Synthetic Biology and Metabolic Engineering for Marine Carotenoids: New Opportunities and Future Prospects

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Diversity of Marine Carotenoids
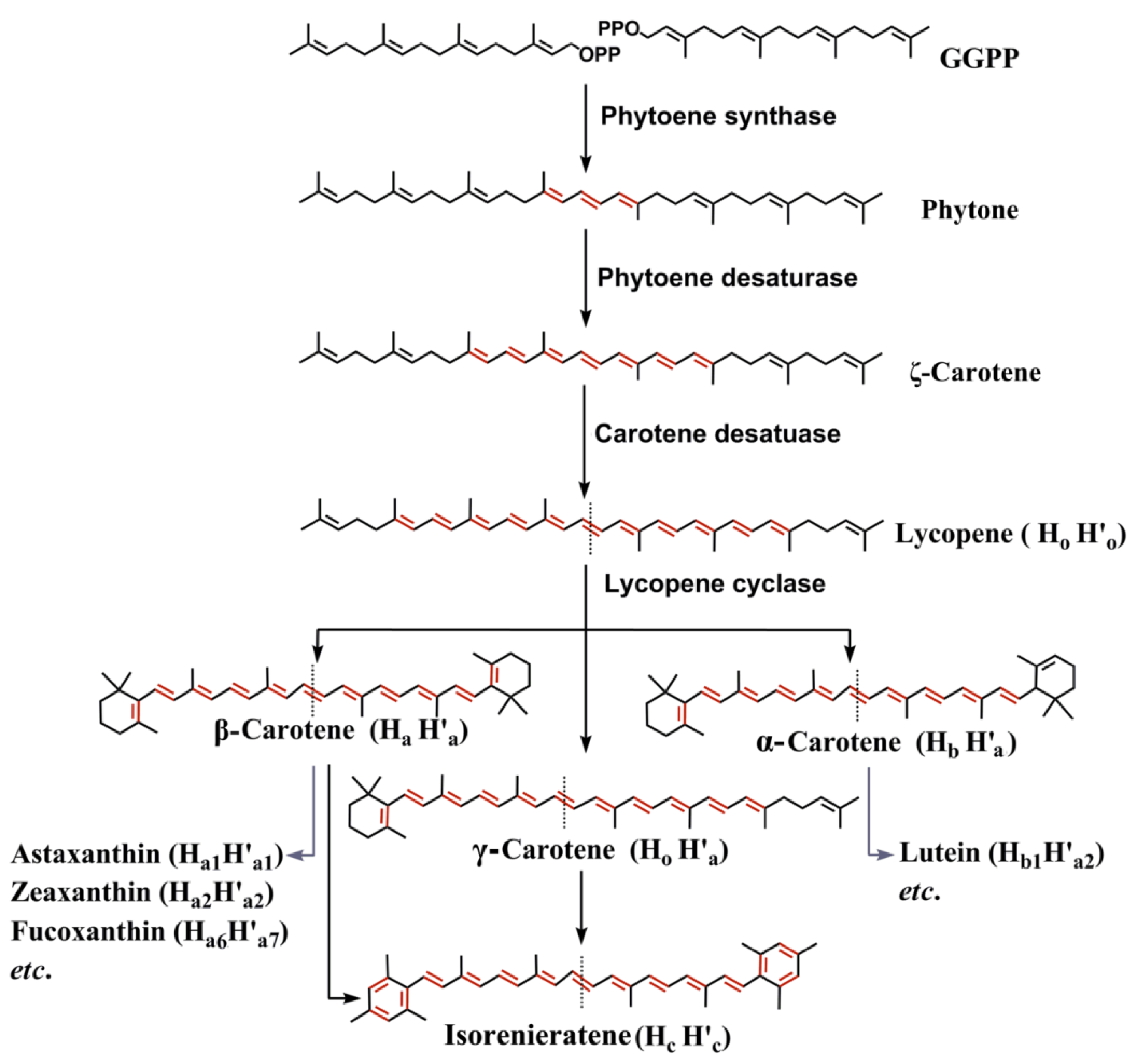
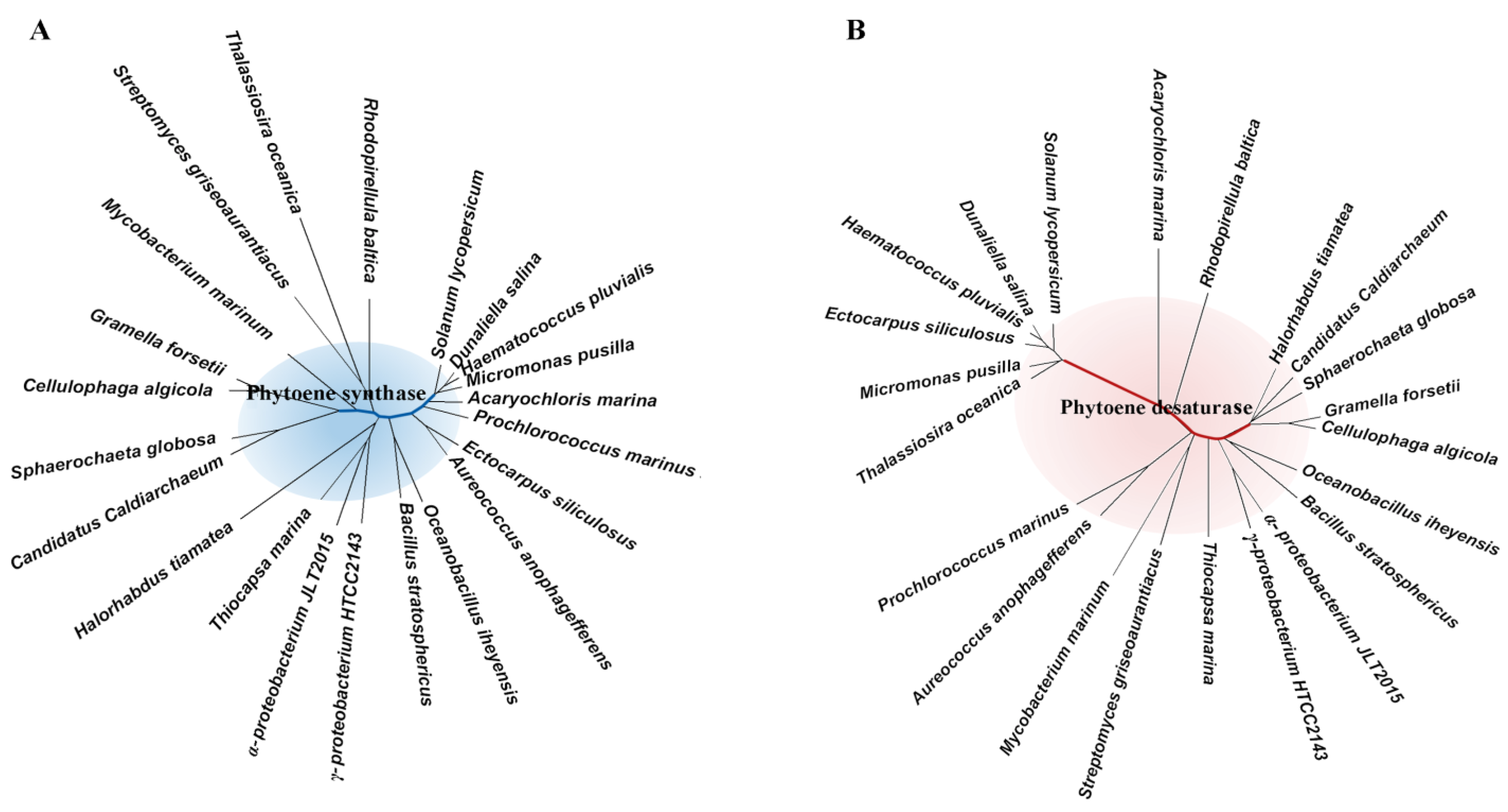
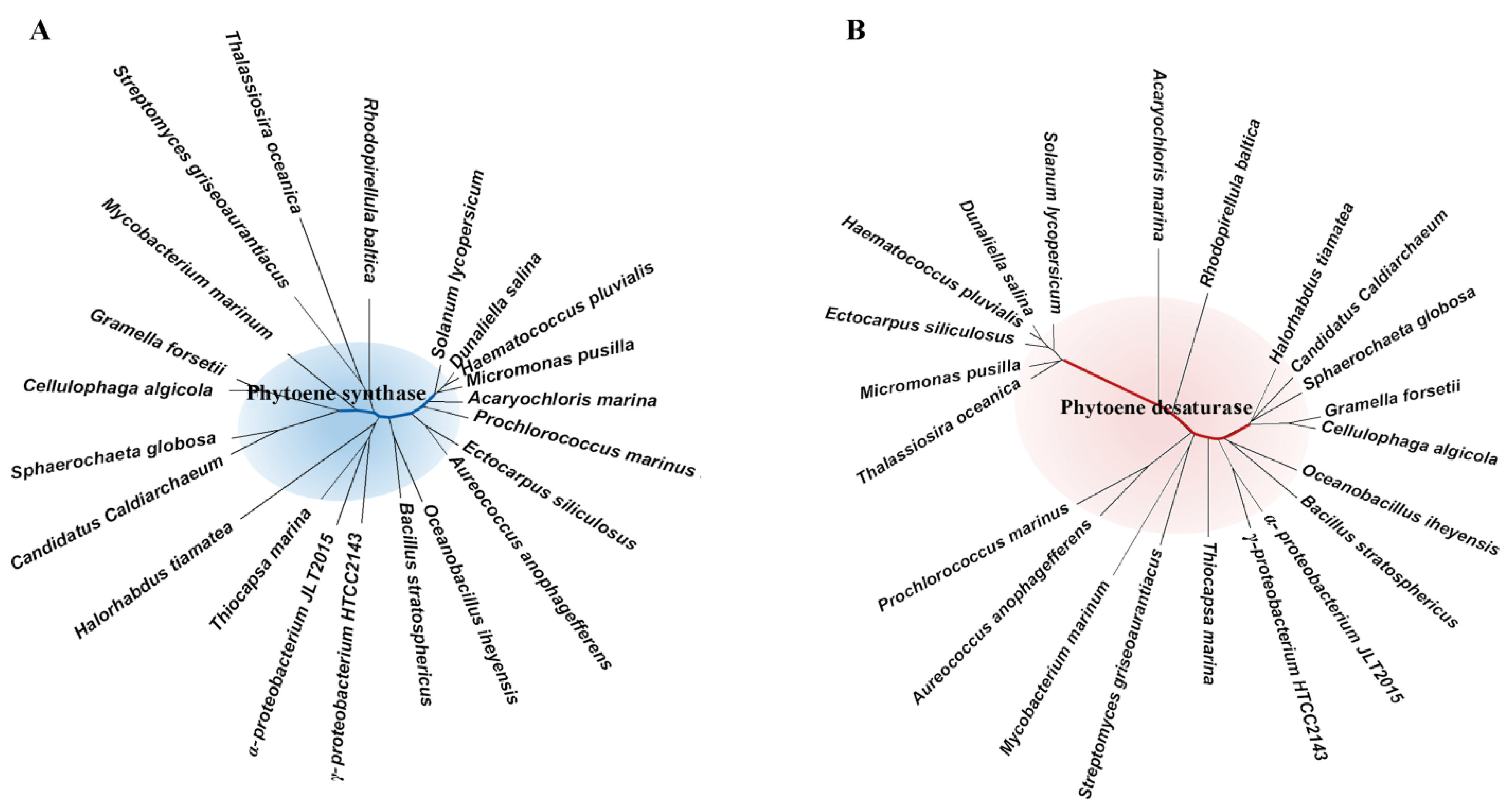
2.1. Pathways and Diverse Enzymes for Biosynthesis of Carotenoids
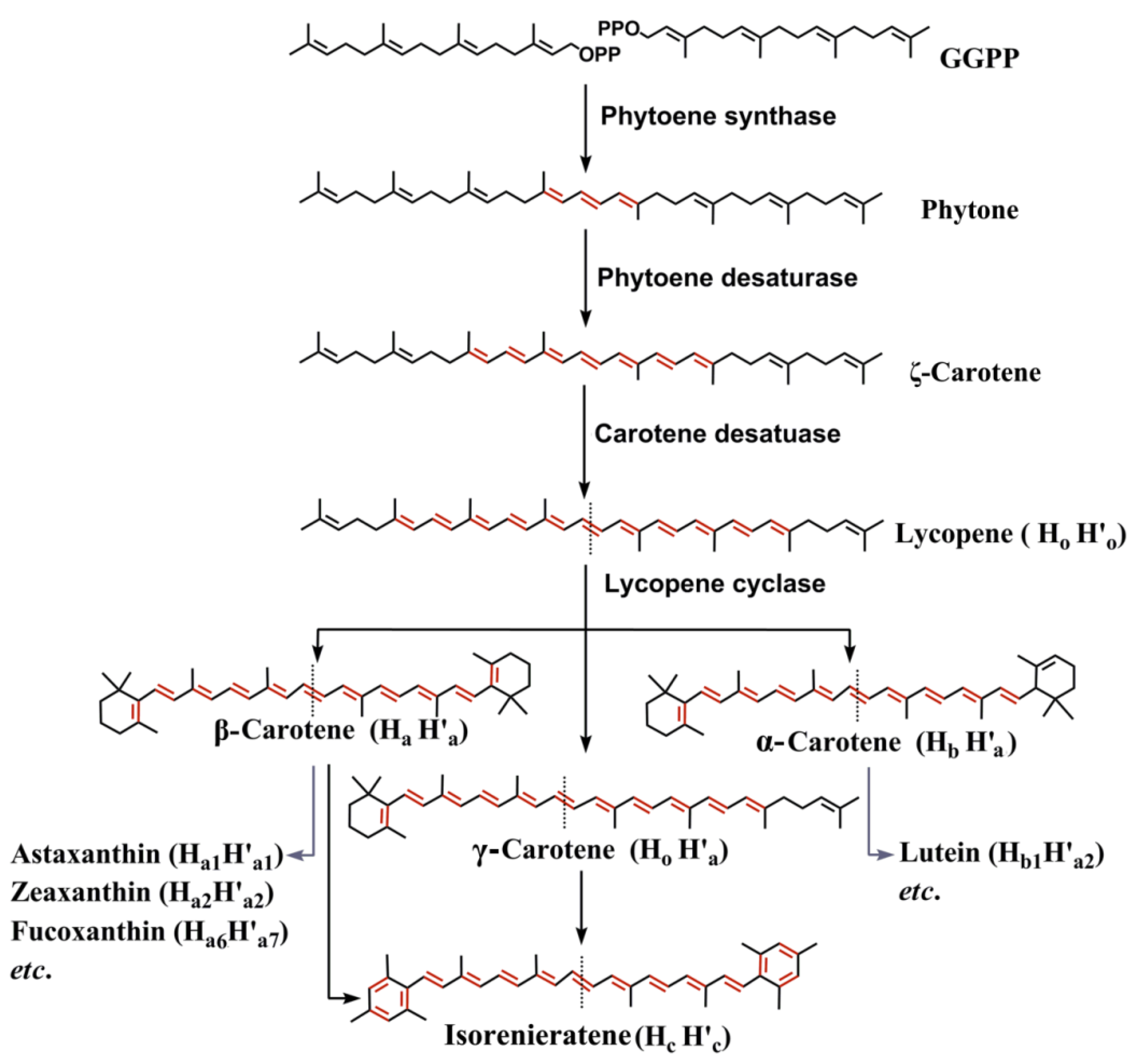
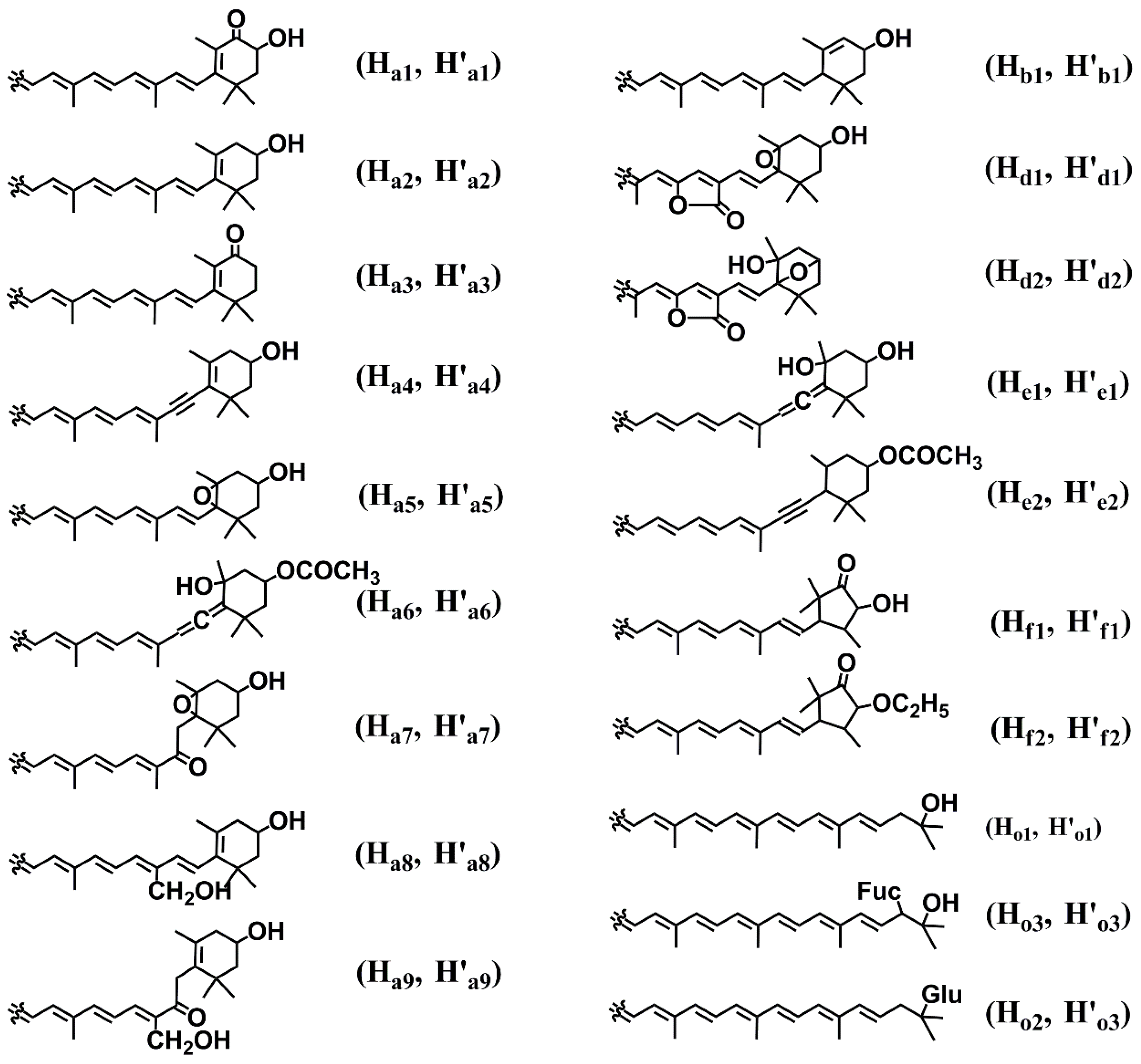
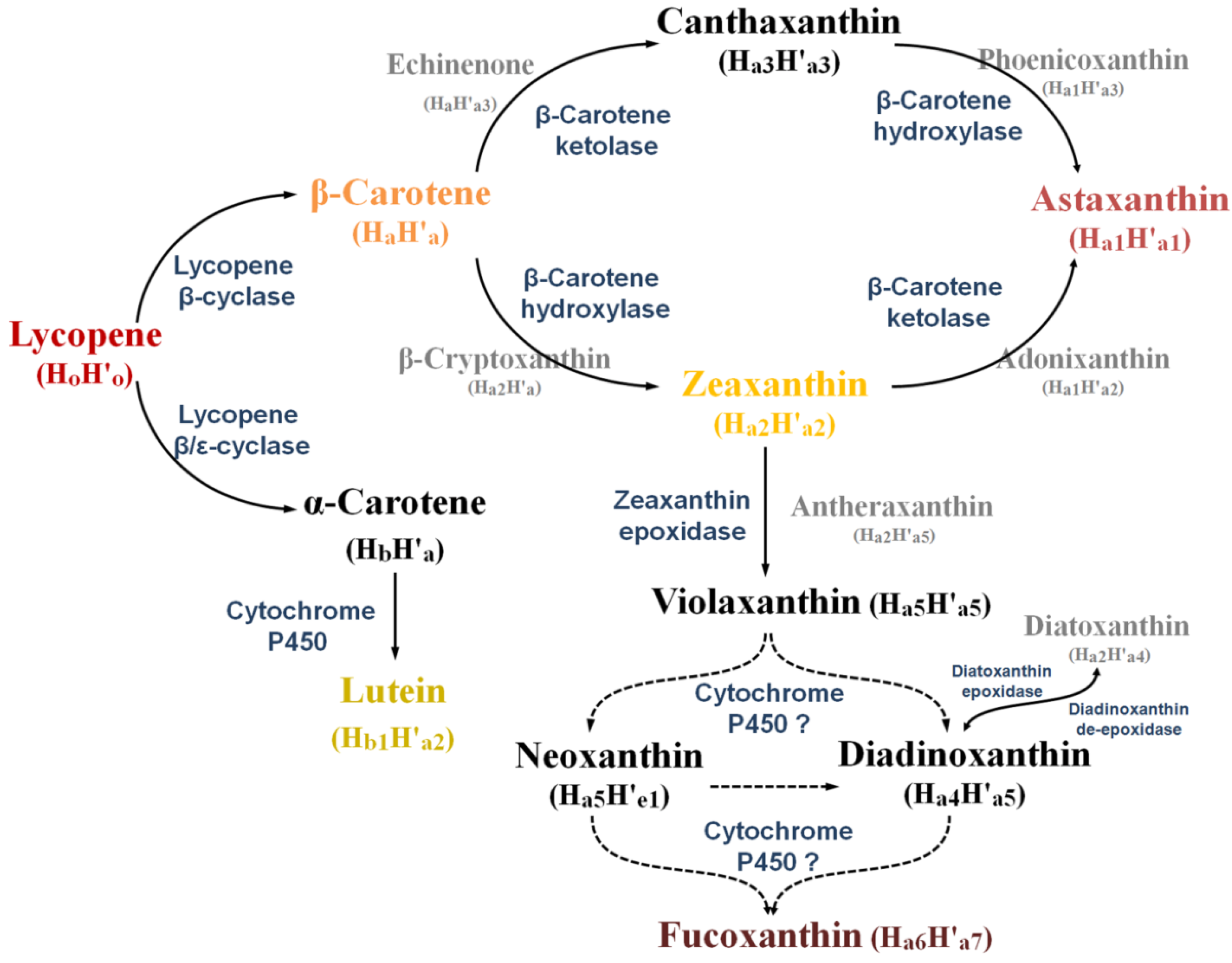
2.2. Diversity of Marine Carotenoids
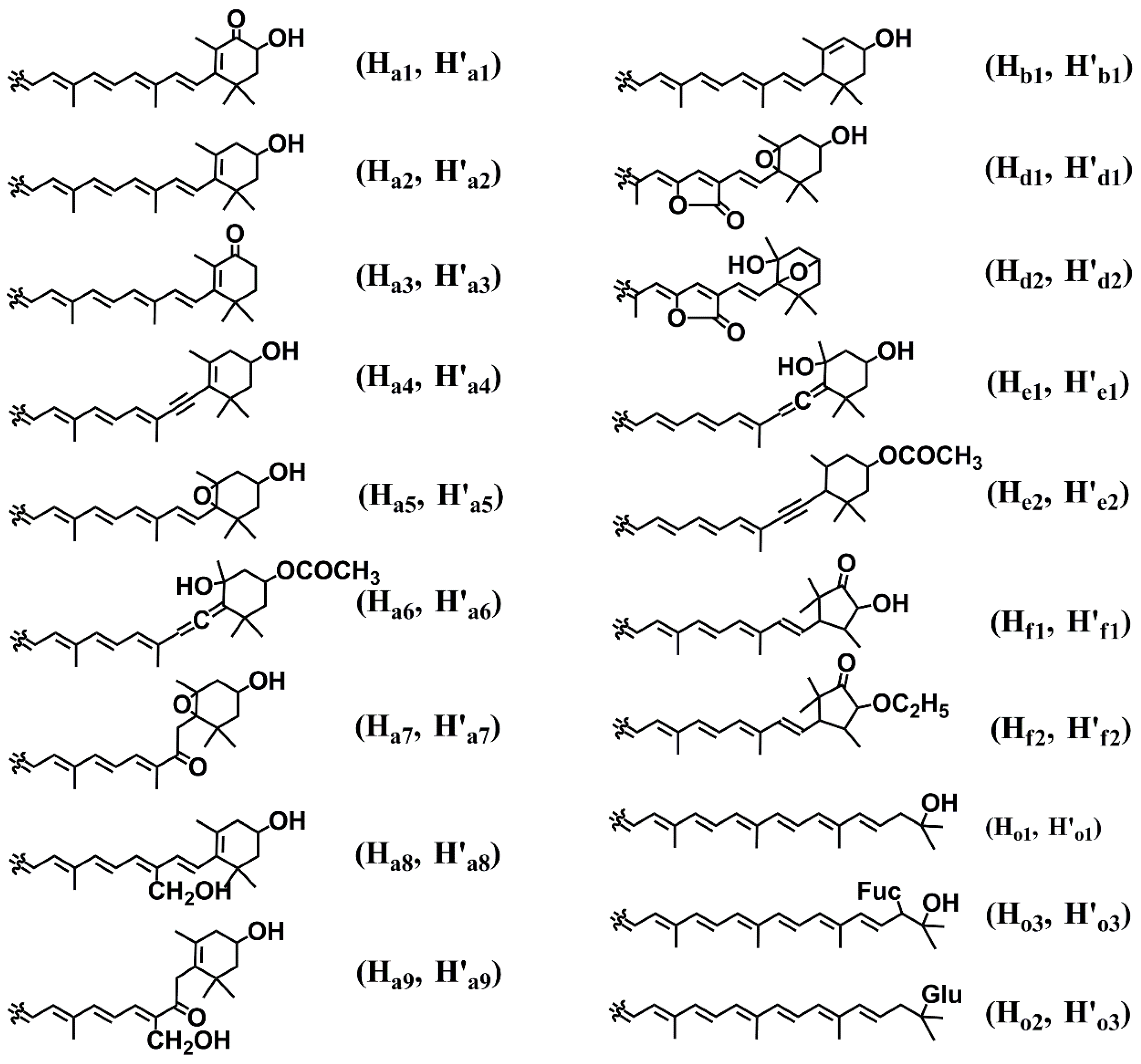
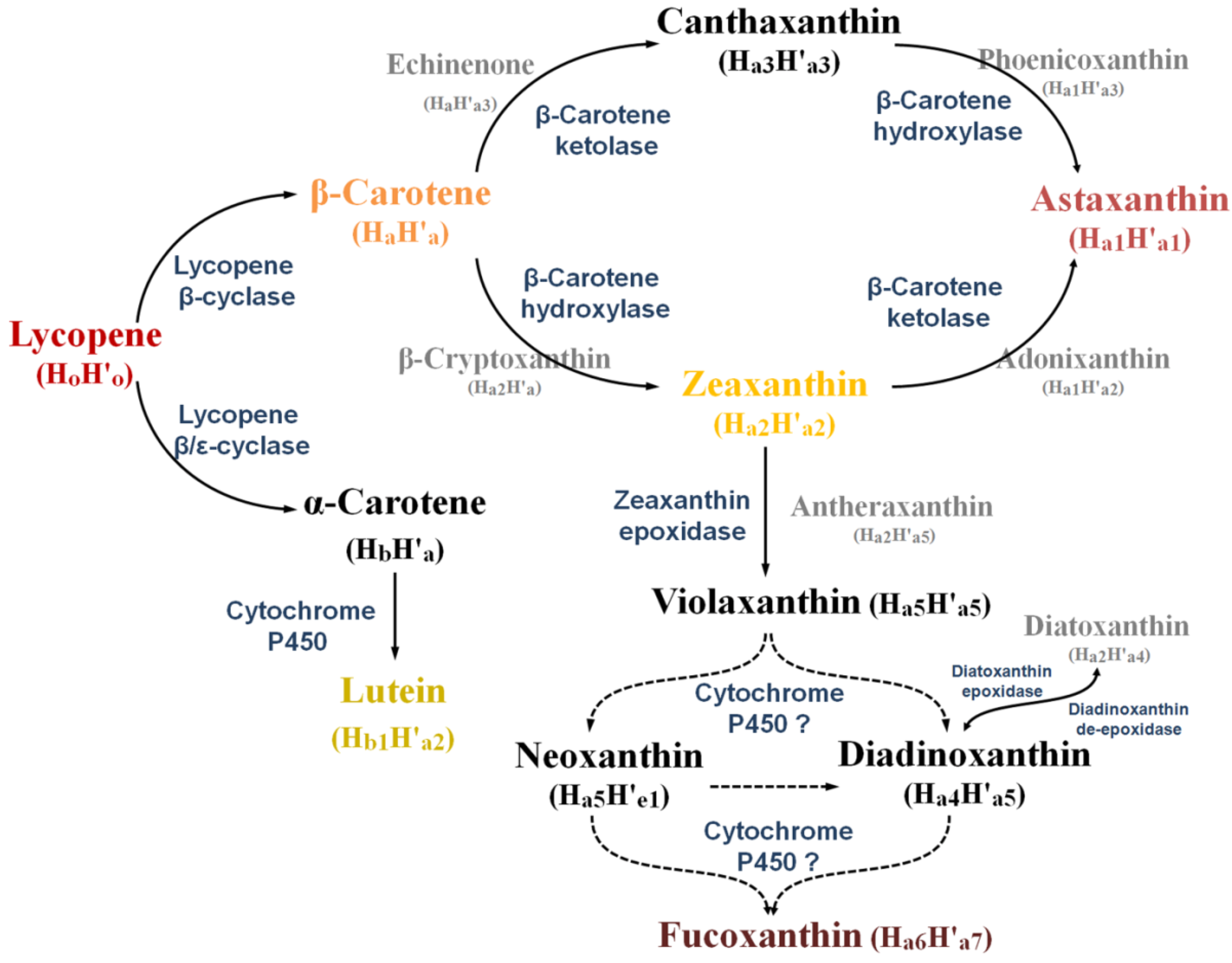
2.3. Synthesis of Some Important Marine Carotenoids and Enzymes
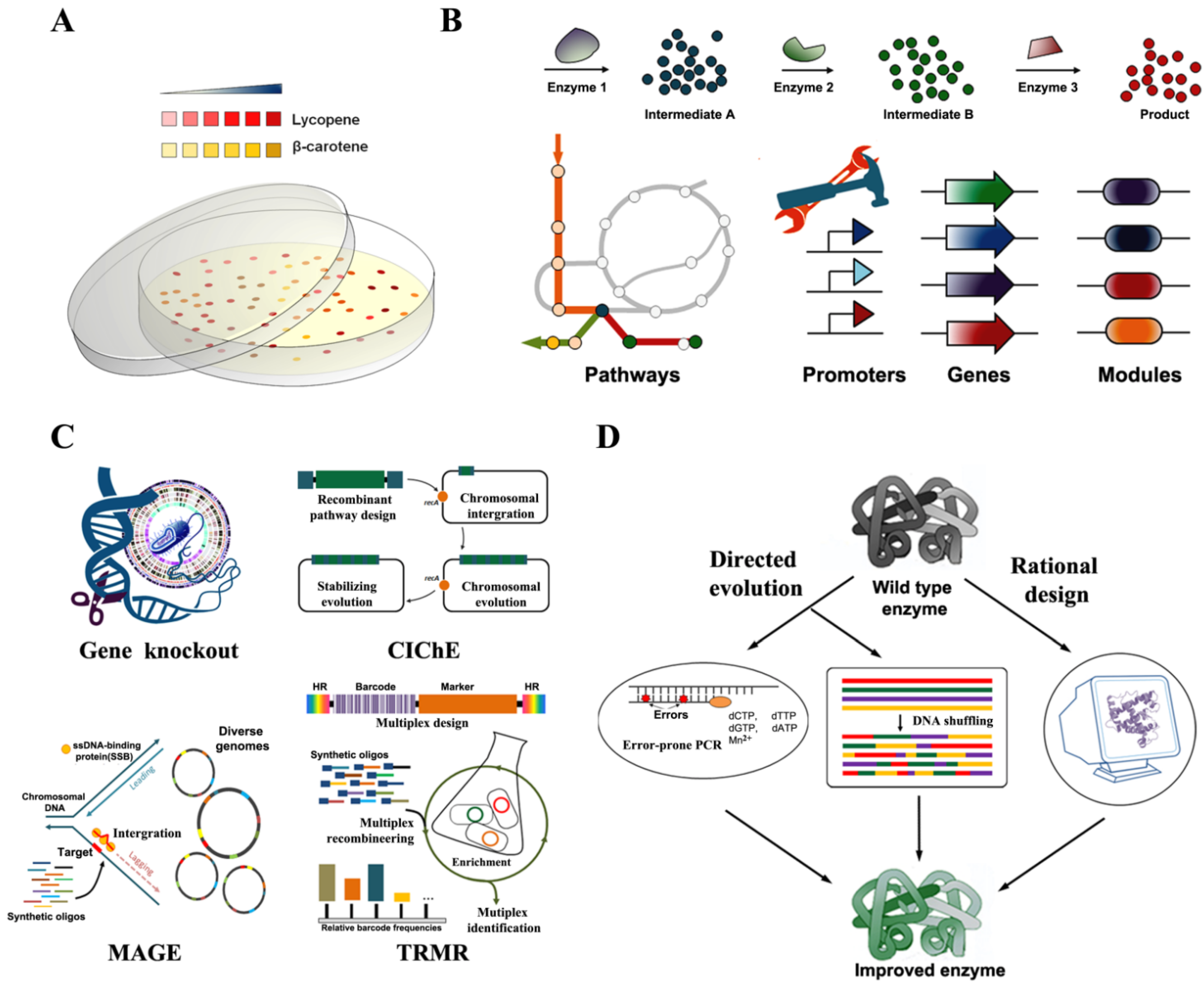
3. Technology Developments for Production of Carotenoids
3.1. Easy Colorimetric Screening of Production of Carotenoids

3.2. Pathway Engineering for Production of Carotenoids
3.3. Genome Engineering for Strain Development
3.4. Protein Engineering for Improvement of Carotenoid Production Enzymes
3.5. Development of Microalgae for Carotenoid Production
