Isolation and Identification of Antitrypanosomal and Antimycobacterial Active Steroids from the Sponge Haliclona simulans

 and
and 
Abstract
:
1. Introduction
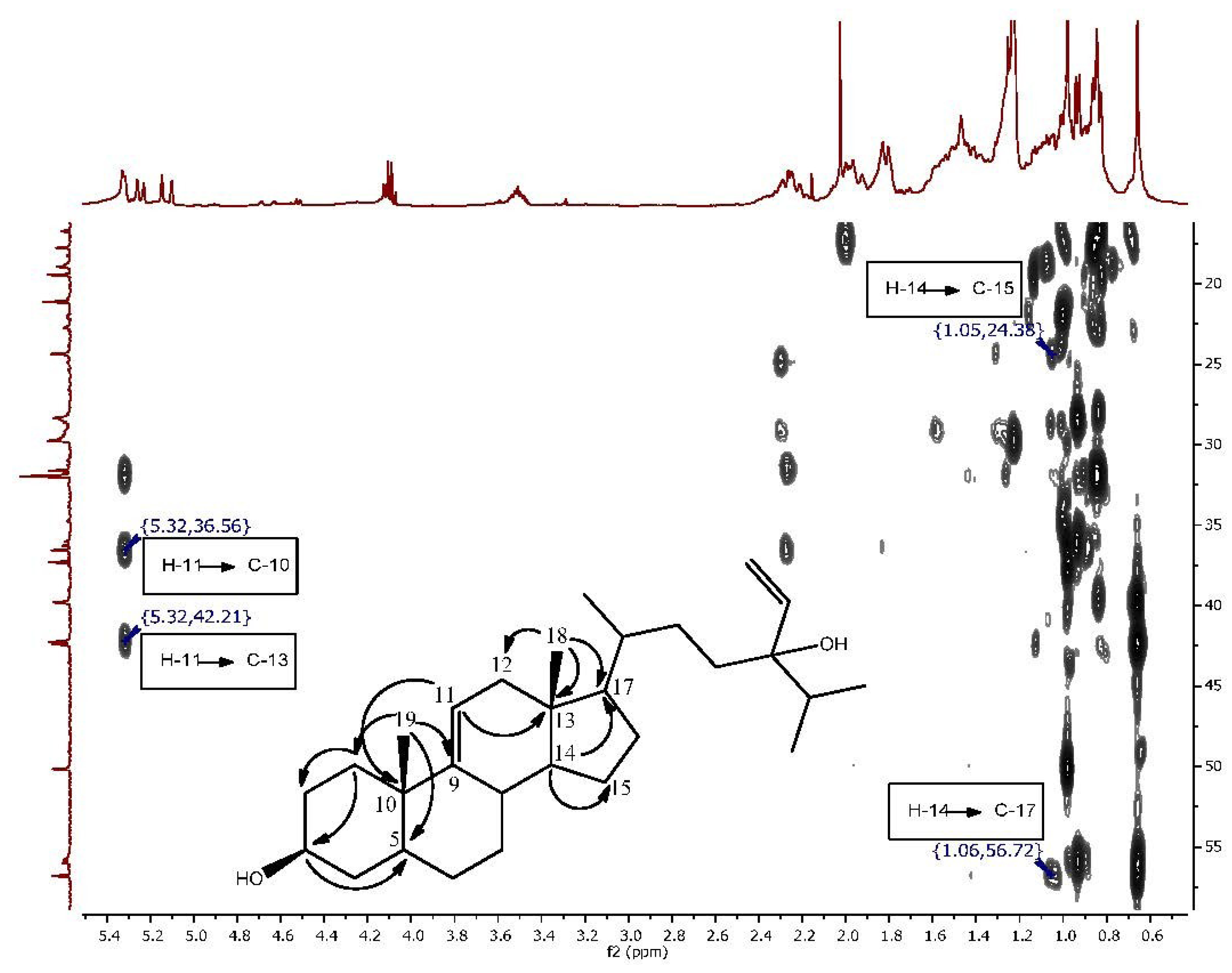
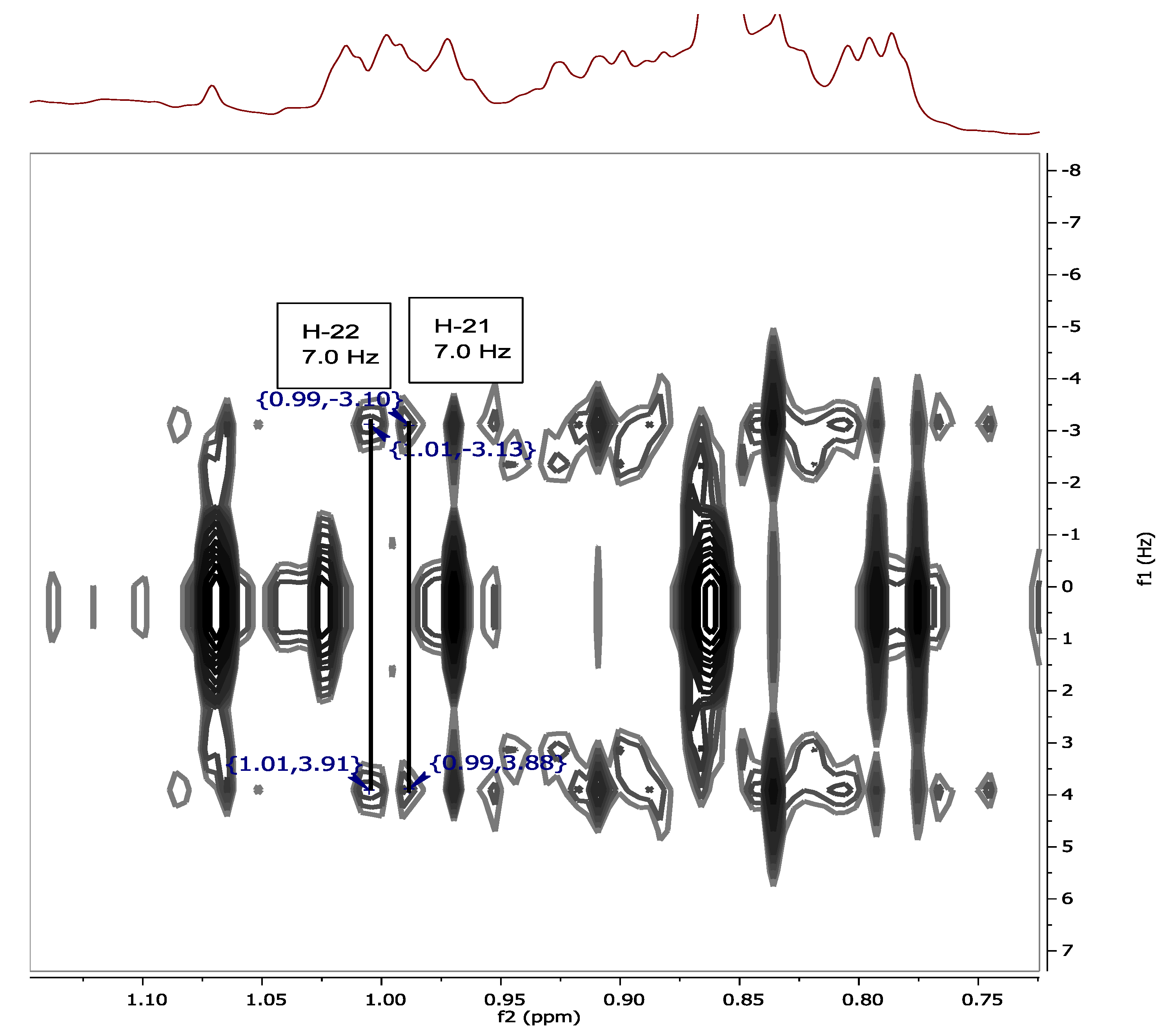
2. Results and Discussion
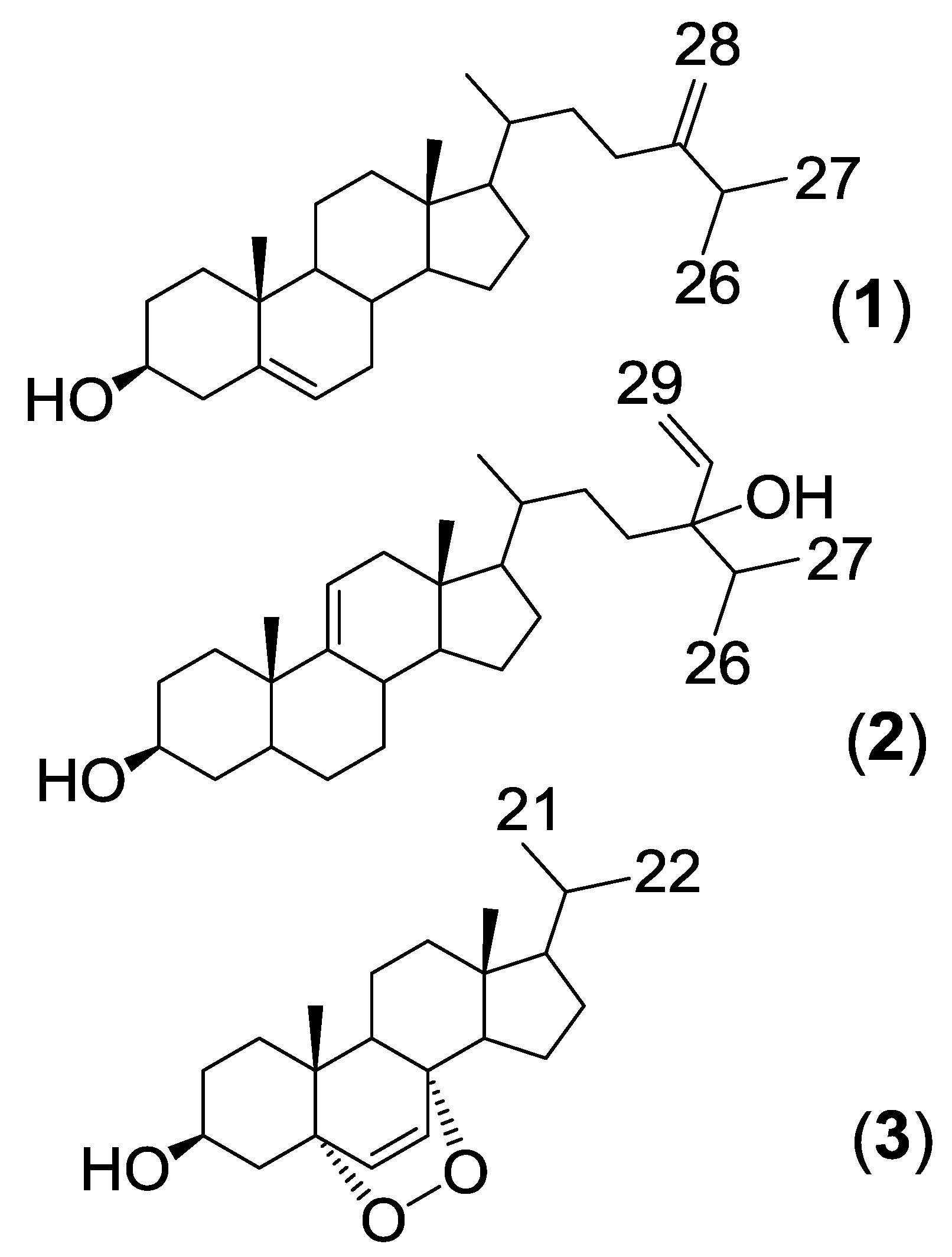

{kind=link}
{kind=link}
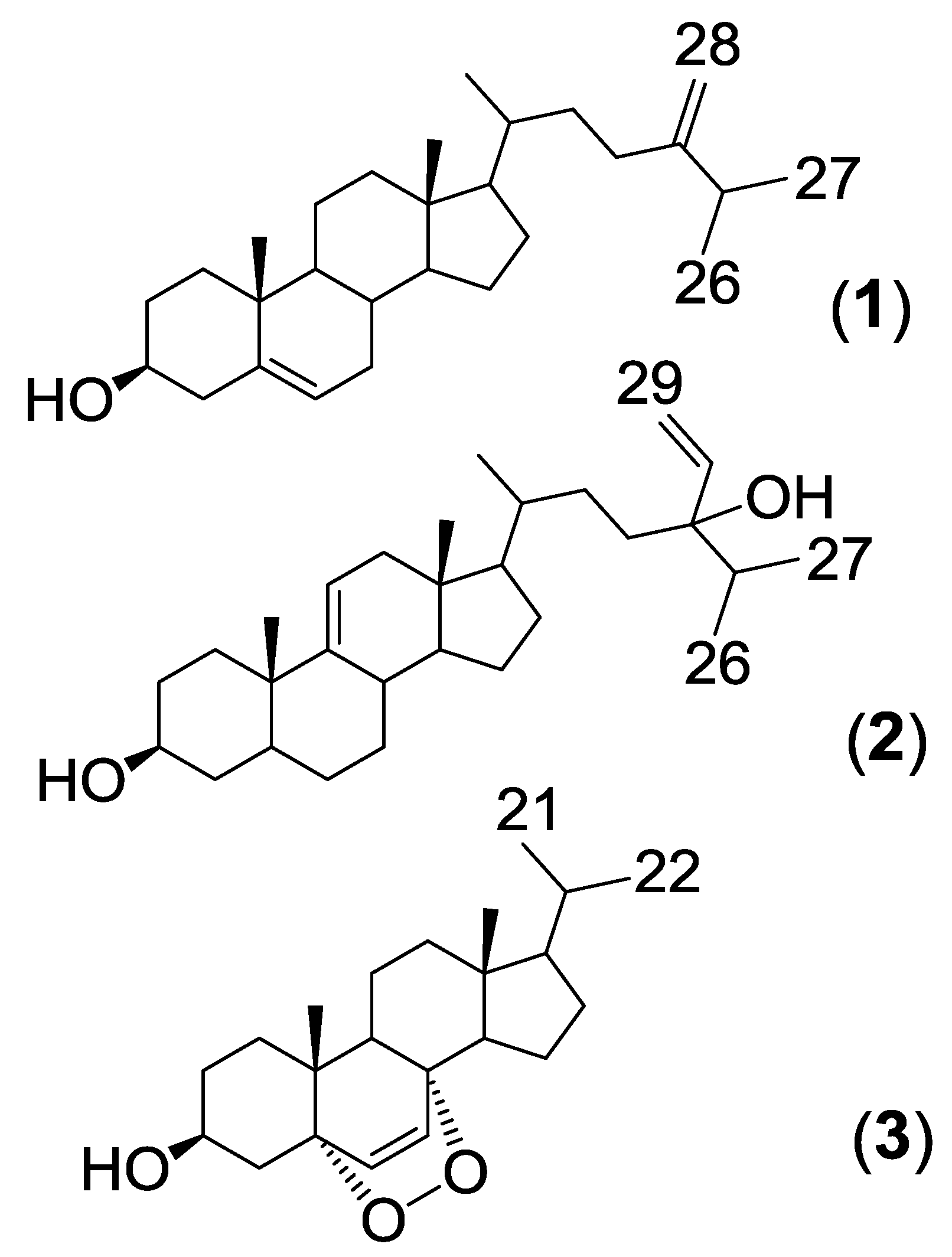{kind=link}
{kind=link}
