the concentrations of interleukin (IL)-6, IL-8 and tumor necrosis factor-α (TNF-α) in the culture medium were measured. All fucoidans significantly increased the production of IL-6, IL-8 and TNF-α from neutrophils. Consistent with the delay of apoptosis, fucoidans from
M. pyrifera or
U. pinnatifida induced the highest amount of cytokine production. These data demonstrated that fucoidans delay the apoptosis and promote pro-inflammatory cytokine production in human neutrophils, and fucoidans from
M. pyrifera and
U. pinnatifida have the strongest effect on both apoptosis and cytokine production.
Figure 1.
Effect of fucoidans on spontaneous apoptosis and pro-inflammatory cytokine production of human neutrophils. Isolated peripheral blood neutrophils (2 × 105) were cultured with or without fucoidans (10 μg/mL) for 24 h. (A) Cell apoptosis was assessed by Annexin V-FITC and PI staining. Numbers in the plots show the percentage of the cells in the respective quadrant among the total cells shown in the plots; (B) Percentage of early apoptotic cells (Annexin V+PI− cells) was shown. Data are representative or the average of analyses of five samples from five donors for each group; (C) Dose-dependent delay of neutrophil apoptosis by fucoidans was measured by morphological changes. Data are the average of analyses of five samples from five donors; (D) IL-6, IL-8 and TNF-α concentrations in the cultured supernatants were measured by ELISA. Data are the average of analyses of 5 samples from 5 donors. Data shown are the mean ± SEM. * p < 0.05; ** p < 0.01 versus PBS (phosphate buffered saline) group.
Figure 1.
Effect of fucoidans on spontaneous apoptosis and pro-inflammatory cytokine production of human neutrophils. Isolated peripheral blood neutrophils (2 × 105) were cultured with or without fucoidans (10 μg/mL) for 24 h. (A) Cell apoptosis was assessed by Annexin V-FITC and PI staining. Numbers in the plots show the percentage of the cells in the respective quadrant among the total cells shown in the plots; (B) Percentage of early apoptotic cells (Annexin V+PI− cells) was shown. Data are representative or the average of analyses of five samples from five donors for each group; (C) Dose-dependent delay of neutrophil apoptosis by fucoidans was measured by morphological changes. Data are the average of analyses of five samples from five donors; (D) IL-6, IL-8 and TNF-α concentrations in the cultured supernatants were measured by ELISA. Data are the average of analyses of 5 samples from 5 donors. Data shown are the mean ± SEM. * p < 0.05; ** p < 0.01 versus PBS (phosphate buffered saline) group.
![Marinedrugs 13 01084 g001]()
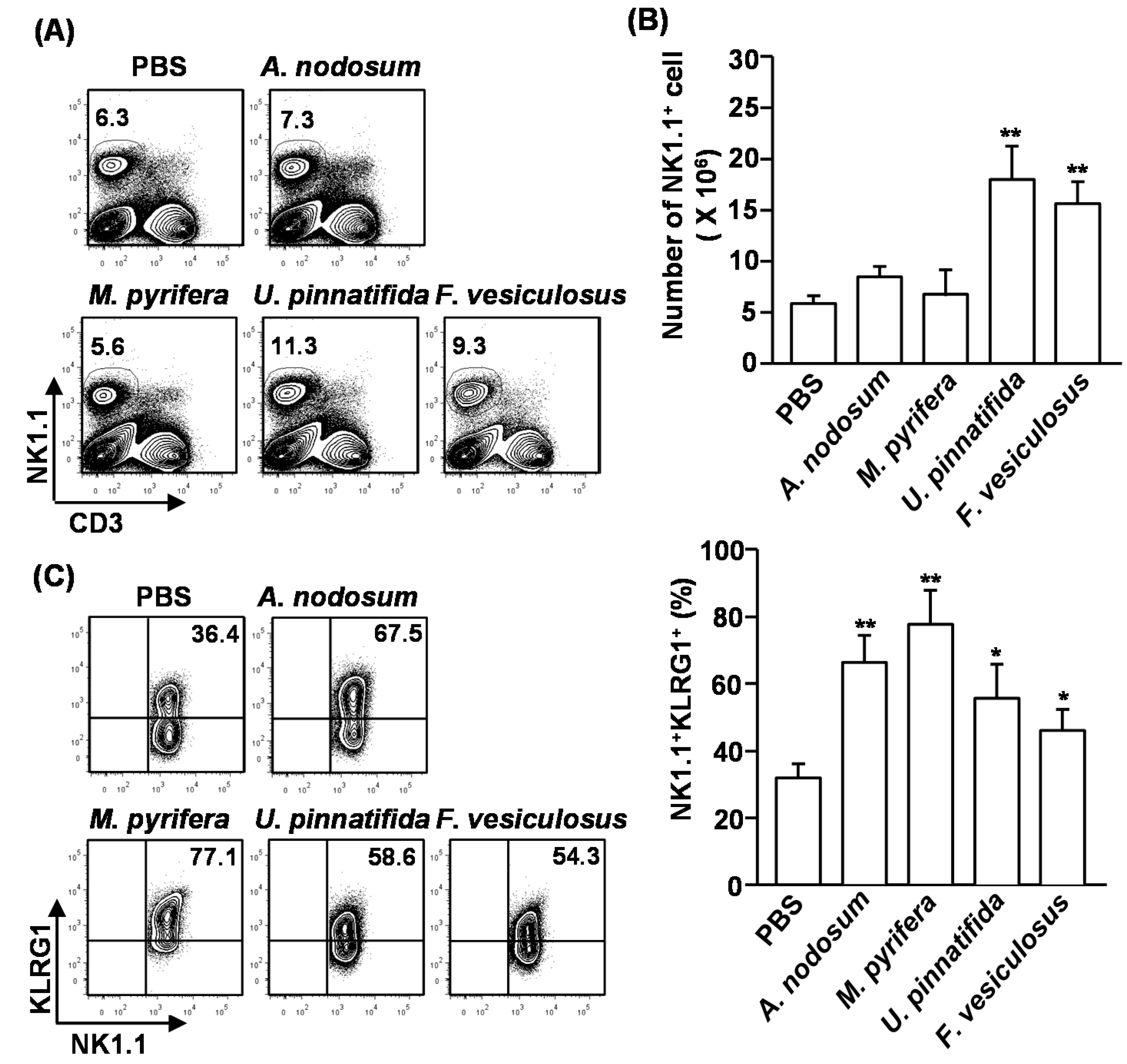
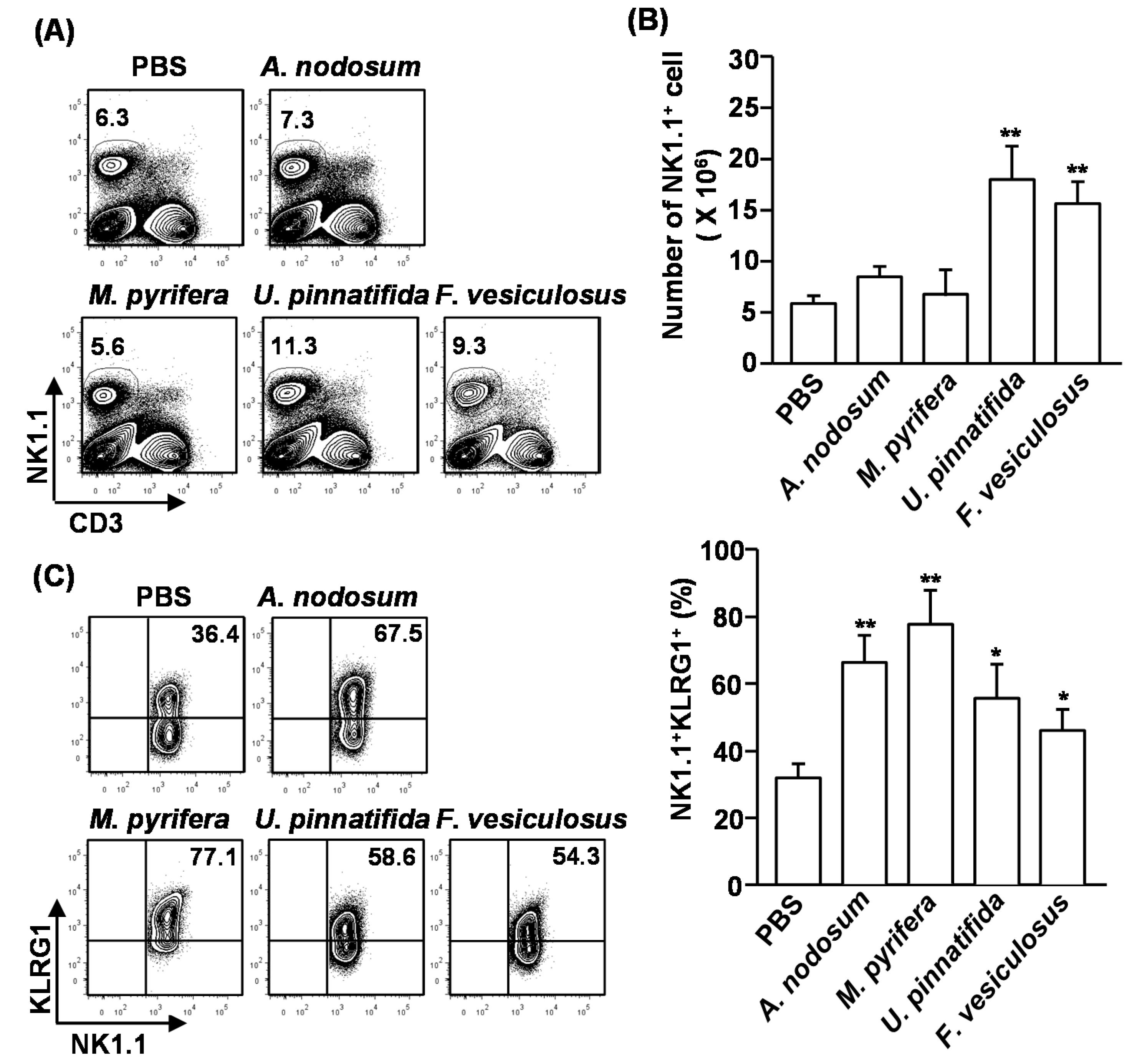
2.3. Effect of Fucoidans on the Activation and Cytotoxicity of NK Cells
NK cells play crucial roles in cell-mediated immunity and elimination of tumor cells. Since polysaccharide can induce activation and cytotoxicity on NK cells, we assessed whether fucoidans also can promote NK cell activation and cytotoxicity. Moreover, previous study showed 50 mg/kg of fucoidan from
F. vesiculosus induced NK cell activation mouse
in vivo [
16], we injected
intraperitoneally (
i.p.) C57BL/6 mice with 50 mg/kg of fucoidans for four successive days and the percentages and cell numbers of CD3
−NK1.1
+ cells in the spleen were measured. Upon administration of fucoidans from
U. pinnatifida and
F. vesiculosus, the percentages and cell numbers of CD3
−NK1.1
+ cells were significantly increased compared to the controls. In contrast, the other two fucoidans had no effect (
Figure 2A,B). Next, we examined the activation of NK cells after fucoidan injection. Killer cell lectin-like receptor subfamily G member 1 (KLRG1), which is the terminal differentiation and maturation marker in NK cells, was substantially up-regulated in NK cells that were treated with all type of fucoidans for 4 days (
Figure 2C). Moreover, intracellular interferon-γ (IFN-γ) levels, which is normally produced by activated NK cells, were markedly increased in NK cells by fucoidan treatment (
Figure 2D). Interestingly, fucoidan from
M. pyrifera showed the strongest effect on the maturation and activation of NK cells, although it did not increase the number of NK cells in spleen.