4. Opportunities and Challenges
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Cazzonelli, C. Carotenoids in nature: Insights from plants and beyond. Funct. Plant Biol. 2011, 38, 833–847. [Google Scholar] [CrossRef]
- Bertrand, M. Carotenoid biosynthesis in diatoms. Photosynth. Res. 2010, 106, 89–102. [Google Scholar] [CrossRef] [PubMed]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef] [PubMed]
- Shindo, K.; Misawa, N. New and rare carotenoids isolated from marine bacteria and their antioxidant activities. Mar. Drugs 2014, 12, 1690–1698. [Google Scholar] [CrossRef] [PubMed]
- Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids Handbook; Birkhäuser: Basel, Switzerland, 2004. [Google Scholar]
- Armstrong, G.A.; Hearst, J.E. Carotenoids 2: Genetics and molecular biology of carotenoid pigment biosynthesis. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1996, 10, 228–237. [Google Scholar]
- Mathews, M.M.; Sistrom, W.R. The function of the carotenoid pigments of Sarcina lutea. Archiv. Mikrobiol. 1960, 35, 139–146. [Google Scholar] [CrossRef]
- Liu, J.; Novero, M.; Charnikhova, T.; Ferrandino, A.; Schubert, A.; Ruyter-Spira, C.; Bonfante, P.; Lovisolo, C.; Bouwmeester, H.J.; Cardinale, F. Carotenoid cleavage dioxygenase 7 modulates plant growth, reproduction, senescence, and determinate nodulation in the model legume Lotus japonicus. J. Exp. Bot. 2013, 64, 1967–1981. [Google Scholar] [CrossRef] [PubMed]
- Bolhassani, A.; Khavari, A.; Bathaie, S.Z. Saffron and natural carotenoids: Biochemical activities and anti-tumor effects. Biochim. Biophys. Acta 2014, 1845, 20–30. [Google Scholar]
- Rodrigues, E.; Mariutti, L.R.; Mercadante, A.Z. Scavenging capacity of marine carotenoids against reactive oxygen and nitrogen species in a membrane-mimicking system. Mar. Drugs 2012, 10, 1784–1798. [Google Scholar] [CrossRef] [PubMed]
- Van Den Berg, H.; Faulks, R.; Fernando Granado, H.; Hirschberg, J.; Olmedilla, B.; Sandmann, G.; Southon, S.; Stahl, W. The potential for the improvement of carotenoid levels in foods and the likely systemic effects. J. Sci. Food Agric. 2000, 80, 880–912. [Google Scholar]