{kind=link}
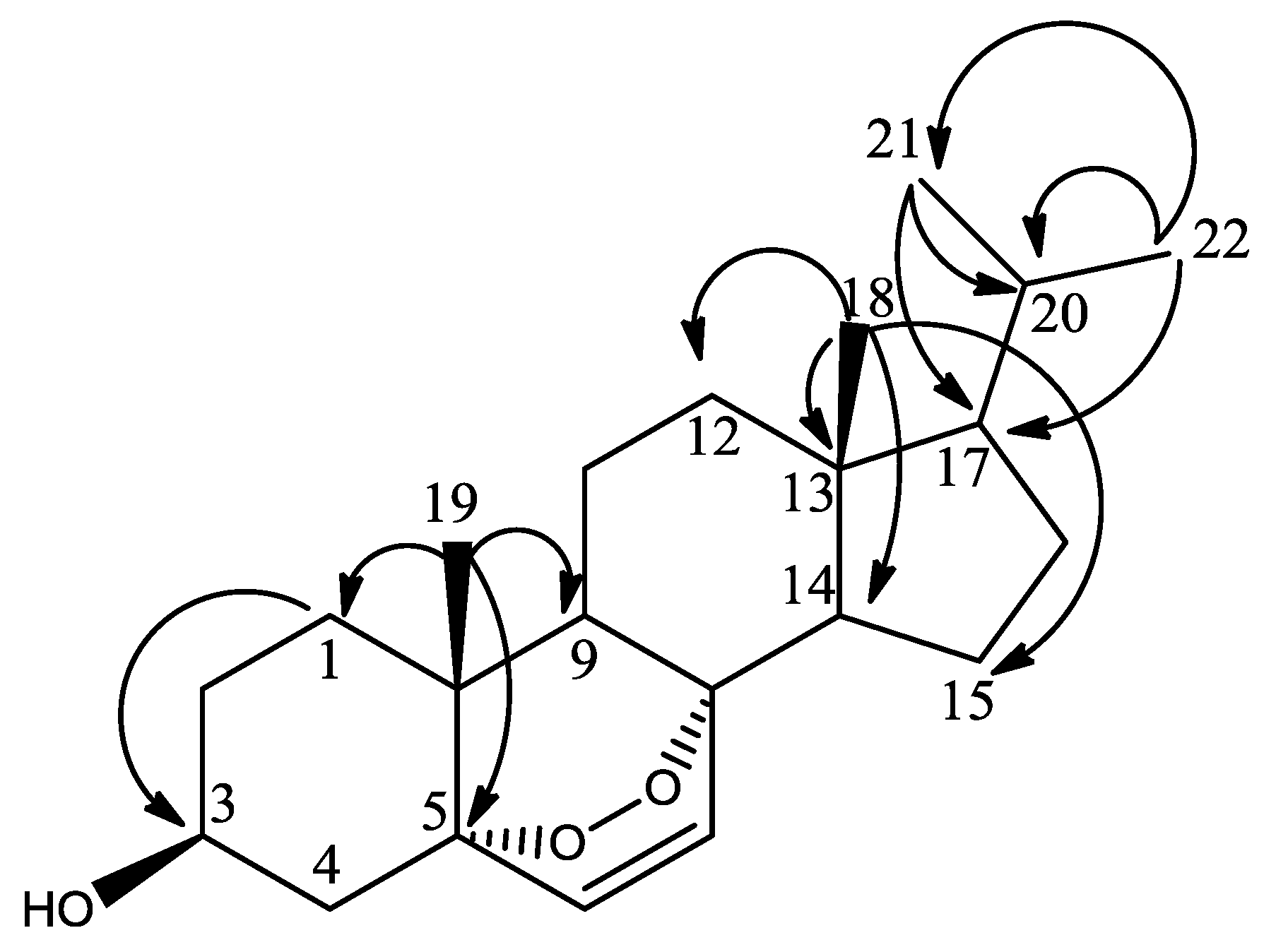{kind=link}
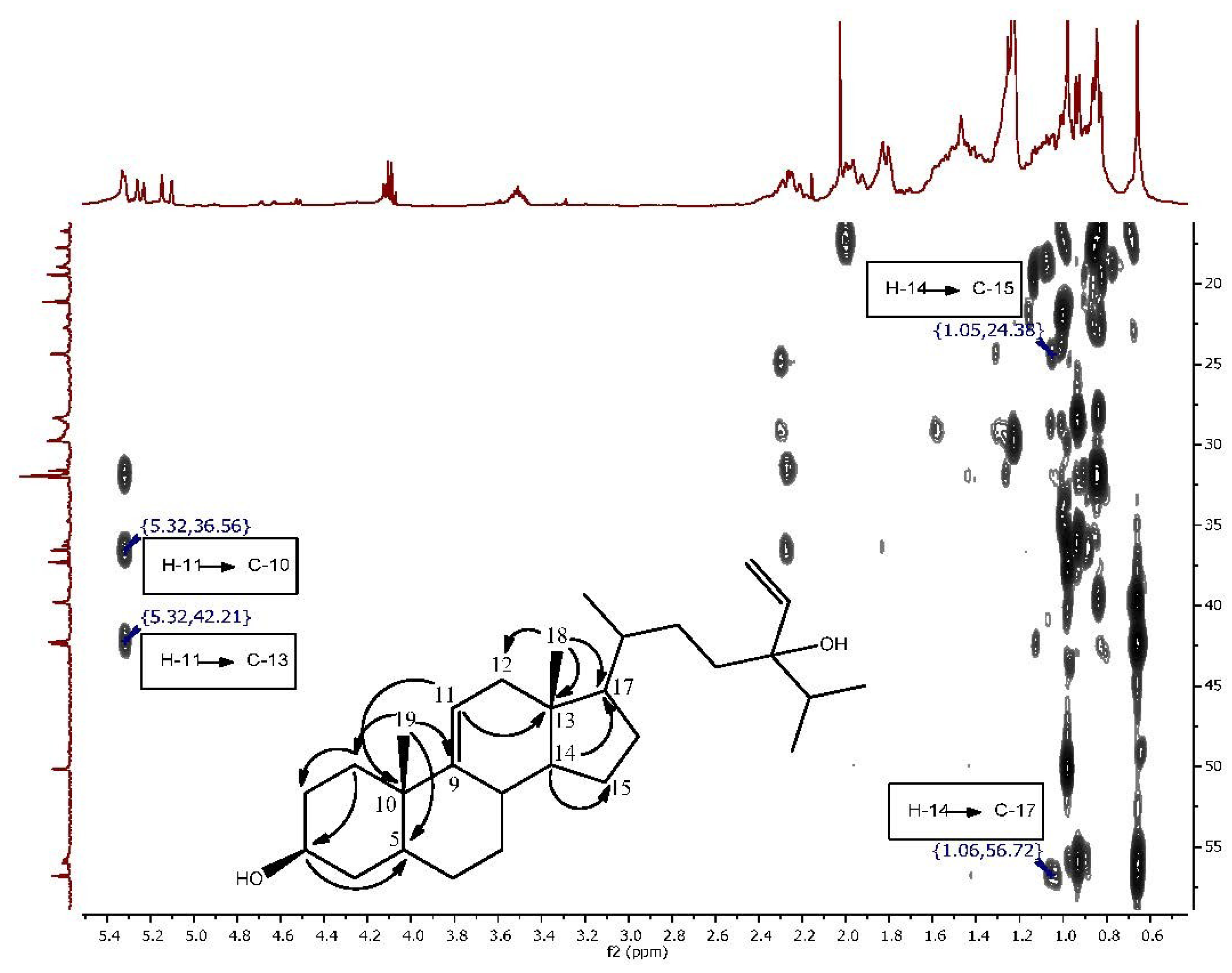
{kind=link}
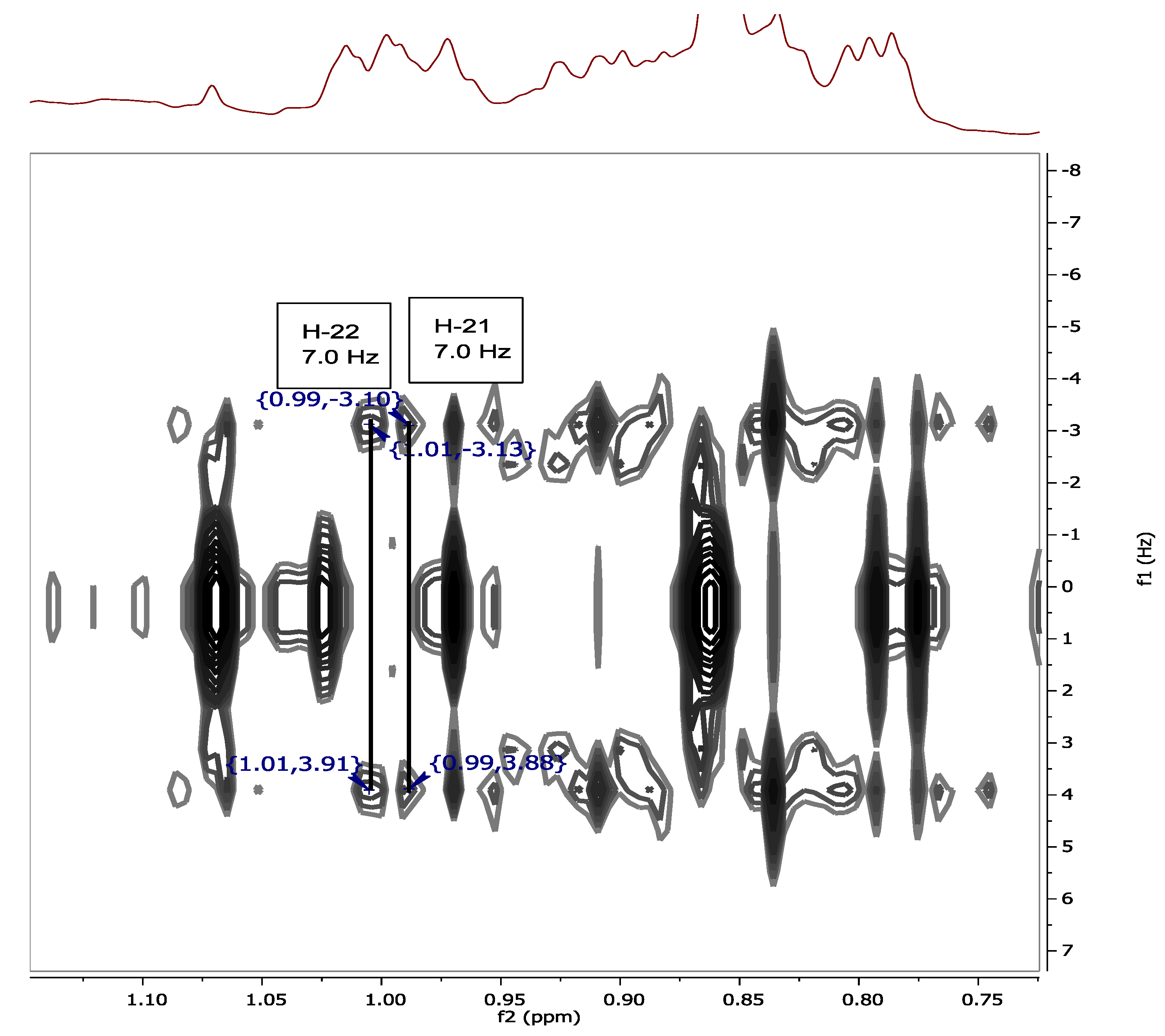
{kind=link}
| Position | 1 | 2 | 3 |
|---|---|---|---|
| 1 | 1.81 (d, 5.1) | 1.82 (d, 11.5) | 1.68, 1.94 (d, 4.6) |
| 2 | 1.50 | 1.45 (d, 12.9) | 1.52, 1.83 |
| 3 | 3.52 (td, 5.3, 11.0) | 3.51 (tt, 4.7, 10.6) | 3.96 (tt, 5.1, 11.1) |