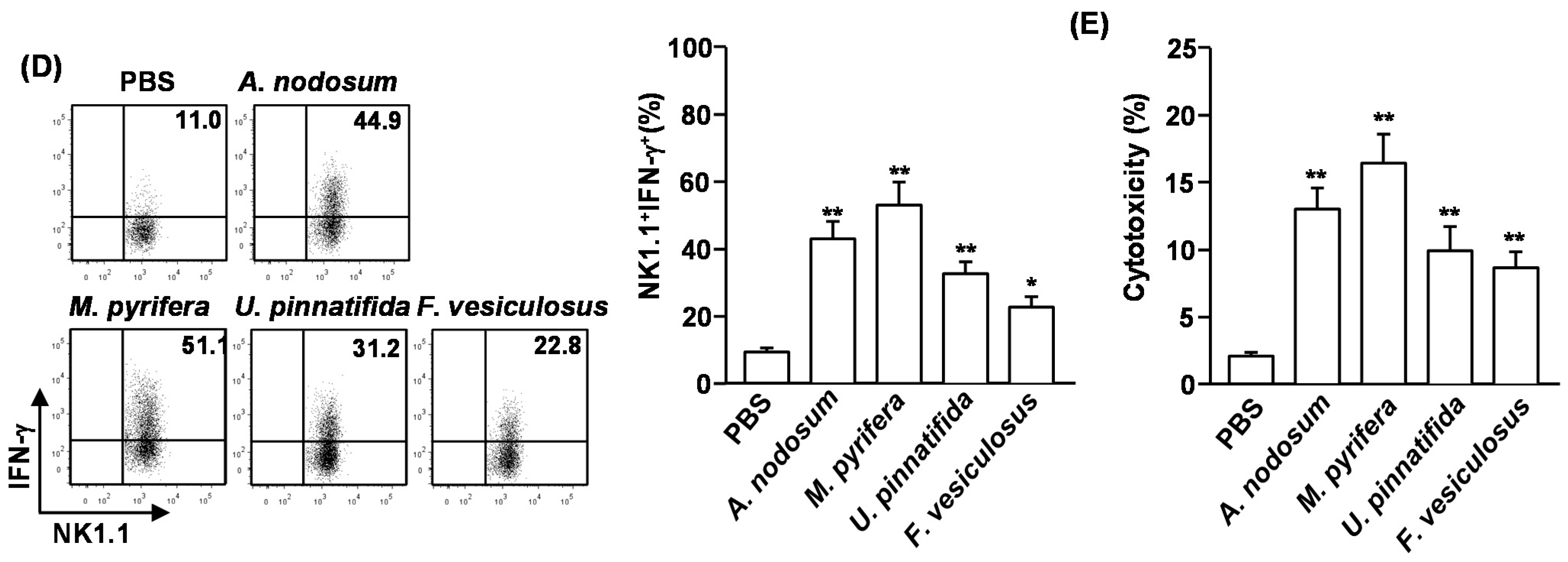
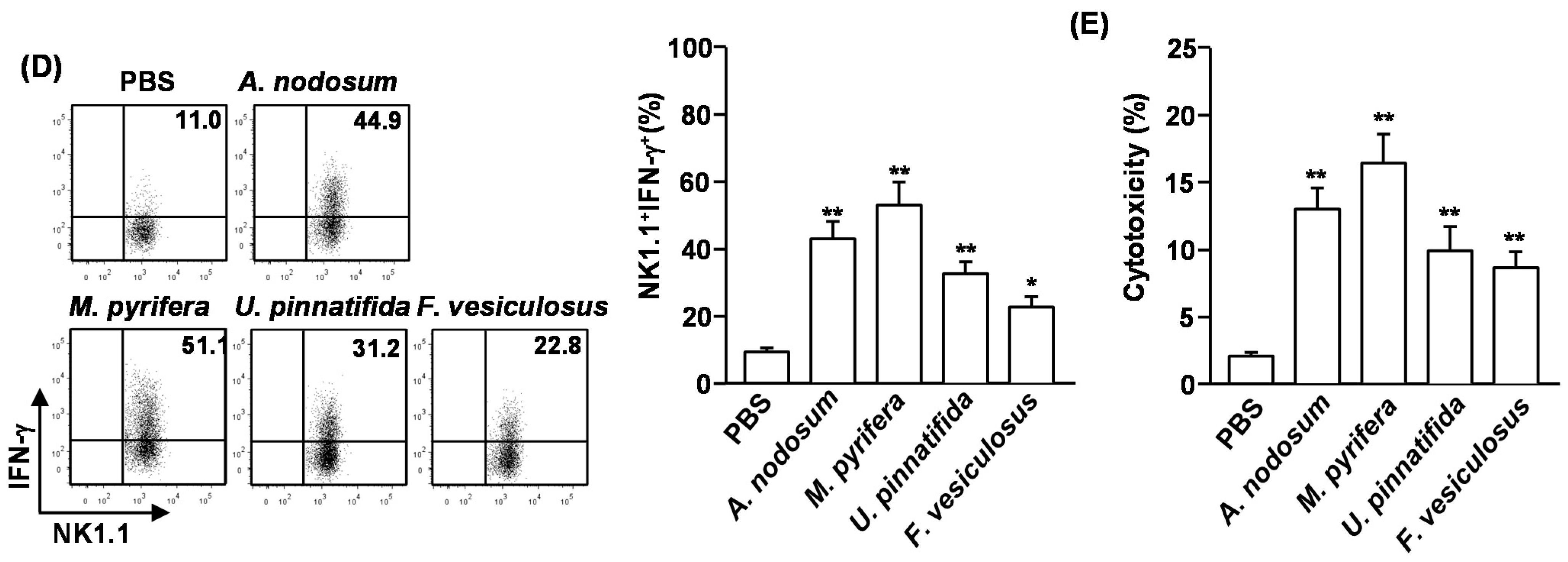
We then assessed whether systemic administration of fucoidans can induce cytotoxic activity in NK cells against YAC-1 cells, which are sensitive to lysis by activated murine NK cells. CD3
−NK1.1
+ cells were isolated from spleen of fucoidan-treated mice and co-cultured with YAC-1 cells for 4 h. All fucoidans induced significant increases in the cytotoxic activity of NK cells against YAC-1 cells. Consistent with the activation and maturation of NK cells, fucoidan from
M. pyrifera induced the highest cytotoxicity in NK cells against YAC-1 cells (
Figure 2E). These results indicate that systemic administration of fucoidans promotes NK cell activation and cytotoxicity.
Figure 2.
Fucoidans promote activation and maturation of mouse NK cells in vivo. C57BL/6 mice were injected with fucoidans (50 mg/kg) for four consecutive days. (A) Frequency of NK cells, defined as CD3−NK1.1+, in the spleen is indicated by the number shown in each plot; (B) Absolute number of NK cells in the spleen; (C) Expression levels of KLRG1 in CD3−NK1.1+ cells (left panel). Mean percentage of NK1.1+KLRG1+ cells (right panel); (D) Percentage of IFN-γ-producing cells among CD3−NK1.1+ cells in the spleen was assessed by flow cytometric analysis cells (left panel). Mean percentage of NK1.1+IFN-γ+ cells (right panel); (E) CD3−NK1.1+ cells were cultured with YAC-1 target cells at an E:T ration of 10:1 for 4 h. Percent specific lysis was measured as described at Method. All data are representative of or the average of analyses of six independent samples (2 mice per experiment, total 3 independent experiments). Data shown are the mean ± SEM. * p < 0.05; ** p < 0.01 versus PBS group.
Figure 2.
Fucoidans promote activation and maturation of mouse NK cells in vivo. C57BL/6 mice were injected with fucoidans (50 mg/kg) for four consecutive days. (A) Frequency of NK cells, defined as CD3−NK1.1+, in the spleen is indicated by the number shown in each plot; (B) Absolute number of NK cells in the spleen; (C) Expression levels of KLRG1 in CD3−NK1.1+ cells (left panel). Mean percentage of NK1.1+KLRG1+ cells (right panel); (D) Percentage of IFN-γ-producing cells among CD3−NK1.1+ cells in the spleen was assessed by flow cytometric analysis cells (left panel). Mean percentage of NK1.1+IFN-γ+ cells (right panel); (E) CD3−NK1.1+ cells were cultured with YAC-1 target cells at an E:T ration of 10:1 for 4 h. Percent specific lysis was measured as described at Method. All data are representative of or the average of analyses of six independent samples (2 mice per experiment, total 3 independent experiments). Data shown are the mean ± SEM. * p < 0.05; ** p < 0.01 versus PBS group.