- Fernandez-Sevilla, J.M.; Acien Fernandez, F.G.; Molina Grima, E. Biotechnological production of lutein and its applications. Appl. Microbiol. Biotechnol. 2010, 86, 27–40. [Google Scholar]
- Kirsh, V.A.; Mayne, S.T.; Peters, U.; Chatterjee, N.; Leitzmann, M.F.; Dixon, L.B.; Urban, D.A.; Crawford, E.D.; Hayes, R.B. A prospective study of lycopene and tomato product intake and risk of prostate cancer. Cancer Epidemiol. Biomark. Prev. Publ. Am. Assoc. Cancer Res. Cosponsored Am. Soc. Prev. Oncol. 2006, 15, 92–98. [Google Scholar] [CrossRef]
- Wang, W.; Shinto, L.; Connor, W.E.; Quinn, J.F. Nutritional biomarkers in alzheimer’s disease: The association between carotenoids, n-3 fatty acids, and dementia severity. J. Alzheimer’s Dis. JAD 2008, 13, 31–38. [Google Scholar]
- Cosgrove, J. The carotenoid market: Beyond beta-carotene. Nutraceuticals World. 13 December 2010. Available online: http://www.nutraceuticalsworld.com/contents/view_online-exclusives/2010-12-13/the-carotenoid-market-beyond-beta-carotene/ (accessed on 5 September 2014).
