
Influence of Temperature on Growth and Production of Pectenotoxin-2 by a Monoclonal Culture of Dinophysis caudata

Abstract
:1. Introduction
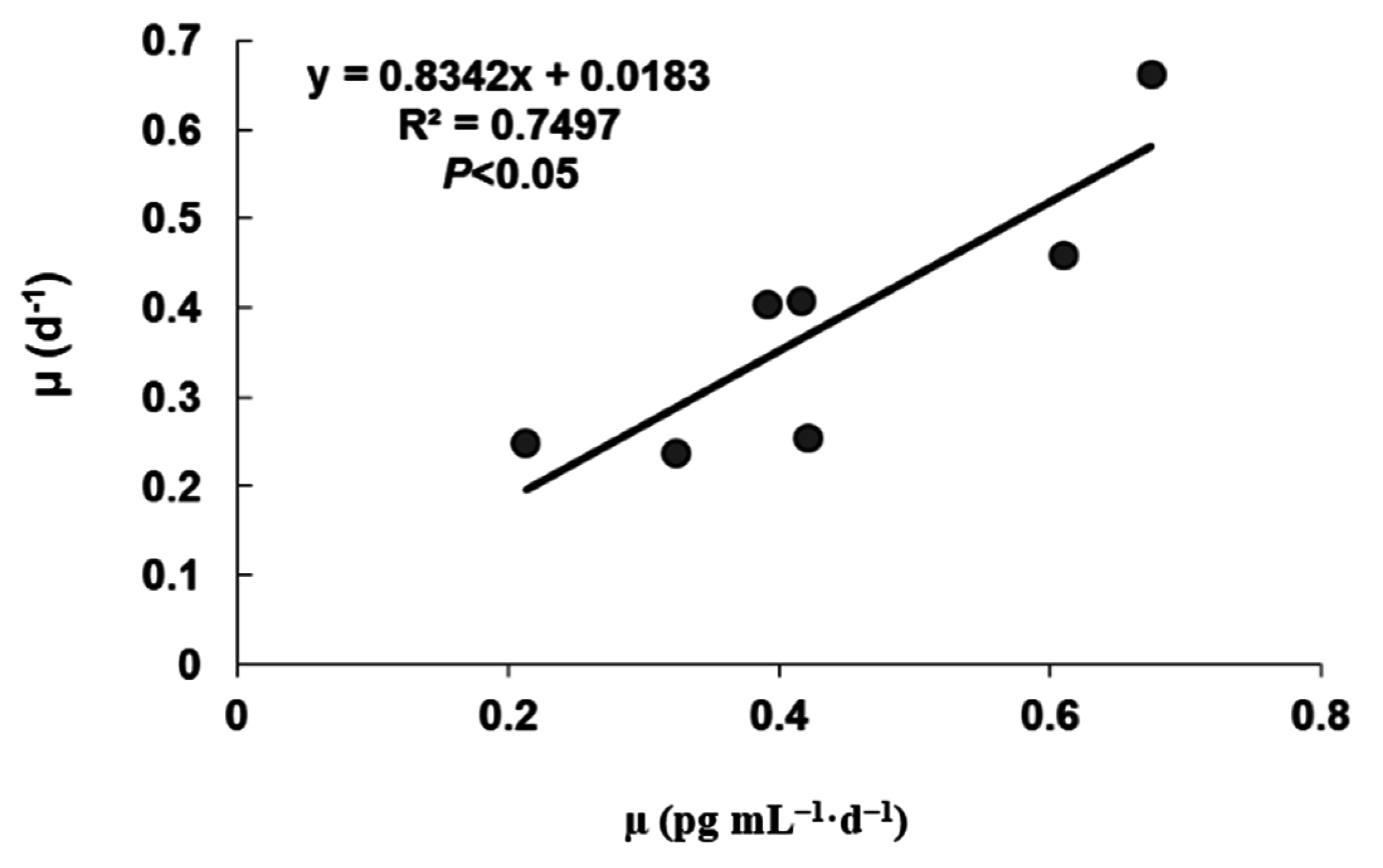
2. Results and Discussion
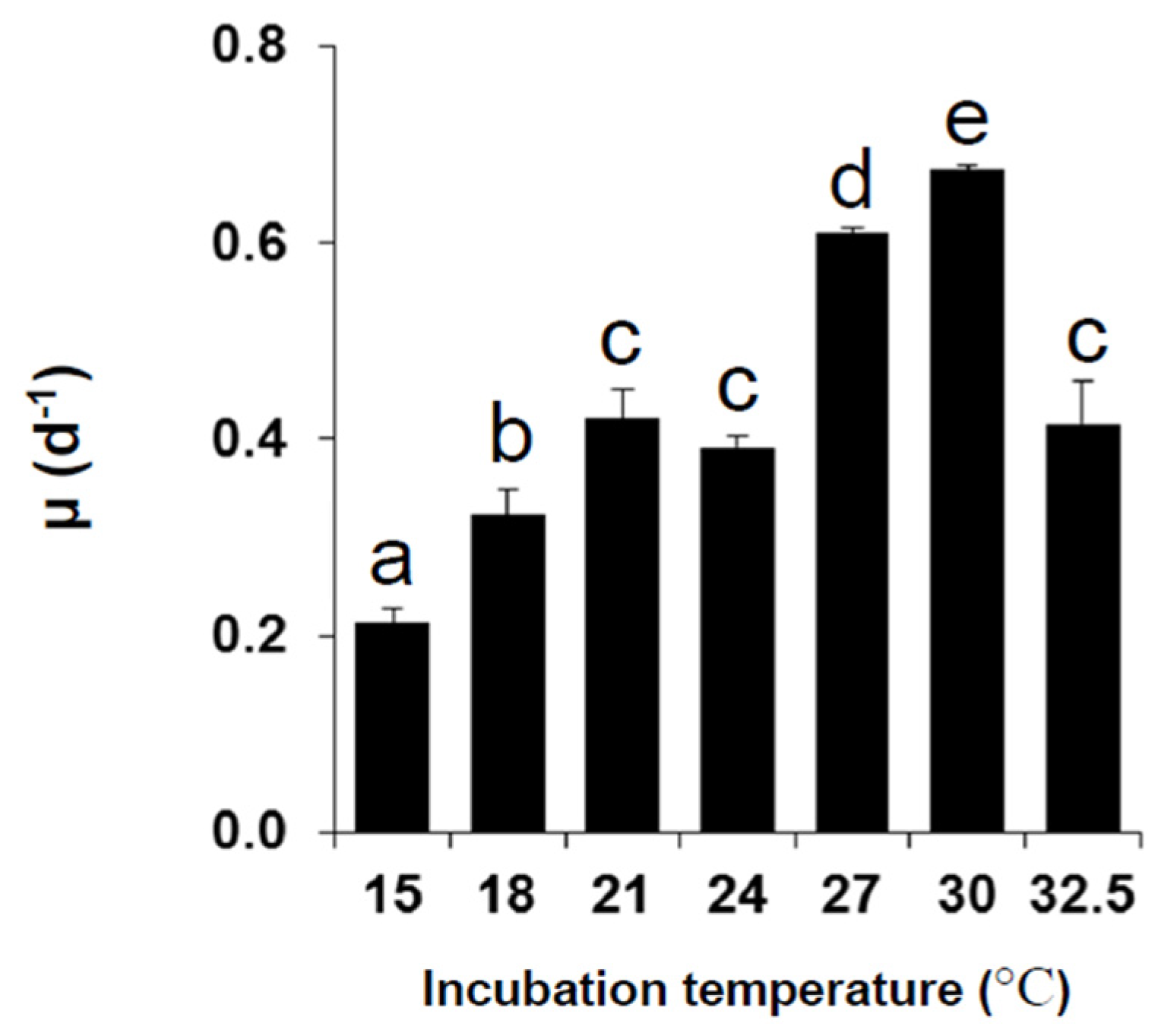
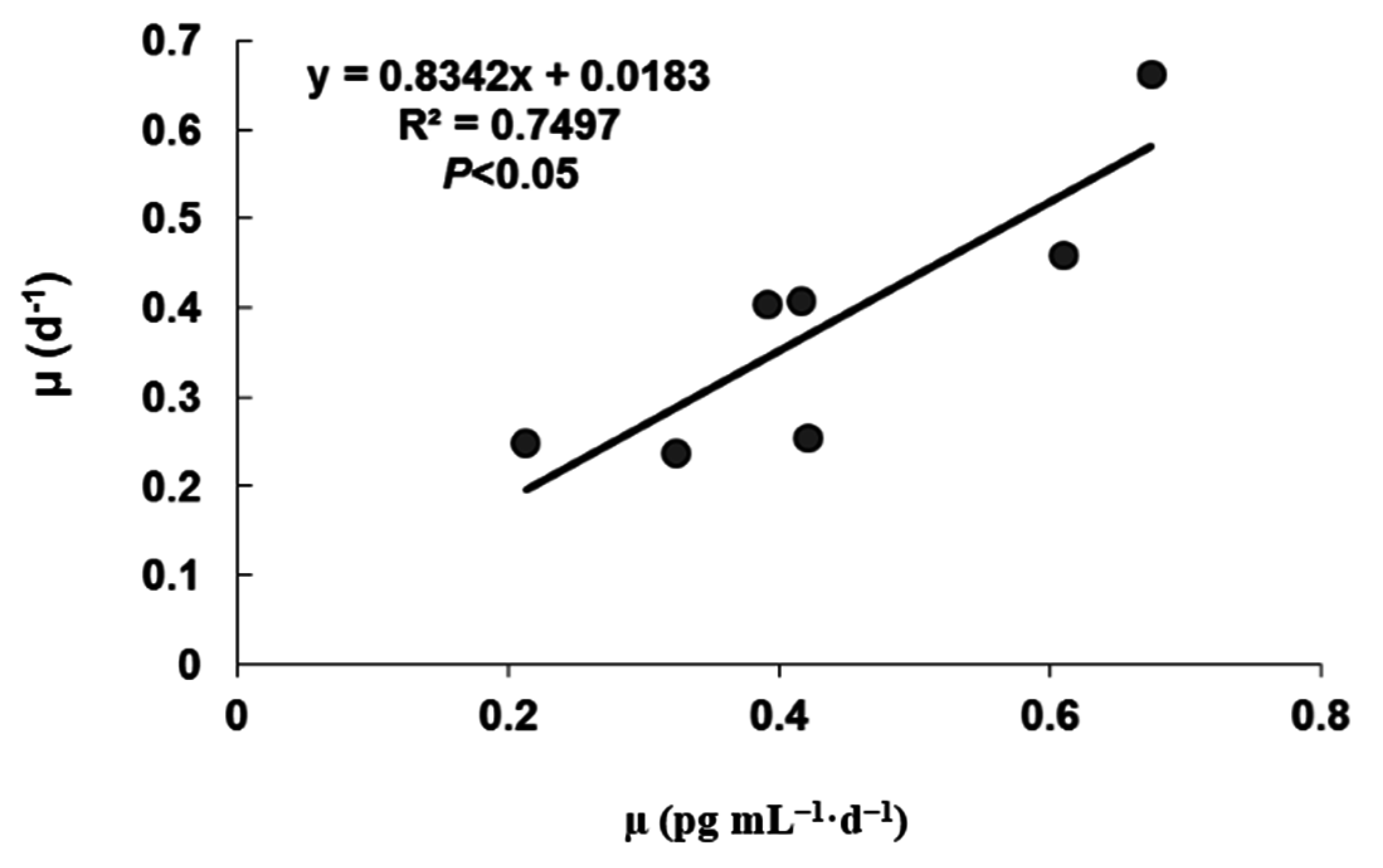
2.1. Growth under Different Temperatures
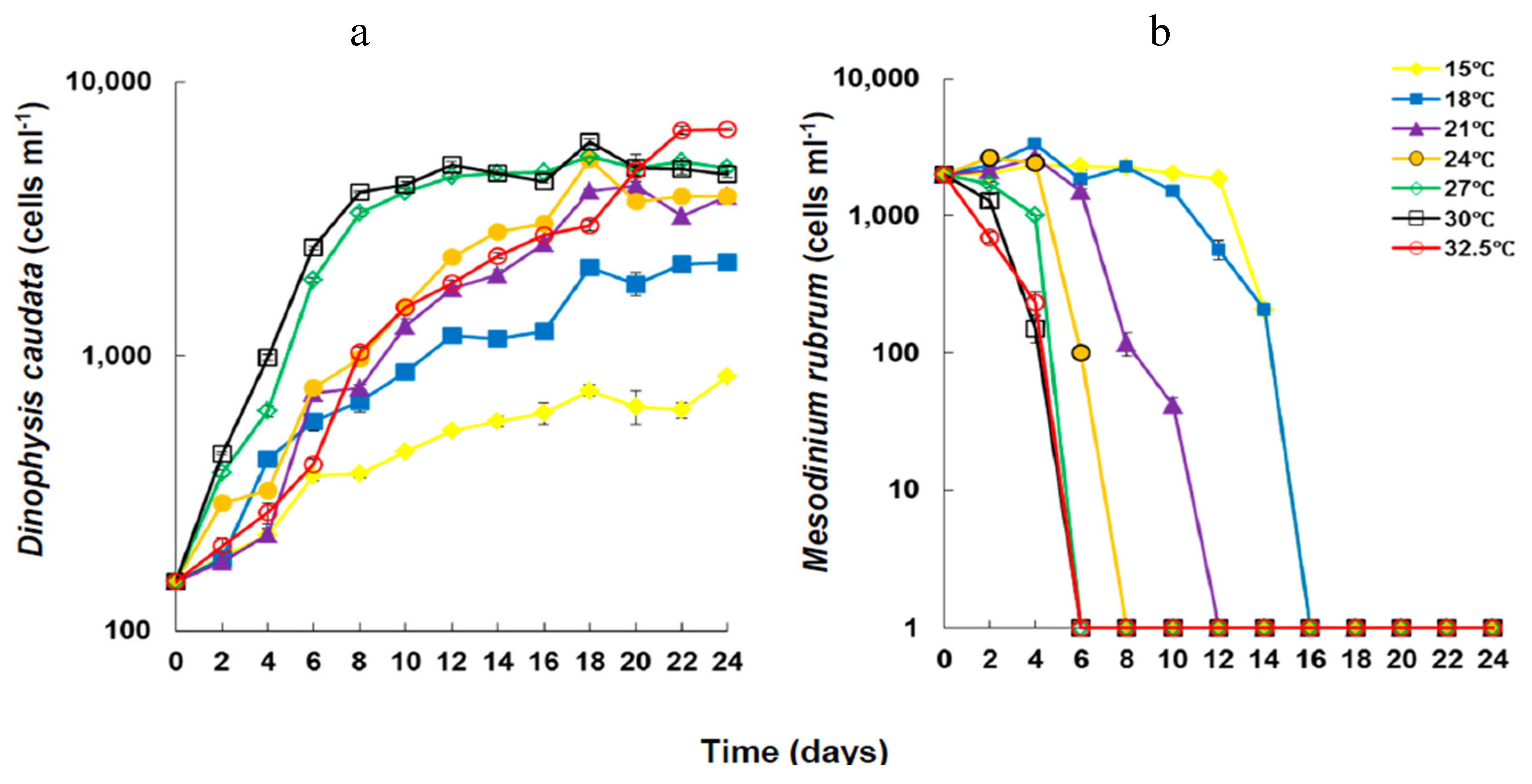

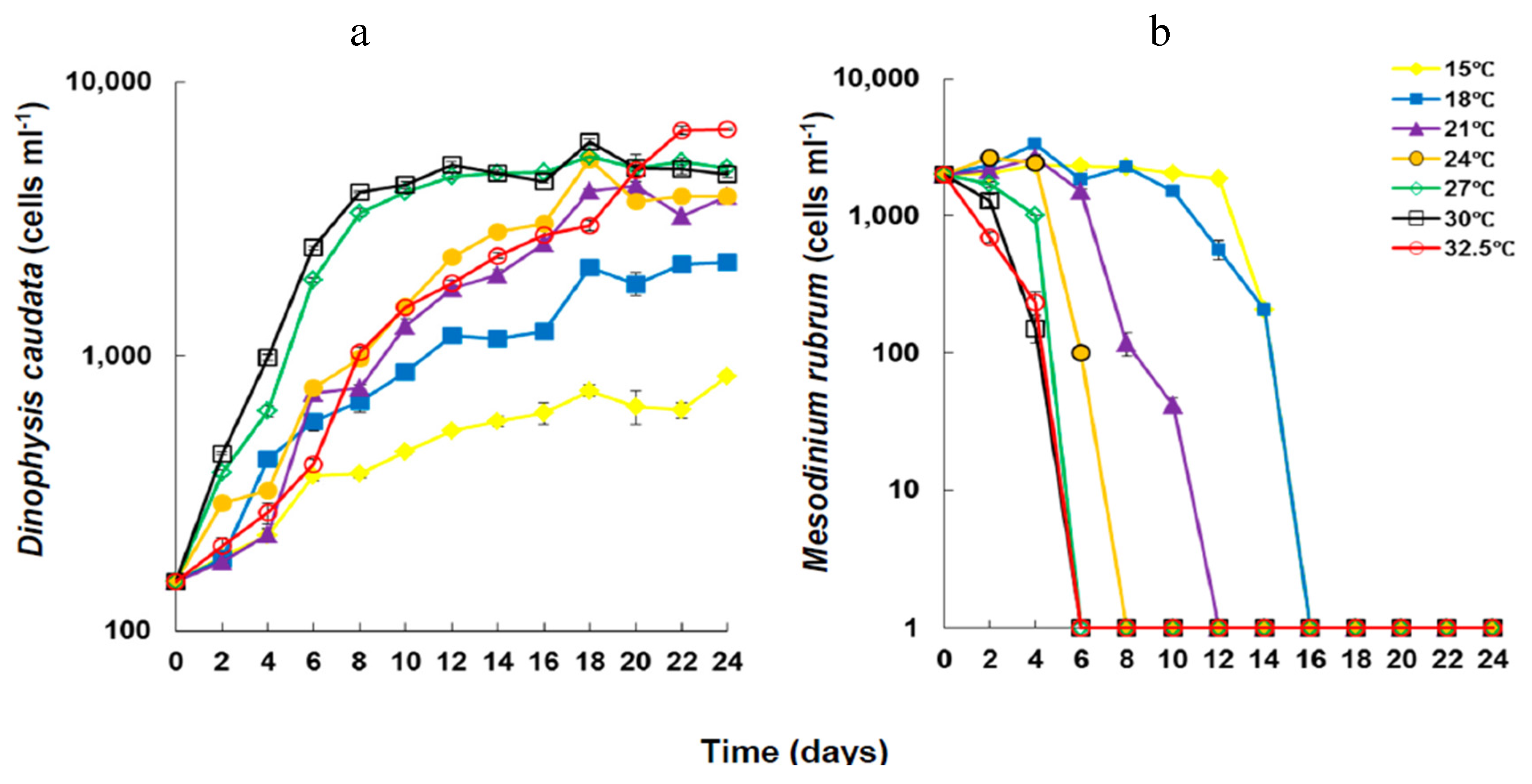{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | |||||||
|---|---|---|---|---|---|---|---|
| 15 | 18 | 21 | 24 | 27 | 30 | 32.5 | |
| 15 | |||||||
| 18 | NS | ||||||
| 21 | * | NS | |||||
| 24 | *** | NS | NS | ||||
| 27 | *** | *** | * | NS | |||
| 30 | *** | *** | * | NS | NS | ||
| 32.5 | *** | NS | NS | NS | NS | NS | |
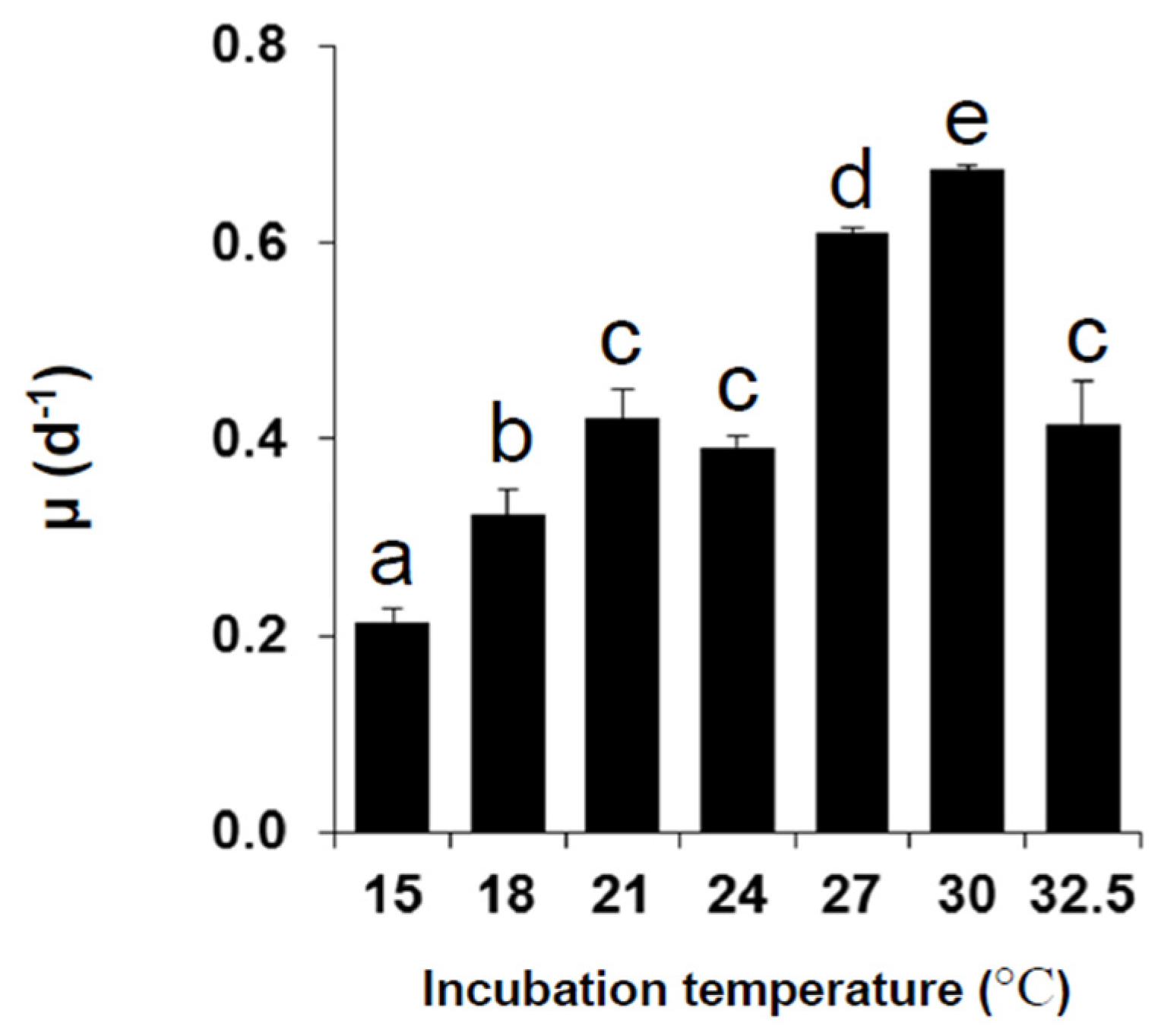
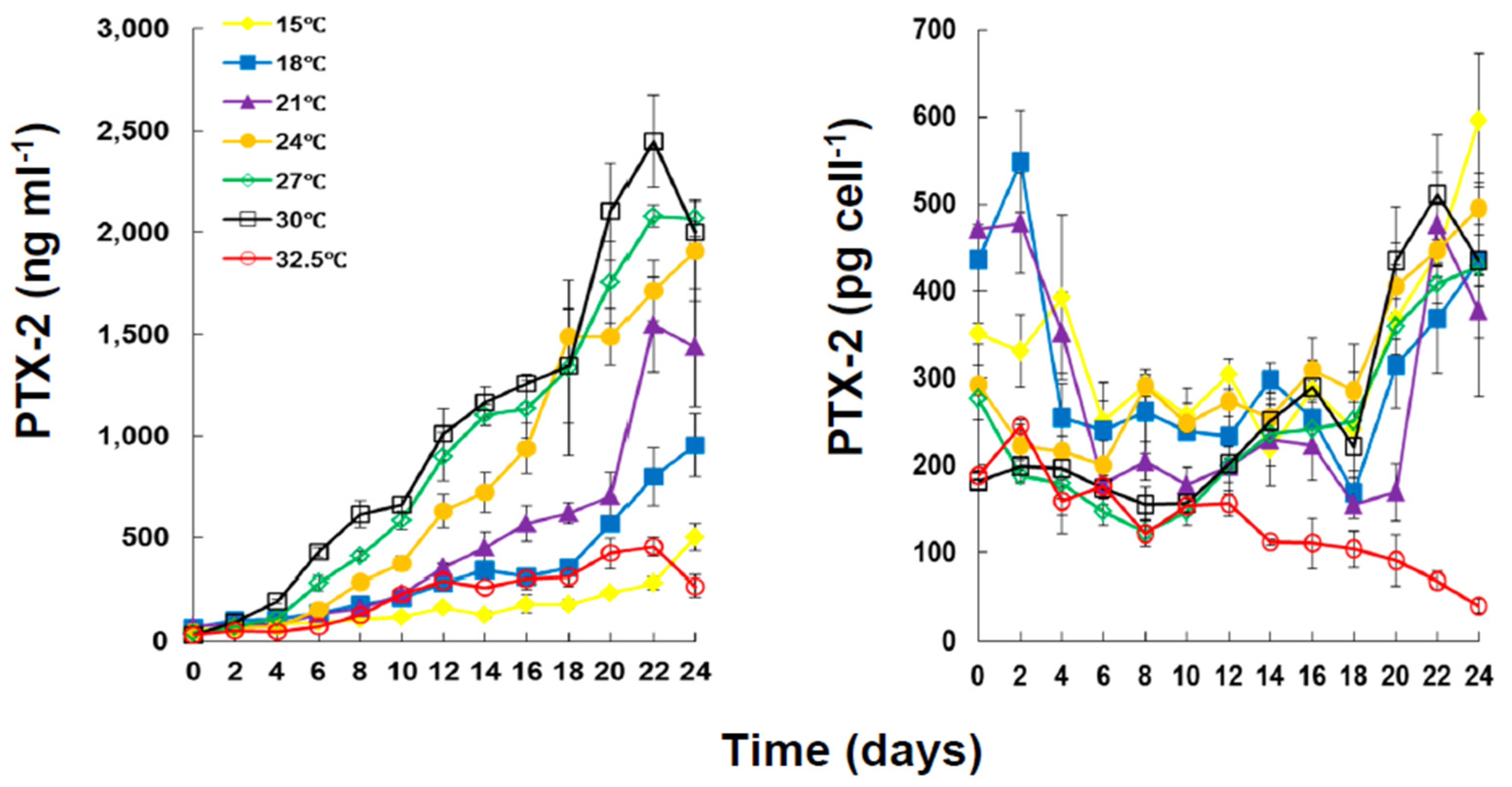
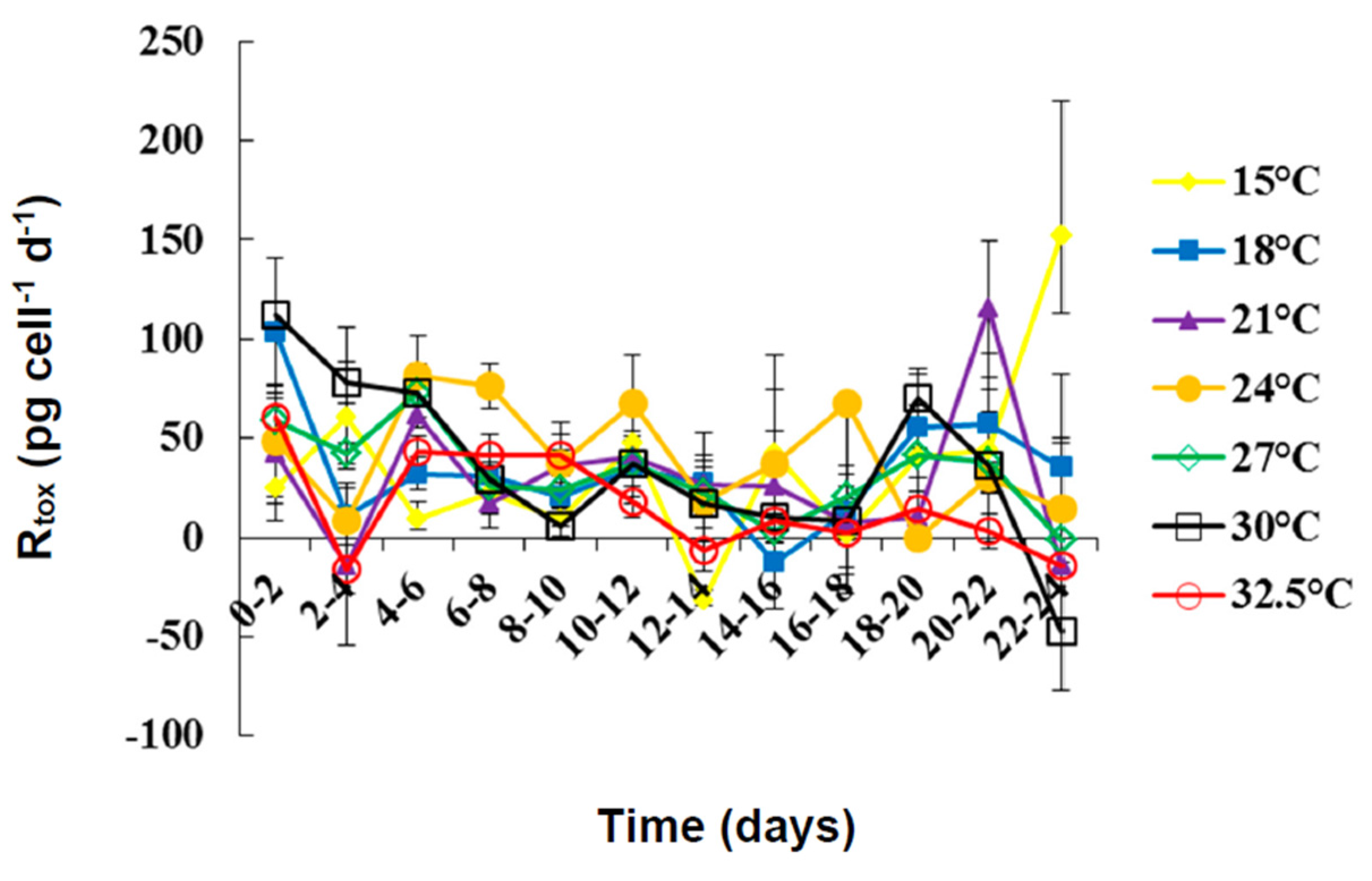
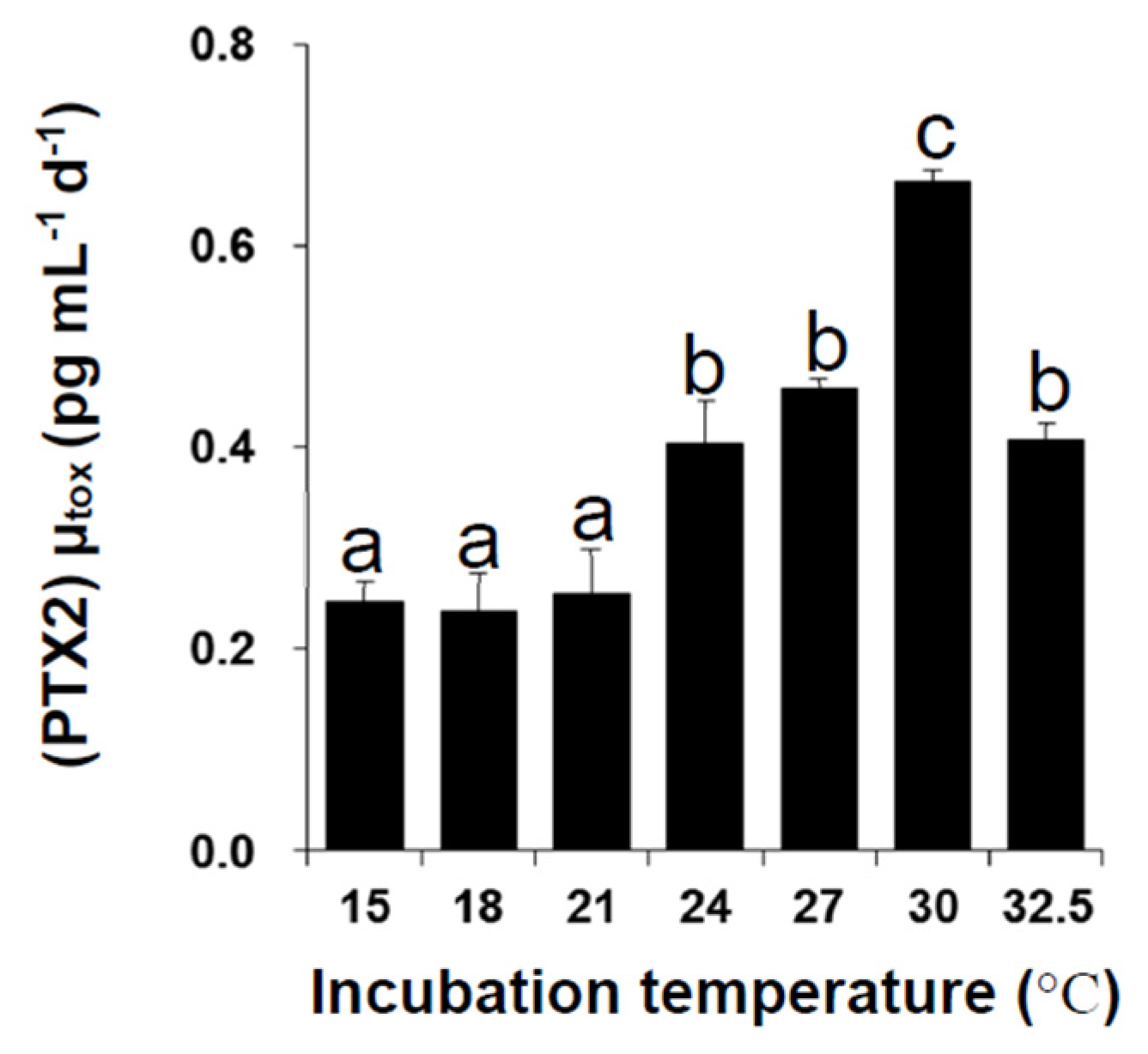
2.2. Toxin Content and Production under Different Temperatures
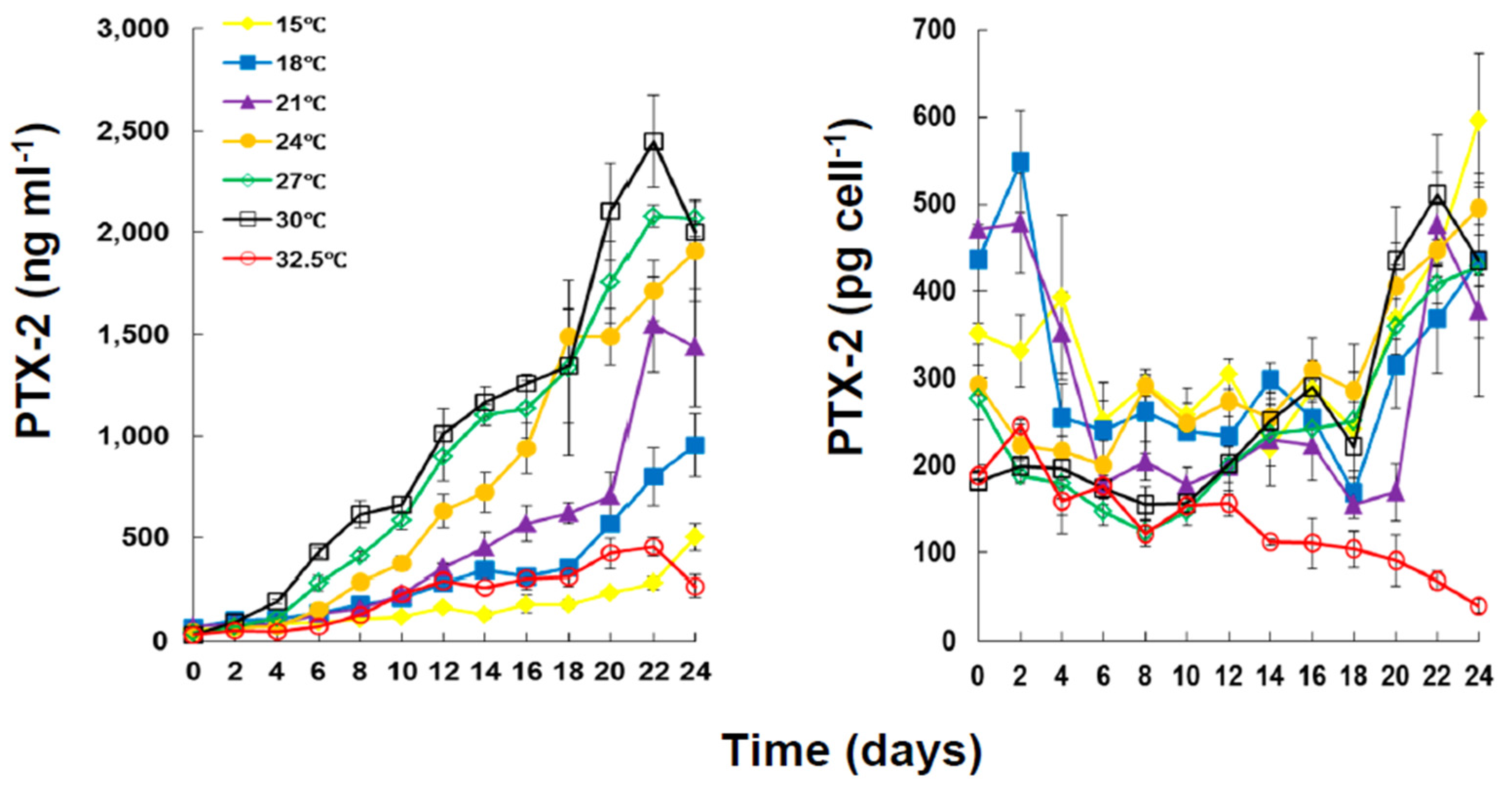
| Temperature (°C) | |||||||
|---|---|---|---|---|---|---|---|
| 15 | 18 | 21 | 24 | 27 | 30 | 32.5 | |
| 15 | |||||||
| 18 | NS | ||||||
| 21 | * | NS | |||||
| 24 | *** | NS | NS | ||||
| 27 | *** | NS | NS | NS | |||
| 30 | *** | * | NS | NS | NS | ||
| 32.5 | NS | NS | NS | * | * | * | |
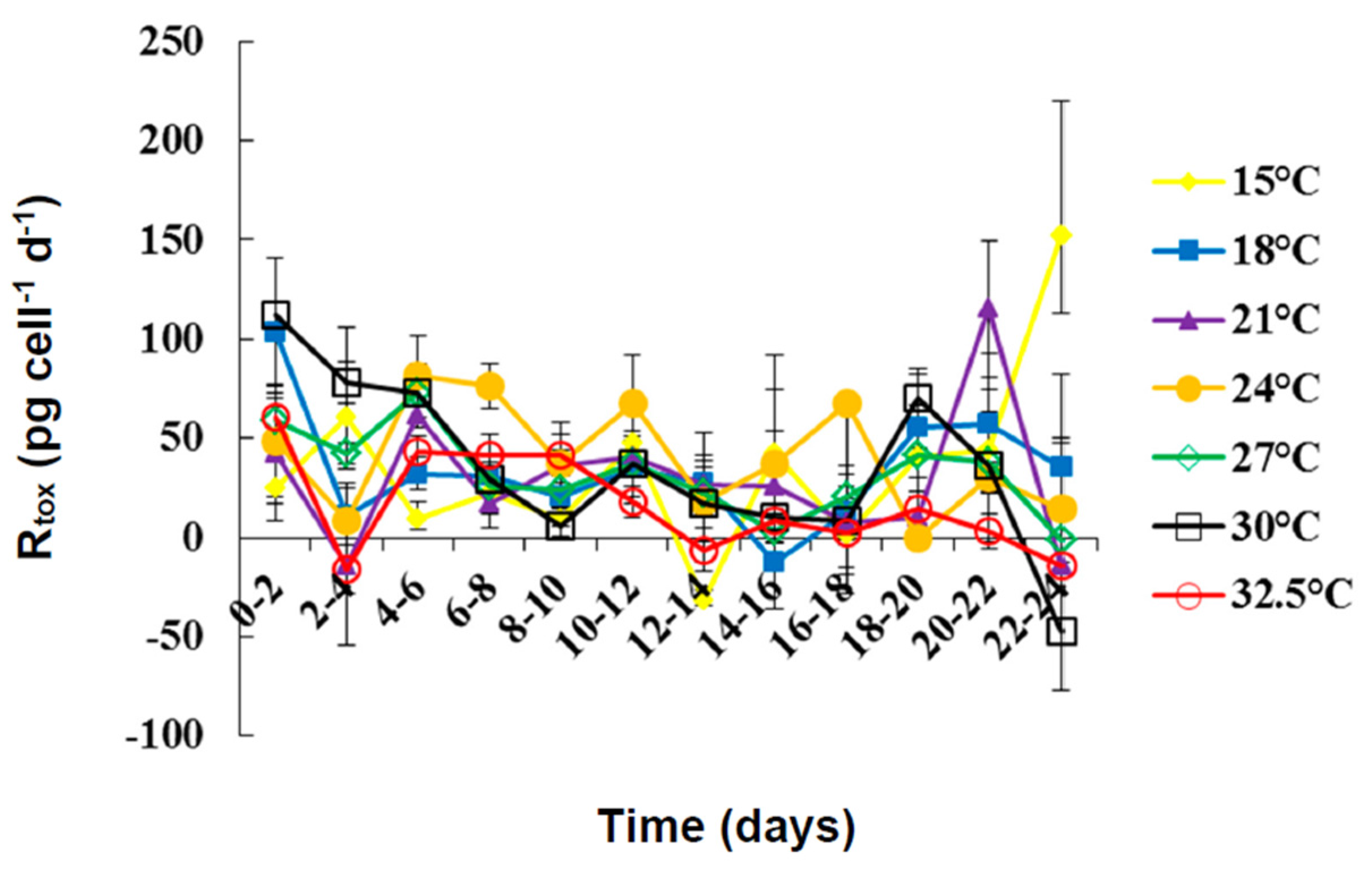
| Temperature (°C) | ||||||||
|---|---|---|---|---|---|---|---|---|
| 15 | 18 | 21 | 24 | 27 | 30 | 32.5 | ||
| Rtox | Min | −31.67 | −12.31 | −14.77 | 0 | −1.35 | −47.60 | −16.04 |
| Max | 152.04 | 103.32 | 115.33 | 81.39 | 72.86 | 112.30 | 60.65 | |
| Mean | 35.16 | 33.98 | 29.46 | 22.93 | 32.06 | 35.79 | 16.29 | |
| SE | 12.87 | 8.35 | 101.63 | 8.01 | 6.10 | 12.27 | 7.24 | |
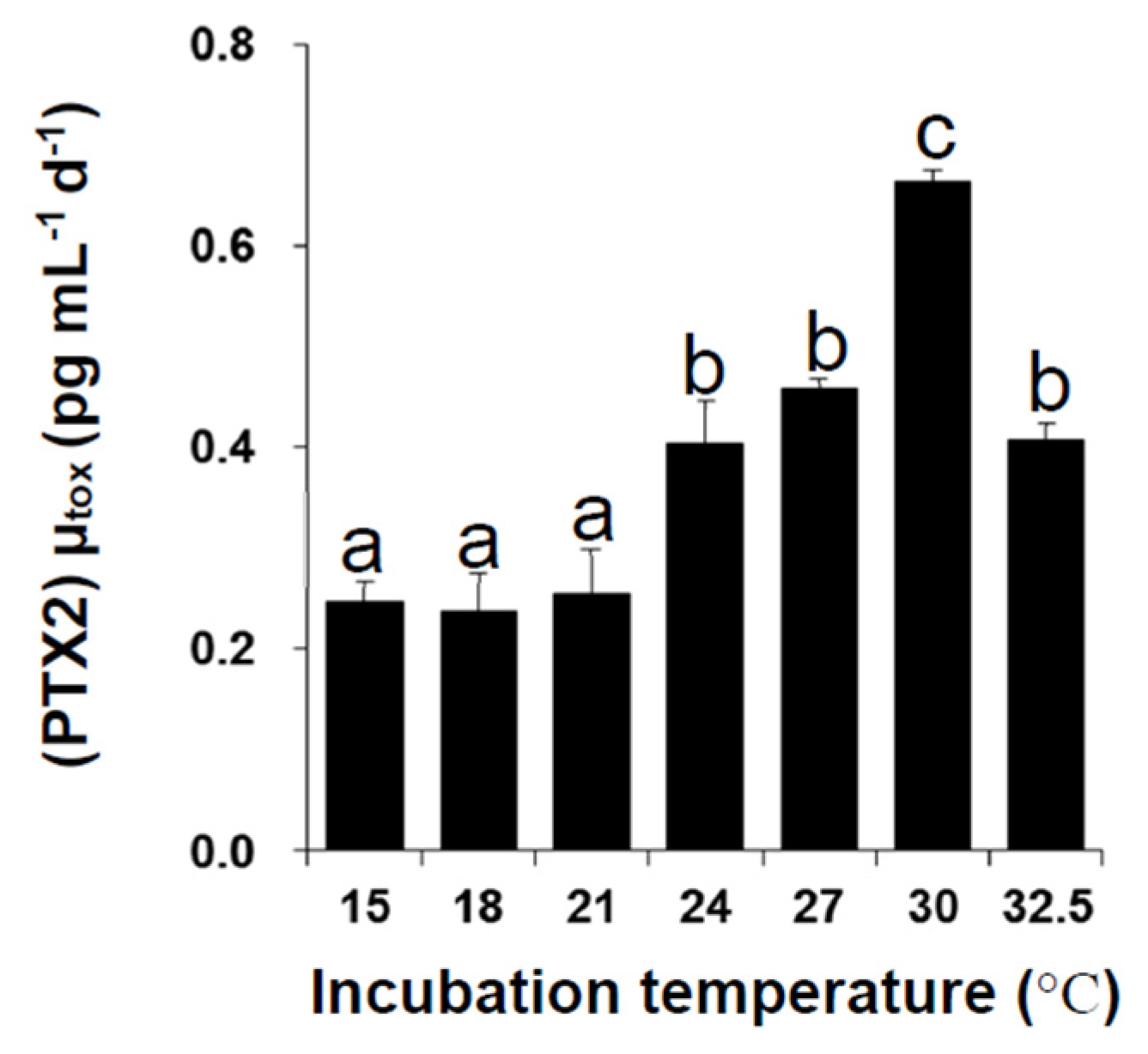
3. Experimental Section
3.1. Isolation and Maintenance of Clonal Strains
3.2. Growth Experiments
3.3. Toxin Analyses
3.4. Data Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Yasumoto, T.; Oshima, Y.; Yamaguchi, M. Occurrence of a new type of shellfish poisoning in the Tohoku district. Bull. Jpn. Soc. Sci. Fish. 1978, 44, 1249–1255. [Google Scholar]
- Yasumoto, T.; Murata, M. Marine toxins. Chem. Rev. 1993, 93, 1897–1993. [Google Scholar]
- Yasumoto, T.; Murata, M.; Oshima, Y.; Sano, M.; Matsumoto, G.; Clardy, J. Diarrhetic shellfish toxins. Tetrahedron 1985, 41, 1019–1025. [Google Scholar] [CrossRef]
- Terao, K.; Ito, E.; Yanagi, T.; Yasumoto, T. Histopatholgical studies on marine toxin poisoning. 1. Ultrastructural changes in the small intestine and liver of suckling mice induced by dinophysistoxin-1 and pectenotoxin-1. Toxicon 1986, 24, 1141–1151. [Google Scholar] [CrossRef]
- Bialojan, C.; Takai, A. Inhibitory effect of a marine-sponge toxin, okadaic acid, on protein phosphatases. Biochem. J. 1998, 256, 283–290. [Google Scholar]
- Fernández, J.J.; Candenas, M.L.; Souto, M.L.; Trujillo, M.M.; Norte, M. Okadaic acid, useful tool for studying cellular processes. Curr. Med. Chem. 2002, 9, 229–262. [Google Scholar] [CrossRef] [PubMed]