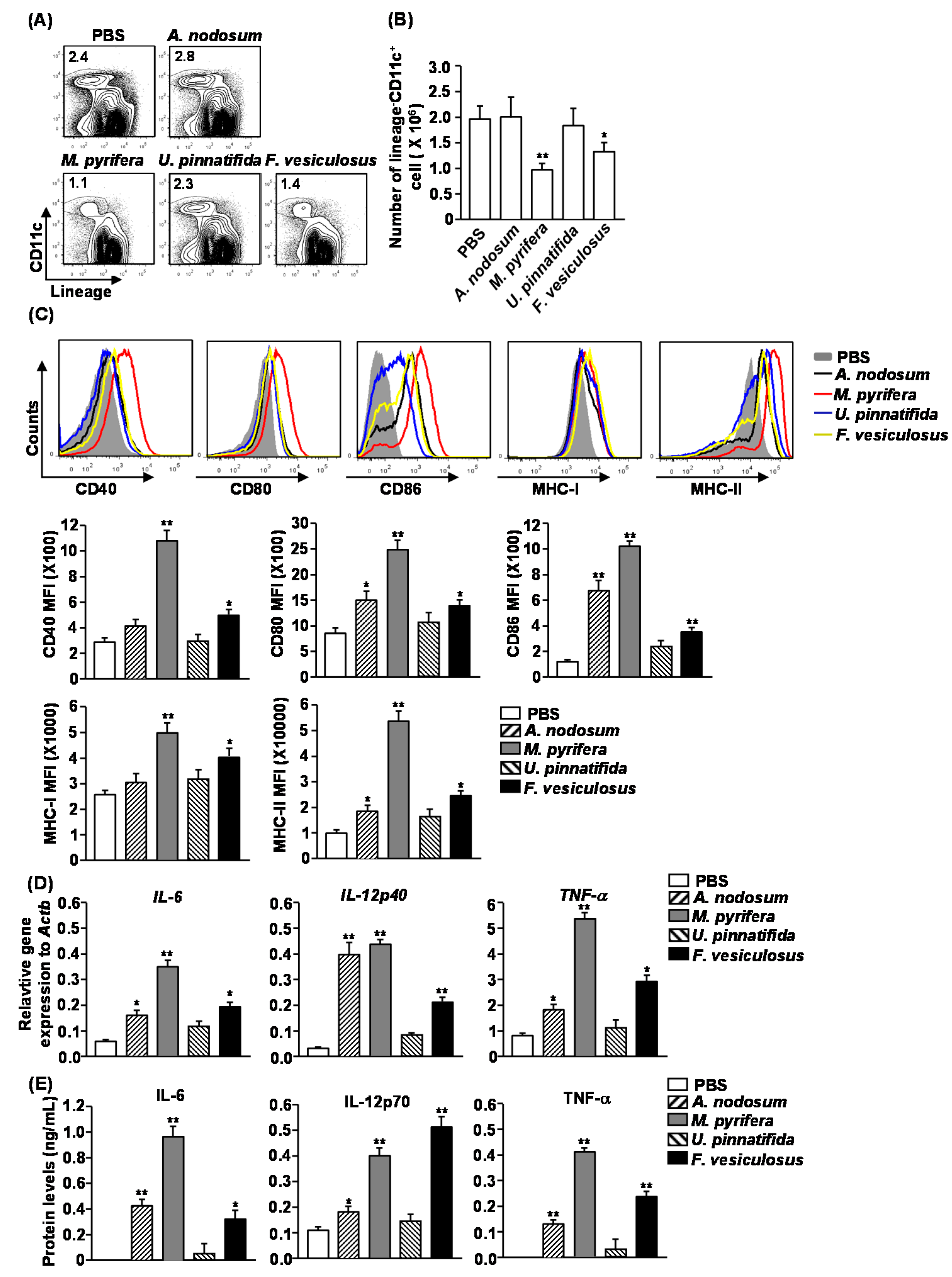
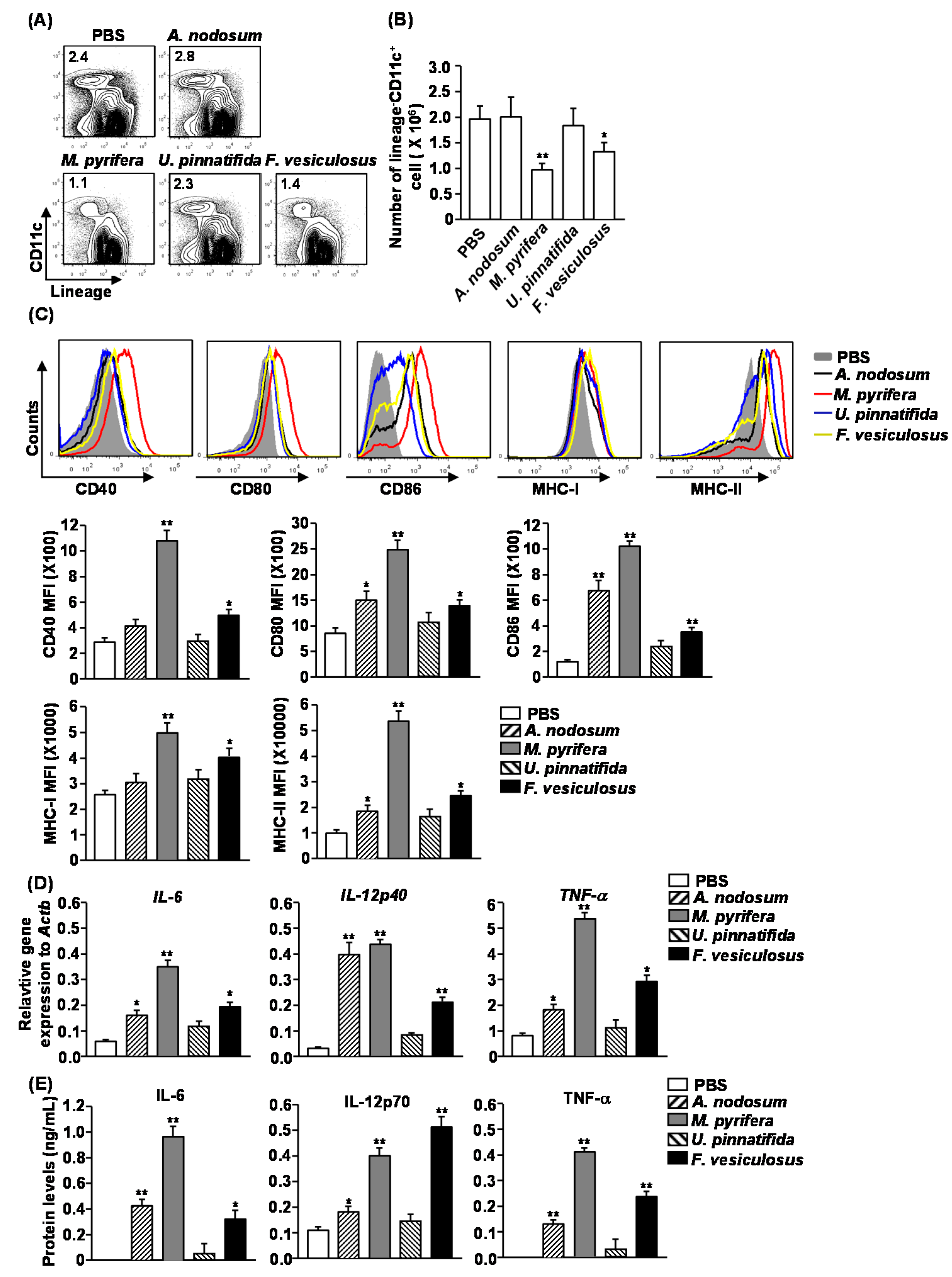
2.4. Effect of Fucoidans on the Activation and Maturation of Spleen Dendritic Cells (DCs) in Vivo
Our previous observation that fucoidan from
F. vesiculosus promotes spleen DC activation prompted us to investigate the effect of different fucoidans on spleen DC activation
in vivo [
4]. We injected 50 mg/kg fucoidans
intravenously (
i.v.) to C57BL/6 mice and analyzed spleen DCs 24 h later. Treatment with fucoidans from
M. pyrifera and
F. vesiculosus led to a significant decrease in the proportion and numbers of spleen DCs, which were identified as lineage
−CD11c
+ cells, whereas fucoidans from
A. nodosum and
U. pinnatifida had no significant effect (
Figure 3A,B). Moreover, all fucoidans induced substantial increases in the surface levels of CD80, CD86 and MHC class II in spleen DCs (
Figure 3C). The most active stimulator was fucoidan from
M. pyrifera, which induced up-regulation of CD40, CD80, CD86, MHC class I and MHC class II in spleen DCs (
Figure 3C). The least active compounds were fucoidans from
A. nodosum and
U. pinnatifida, which did not induce up-regulation of CD40 and MHC class I (
Figure 3C).
Figure 3.
Administration of fucoidans induces spleen DC activation. C57BL/6 mice were injected i.v. with 50 mg/kg fucoidans for 24 h. (A) Percentage of DCs, defined as lineage−CD11c+, was analyzed by flow cytometry; (B) Absolute number of lineage−CD11c+ cells within live cells was shown; (C) Expression levels of CD40, CD80, CD86, MHC class I and MHC class II were measured by flow cytometry (upper panel). Mean fluorescence intensity (MFI) of these molecules is shown (lower panel). Data are representative of or the average of analyses of six independent samples (two mice per experiment, total of three independent experiments); (D) Expression levels of IL-6, IL-12p40 and TNF-α mRNA were measured from spleen 2 h after 50 mg/kg fucoidans injection; (E) IL-6, IL-12p70 and TNF-α concentrations in sera were shown from fucoidan-treated mice at 24 h after treatment. Data are representative of or the average of analyses of six independent samples (two mice per experiment, total of three independent experiments). Data shown are the mean ± SEM. * p < 0.05; ** p < 0.01 versus PBS group.
Figure 3.
Administration of fucoidans induces spleen DC activation. C57BL/6 mice were injected i.v. with 50 mg/kg fucoidans for 24 h. (A) Percentage of DCs, defined as lineage−CD11c+, was analyzed by flow cytometry; (B) Absolute number of lineage−CD11c+ cells within live cells was shown; (C) Expression levels of CD40, CD80, CD86, MHC class I and MHC class II were measured by flow cytometry (upper panel). Mean fluorescence intensity (MFI) of these molecules is shown (lower panel). Data are representative of or the average of analyses of six independent samples (two mice per experiment, total of three independent experiments); (D) Expression levels of IL-6, IL-12p40 and TNF-α mRNA were measured from spleen 2 h after 50 mg/kg fucoidans injection; (E) IL-6, IL-12p70 and TNF-α concentrations in sera were shown from fucoidan-treated mice at 24 h after treatment. Data are representative of or the average of analyses of six independent samples (two mice per experiment, total of three independent experiments). Data shown are the mean ± SEM. * p < 0.05; ** p < 0.01 versus PBS group.
![Marinedrugs 13 01084 g003]()
To determine whether fucoidans affect production of cytokines, we injected C57BL/6 mice with fucoidans and analyzed the production of pro-inflammatory cytokines after 2 or 24 h.
In vivo administration of all fucoidans, except for that from
U. pinnatifida, caused a marked increase in mRNA levels of IL-6, IL-12p40 and TNF-α in splenocytes 2 h post-injection compared to PBS-treated control mice (
Figure 3D). Moreover, serum levels of IL-6, IL-12p70 and TNF-α were also dramatically increased in mice treated with fucoidans, except for those treated with fucoidan from
U. pinnatifida (
Figure 3E). Among the fucoidans, fucoidan from
M. pyrifera had the strongest effect on promoting cytokine production. Therefore based on both the induction of co-stimulatory molecules and cytokine production, fucoidan from
M. pyrifera was the most potent inducer of spleen DC activation and maturation
in vivo.