- Rodriguez-Saiz, M.; de la Fuente, J.L.; Barredo, J.L. Xanthophyllomyces dendrorhous for the industrial production of astaxanthin. Appl. Microbiol. Biotechnol. 2010, 88, 645–658. [Google Scholar]
- Kim, Y.S.; Lee, J.H.; Kim, N.H.; Yeom, S.J.; Kim, S.W.; Oh, D.K. Increase of lycopene production by supplementing auxiliary carbon sources in metabolically engineered Escherichia coli. Appl. Microbiol. Biotechnol. 2011, 90, 489–497. [Google Scholar] [CrossRef] [PubMed]
- Vilchez, C.; Forjan, E.; Cuaresma, M.; Bedmar, F.; Garbayo, I.; Vega, J.M. Marine carotenoids: Biological functions and commercial applications. Mar. Drugs 2011, 9, 319–333. [Google Scholar] [CrossRef] [PubMed]
- Miziorko, H.M. Enzymes of the mevalonate pathway of isoprenoid biosynthesis. Arch. Biochem. Biophys. 2011, 505, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Hunter, W.N. The non-mevalonate pathway of isoprenoid precursor biosynthesis. J. Biol. Chem. 2007, 282, 21573–21577. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Kim, J.Y.; Choi, E.S.; Kim, S.W. Microbial production of farnesol (FOH): Current states and beyond. Process. Biochem. 2011, 46, 1221–1229. [Google Scholar] [CrossRef]
- Rohdich, F.; Kis, K.; Bacher, A.; Eisenreich, W. The non-mevalonate pathway of isoprenoids: Genes, enzymes and intermediates. Curr. Opin. Chem. Biol. 2001, 5, 535–540. [Google Scholar] [CrossRef] [PubMed]
- Fujisaki, S.; Hara, H.; Nishimura, Y.; Horiuchi, K.; Nishino, T. Cloning and nucleotide sequence of the ispA gene responsible for farnesyl diphosphate synthase activity in Escherichia coli. J. Biochem. 1990, 108, 995–1000. [Google Scholar] [PubMed]
- Math, S.K.; Hearst, J.E.; Poulter, C.D. The crtE gene in Erwinia herbicola encodes geranylgeranyl diphosphate synthase. Proc. Natl. Acad. Sci. USA 1992, 89, 6761–6764. [Google Scholar] [CrossRef] [PubMed]
- Raisig, A.; Bartley, G.; Scolnik, P.; Sandmann, G. Purification in an active state and properties of the 3-step phytoene desaturase from Rhodobacter capsulatus overexpressed in Escherichia coli. J. Biochem. 1996, 119, 559–564. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Asuaa, G.; Langa, H.; Cogdellb, R.; Hunter, C.N. Carotenoid diversity: A modular role for the phytoene desaturase step. Trends Plant Sci. 1998, 3, 445–449. [Google Scholar]
- Takaichi, S.; Mochimaru, M. Carotenoids and carotenogenesis in cyanobacteria: Unique ketocarotenoids and carotenoid glycosides. Cell. Mol. Life Sci. CMLS 2007, 64, 2607–2619. [Google Scholar] [CrossRef]
- Takaichi, S. Carotenoids in algae: Distributions, biosyntheses and functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef] [PubMed]
- Umeno, D.; Tobias, A.V.; Arnold, F.H. Diversifying carotenoid biosynthetic pathways by directed evolution. Microbiol. Mol. Biol. Rev. MMBR 2005, 69, 51–78. [Google Scholar] [CrossRef]
- Schaub, P.; Yu, Q.; Gemmecker, S.; Poussin-Courmontagne, P.; Mailliot, J.; McEwen, A.G.; Ghisla, S.; Al-Babili, S.; Cavarelli, J.; Beyer, P. On the structure and function of the phytoene desaturase CrtI from Pantoea ananatis, a membrane-peripheral and FAD-dependent oxidase/isomerase. PLoS One 2012, 7, e39550. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.I.; Liu, G.Y.; Song, Y.; Yin, F.; Hensler, M.E.; Jeng, W.Y.; Nizet, V.; Wang, A.H.; Oldfield, E. A cholesterol biosynthesis inhibitor blocks Staphylococcus aureus virulence. Science 2008, 319, 1391–1394. [Google Scholar] [CrossRef] [PubMed]
- Tobias, A.V.; Arnold, F.H. Biosynthesis of novel carotenoid families based on unnatural carbon backbones: A model for diversification of natural product pathways. Biochim. Biophys. Acta 2006, 1761, 235–246. [Google Scholar] [CrossRef]
- Martin, W.; Rujan, T.; Richly, E.; Hansen, A.; Cornelsen, S.; Lins, T.; Leister, D.; Stoebe, B.; Hasegawa, M.; Penny, D. Evolutionary analysis of Arabidopsis, cyanobacterial, and chloroplast genomes reveals plastid phylogeny and thousands of cyanobacterial genes in the nucleus. Proc. Natl. Acad. Sci. USA 2002, 99, 12246–12251. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Prieto, A.; Hackett, J.D.; Soares, M.B.; Bonaldo, M.F.; Bhattacharya, D. Cyanobacterial contribution to algal nuclear genomes is primarily limited to plastid functions. Curr. Biol. CB 2006, 16, 2320–2325. [Google Scholar]
- Sandmann, G. Evolution of carotene desaturation: The complication of a simple pathway. Arch. Biochem. Biophys. 2009, 483, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Steiger, S.; Jackisch, Y.; Sandmann, G. Carotenoid biosynthesis in Gloeobacter violaceus PCC4721 involves a single CrtI-type phytoene desaturase instead of typical cyanobacterial enzymes. Arch. Microbiol. 2005, 184, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Tsuchiya, T.; Takaichi, S.; Misawa, N.; Maoka, T.; Miyashita, H.; Mimuro, M. The cyanobacterium Gloeobacter violaceus PCC4721 uses bacterial-type phytoene desaturase in carotenoid biosynthesis. FEBS Lett. 2005, 579, 2125–2129. [Google Scholar] [CrossRef] [PubMed]
- Harada, J.; Nagashima, K.V.; Takaichi, S.; Misawa, N.; Matsuura, K.; Shimada, K. Phytoene desaturase, CrtI, of the purple photosynthetic bacterium, Rubrivivax gelatinosus, produces both neurosporene and lycopene. Plant Cell Physiol. 2001, 42, 1112–1118. [Google Scholar] [CrossRef] [PubMed]
- Frigaard, N.U.; Maresca, J.A.; Yunker, C.E.; Jones, A.D.; Bryant, D.A. Genetic manipulation of carotenoid biosynthesis in the green sulfur bacterium Chlorobium tepidum. J. Bacterial. 2004, 186, 5210–5220. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar]
- Lamers, P.P.; Janssen, M.; de Vos, R.C.; Bino, R.J.; Wijffels, R.H. Exploring and exploiting carotenoid accumulation in Dunaliella salina for cell-factory applications. Trends Biotechnol. 2008, 26, 631–638. [Google Scholar] [CrossRef] [PubMed]
- Rabbani, S.; Beyer, P.; Lintig, J.; Hugueney, P.; Kleinig, H. Induced β-carotene synthesis driven by triacylglycerol deposition in the unicellular alga Dunaliella bardawil. Plant Physiol. 1998, 116, 1239–1248. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.; Yu, X.; Wang, Y.; Cui, Y.; Li, X.; Liu, Z.; Qin, S. Evolutionary origins, molecular cloning and expression of carotenoid hydroxylases in eukaryotic photosynthetic algae. BMC Genomics 2013, 14, 457. [Google Scholar] [CrossRef] [PubMed]
- El-Baky, H.A.; BaZ, F.E.; El-Baroty, G. Spirulina species as a source of carotenoids and α-tocopherol and its anticarcinoma factors. Biotechnology 2003, 2, 222–240. [Google Scholar]
- Choubert, G.; Heinrich, O. Carotenoid pigments of the green alga Haematococcus pluvialis: Assay on rainbow trout, Oncorhynchus mykiss, pigmentation in comparison with synthetic astaxanthin and canthaxanthin. Aquaculture 1993, 112, 217–226. [Google Scholar] [CrossRef]
- Abe, K.; Hattori, H.; Hirano, M. Accumulation and antioxidant activity of secondary carotenoids in the aerial microalga Coelastrella striolata var. Multistriata. Food Chem. 2007, 100, 656–661. [Google Scholar] [CrossRef]
- Sanchez, J.F.; Fernandez-Sevilla, J.M.; Acien, F.G.; Ceron, M.C.; Perez-Parra, J.; Molina-Grima, E. Biomass and lutein productivity of Scenedesmus almeriensis: Influence of irradiance, dilution rate and temperature. Appl. Microbiol. Biotechnol. 2008, 79, 719–729. [Google Scholar] [CrossRef] [PubMed]