- Fujiki, H.; Suganuma, M.; Suguri, H.; Yoshizawa, S.; Takagi, K.; Uda, N.; Wakamatsu, K.; Yamada, K.; Murata, M.; Yasumoto, T.; et al. Diarrhetic shellfish toxin, dinophysistoxin-1, is a potent tumor promoter on mouse skin. Jpn. J. Cancer Res. 1988, 79, 1089–1093. [Google Scholar] [CrossRef] [PubMed]
- Tubaro, A.; Sosa, S.; Bornancin, A.; Hungerford, J. Pharmacology and toxicology of diarrheic shellfish toxins. In Seafood and Freshwater Toxins. Pharmacology, Physiology and Detection, 2nd ed.; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 229–253. [Google Scholar]
- Ogino, H.; Kumagai, M.; Yasumoto, T. Toxicologic evaluation of yessotoxin. Nat. Toxins 1997, 5, 255–259. [Google Scholar] [CrossRef]
- Terao, K.; Ito, E.; Orada, M.; Murata, M.; Yasumoto, T. Histopathological studies on experimental marine toxin poisoning—5. The effects in mice of yessotoxin isolated from Patinopecten yessoensis and of a desulfated derivative. Toxicon 1990, 28, 1095–1104. [Google Scholar] [CrossRef]
- Satake, M.; Ichimura, T.; Sekiguchi, K.; Yoshimatsu, S.; Oshima, Y. Confirmation of yessotoxin and 45,46,47-trinoryessotoxin production by Protoceratium reticulatum collected in Japan. Nat. Toxins 1999, 7, 147–150. [Google Scholar] [CrossRef]
- Lawrence, J.; Loreal, H.; Toyofuku, H.; Hess, P.; Iddya, K.; Ababouch, L. Assessment and Management of Biotoxin Risks in Bivalve Molluscs; FAO Fisheries and Aquaculture Technical Paper 551; FAO: Rome, Italy, 2011; p. 337. [Google Scholar]
- Yoon, M.Y.; Kim, Y.Z. Acute toxicity of pectenotoxin 2 and its effects on hepatic metabolizing enzyme in mice. Korean J. Toxicol. 1997, 13, 183–186. [Google Scholar]
- Ito, E.; Suzuki, T.; Oshima, Y.; Yasumoto, T. Studies on diarrheic activity on pectenotoxin-6 in the mouse and rat. Toxicon 2008, 15, 707–716. [Google Scholar] [CrossRef] [PubMed]
- Jun, J.H.; Sim, C.S.; Lee, C.O. Cytotoxic compounds from a two-sponge association. J. Nat. Prod. 1995, 58, 1722–1726. [Google Scholar]
- Vilariño, N.; Espiña, B. Pharmacology of pectenotoxins. In Seafood and Freshwater Toxins. Pharmacology, Physiology and Detection, 2nd ed.; Botana, L.M., Ed.; CRC Press: Boca Raton, FL, USA, 2008; pp. 361–369. [Google Scholar]
- Miles, C.O.; Wilkins, A.L.; Munday, R.; Dines, M.H.; Hawkes, A.D.; Briggs, L.R.; Quilliam, M.A.; MacKenzie, A.L.; Beuzenberg, V.; Towers, N.R. Isolation of pectenotpxin-2 from Dinophysis acuta and its conversion to pectenotoxin-2 seco acid, and preliminary assessment of their acute toxicities. Toxicon 2004, 43, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Miles, C.O.; Wilkins, A.L.; Munday, J.S.; Munday, R.; Hawkes, A.D.; Jensen, D.J.; Cooney, J.M.; Beuzenberg, V. Production of 7-epi-pectenotoxin-2 seco acid and assessment of its acute toxicity to mice. J. Agric. Food Chem. 2006, 54, 1530–1534. [Google Scholar] [CrossRef] [PubMed]