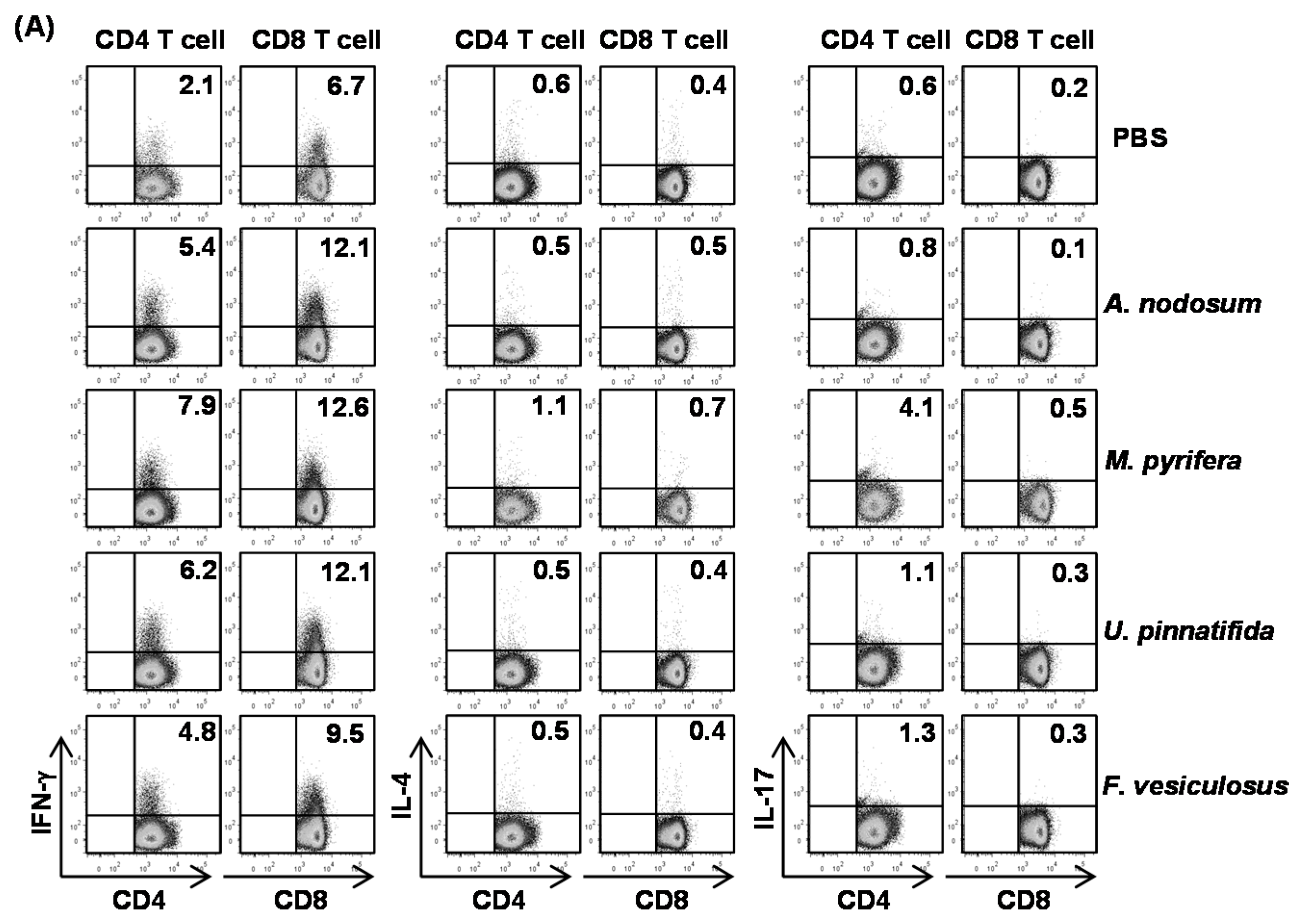
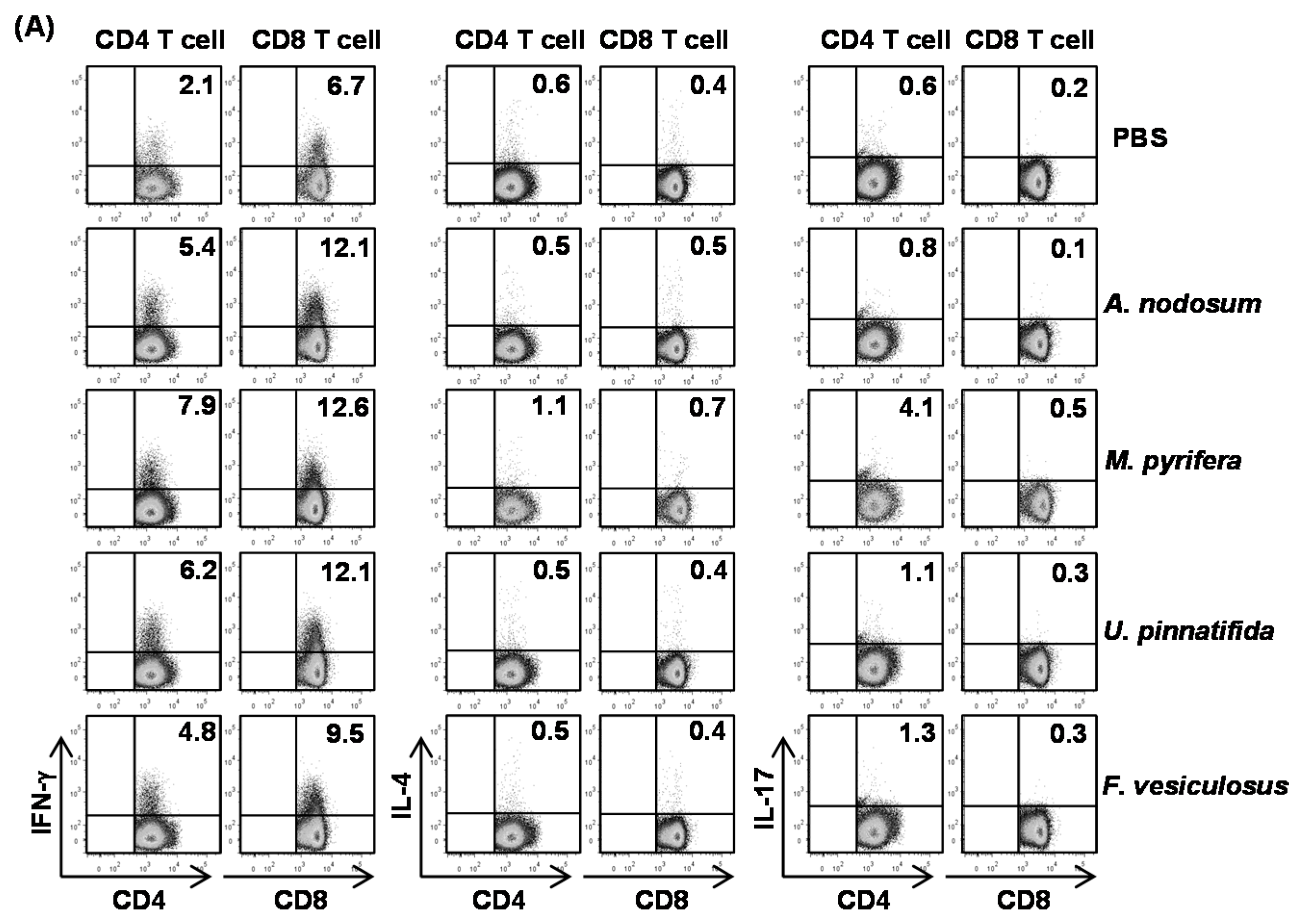
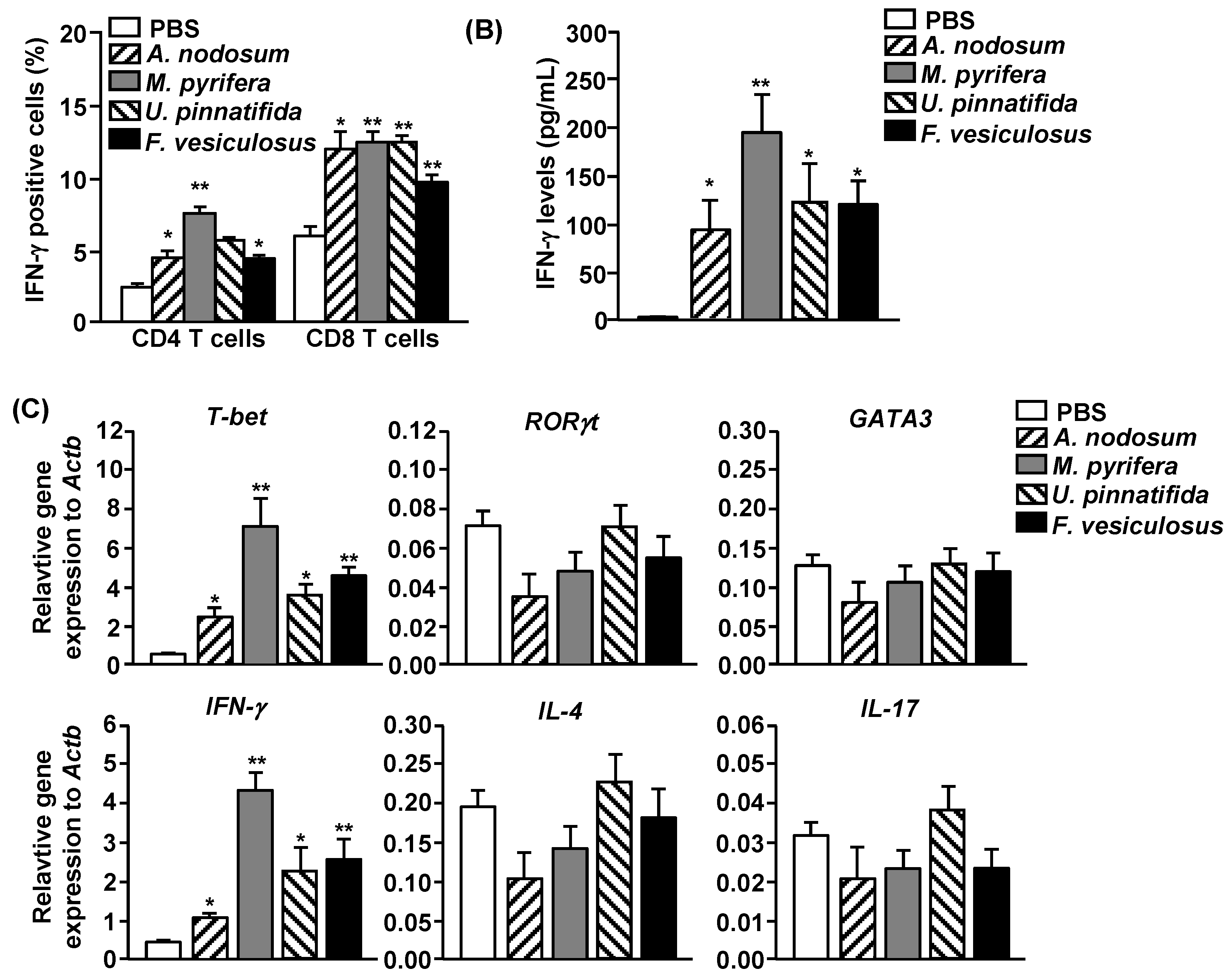
2.5. Effect of Fucoidans on T Cell Activation and Differentiation in Vivo
To determine whether fucoidans can have an impact on T cell responses, we
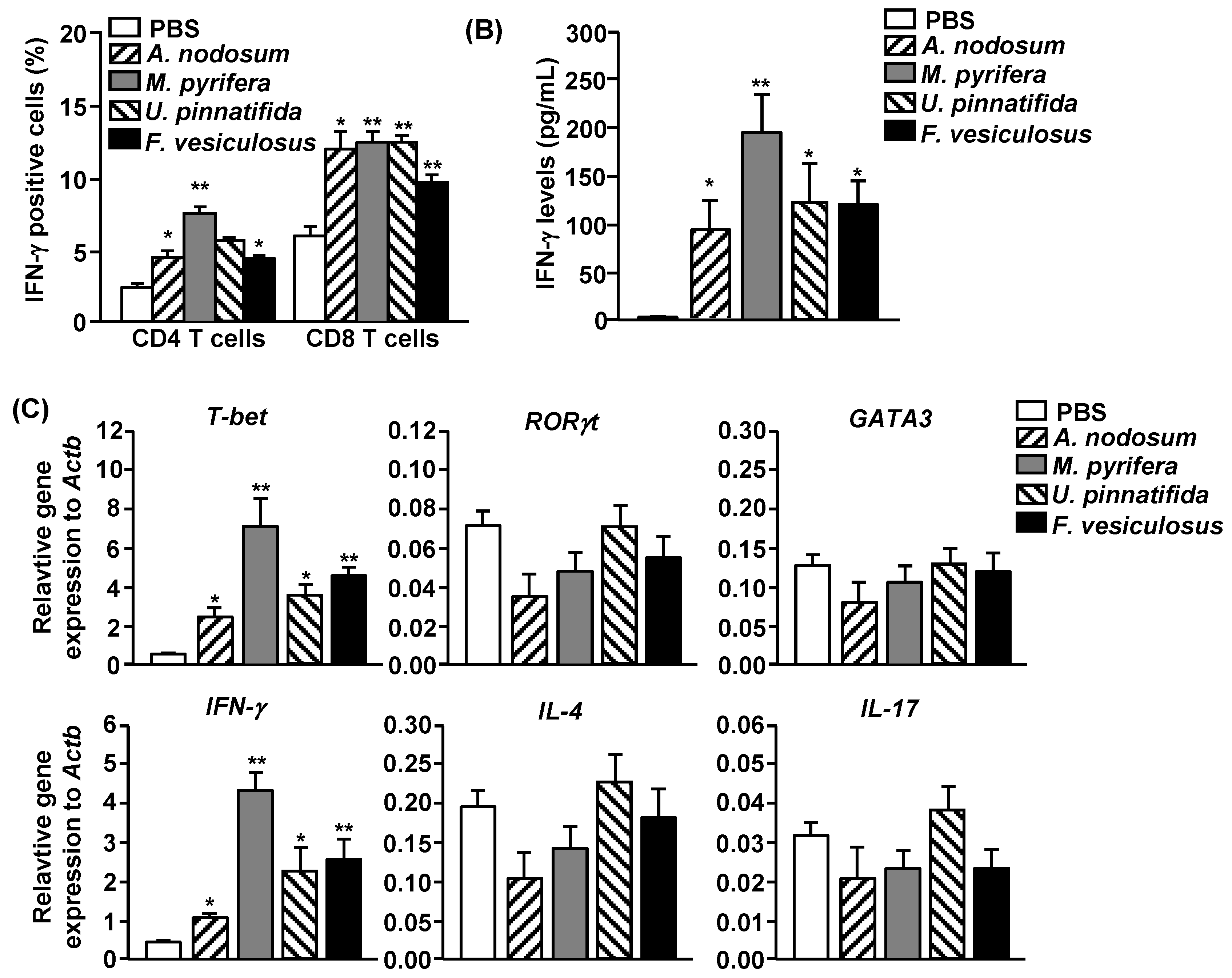
i.p. injected 20 mg/kg fucoidans to C57BL/6 and three days later, injected the same amount of fucoidans again. All fucoidan treatment led to marked increases in the proportions of splenic CD4 and CD8 T cells that produced IFN-γ and TNF-α, the signature cytokines of Th1 and Tc1 cells, as compared to PBS treatment (
Figure 4A). In contrast, the percentages of IL-17- or IL-4-producing CD4 and CD8 T cells in the spleen were not significantly increased by fucoidans (
Figure 4A). Serum levels of IFN-γ and TNF-α were also markedly increased by fucoidan (
Figure 4B). Moreover, fucoidan-treated mice had significantly higher mRNA amounts of T-bet, the critical transcription factor for Th1 and Tc1 cells, and IFN-γ in the spleen than control mice (
Figure 4C). In contrast, the mRNA levels of GATA3 and RORγt, transcription factor for Th2 and Th17, were not changed by fucoidan treatment (
Figure 4C). Consistent with its effect on DC activation and maturation, fucoidan from
M. pyrifera showed the strongest effect among the four fucoidans on T cell activation
in vivo. Taken together, these results showed that fucoidan from
M. Pyrifera is the most active immune modulator of NK cell, DC and T cell activation and function among the four fucoidans tested.
Figure 4.
Fucoidans treatment increases IFN-γ-producing CD4 and CD8 T cells in vivo. C57BL/6 mice were injected i.p. with 50 mg/kg fucoidans and 3 days later, injected again with same amount of fucoidans. (A) Percentage of IFN-γ, IL-4 and IL-17 positive cells within CD4 and CD8 T cells, identified