| 4 | 2.27 (m) | 2.24 (m) | 1.90 (s), 2.12 (d, 4.8) |
| 6 | 5.34 (d, 5.7) | 0.90 (d, 4.0) | 6.23 (d, 8.5) |
| 7 | 1.95 | 6.49 (d, 8.5) | |
| 9 | 1.50 (m) | ||
| 11 | 5.32 (d, 5.0) | 1.20, 1.49 | |
| 12 | 1.14, 1.94 | 1.21, 1.97 | |
| 14 | 1.06 | 1.57, (d, 3.6) | |
| 15 | 0.99 | ||
| 16 | 1.24 (s) | ||
| 17 | 1.20 | ||
| 18 | 0.67 (t, 3.1) | 0.66 (s) | 0.80 (s) |
| 19 | 0.99 (s) | 0.98 (s) | 0.87 (s) |
| 20 | 1.41 (d, 3.9) | 1.98 (m) | |
| 21 | 0.93 (d, 6.7) | 0.93 (d, 6.3) | 0.98 (d, 7.0) |
| 22 | 1.85 | 1.02 (d, 7.0) | |
| 23 | 1.11, 1.99 | ||
| 25 | 1.99 | ||
| 26 | 1.02 (d, 2.3) | 0.98 | |
| 27 | 1.02 (d, 2.3) | 0.87 | |
| 28 | 4.64 (s), 4.70 (s) | 5.72 (m) | |
| 29 | 5.12 (d, 17.9) 5.25 (d, 11.4) |
| Position | 1 | 2 | 3 |
|---|---|---|---|
| 1 | 37.3 (CH2) | 37.3 (CH2) | 34.8 (CH2) |