- Draisci, R.; Ferretti, E.; Palleschi, L.; Marchiafava, C.; Poletti, R.; Milandri, A.; Ceredi, A.; Popei, M. High levels of yessotoxin in mussels and presence of yessotoxin and homoyessotoxin in dinoflagellates of the Adriatic Sea. Toxicon 1999, 37, 1187–1193. [Google Scholar] [CrossRef]
- Paz, B.; Riobó, P.; Fernández, M.L.; Fraga, S.; Franco, J.M. Production and release of yessotoxins by the dinoflagellates Protoceratium reticulatum and Lingulodinium polyedrum in culture. Toxicon 2004, 44, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Rhodes, L.; McNabb, P.; de Salas, M.; Briggs, L.; Beuzenberg, V.; Gladstone, M. Yessotoxin production by Gonyaulax spinifera. Harmful Algae 2006, 5, 148–155. [Google Scholar] [CrossRef]
- Camacho, F.G.; Rodríguez, J.G.; Miron, S.S.; Ceron García, M.C.; Belarbi, E.H.; Christi, Y.; Molima Grima, E. Biotechnological significance of toxic marine dinoflagellates. Biotechnol. Adv. 2007, 25, 176–194. [Google Scholar] [CrossRef] [PubMed]
- Gómez, F. A list of free-living dinoflagellate species in the world’s oceans. Acta Bot. Croat. 2005, 64, 129–212. [Google Scholar]
- Jacobson, D.M.; Anderson, R.A. The discovery of mixotrophy in photosynthetic species of Dinophysis (Dinophyceae): Light and electron microscopical observations of food vacuoles in Dinophysis acuminata, D. norvegica and two heterotrophic dinophysoid dinoflagellates. Phycologia 1994, 33, 97–110. [Google Scholar] [CrossRef]
- Reguera, B.; Gil, S. Small cell and intermediate cell formation in species of Dinophysis (Dinophyceae, Dinophysiales). J. Phycol. 2001, 37, 318–333. [Google Scholar] [CrossRef]
- Reguera, B.; Velo-Suárez, L.; Raine, R.; Park, M. Harmful Dinophysis species: A review. Harmful Algae 2012, 14, 87–106. [Google Scholar] [CrossRef]
- Reguera, B.; Garcés, E.; Pazos, I.; Bravo, I.; Ramilo, I.; González-Gil, S. Cell cycle patterns and estimates of in situ division rates of dinoflagellates of the genus Dinophysis by a postmitotic index. Mar. Ecol. Prog. Ser. 2003, 249, 117–131. [Google Scholar] [CrossRef]
- Reguera, B.; Pizarro, G. Planktonic Dinoflagellates Which Produce Polyether Toxins of the Old―DSP Complex. In Seafood and Freshwater Toxins: Pharmacology, Physiology and Detection, 2nd ed.; Botana, L.M., Ed.; Taylor & Francis: London, UK, 2008; pp. 257–284. [Google Scholar]
- Guzmán, L.; Campodonico, I. Marea roja en la región de Magallanes. Publ. Inst. Pat. Ser. Mon. 1975, 9, 44. [Google Scholar]
- MacDonald, E. Dinophysis Bloom in West Scotland; Harmful Algae News Series No.9; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 1994. [Google Scholar]
- Subba Rao, D.V.; Pan, Y.; Zitko, V.; Bugden, G.; Mackeigan, K. Diarrhetic shellfish poisoning (DSP) associated with a subsurface bloom of Dinophysis norvegica in Bedford Basin, eastern Canada. Mar. Ecol. Prog. Ser. 1993, 97, 117–126. [Google Scholar] [CrossRef]
- Dahl, E.; Aune, T.; Aase, B. Reddish water due to mass occurrence of Dinophysis spp. In Harmful and Toxic Algal Blooms; Yasumoto, T., Oshima, Y., Fukuyo, Y., Eds.; Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 1996; pp. 265–267. [Google Scholar]