by gating on the CD4+CD8− and CD4−CD8+ cells respectively, in the spleen was assessed by flow cytometric analysis (upper panel). Mean percentage of IFN-γ+ cells (lower panel); (B) IFN-γ concentrations in the serum. All data are representative of or the average of analyses of six independent samples (two mice per experiment, total of three independent experiments); (C) Gene expression in spleens was measured 24 h after fucoidan injection. Data are the average of analyses of six independent samples (two mice per experiment, total of three independent experiments). Data shown are the mean ± SEM. * p < 0.05; ** p < 0.01 versus PBS group.
Figure 4.
Fucoidans treatment increases IFN-γ-producing CD4 and CD8 T cells in vivo. C57BL/6 mice were injected i.p. with 50 mg/kg fucoidans and 3 days later, injected again with same amount of fucoidans. (A) Percentage of IFN-γ, IL-4 and IL-17 positive cells within CD4 and CD8 T cells, identified by gating on the CD4+CD8− and CD4−CD8+ cells respectively, in the spleen was assessed by flow cytometric analysis (upper panel). Mean percentage of IFN-γ+ cells (lower panel); (B) IFN-γ concentrations in the serum. All data are representative of or the average of analyses of six independent samples (two mice per experiment, total of three independent experiments); (C) Gene expression in spleens was measured 24 h after fucoidan injection. Data are the average of analyses of six independent samples (two mice per experiment, total of three independent experiments). Data shown are the mean ± SEM. * p < 0.05; ** p < 0.01 versus PBS group.
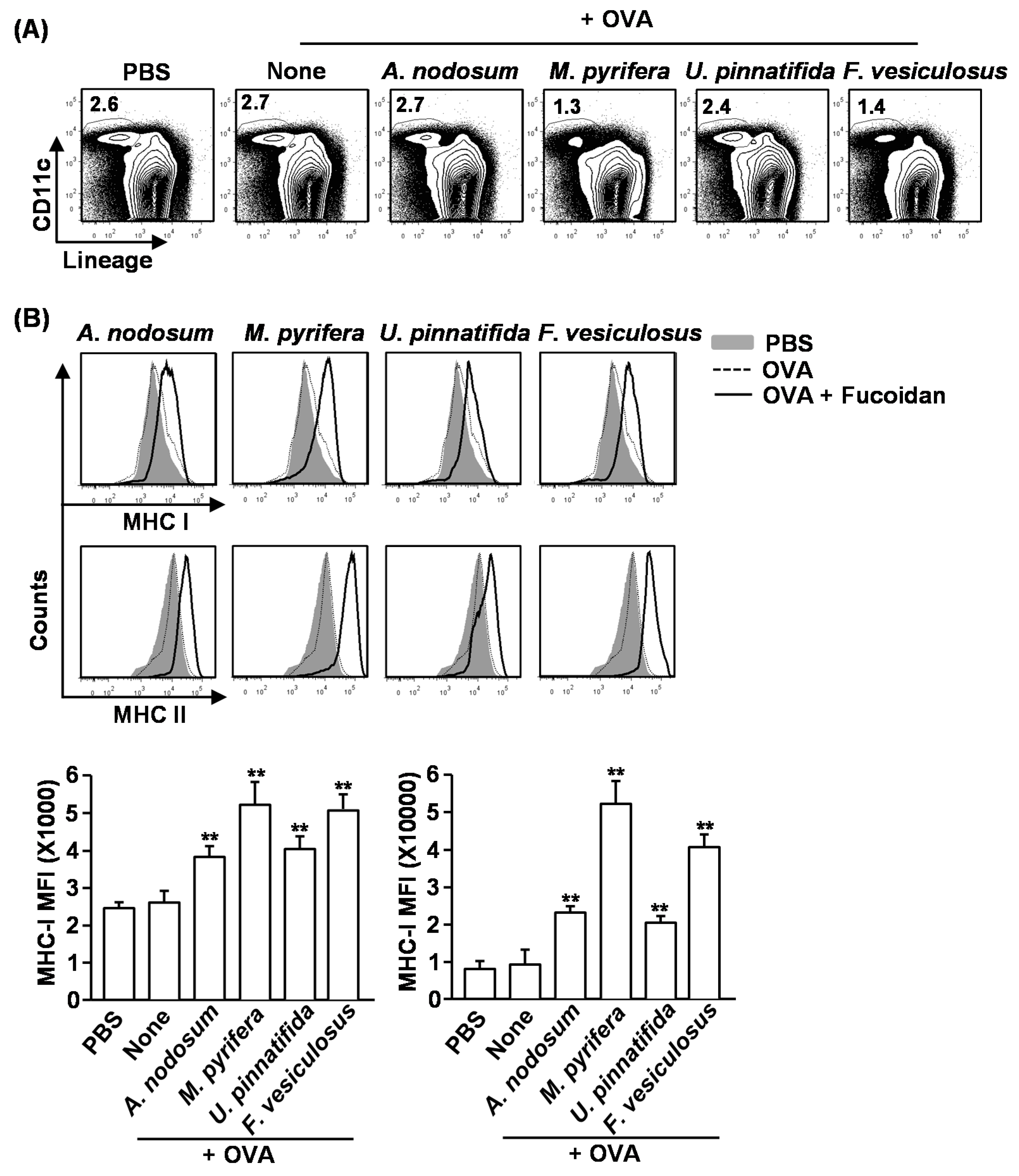
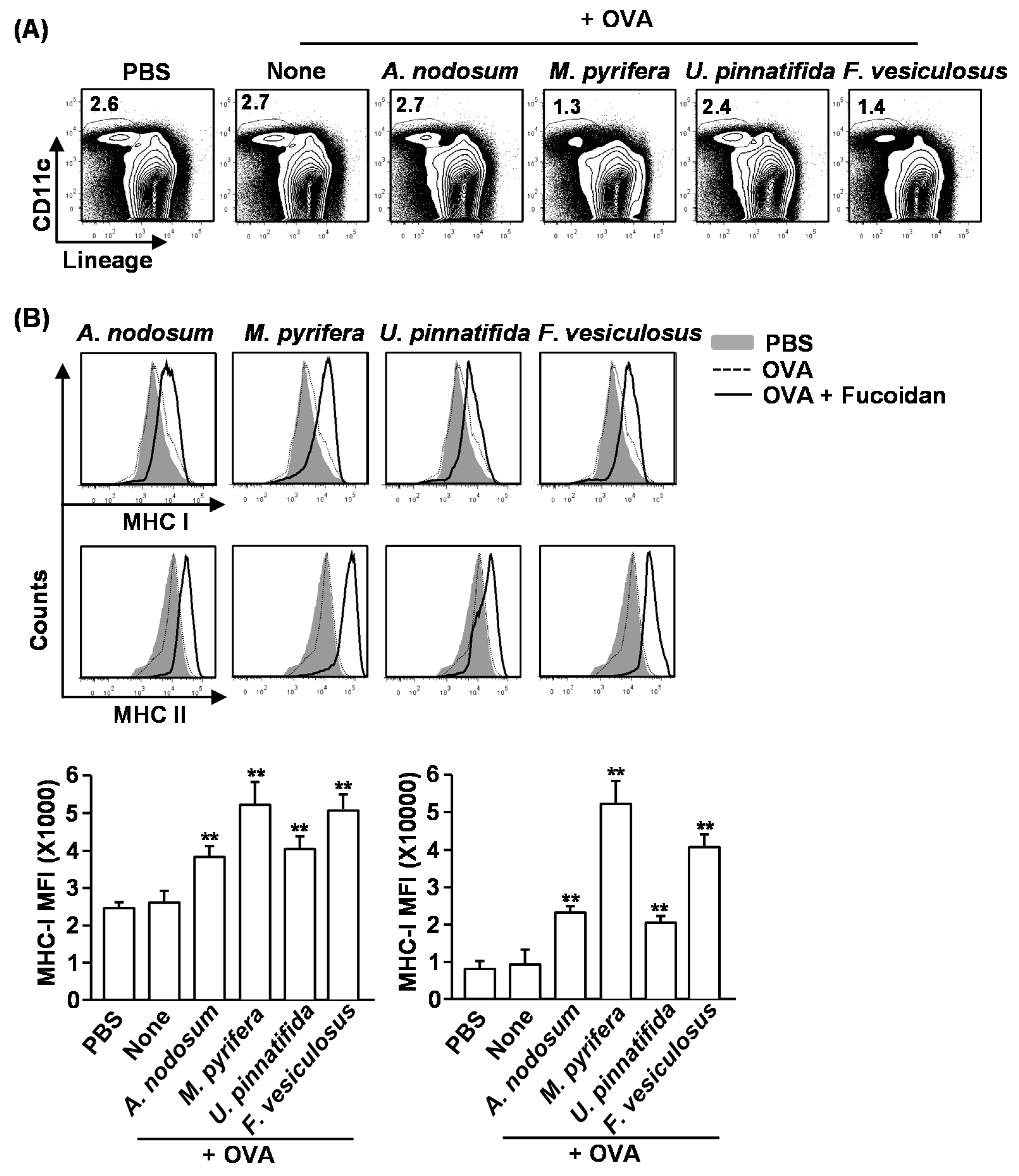
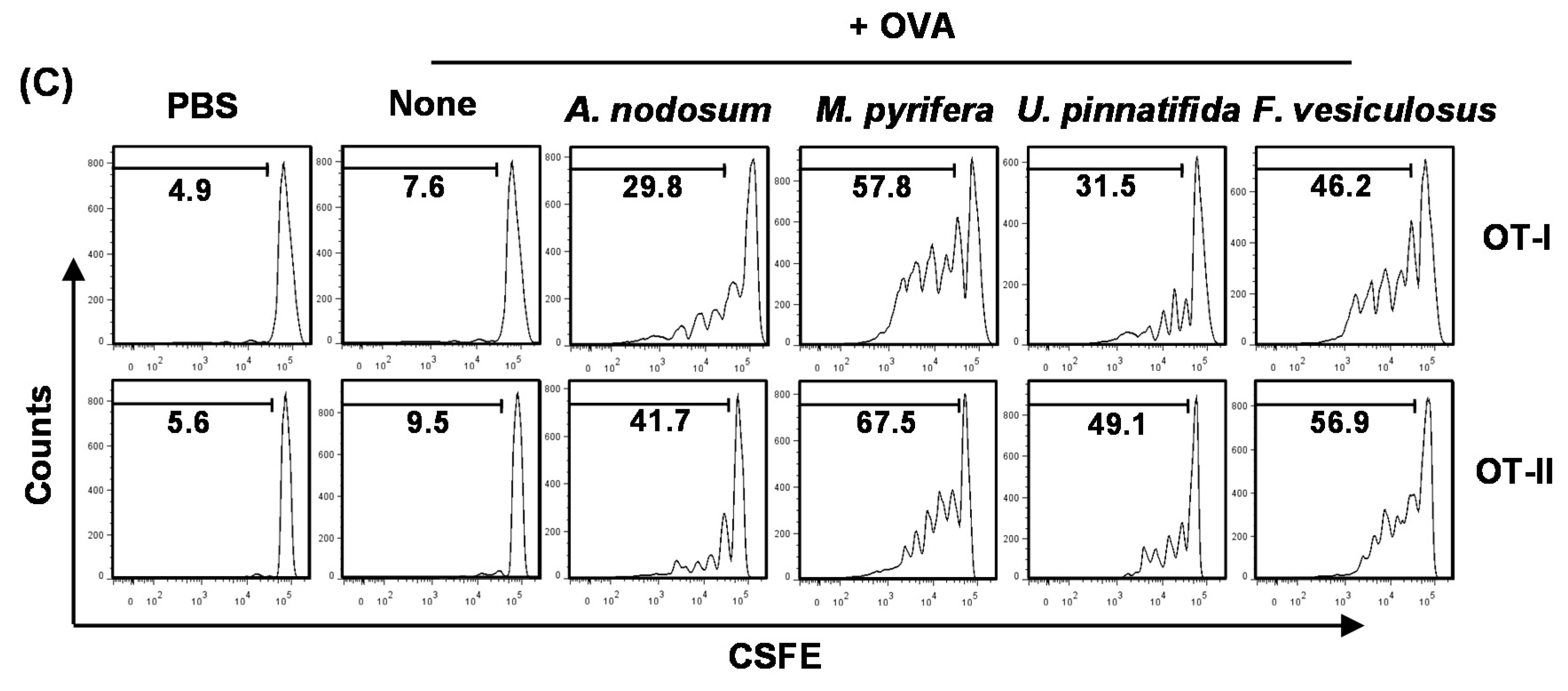
2.6. Effect of Fucoidans on Antigen Presentation by DCs and Antigen-Specific T Cell Proliferation
Our finding that fucoidans induce spleen DC and T cell activation