| 2 | 31.7 (CH2) | 31.6 (CH2) | 30.2 (CH2) |
| 3 | 71.9 (CH) | 71.9 (CH) | 66.6 (CH) |
| 4 | 42.3 (CH2) | 42.3 (CH2) | 37.0 (CH2) |
| 5 | 140.8 (C) | 50.2 (CH) | 82.2 (C) |
| 6 | 121.8 (CH) | 34.8 (CH2) | 135.50 (CH) |
| 7 | 31.0 (CH2) | 22.0 (CH2) | 130.8 (CH) |
| 8 | 32.0 (CH) | 21.1 (CH) | 79.5 (C) |
| 9 | 50.2 (CH) | 140.8 (C) | 51.1 (CH) |
| 10 | 36.6 (C) | 36.6 (C) | 36.0 (C) |
| 11 | 21.2 (CH2) | 121.8 (CH) | 23.5 (CH2) |
| 12 | 39.8 (CH2) | 39.8 (CH2) | 39.5 (CH2) |
| 13 | 42.4 (C) | 42.4 (C) | 44.8 (C) |
| 14 | 56.8 (CH) | 56.0 (CH) | 51.7 (CH) |
| 15 | 24.4 (CH2) | 24.4 (CH2) | 20.7 (CH2) |
| 16 | 28.3 (CH2) | 29.8 (CH2) | 29.8 (CH2) |
| 17 | 56.1 (CH) | 56.8 (CH) | 56.2 (CH) |
| 18 | 11.9 (CH3) | 11.9 (CH3) | 12.7 (CH3) |
| 19 | 19.5 (CH3) | 19.5 (CH3) | 18.3 (CH3) |