- Reguera, B.; Riobó, P.; Rodríguez, F.; Díaz, P.A.; Piazarro, G.; Paz, B.; Franco, J.M.; Blanco, J. Dinophysis toxins: Causative organisms, distribution and fate in shellfish. Mar. Drugs 2014, 12, 394–461. [Google Scholar] [CrossRef] [PubMed]
- Santhanam, R.; Srinivasan, A. Impact of dinoflagellate Dinophysis caudata bloom on the hydrography and fishery potentials of Tuticorin Bay, South India. In Harmful and Toxic Algal Blooms; Yasumoto, Y., Oshima, Y., Fukuyo, Y., Eds.; IOC of UNESCO: Sendai, Japan, 1996; pp. 41–44. [Google Scholar]
- Poletti, R.; Cettul, K.; Bovo, F.; Milandri, A.; Pompei, M.; Frate, R. Distribution of toxic dinoflagellates and their impact on shellfish along the northwest Adriatic coast. In Harmful Algae; Reguera, B., Blanco, J., Fernández, M.L., Wyatt, T., Eds.; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Santiago de Compostela, Spain, 1998; pp. 88–90. [Google Scholar]
- Morton, S.T.; Vershinin, A.; Smith, L.L.; Leighfield, T.A.; Pankov, S.; Quilliam, M.A. Seasonality of Dinophysis spp. and Prorocentrum lima in Black Sea phytoplankton and associated shellfish toxicity. Harmful Algae 2009, 8, 629–639. [Google Scholar] [CrossRef]
- Pizarro, G.; Moroño, A.; Paz, P.; Franco, J.M.; Pazos, Y.; Reguera, B. Evaluation of passive samplers as a monitoring tool for early warning of Dinophysis toxins in shellfish. Mar. Drugs 2013, 11, 3823–3845. [Google Scholar] [CrossRef] [PubMed]
- Tahri-Joutei, L. Gymnodinium catenatum Graham blooms on Moroccan waters. In Harmful Algae; Reguera, B., Blanco, J., Fernández, M.L., Wyatt, T., Eds.; Xunta de Galicia and Intergovernmental Oceanographic Commission of UNESCO: Santiago de Compostela, Spain, 1998; pp. 66–67. [Google Scholar]
- Flores, M.; Franco, J.M.; Lluch Cota, S.E.; Lluch Cota, D.B.; Cortés-Altamirano, R.; Sierra-Beltrán, A.P. Recent species shifts on the HAB occurrences in Acapulco Bay, Mexico. In Harmful Algae 2002, Proceedings of the 10th International Conference on Harmful Algae, St. Pete Beach, FL, USA, 21–25 October 2002; Steidinger, K.A., Landsberg, J.H., Tomas, C.R., Vargo, G.A., Eds.; Florida Fish and Wildlife Conservation Commission, Florida Institute of Oceanography and Intergovernmental Oceanographic Commission of UNESCO: Paris, France, 2002; p. 96. [Google Scholar]
- Dickey, R.; Fryxell, G.; Granade, R.; Roelke, D. Detection of the marine toxins okadaic acid and domoic acid in shellfish and phytoplankton in the Gulf of Mexico. Toxicon 1992, 30, 355–359. [Google Scholar] [CrossRef]
- Méndez, S.; Ferrari, G. Floraciones algales nocivas en Uruguay: Antecedentes, proyectos en curso y revisión de resultados. In Floraciones Algales Nocivas en el Cono Sur Americano; Sar, E.A., Ferrario, M.E., Reguera, B., Eds.; Instituto Español de Oceanografía: Madrid, Spain, 2002; pp. 271–288. (In Spanish) [Google Scholar]
- Sar, E.A.; Sunesen, I.; Lavigne, A.; Goya, A. Dinophysis spp. asociadas a detección de toxinas diarreicas (DSTs) en moluscos y a intoxicación diarreica en humanos (Provincia de Buenos Aires, Argentina). Rev. Biol. Mar. Oceanogr. 2010, 45, 451–460. (In Spanish) [Google Scholar] [CrossRef]