in vivo prompted us to further investigate the adjuvant effect of fucoidan in antigen-specific T cell response
in vivo. We first examined whether fucoidans can promote antigen-presentation by DCs. Mice were injected with PBS, ovalbumin (OVA) or OVA + fucoidans for 24 h, and then measured for frequency of DCs and expression of MHC classes I and II on spleen lineage
−CD11c
+ DCs. Consistent with the results shown in
Figure 3, the frequency of spleen CD11c
+ DCs was dramatically decreased by injection of fucoidans from
M. pyrifera or
F. vesiculosus (
Figure 5A). Moreover, all fucoidans promoted up-regulation of MHC class I and II expression on CD11c
+ DCs (
Figure 5B). As expected, fucoidan from
M. pyrifera induced the greatest up-regulation of these surface proteins among the four fucoidans (
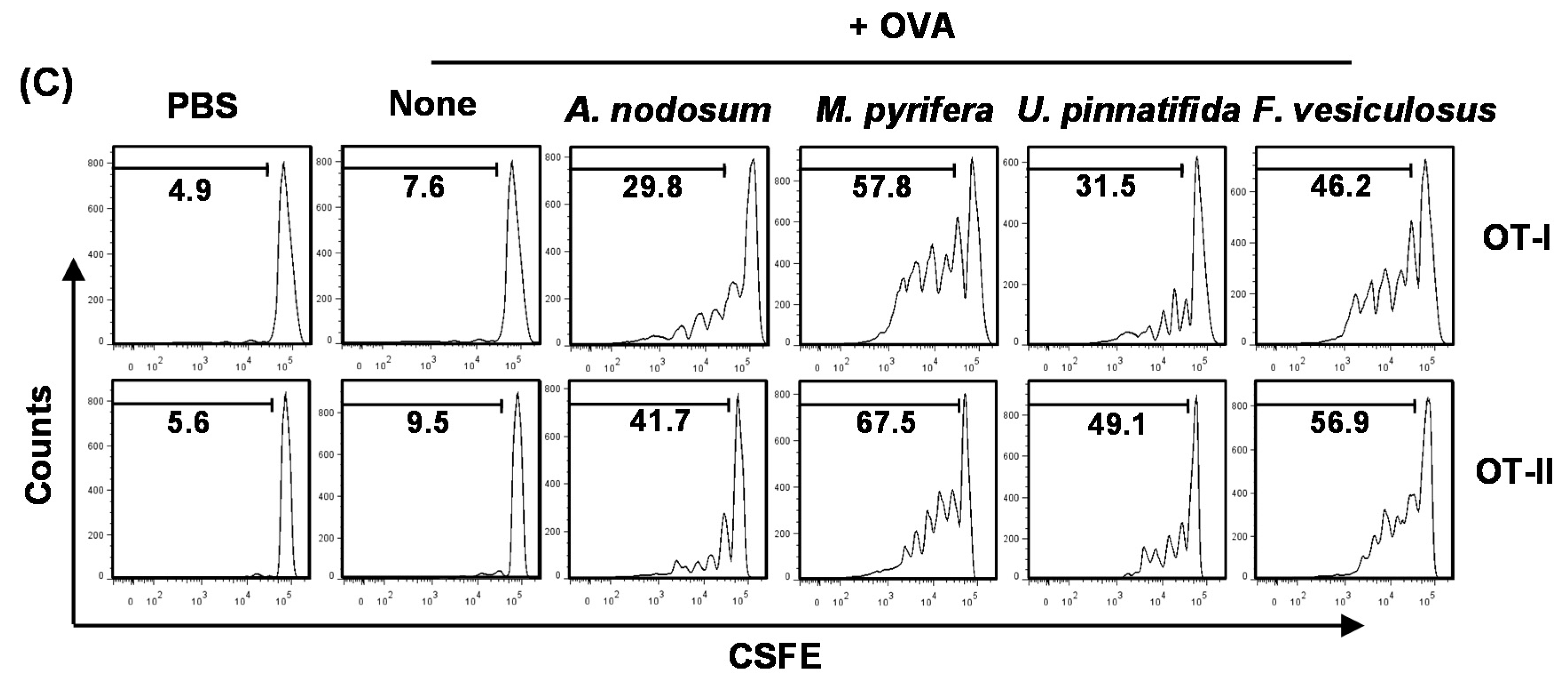
Figure 5B). Next, we performed an adoptive transfer experiment to detect OVA-specific OT-I and OT-II T cell proliferation. CFSE-labeled OT-I CD8
+ or OT-II CD4
+ T cells from CD45.2 TCR-transgenic mice were transferred into CD45.1 congenic mice, and 24 h later, the mice received injection of PBS, OVA or OVA + fucoidans. After 3 days, the proliferation of CD45.2
+ OT-I or OT-II cells was determined by CFSE dilution assay. OT-I and OT-II T cell proliferation was robustly increased in mice immunized with OVA + fucoidans compared to those in mice immunized with OVA alone (
Figure 5C). Consistent with its effect on DC activation and maturation, fucoidan from
M. pyrifera stimulated the strongest antigen-specific T cell proliferation. These data demonstrated that fucoidan functions as an adjuvant to enhance antigen presentation and antigen-specific CD4 and CD8 T cell activation. Moreover, fucoidan from
M. pyrifera is the most effective adjuvant
in vivo for antigen presentation among the four fucoidans tested.