| 20 | 35.9 (CH2) | 36.3 (CH) | 33.9 (CH) |
| 21 | 18.8 (CH2) | 19.0(CH3) | 22.1 (CH3) |
| 22 | 23.9 (CH2) | 28.3 (CH2) | 28.4 (CH2) |
| 23 | 34.8 (CH2) | 28.5 (CH2) | 28.1 (CH3) |
| 24 | 157.0 (C) | 89.2 (C) | |
| 25 | 33.9 (CH) | 32.0 (CH) | |
| 26 | 22.1 (CH3) | 17.8 (CH3) | |
| 27 | 28.1 (CH3) | 16.7 (CH3) | |
| 28 | 106.0 (CH2) | 137.2 (CH) | |
| 29 | 116.3 (CH2) |
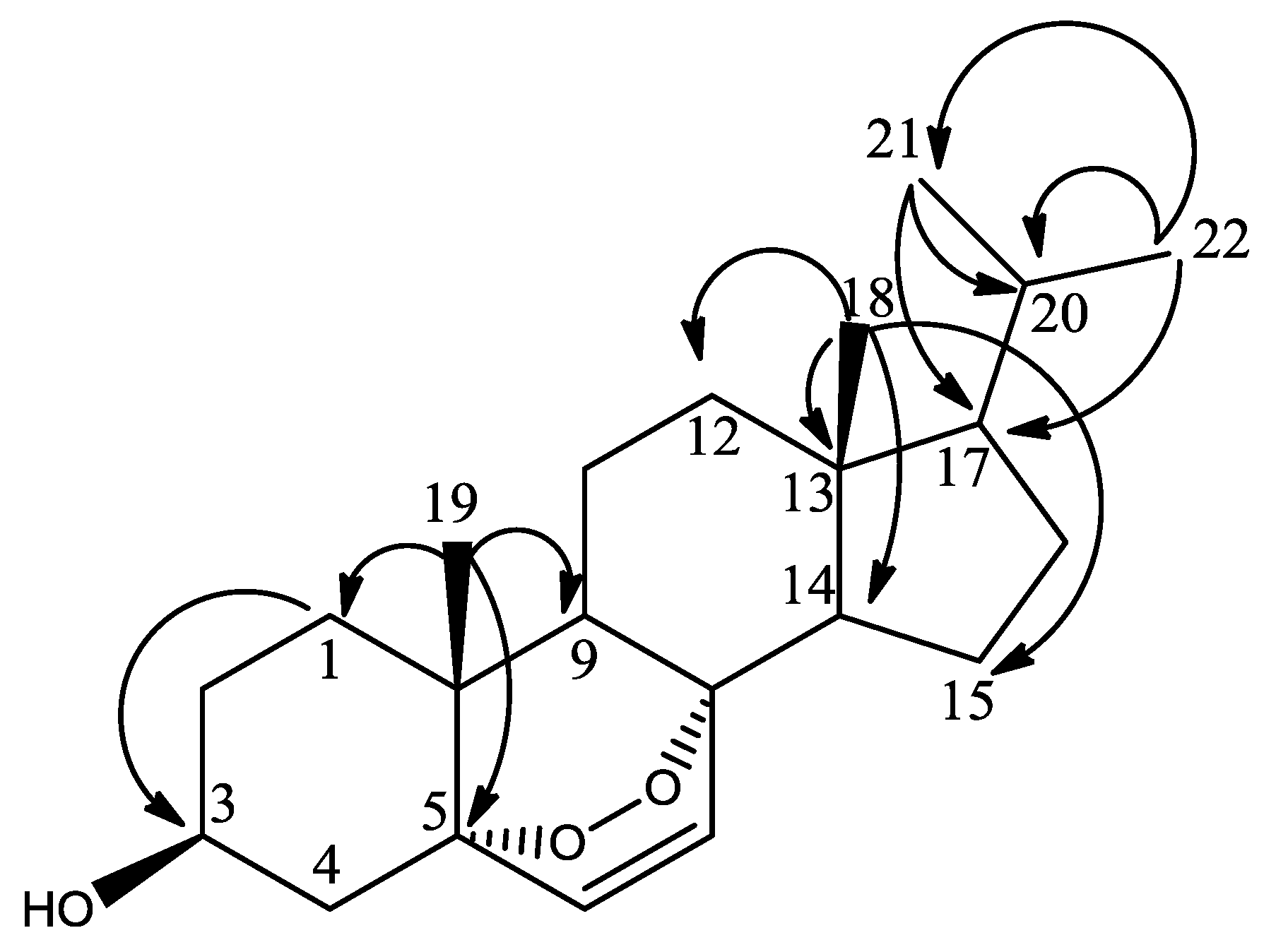