- Fukuyo, Y.; Toyoda, Y.; Miyazaki, S. Dinoflagellates found in Sanriku Coast—I. Genus Dinophysis. Otsuchi Mar. Res. Cent. Rep. 1981, 7, 13–23. [Google Scholar]
- Tseng, C.K.; Zhou, M.J.; Zou, J.Z. Toxic Phytoplankton Studies in China. In Toxic Phytoplankton Blooms in the Sea; Smayda, T.J., Shimizu, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 1993; pp. 347–352. [Google Scholar]
- Holmes, M.J.; Teo, S.L.M. Toxic marine dinoflagellates in Singapore waters that cause seafood poisonings. Clin. Exp. Pharmacol. Physiol. 2002, 29, 829–836. [Google Scholar] [CrossRef] [PubMed]
- Karunasagar, I.; Segar, K.; Karunasagar, I. Incidence of PSP and DSP in shellfish along the coast of Karnataka State (India). In Red Tides. Biology, Enviromental Science and Toxicology; Okaichi, T., Anderson, D.M., Nemoto, T., Eds.; Elsevier: New York, NY, USA, 1989; pp. 61–64. [Google Scholar]
- Burgess, V.; Shaw, G. Pectenotoxins—An issue for public health. A review of their comparative toxicology and metabolism. Environ. Int. 2001, 27, 275–283. [Google Scholar] [CrossRef]
- Lee, J.S.; Tangen, K.; Dahl, E.; Hovgaard, P.; Yasumoto, T. Diarrhetic shellfish toxins in Norwegian mussels. Nippon Suisan Gakk. 1989, 54, 1953–1957. [Google Scholar] [CrossRef]
- Marasigan, A.N.; Sato, S.; Fukuyo, Y.; Kodama, M. Accumulation of a high level of diarrhetic shellfish toxins in the green mussel Perna viridis during a bloom of Dinophysis caudata and Dinophysis miles in Saipan Bay, Panay Island, the Philippines. Fish. Sci. 2001, 67, 994–996. [Google Scholar] [CrossRef]
- Fernández, M.L.; Reguera, B.; González-Gil, S.; Míguez, A. Pectenotoxin-2 in single-cell isolates of Dinophysis caudata and Dinophysis acuta from the Galician Rías (NW Spain). Toxicon 2006, 48, 477–490. [Google Scholar] [CrossRef] [PubMed]
- Fernández, M.L.; Reguera, B.; Ramilo, I.; Martínez, A. Toxin content of Dinophysis acuminata, D. acuta and D. caudata from the Galician Rías Baixas. In Harmful Algal Blooms; Hallegraeff, G.M., Blackburn, S.I., Bolch, C.J., Lewis, R.J., Eds.; Intergovernmental Commission of UNESCO: Paris, France, 2001; pp. 360–363. [Google Scholar]
- Okaichi, T. Red tides found in and around the Seto Inland Sea in 1965. Tech. Bull. Fac. Agric. Kagawa Univ. 1967, 18, 181–185. [Google Scholar]
- Holmes, M.; Teo, S.; Lee, F.; Khoo, H. Persistent low concentrations of diarrhetic shellfish toxins in green mussels Perna viridis from the Johor Strait, Singapore: First record of diarrhetic shellfish toxins from South-East Asia. Mar. Ecol. Prog. Ser. 1999, 181, 257–268. [Google Scholar] [CrossRef]