Figure 5.
Fucoidans promote antigen-specific T cell proliferation in vivo. C57BL/6 mice were injected with PBS, ovalbumin (OVA) or OVA + fucoidan for 24 h. (A) Frequency of DCs, defined as lineage−CD11c+, were analyzed by flow cytometry; (B) Expression levels of MHC class I and II on the gated lineage−CD11c+ DCs in the spleen (upper panel). MFI of MHC class I and MHC class II is shown (lower panels); (C) Purified CD8 T cells from OT-I or CD4 T cells from OT-II mice were labeled with CFSE and transferred into CD45.1 congenic mice, and 24 h later, mice were injected with PBS, OVA or OVA + fucoidans. After 3 day treatment, splenocytes from these mice were stained for CD45.2 to identify the donor OT-I or OT-II cells and the proliferation of these cells was determined by CFSE dilution. All data are from analyses of six individual mice each group (two mice per experiment, total of three independent experiments). Data shown are the mean ± SEM. * p < 0.05; ** p < 0.01 versus OVA group.
Figure 5.
Fucoidans promote antigen-specific T cell proliferation in vivo. C57BL/6 mice were injected with PBS, ovalbumin (OVA) or OVA + fucoidan for 24 h. (A) Frequency of DCs, defined as lineage−CD11c+, were analyzed by flow cytometry; (B) Expression levels of MHC class I and II on the gated lineage−CD11c+ DCs in the spleen (upper panel). MFI of MHC class I and MHC class II is shown (lower panels); (C) Purified CD8 T cells from OT-I or CD4 T cells from OT-II mice were labeled with CFSE and transferred into CD45.1 congenic mice, and 24 h later, mice were injected with PBS, OVA or OVA + fucoidans. After 3 day treatment, splenocytes from these mice were stained for CD45.2 to identify the donor OT-I or OT-II cells and the proliferation of these cells was determined by CFSE dilution. All data are from analyses of six individual mice each group (two mice per experiment, total of three independent experiments). Data shown are the mean ± SEM. * p < 0.05; ** p < 0.01 versus OVA group.
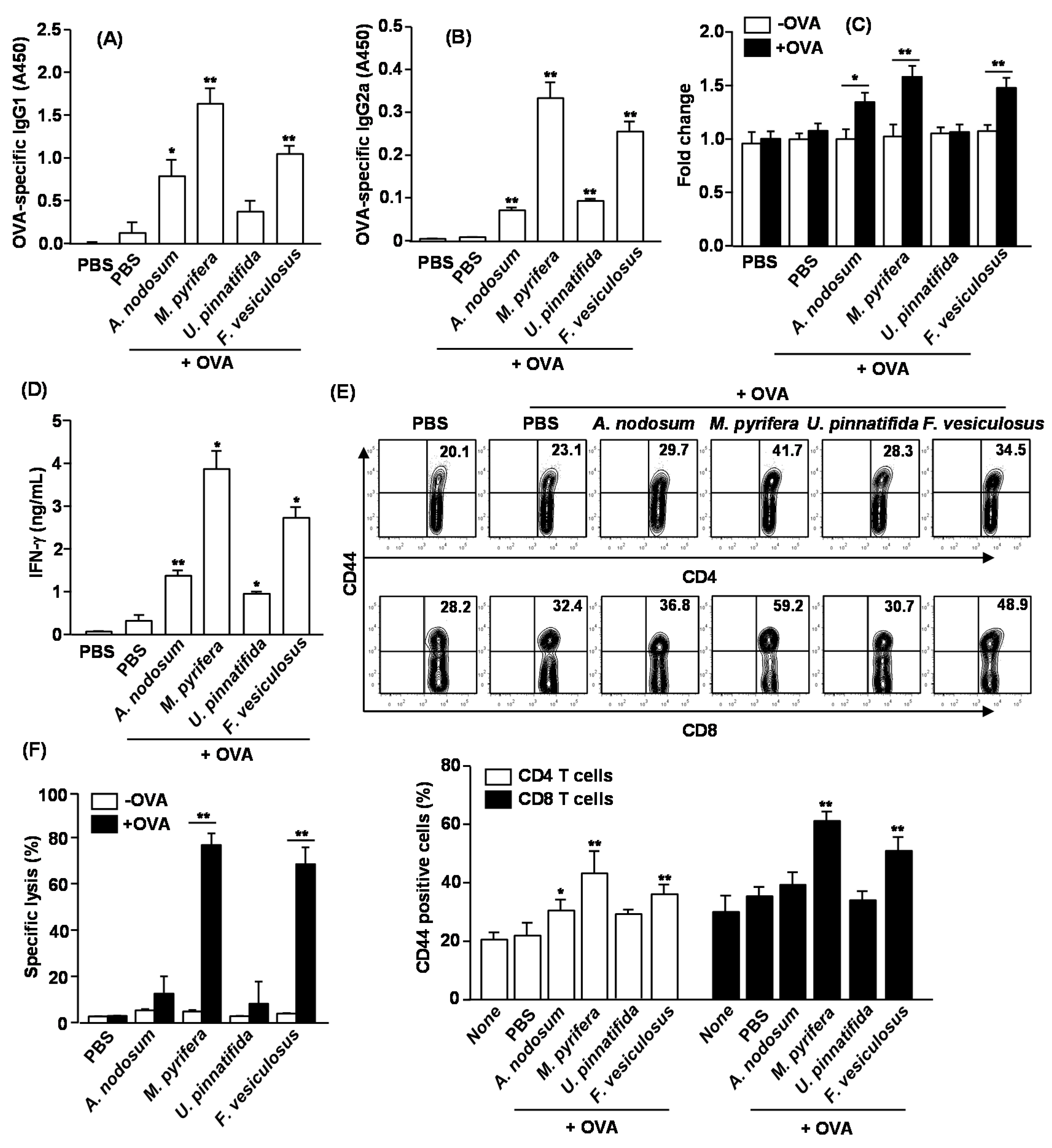
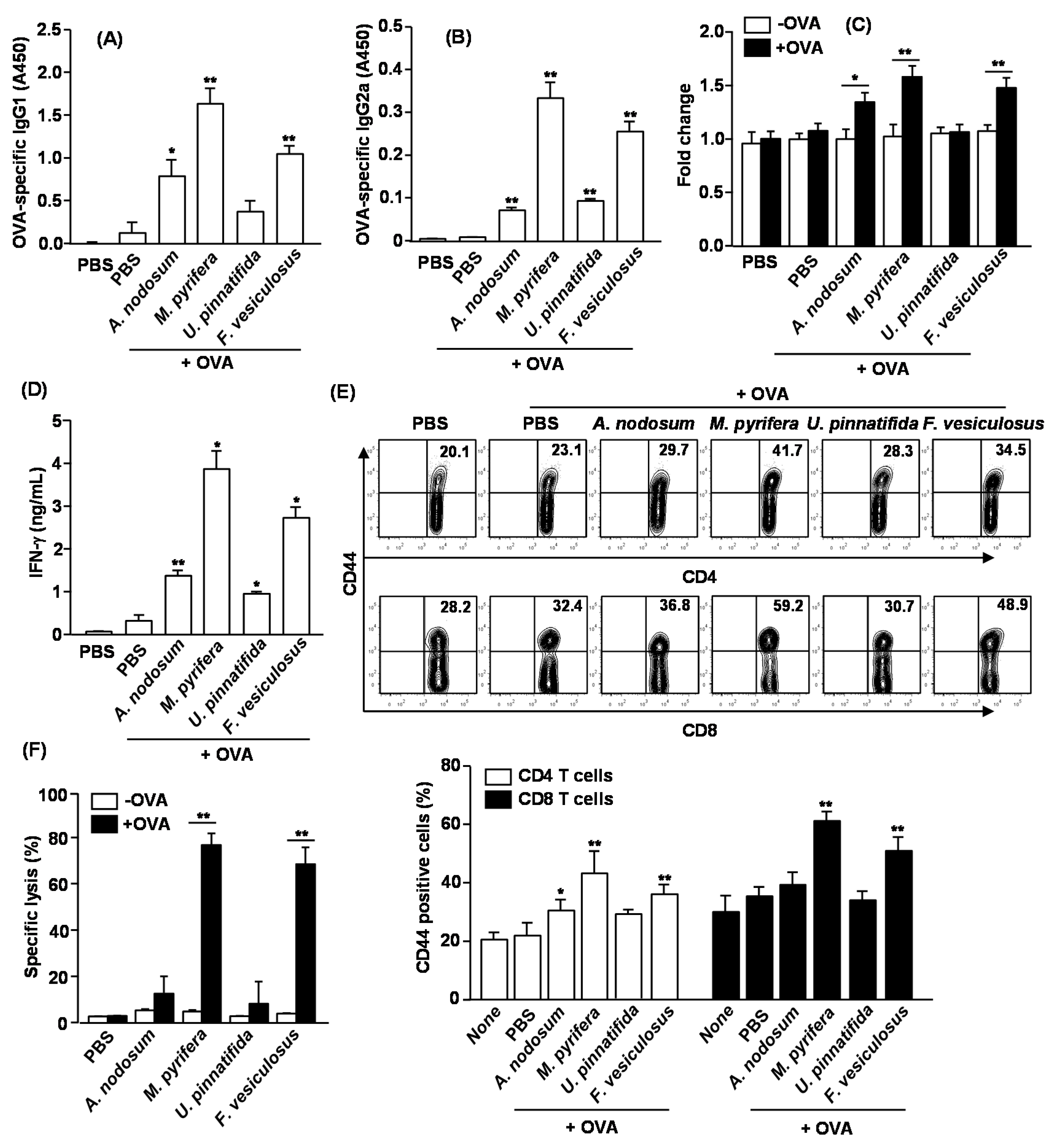
2.7. Adjuvant Effect of Fucoidans on Antigen-Specific Immune Responses in Vivo
To determine whether fucoidans exhibit adjuvant effect
in vivo and which fucoidan is most effective, we immunized C57BL/6 mice with OVA and fucoidans, and examined the adjuvant effect including specific antibody production and T cell responses against OVA. C57BL/6 mice were injected
i.p. with OVA alone or in combination with 50 mg/kg fucoidan on day 0, 14 and 28. On Day 33, sera were analyzed for OVA-specific IgG1 and IgG2a levels. Mice immunized with OVA +
M. pyrifera fucoidan produced remarkably higher amounts of anti-OVA IgG1 and IgG2a than control mice immunized with OVA alone (
Figure 6A,B). Fucoidan from
U. pinnatifida showed less effect on anti-OVA IgG1 and IgG2a production. On day 33, splenocytes were also harvested, re-stimulated with OVA
in vitro for four days, and analyzed for OVA-induced T cell proliferation, IFN-γ production and memory T cell generation. Splenocytes from mice immunized with OVA +
A. nodosum,
M. pyrifera and
F. vesiculosus fucoidan showed significantly greater cell proliferation and IFN-γ production than those from control mice immunized with OVA alone (
Figure 4C,D). These results indicate that fucoidans could function as an adjuvant by promoting Th1 type immune responses. We next examined whether fucoidans promote the generation of effector/memory T cells in OVA immunized mice based on the surface expression of CD44. Fucoidans from
A. nodosum,
M. pyrifera or
F. vesiculosus led to marked increases in the proportions of CD44
+ CD4 and CD8 T cells, whereas fucoidan from
U. pinnatifida did not show such effect (
Figure 6E). We also assayed for CTL activity in an
in vivo cytotoxicity assay. On day 33 after the initial immunization, the immunized mice received SIINFEKL-pulsed and CFSE-labeled splenocytes from C57BL/6 donor mice. Higher than 70% of specific target cell lysis was observed in mice immunized with OVA + fucoidan from
M. pyrifera or
F. vesiculosus, indicative of T cell memory induction (
Figure 6F). No significant target cell killing was observed in mice immunized with OVA + fucoidan from
A. nodosum or
U. pinnatifida. Collectively, these data suggest that fucoidans function as an adjuvant to enhance antigen specific T and B cell immune responses, and among the fucoidans tested in this study, fucoidan from