- Fietz, S.; Nicklisch, A. Acclimation of the diatom Stephanodiscus neoastraea and the cyanobacterium Planktothrix agardhii to simulated natural light fluctuations. Photosynth. Res. 2002, 72, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S. Distributions, biosyntheses and functions of carotenoids in algae. Agro FOOD Industry Hi Tech. 2013, 24, 55–58. [Google Scholar]
- Kupper, H.; Seibert, S.; Parameswaran, A. Fast, sensitive, and inexpensive alternative to analytical pigment HPLC: Quantification of chlorophylls and carotenoids in crude extracts by fitting with Gauss peak spectra. Anal. Chem. 2007, 79, 7611–7627. [Google Scholar] [CrossRef] [PubMed]
- Miyashita, K.; Nishikawa, S.; Beppu, F.; Tsukui, T.; Abe, M.; Hosokawa, M. The allenic carotenoid fucoxanthin, a novel marine nutraceutical from brown seaweeds. J. Sci. Food Agric. 2011, 91, 1166–1174. [Google Scholar] [CrossRef] [PubMed]
- Aitzetmüller, K.; Strain, H.H.; Svec, W.A.; Grandolfo, M.; Katz, J.J. Loroxanthin, a unique xanthophyll from Scenedesmus obliquus and Chlorella vulgaris. Phytochemistry 1969, 1761–1770. [Google Scholar]
- Ganesan, P.; Matsubara, K.; Ohkubo, T.; Tanaka, Y.; Noda, K.; Sugawara, T.; Hirata, T. Anti-angiogenic effect of siphonaxanthin from green alga, Codium fragile. Phytomed. Int. J. Phytother. Phytopharmacol. 2010, 17, 1140–1144. [Google Scholar] [CrossRef] [Green Version]
- Maresca, J.A.; Romberger, S.P.; Bryant, D.A. Isorenieratene biosynthesis in green sulfur bacteria requires the cooperative actions of two carotenoid cyclases. J. Bacterial. 2008, 190, 6384–6391. [Google Scholar] [CrossRef]
- Brocks, J.J.; Love, G.D.; Summons, R.E.; Knoll, A.H.; Logan, G.A.; Bowden, S.A. Biomarker evidence for green and purple sulphur bacteria in a stratified Palaeoproterozoic sea. Nature 2005, 437, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.E.; Bryant, D.A. The biosynthetic pathway for myxol-2' fucoside (myxoxanthophyll) in the cyanobacterium Synechococcus sp. strain PCC 7002. J. Bacterial. 2009, 191, 3292–3300. [Google Scholar]
- Song, P.S.; Koka, P.; Prezelin, B.B.; Haxo, F.T. Molecular topology of the photosynthetic light-harvesting pigment complex, peridinin-chlorophyll A-protein, from marine dinoflagellates. Biochemistry 1976, 15, 4422–4427. [Google Scholar] [CrossRef] [PubMed]
- Maoka, T.; Hashimoto, K.; Akimoto, N.; Fujiwara, Y. Structures of five new carotenoids from the oyster Crassostrea gigas. J. Nat. Prod. 2001, 64, 578–581. [Google Scholar] [CrossRef] [PubMed]
- Maoka, T.; Akimoto, N.; Yim, M.J.; Hosokawa, M.; Miyashita, K. New C37 skeletal carotenoid from the clam, Paphia amabillis. J. Agric. Food Chem. 2008, 56, 12069–12072. [Google Scholar] [CrossRef] [PubMed]
- Maoka, T.; Fujiwara, Y.; Hashimoto, K.; Akimoto, N. Structure of new carotenoids from corbicula clam Corbicula japonica. J. Nat. Prod. 2005, 68, 1341–1344. [Google Scholar] [CrossRef] [PubMed]
- Maoka, T. Carotenoids in marine animals. Mar. Drugs 2011, 9, 278–293. [Google Scholar] [CrossRef] [PubMed]
- Maresca, J.A.; Graham, J.E.; Wu, M.; Eisen, J.A.; Bryant, D.A. Identification of a fourth family of lycopene cyclases in photosynthetic bacteria. Proc. Natl. Acad. Sci. USA 2007, 104, 11784–11789. [Google Scholar] [CrossRef] [PubMed]
- Misawa, N. Carotenoid beta-ring hydroxylase and ketolase from marine bacteria-promiscuous enzymes for synthesizing functional xanthophylls. Mar. Drugs 2011, 9, 757–771. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; DellaPenna, D. Defining the primary route for lutein synthesis in plants: The role of Arabidopsis carotenoid β-ring hydroxylase CYP97A3. Proc. Natl. Acad. Sci. USA 2006, 103, 3474–3479. [Google Scholar] [CrossRef] [PubMed]
- Coesel, S.; Obornik, M.; Varela, J.; Falciatore, A.; Bowler, C. Evolutionary origins and functions of the carotenoid biosynthetic pathway in marine diatoms. PLoS One 2008, 3, e2896. [Google Scholar] [CrossRef] [PubMed]
- Mikami, K.; Hosokawa, M. Biosynthetic pathway and health benefits of fucoxanthin, an algae-specific xanthophyll in brown seaweeds. Int. J. Mol. Sci. 2013, 14, 13763–13781. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.E.; Huang, X.Q.; Hang, Y.; Deng, Y.Y.; Lu, Q.Q.; Lu, S. The P450-type carotene hydroxylase PuCHY1 from Porphyra suggested the evolution of carotenoid metabolism in red algae. J. Integr. Plant Boil. 2014, in press. [Google Scholar]
- Misawa, N.; Satomi, Y.; Kondo, K.; Yokoyama, A.; Kajiwara, S.; Saito, T.; Ohtani, T.; Miki, W. Structure and functional analysis of a marine bacterial carotenoid biosynthesis gene cluster and astaxanthin biosynthetic pathway proposed at the gene level. J. Bacteriol. 1995, 177, 6575–6584. [Google Scholar] [PubMed]
- Netzer, R.; Stafsnes, M.H.; Andreassen, T.; Goksoyr, A.; Bruheim, P.; Brautaset, T. Biosynthetic pathway for γ-cyclic sarcinaxanthin in Micrococcus luteus: Heterologous expression and evidence for diverse and multiple catalytic functions of C50 carotenoid cyclases. J. Bacteriol. 2010, 192, 5688–5699. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Hu, X.; Wang, L.; Wang, X. Reconstruction of the carotenoid biosynthetic pathway of Cronobacter sakazakii BAA894 in Escherichia coli. PLoS One 2014, 9, e86739. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.W.; Keasling, J.D. Metabolic engineering of the nonmevalonate isopentenyl diphosphate synthesis pathway in Escherichia coli enhances lycopene production. Biotechnol. Bioeng. 2001, 72, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Harker, M.; Bramley, P.M. Expression of prokaryotic 1-deoxy-d-xylulose-5-phosphatases in Escherichia coli increases carotenoid and ubiquinone biosynthesis. FEBS Lett. 1999, 448, 115–119. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.S.; Grammel, H.; Abou-Aisha, K.; Sagesser, R.; Ghosh, R. High-Level production of the industrial product lycopene by the photosynthetic bacterium Rhodospirillum rubrum. Appl. Environ. Microbiol. 2012, 78, 7205–7215. [Google Scholar] [CrossRef] [PubMed]
- Miura, Y.; Kondo, K.; Saito, T.; Shimada, H.; Fraser, P.D.; Misawa, N. Production of the carotenoids lycopene, β-carotene, and astaxanthin in the food yeast Candida utilis. Appl. Environ. Microbiol. 1998, 64, 1226–1229. [Google Scholar] [PubMed]
- Harada, H.; Misawa, N. Novel approaches and achievements in biosynthesis of functional isoprenoids in Escherichia coli. Appl. Microbiol. Biotechnol. 2009, 84, 1021–1031. [Google Scholar] [CrossRef] [PubMed]
- Yamano, S.; Ishii, T.; Nakagawa, M.; Ikenaga, H.; Misawa, N. Metabolic engineering for production of β-carotene and lycopene in Saccharomyces cerevisiae. Biosci. Biotechnol. Biochem. 1994, 58, 1112–1114. [Google Scholar] [CrossRef] [PubMed]
- Heider, S.A.; Peters-Wendisch, P.; Netzer, R.; Stafnes, M.; Brautaset, T.; Wendisch, V.F. Production and glucosylation of C50 and C40 carotenoids by metabolically engineered Corynebacterium glutamicum. Appl. Microbiol. Biotechnol. 2014, 98, 1223–1235. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Kim, J.E.; Lee, S.H.; Park, H.M.; Choi, M.S.; Kim, J.Y.; Shin, Y.C.; Keasling, J.D.; Kim, S.W. Engineering the lycopene synthetic pathway in E. coli by comparison of the carotenoid genes of Pantoea agglomerans and Pantoea ananatis. Appl. Microbiol. Biotechnol. 2007, 74, 131–139. [Google Scholar]