- Basti, L.; Uchida, H.; Kanamori, M.; Matsushima, R.; Suzuki, T.; Nagai, S. Mortality and pathology of Japanese scallop, Patinopecten (Mizuhopecten) yessoensis, and noble scallop, Mimachlamys nobilis, fed monoclonal culture of PTX-producer, Dinophysis caudata. In Marine and Freshwater Harmful Algae, Proceedings of the 16th International Conference on Harmful Algae, Wellington, New Zealand, 27–30 October 2014; MacKenzie, A.L., Ed.; Cawthron Institute, Nelson, New Zealand and International Society for the Study of Harmful Algae: Wellington, New Zealand, 2014; pp. 105–108. [Google Scholar]
- Nishitani, G.; Nagai, S.; Sakiyama, S.; Kamiyama, T. Successful cultivation of the toxic dinoflagellate Dinophysis caudata (Dinophyceae). Plankton Benthos Res. 2008, 3, 78–85. [Google Scholar] [CrossRef]
- Park, M.G.; Kim, S.; Kim, H.S.; Myung, G.; Kang, Y.G.; Yih, W. First successful culture of the marine dinoflagellate Dinophysis acuminata. Aquat. Microb. Ecol. 2006, 45, 101–106. [Google Scholar] [CrossRef]
- Nagai, S.; Nitshitani, G.; Tomaru, Y.; Sakiyama, S.; Kamiyama, T. Predation by the toxic dinoflagellate Dinophysis fortii on the ciliate Myrionecta rubra and observation of sequestration of ciliate chloroplasts. J. Phycol. 2008, 44, 909–922. [Google Scholar] [CrossRef]
- Nishitani, G.; Nagai, S.; Takano, Y.; Sakiyama, S.; Baba, K.; Kamiyama, T. Growth characteristics and phylogenetic analysis of the marine dinoflagellate Dinophysis infundibulus (Dinophyceae). Aquat. Microb. Ecol. 2008, 52, 209–221. [Google Scholar] [CrossRef]
- Rodríguez, F.; Escalera, L.; Reguera, B.; Rial, P.; Riobó, P.; da Silva, T.J. Morphological variability, toxinology and genetics of the dinoflagellate Dinophysis tripos (Dinophysiaceae, Dinophysiales). Harmful Algae 2012, 13, 26–33. [Google Scholar] [CrossRef]
- Nagai, S.; Suzuki, T.; Kamiyama, T. Successful cultivation of the toxic dinoflagellate Dinophysis tripos (Dinophyceae). Plankton Benthos Res. 2013, 8, 171–177. [Google Scholar] [CrossRef]
- Jaén, D.; Mamán, L.; Domínguez, R.; Martín, E. First report of Dinophysis acuta in culture. Harmful Algal News 2009, 39, 1–2. [Google Scholar]
- Mafra, L.L., Jr. Detecção de Toxinas eEfeitos Tóxicos em Microalgas Cultivadas ou Coletadas na Costa sul do Brasil: Estado da Arte em 2013. In Livro de Resumos da Reunião Latino-Americana sobre Algas Nocivas, Santa Catarina, Brasil, 7–9 de Outubro, 2013; Proença, L.A.O., Renan de Souza, K., Eds.; Laboratory of Research and Monitoring of Harmful Algae and Phycotoxins: Santa Catarina, Brazil, 2013; p. 18. (In Portuguese) [Google Scholar]
- Riobó, P.; Reguera, B.; Franco, J.M.; Rodríguez, F. First report of the toxin profile of Dinophysis sacculus Stein from LC-MS analysis of laboratory cultures. Toxicon 2013, 76, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Cembella, A.D.; John, U. Molecular physiology of toxin production and growth regulation in harmful algae. In Ecology of Harmful Algae; Graneli, E., Turner, J.T., Eds.; Springer-Verlag: Heidelberg, Germany, 2006; pp. 215–227. [Google Scholar]
- Kamiyama, T.; Suzuki, T. Production of dinophysis-1 and pectenotoxin-2 by a culture of Dinophysis acuminata (Dinophyceae). Harmful Algae 2009, 8, 312–317. [Google Scholar] [CrossRef]