M. pyrifera is the most effective adjuvant for induction antigen specific immune responses.
2.8. Discussion
Fucoidan, a sulfated polysaccharide purified from brown algae, has been reported to modulate immunity based on both
in vitro and
in vivo studies [
3,
4,
17,
18]. Although a variety of biological activities of fucoidan have been reported, immune modulation effect of different species of fucoidans has been not compared. In this study, we demonstrated that fucoidan from
M. pyrifera has the most potent immune activating effect on human neutrophils, and on mouse NK cells, DCs and T cells compared to three other fucoidans.
Our previous study has shown that fucoidan from
U. pinnatifida induces delay of apoptosis and production of pro-inflammatory cytokines in human neutrophils and may be potential therapeutic compounds for bacterial infectious diseases and neutropenia by controlling neutrophil homeostasis and function with fucoidan [
13]. In this regard, we confirmed here that all fucoidans were able to delay spontaneous neutrophil apoptosis at high concentration. However, we demonstrated that only fucoidans derived from
U. pinnatifida and
M. pyrifera could inhibit neutrophil apoptosis at low concentration. The most plausible hypothesis to explain the specific inhibitory effect of fucoidans on neutrophil apoptosis could be their ability to react with L- and P-selectins on neutrophils [
19]. It is well-established that L-selectin is efficiently shed from the surface of neutrophils and down-regulation of L-selectin is induced during programmed neutrophil apoptosis [
20]. Fucoidan may block L-selectin shedding, which can contribute to the delay of neutrophil apoptosis. Whether fucoidans from
U. pinnatifida and
M. pyrifera can inhibit L-selectin shedding more efficiently than other fucoidans will be addressed in our future studies.
Activation of NK cells have been historically proposed as a complementary treatment for infectious diseases and cancer [
21]. Interestingly, we found that different fucoidan shows different effect on NK cell proliferation and activation. Fucoidan from
U. pinnatifida showed the strongest effect of NK cell expansion. However, fucoidan from
M. pyrifera showed the strongest effect on the activation and cytotoxic activity of NK cells than that from
U. pinnatifida. These differences may be caused by the ability of different fucoidans to induce different panels of cytokines
in vivo. Since NK cell activation and proliferation are differentially regulated by different cytokines [
22], fucoidan from
M. pyrifera and
U. pinnatifida may preferentially induce cytokines that promote NK cell activation or proliferation, respectively.
Figure 6.
Immunization with OVA and fucoidans enhances OVA specific immunity. C57BL/6 mice were immunized i.p. with PBS, OVA or OVA + fucoidan on days 0, 14, 28. On Day 33, serum (A) OVA-specific IgG1 and (B) IgG2a concentrations were measured by ELISA. * p < 0.05; ** p < 0.01 versus OVA group; (C) Splenocytes were harvested from immunized mice on day 33, and re-stimulated with or without OVA (50 μg/mL) for 4 days. Cell proliferation from re-stimulated splenocytes was measured; (D) IFN-γ concentrations in the above splenocytes culture supernatants were shown; (E) CD44 expression on CD4 or CD8 T cells was analyzed by flow cytometry. The number in each plot indicates the percentage of CD44+ cells (upper panel). Mean percentage of CD44+ cells in CD4 or CD8 T cells was shown (lower panel). Data are representative of or the average of analyses of six independent samples (two mice per experiment, total of three independent experiments). * p < 0.05; ** p < 0.01 versus OVA group; (F) On Day 33, in vivo killing of adoptively transferred SIINFEK-coated and CFSE-labeled target cells by CTLs in the immunized mice was measured. Data are from analyses of six individual mice in each group (two mice per experiment, total of six independent experiments). * p < 0.05; ** p < 0.01.
Figure 6.
Immunization with OVA and fucoidans enhances OVA specific immunity. C57BL/6 mice were immunized i.p. with PBS, OVA or OVA + fucoidan on days 0, 14, 28. On Day 33, serum (A) OVA-specific IgG1 and (B) IgG2a concentrations were measured by ELISA. * p < 0.05; ** p < 0.01 versus OVA group; (C) Splenocytes were harvested from immunized mice on day 33, and re-stimulated with or without OVA (50 μg/mL) for 4 days. Cell proliferation from re-stimulated splenocytes was measured; (D) IFN-γ concentrations in the above splenocytes culture supernatants were shown; (E) CD44 expression on CD4 or CD8 T cells was analyzed by flow cytometry. The number in each plot indicates the percentage of CD44+ cells (upper panel). Mean percentage of CD44+ cells in CD4 or CD8 T cells was shown (lower panel). Data are representative of or the average of analyses of six independent samples (two mice per experiment, total of three independent experiments). * p < 0.05; ** p < 0.01 versus OVA group; (F) On Day 33, in vivo killing of adoptively transferred SIINFEK-coated and CFSE-labeled target cells by CTLs in the immunized mice was measured. Data are from analyses of six individual mice in each group (two mice per experiment, total of six independent experiments). * p < 0.05; ** p < 0.01.
![Marinedrugs 13 01084 g006]()
The present study also investigated the adjuvant effect of different fucoidans on immune responses in mice. An ideal vaccine adjuvant should boost cell-mediated immune responses, such as specific antigen presentation by DCs and antigen-specific T cell activation and proliferation, in order to effectively eliminate pathogens [
23,
24]. We showed that treatment with fucoidans from
M. pyrifera and
F. vesiculosus cause decreases in the number of spleen DCs. It has been shown that fully matured DCs can undergo apoptosis or anergy when they induced T cell activation [
25,
26]. Therefore, treatment of fucoidans from
M. pyrifera and
F. vesiculosus may induce full maturation of spleen DCs that in turn stimulate T cells, which can happen rapidly within 24 h of fucoidan treatment. Consistent with this, fucoidan from
M. pyrifera and
F. vesiculosus showed effective T cell stimulation effect compared with other fucoidans. Moreover, fucoidans from
M. pyrifera and
F. vesiculosus exhibit an effective adjuvant activity to facilitate soluble OVA antigen-induced Th1 and CTL responses, OVA-specific IgG1 and IgG2a production and memory T cell differentiation, consistent with previous report that crude extract of fucoidan from
F. vesiculosus can functions as an adjuvant [
4].
According to the previous reports, the bioactivities of fucoidan are depending on the sulfate content, and fucoidan with higher sulfate shows higher activity [
16]. However, we found that fucoidan from
M. pyrifera has the most effective function in activating NK cells, DCs and T cells and has the strongest adjuvant effect, while its sulfate content is not substantially difference from the other fucoidans. On the other hand, based on composition study, the uronic acid content in
M. pyrifera fucoidan is higher than the other fucoidans. Interestingly, we have previously shown that ascophyllan purified from
A. nodosum has higher uronic acid levels than fucoidan and can promotes stronger DC maturation and T cell activation than fucoidan [
26,
27]. Hence, the uronic acid levels in fucoidan may be a factor that determines the immune-activating capacity, at least the capacity of activating DCs and T cells. Further investigation of this possibility and elucidation of the molecular and structural basis determining the immune-modulatory activity of fucoidan will be crucial to the development and optimization of fucoidan-based therapeutics and adjuvants in future.
In this study, we investigated the immune modulatory effect of four fucoidans including their effect on the apoptosis of human neutrophil in vitro and activation of mouse NK, DC and T cells in vivo, as well as their adjuvant effect on antigen-specific immune responses. Our results provide evidence that the fucoidan derived from M. pyrifera is a powerful immune modulator, which can delay human neutrophil apoptosis, and enhance mouse NK cell activation, DC maturation, T cell immune responses, antigen specific antibody production and memory T cell generation. Fucoidan from M. pyrifera may be a potentially useful novel drug for infectious diseases and cancer, and an effective adjuvant for vaccine.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}