- Sandmann, G.; Albrecht, M.; Schnurr, G.; Knorzer, O.; Boger, P. The biotechnological potential and design of novel carotenoids by gene combination in Escherichia coli. Trends Biotechnol. 1999, 17, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Misawa, N. Pathway engineering for functional isoprenoids. Curr. Opin. Biotechnol. 2011, 22, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Park, H.M.; Kim, J.E.; Lee, S.H.; Choi, M.S.; Kim, J.Y.; Oh, D.K.; Keasling, J.D.; Kim, S.W. Increased β-carotene production in recombinant Escherichia coli harboring an engineered isoprenoid precursor pathway with mevalonate addition. Biotechnol. Prog. 2007, 23, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Verwaal, R.; Wang, J.; Meijnen, J.P.; Visser, H.; Sandmann, G.; van den Berg, J.A.; van Ooyen, A.J. High-level production of beta-carotene in Saccharomyces cerevisiae by successive transformation with carotenogenic genes from Xanthophyllomyces dendrorhous. Appl. Environ. Microbiol. 2007, 73, 4342–4350. [Google Scholar] [CrossRef] [PubMed]
- Yoon, S.H.; Lee, S.H.; Das, A.; Ryu, H.K.; Jang, H.J.; Kim, J.Y.; Oh, D.K.; Keasling, J.D.; Kim, S.W. Combinatorial expression of bacterial whole mevalonate pathway for the production of β-carotene in E. coli. J. Biotechnol. 2009, 140, 218–226. [Google Scholar] [CrossRef]
- Yoshida, K.; Ueda, S.; Maeda, I. Carotenoid production in Bacillus subtilis achieved by metabolic engineering. Biotechnol. Lett. 2009, 31, 1789–1793. [Google Scholar] [CrossRef] [PubMed]
- Beuttler, H.; Hoffmann, J.; Jeske, M.; Hauer, B.; Schmid, R.D.; Altenbuchner, J.; Urlacher, V.B. Biosynthesis of zeaxanthin in recombinant Pseudomonas putida. Appl. Microbiol. Biotechnol. 2011, 89, 1137–1147. [Google Scholar] [CrossRef] [PubMed]
- Ye, V.M.; Bhatia, S.K. Pathway engineering strategies for production of beneficial carotenoids in microbial hosts. Biotechnol. Lett. 2012, 34, 1405–1414. [Google Scholar] [CrossRef] [PubMed]
- Vick, J.E.; Johnson, E.T.; Choudhary, S.; Bloch, S.E.; Lopez-Gallego, F.; Srivastava, P.; Tikh, I.B.; Wawrzyn, G.T.; Schmidt-Dannert, C. Optimized compatible set of Biobrick vectors for metabolic pathway engineering. Appl. Microbiol. Biotechnol. 2011, 92, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Z.; Elledge, S.J. Harnessing homologous recombination in vitro to generate recombinant DNA via SLIC. Nat. Methods 2007, 4, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Gibson, D.G.; Young, L.; Chuang, R.Y.; Venter, J.C.; Hutchison, C.A., 3rd; Smith, H.O. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat. Methods 2009, 6, 343–345. [Google Scholar]
- Wingler, L.M.; Cornish, V.W. Reiterative recombination for the in vivo assembly of libraries of multigene pathways. Proc. Natl. Acad. Sci. USA 2011, 108, 15135–15140. [Google Scholar] [CrossRef] [PubMed]
- Sleight, S.C.; Sauro, H.M. Randomized Biobrick assembly: A novel DNA assembly method for randomizing and optimizing genetic circuits and metabolic pathways. ACS Synth. Boil. 2013, 2, 506–518. [Google Scholar] [CrossRef]
- Yadav, V.G.; de Mey, M.; Lim, C.G.; Ajikumar, P.K.; Stephanopoulos, G. The future of metabolic engineering and synthetic biology: Towards a systematic practice. Metab. Eng. 2012, 14, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Ajikumar, P.K.; Xiao, W.H.; Tyo, K.E.; Wang, Y.; Simeon, F.; Leonard, E.; Mucha, O.; Phon, T.H.; Pfeifer, B.; Stephanopoulos, G. Isoprenoid pathway optimization for taxol precursor overproduction in Escherichia coli. Science 2010, 330, 70–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Temme, K.; Hill, R.; Segall-Shapiro, T.H.; Moser, F.; Voigt, C.A. Modular control of multiple pathways using engineered orthogonal T7 polymerases. Nucleic Acids Res. 2012, 40, 8773–8781. [Google Scholar] [CrossRef] [PubMed]
- Salis, H.M.; Mirsky, E.A.; Voigt, C.A. Automated design of synthetic ribosome binding sites to control protein expression. Nat. Biotechnol. 2009, 27, 946–950. [Google Scholar] [CrossRef] [PubMed]
- Pfleger, B.F.; Pitera, D.J.; Smolke, C.D.; Keasling, J.D. Combinatorial engineering of intergenic regions in operons tunes expression of multiple genes. Nat. Biotechnol. 2006, 24, 1027–1032. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Haurwitz, R.E.; Shao, W.; Doudna, J.A.; Arkin, A.P. RNA processing enables predictable programming of gene expression. Nat. Biotechnol. 2012, 30, 1002–1006. [Google Scholar] [CrossRef] [PubMed]
- Dueber, J.E.; Wu, G.C.; Malmirchegini, G.R.; Moon, T.S.; Petzold, C.J.; Ullal, A.V.; Prather, K.L.; Keasling, J.D. Synthetic protein scaffolds provide modular control over metabolic flux. Nat. Biotechnol. 2009, 27, 753–759. [Google Scholar] [CrossRef] [PubMed]
- Parsons, J.B.; Dinesh, S.D.; Deery, E.; Leech, H.K.; Brindley, A.A.; Heldt, D.; Frank, S.; Smales, C.M.; Lunsdorf, H.; Rambach, A.; et al. Biochemical and structural insights into bacterial organelle form and biogenesis. J. Biol. Chem. 2008, 283, 14366–14375. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.J.; Pitera, D.J.; Withers, S.T.; Newman, J.D.; Keasling, J.D. Engineering a mevalonate pathway in Escherichia coli for production of terpenoids. Nat. Biotechnol. 2003, 21, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Carothers, J.M.; Keasling, J.D. Design of a dynamic sensor-regulator system for production of chemicals and fuels derived from fatty acids. Nat. Biotechnol. 2012, 30, 354–359. [Google Scholar] [CrossRef] [PubMed]
- Farmer, W.R.; Liao, J.C. Improving lycopene production in Escherichia coli by engineering metabolic control. Nat. Biotechnol. 2000, 18, 533–537. [Google Scholar] [CrossRef] [PubMed]
- Farmer, W.R.; Liao, J.C. Precursor balancing for metabolic engineering of lycopene production in Escherichia coli. Biotechnol. Prog. 2001, 17, 57–61. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Nambou, K.; Wei, L.; Cao, J.; Imanaka, T.; Hua, Q. Lycopene production in recombinant strains of Escherichia coli is improved by knockout of the central carbon metabolism gene coding for glucose-6-phosphate dehydrogenase. Biotechnol. Lett. 2013, 35, 2137–2145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, X.; Zou, R.; Zhou, K.; Stephanopoulos, G.; Too, H.P. Combining genotype improvement and statistical media optimization for isoprenoid production in E. coli. PLoS One 2013, 8, e75164. [Google Scholar] [CrossRef]
- Suh, W. High isoprenoid flux Escherichia coli as a host for carotenoids production. Methods Mol. Boil. 2012, 834, 49–62. [Google Scholar]
- Reyes, L.H.; Gomez, J.M.; Kao, K.C. Improving carotenoids production in yeast via adaptive laboratory evolution. Metab. Eng. 2014, 21, 26–33. [Google Scholar] [CrossRef] [PubMed]
- Ozaydin, B.; Burd, H.; Lee, T.S.; Keasling, J.D. Carotenoid-Based phenotypic screen of the yeast deletion collection reveals new genes with roles in isoprenoid production. Metab. Eng. 2013, 15, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Alper, H.; Miyaoku, K.; Stephanopoulos, G. Construction of lycopene-overproducing E. coli strains by combining systematic and combinatorial gene knockout targets. Nat. Biotechnol. 2005, 23, 612–616. [Google Scholar]