- Nagai, S.; Suzuki, T.; Nishikawa, T.; Kamiyama, T. Differences in the production and excretion kinetics of okadaic acid, dinophysistoxin-1, and pectenotoxin-2 between cultures of Dinophysis acuminata and Dinophysis fortii isolated from western Japan. J. Phycol. 2011, 47, 1326–13337. [Google Scholar] [CrossRef]
- Kamiyama, T.; Nagai, S.; Suzuki, T.; Miyamura, K. Effect of temperature on production of okadaic acid, dinophysistoxin-1, and pectenotoxin-2 by Dinophysis acuminata in culture experiments. Aquat. Microb. Ecol. 2010, 60, 193–202. [Google Scholar] [CrossRef]
- Tong, M.; Kulis, D.M.; Fux, E.; Smith, J.L.; Hess, P.; Zhou, Q.; Anderson, D.M. The effects of growth phase and light intensity on toxin production by Dinophysis acuminata from northeastern United States. Harmful Algae 2011, 10, 254–264. [Google Scholar] [CrossRef]
- Watton, P.S. Temperature and lake-outlet communities. J. Therm. Biol. 1995, 20, 121–125. [Google Scholar] [CrossRef]
- Hansen, P.J.; Bjørnsen, P.K.; Hansen, B.W. Zooplankton grazing and growth: Scaling within the 2–2000-µm body size range. Limnol. Oceanogr. 1997, 42, 687–704. [Google Scholar] [CrossRef]
- Crawford, D.W. Mesodinium rubrum: The phytoplankter that wasn’t. Mar. Ecol. Prog. Ser. 1989, 58, 161–174. [Google Scholar] [CrossRef]
- Fenchel, T.; Hansen, R.S. Motile behavior of the bloom-forming ciliate Mesodinium rubrum. Mar. Biol. Res. 2006, 2, 33–40. [Google Scholar] [CrossRef]
- Yih, W.; Kim, H.S.; Jeong, H.J.; Myung, G.; Kim, Y.G. Ingestion of cryptophyte cells by the marine photosynthetic ciliate Mesodiunium rubrum. Aquat. Microb. Ecol. 2004, 36, 165–170. [Google Scholar] [CrossRef]
- Jakobsen, H.H.; Everett, L.M.; Strom, S.L. Hydromechanical signaling between the ciliate Mesodinium pulex and motile prey. Mar. Ecol. Prog. Ser. 2006, 44, 197–206. [Google Scholar] [CrossRef]
- Riisgård, H.U.; Larsen, P.S. Ciliary-propelling mechanism, effects of temperature and viscosity on swimming speed, and adaptive significance of “jumping” in the ciliate Mesodinium rubrum. Mar. Biol. Res. 2009, 5, 585–595. [Google Scholar] [CrossRef]
- Reguera, B.; Bravo, I.; McCall, H.; Reyero, M.I. Phased cell division and other biological observations in field populations of Dinophysis spp. during cell cycle studies. In Harmful and Toxic Algal Blooms; Yasumoto, T., Fukuyo, Y., Eds.; IOC of UNESCO: Paris, France, 1996; pp. 257–260. [Google Scholar]
- Sun, F.; Zhou, Q.X. Metal accumulation in the polychaete Hediste japonica with emphasis on interaction between heavy metals and petroleum hydrocarbons. Environ. Pollut. 2007, 149, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Riisgård, K.; Hansen, P.J. Role of food uptake for photosynthesis, growth and survival of the mixotrophic dinoflagellate Dinophysis acuminata. Mar. Ecol. Prog. Ser. 2009, 381, 51–62. [Google Scholar] [CrossRef]
- Kim, S.; Kang, Y.G. Growth and grazing responses of the mixotrophic dinoflagellate Dinophysis acuminata as functions of light intensity and prey concentration. Aquat. Microb. Ecol. 2008, 51, 301–310. [Google Scholar] [CrossRef]
- Chou, R.; Lee, H.B. Commercial marine fish farming in Singapore. Aquac. Res. 1997, 28, 767–776. [Google Scholar] [CrossRef]
- Guillard, R.R.L. Culture of phytoplankton for feeding marine Invertebrates. In Culture of Marine Invertebrate Animals; Smith, W.L., Chanley, M.H., Eds.; Plenum Press: New York, USA, 1975; pp. 26–60. [Google Scholar]
- Nagai, S.; Matsuyama, Y.; Oh, S.J.; Itakura, S. Effects of nutrients and temperate on encystement of the toxic dinoflagellate Alexandrium tamarense (Dinophyceae) isolated from Hiroshima Bay, Japan. Plankton Biol. Ecol. 2004, 51, 103–109. [Google Scholar]
- Guillard, R.R.L. Division rates. In Handbook of Physiological Methods: Culture of Methods and Growth Measurements; Stein, J.R., Ed.; Cambridge University: Cambridge, UK, 1975; pp. 289–311. [Google Scholar]
- Suzuki, T.; Mitsuya, T.; Imai, M.; Yamasaki, M. DSP toxin contents in Dinophysis fortii and scallops collected at Mutsu Bay, Japan. J. Appl. Phycol. 1997, 8, 509–515. [Google Scholar] [CrossRef]
- Suzuki, T.; Mitsuyama, T.; Matsubara, H.; Yamasaki, M. Determination of pectenotoxin-2 after solid-phase extraction from seawater and from the dinoflagellate Dinophysis fortii by liquid chromatography with electrospray mass spectrometry and ultraviolet detection: Evidence of oxidation of pectenotoxin-2 to pectenotoxin-6 in scallops. J. Chromatogr. A 1998, 815, 155–160. [Google Scholar] [PubMed]
- Suzuki, T.; MacKenzie, L.; Stirling, D.; Adamson, J. Conversion of pectenotoxin-2 to pectenotoxin-2 seco acid in the New Zealand scallop, Pecten novaezelandiae. Fish. Sci. 2001, 67, 506–510. [Google Scholar] [CrossRef]
- Anderson, D.M.; Kulis, D.M.; Sullivan, J.J.; Hall, S.; Lee, C. Dynamics and physiology of saxitoxin production by the dinoflagellate Alexandrium spp. Mar. Biol. 1990, 104, 511–524. [Google Scholar] [CrossRef]
- Suzuki, T.; MacKenzie, L.; Stirling, D.; Adamson, J. Pectenotoxin-2 seco acid: A toxin converted from pectenotoxin-2 by the New Zealand Greenshell mussel, Perna canaliculus. Toxicon 2001, 39, 507–514. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basti, L.; Uchida, H.; Matsushima, R.; Watanabe, R.; Suzuki, T.; Yamatogi, T.; Nagai, S. Influence of Temperature on Growth and Production of Pectenotoxin-2 by a Monoclonal Culture of Dinophysis caudata. Mar. Drugs 2015, 13, 7124-7137. https://doi.org/10.3390/md13127061
Basti L, Uchida H, Matsushima R, Watanabe R, Suzuki T, Yamatogi T, Nagai S. Influence of Temperature on Growth and Production of Pectenotoxin-2 by a Monoclonal Culture of Dinophysis caudata. Marine Drugs. 2015; 13(12):7124-7137. https://doi.org/10.3390/md13127061
Chicago/Turabian StyleBasti, Leila, Hajime Uchida, Ryoji Matsushima, Ryuichi Watanabe, Toshiyuki Suzuki, Toshifumi Yamatogi, and Satoshi Nagai. 2015. "Influence of Temperature on Growth and Production of Pectenotoxin-2 by a Monoclonal Culture of Dinophysis caudata" Marine Drugs 13, no. 12: 7124-7137. https://doi.org/10.3390/md13127061