- Alper, H.; Jin, Y.S.; Moxley, J.F.; Stephanopoulos, G. Identifying gene targets for the metabolic engineering of lycopene biosynthesis in Escherichia coli. Metab. Eng. 2005, 7, 155–164. [Google Scholar] [CrossRef] [PubMed]
- Choi, H.S.; Lee, S.Y.; Kim, T.Y.; Woo, H.M. In silico identification of gene amplification targets for improvement of lycopene production. Appl. Environ. Microbiol. 2010, 76, 3097–3105. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.J.; Lee, Y.M.; Yoon, S.H.; Kim, J.H.; Ock, S.W.; Jung, K.H.; Shin, Y.C.; Keasling, J.D.; Kim, S.W. Identification of genes affecting lycopene accumulation in Escherichia coli using a shot-gun method. Biotechnol. Bioeng. 2005, 91, 636–642. [Google Scholar] [CrossRef] [PubMed]
- Alper, H.; Stephanopoulos, G. Uncovering the gene knockout landscape for improved lycopene production in E. coli. Appl. Microbiol. Biotechnol. 2008, 78, 801–810. [Google Scholar] [CrossRef]
- Jin, Y.S.; Stephanopoulos, G. Multi-dimensional gene target search for improving lycopene biosynthesis in Escherichia coli. Metab. Eng. 2007, 9, 337–347. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Li, Q.; Sun, T.; Zhu, X.; Xu, H.; Tang, J.; Zhang, X.; Ma, Y. Engineering central metabolic modules of Escherichia coli for improving β-carotene production. Metab. Eng. 2013, 17, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.H.; Isaacs, F.J.; Carr, P.A.; Sun, Z.Z.; Xu, G.; Forest, C.R.; Church, G.M. Programming cells by multiplex genome engineering and accelerated evolution. Nature 2009, 460, 894–898. [Google Scholar] [CrossRef] [PubMed]
- Warner, J.R.; Reeder, P.J.; Karimpour-Fard, A.; Woodruff, L.B.; Gill, R.T. Rapid profiling of a microbial genome using mixtures of barcoded oligonucleotides. Nat. Biotechnol. 2010, 28, 856–862. [Google Scholar] [CrossRef] [PubMed]
- Kachroo, A.H.; Jayaram, M.; Rowley, P.A. Metabolic engineering without plasmids. Nat. Biotechnol. 2009, 27, 729–731. [Google Scholar] [CrossRef] [PubMed]
- Chiang, C.J.; Chen, P.T.; Chao, Y.P. Replicon-free and markerless methods for genomic insertion of dnas in phage attachment sites and controlled expression of chromosomal genes in Escherichia coli. Biotechnol. Bioeng. 2008, 101, 985–995. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.Y.; Shen, H.J.; Cui, Y.Y.; Chen, S.G.; Weng, Z.M.; Zhao, M.; Liu, J.Z. Chromosomal evolution of Escherichia coli for the efficient production of lycopene. BMC Biotechnol. 2013, 13, 6. [Google Scholar] [CrossRef] [PubMed]
- Shin, J.H.; Kim, H.U.; Kim, D.I.; Lee, S.Y. Production of bulk chemicals via novel metabolic pathways in microorganisms. Biotechnol. Adv. 2013, 31, 925–935. [Google Scholar] [CrossRef] [PubMed]
- Zanghellini, A. De novo computational enzyme design. Curr. Opin. Biotechnol. 2014, 29C, 132–138. [Google Scholar] [CrossRef] [PubMed]
- Damborsky, J.; Brezovsky, J. Computational tools for designing and engineering enzymes. Curr. Opin. Chem. Biol. 2014, 19C, 8–16. [Google Scholar] [CrossRef]
- Tao, L.; Wilczek, J.; Odom, J.M.; Cheng, Q. Engineering a β-carotene ketolase for astaxanthin production. Metab. Eng. 2006, 8, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.W.; Stead, K.J.; Yao, H.; He, H. Mutational and functional analysis of the β-carotene ketolase involved in the production of canthaxanthin and astaxanthin. Appl. Environ. Microbiol. 2006, 72, 5829–5837. [Google Scholar] [CrossRef] [PubMed]
- Umeno, D.; Tobias, A.V.; Arnold, F.H. Evolution of the C30 carotenoid synthase CrtM for function in a C40 pathway. J. Bacteriol. 2002, 184, 6690–6699. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Dannert, C.; Umeno, D.; Arnold, F.H. Molecular breeding of carotenoid biosynthetic pathways. Nat. Biotechnol. 2000, 18, 750–753. [Google Scholar]
- Fuxreiter, M.; Mones, L. The role of reorganization energy in rational enzyme design. Curr. Opin. Chem. Biol. 2014, 21C, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Leonard, E.; Ajikumar, P.K.; Thayer, K.; Xiao, W.H.; Mo, J.D.; Tidor, B.; Stephanopoulos, G.; Prather, K.L. Combining metabolic and protein engineering of a terpenoid biosynthetic pathway for overproduction and selectivity control. Proc. Natl. Acad. Sci. USA 2010, 107, 13654–13659. [Google Scholar] [CrossRef] [PubMed]
- Furubayashi, M.; Ikezumi, M.; Kajiwara, J.; Iwasaki, M.; Fujii, A.; Li, L.; Saito, K.; Umeno, D. A high-throughput colorimetric screening assay for terpene synthase activity based on substrate consumption. PLoS One 2014, 9, e93317. [Google Scholar] [CrossRef] [PubMed]
- Del Campo, J.A.; Garcia-Gonzalez, M.; Guerrero, M.G. Outdoor cultivation of microalgae for carotenoid production: Current state and perspectives. Appl. Microbiol. Biotechnol. 2007, 74, 1163–1174. [Google Scholar]
- Rosenberg, J.N.; Oyler, G.A.; Wilkinson, L.; Betenbaugh, M.J. A green light for engineered algae: Redirecting metabolism to fuel a biotechnology revolution. Curr. Opin. Biotechnol. 2008, 19, 430–436. [Google Scholar] [CrossRef] [PubMed]
- Binti Ibnu Rasid, E.N.; Mohamad, S.E.; Jamaluddin, H.; Salleh, M.M. Screening factors influencing the production of astaxanthin from freshwater and marine microalgae. Appl. Biochem. Biotechnol. 2014, 172, 2160–2174. [Google Scholar]
- Casal, C.; Cuaresma, M.; Vega, J.M.; Vilchez, C. Enhanced productivity of a lutein-enriched novel acidophile microalga grown on urea. Mar. Drugs 2011, 9, 29–42. [Google Scholar] [CrossRef]
- Fu, W.; Paglia, G.; Magnusdottir, M.; Steinarsdottir, E.A.; Gudmundsson, S.; Palsson, B.O.; Andresson, O.S.; Brynjolfsson, S. Effects of abiotic stressors on lutein production in the green microalga Dunaliella salina. Microb. Cell Factories 2014, 13, 3. [Google Scholar] [CrossRef]
- Fu, W.; Guethmundsson, O.; Paglia, G.; Herjolfsson, G.; Andresson, O.S.; Palsson, B.O.; Brynjolfsson, S. Enhancement of carotenoid biosynthesis in the green microalga Dunaliella salina with light-emitting diodes and adaptive laboratory evolution. Appl. Microbiol. Biotechnol. 2013, 97, 2395–2403. [Google Scholar] [CrossRef] [PubMed]
- Leon, R.; Couso, I.; Fernandez, E. Metabolic engineering of ketocarotenoids biosynthesis in the unicelullar microalga Chlamydomonas reinhardtii. J. Biotechnol. 2007, 130, 143–152. [Google Scholar] [CrossRef] [PubMed]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Wang, C.; Kim, J.-H.; Kim, S.-W. Synthetic Biology and Metabolic Engineering for Marine Carotenoids: New Opportunities and Future Prospects. Mar. Drugs 2014, 12, 4810-4832. https://doi.org/10.3390/md12094810
Wang C, Kim J-H, Kim S-W. Synthetic Biology and Metabolic Engineering for Marine Carotenoids: New Opportunities and Future Prospects. Marine Drugs. 2014; 12(9):4810-4832. https://doi.org/10.3390/md12094810
Chicago/Turabian StyleWang, Chonglong, Jung-Hun Kim, and Seon-Won Kim. 2014. "Synthetic Biology and Metabolic Engineering for Marine Carotenoids: New Opportunities and Future Prospects" Marine Drugs 12, no. 9: 4810-4832. https://doi.org/10.3390/md12094810