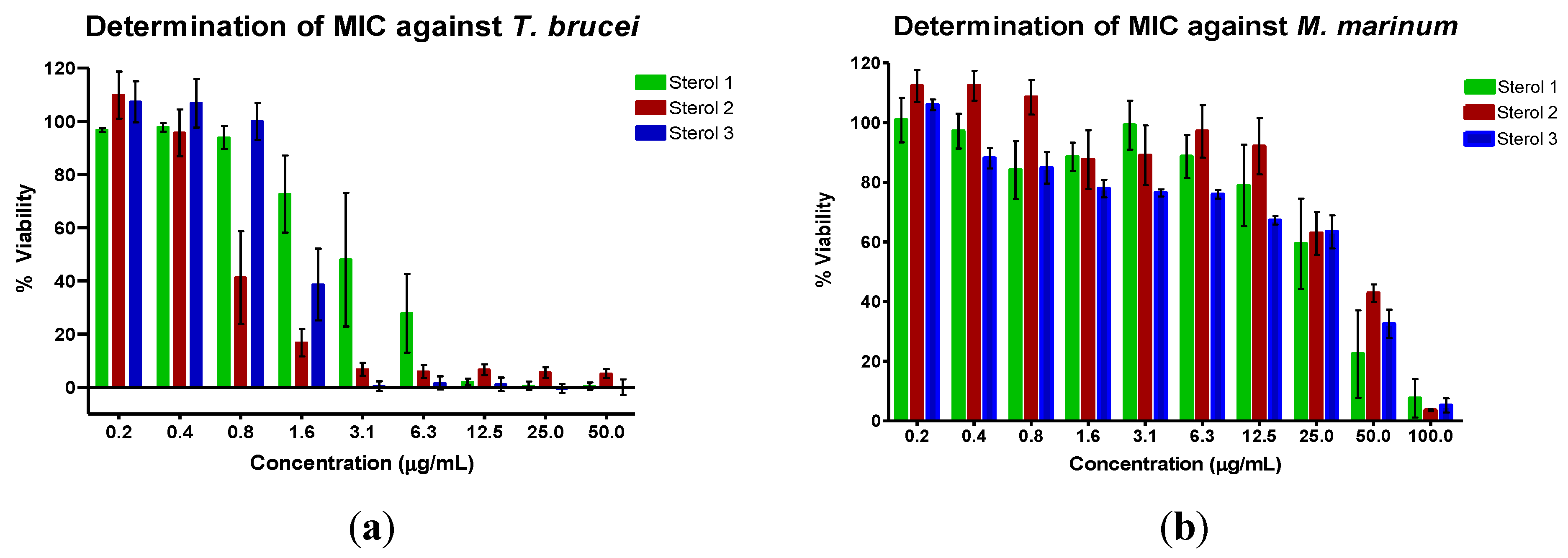
| Sterol | MIC Average ± Std Dev (n = 4) | Cytotoxicity on HS27 Cells | |
|---|---|---|---|
| T. b. brucei (μM) | M. marinum (μM) | IC50 (μM) ± Std Dev (n = 3) | |
| 1 | 21.56 ± 11.80 | 156.90 ± 54.35 | 58 ± 3.53 |
| 2 | 4.58 ± 1.80 | 233.44 ± 0 | >100 ± 4.03 |
| 3 | 9.01 ± 0 | 288.81 ± 0 | 100 ± 2.95 |
| Suramin | 0.11 ± 0 | ||
| Gentamycin | 13.48 ± 0 | ||
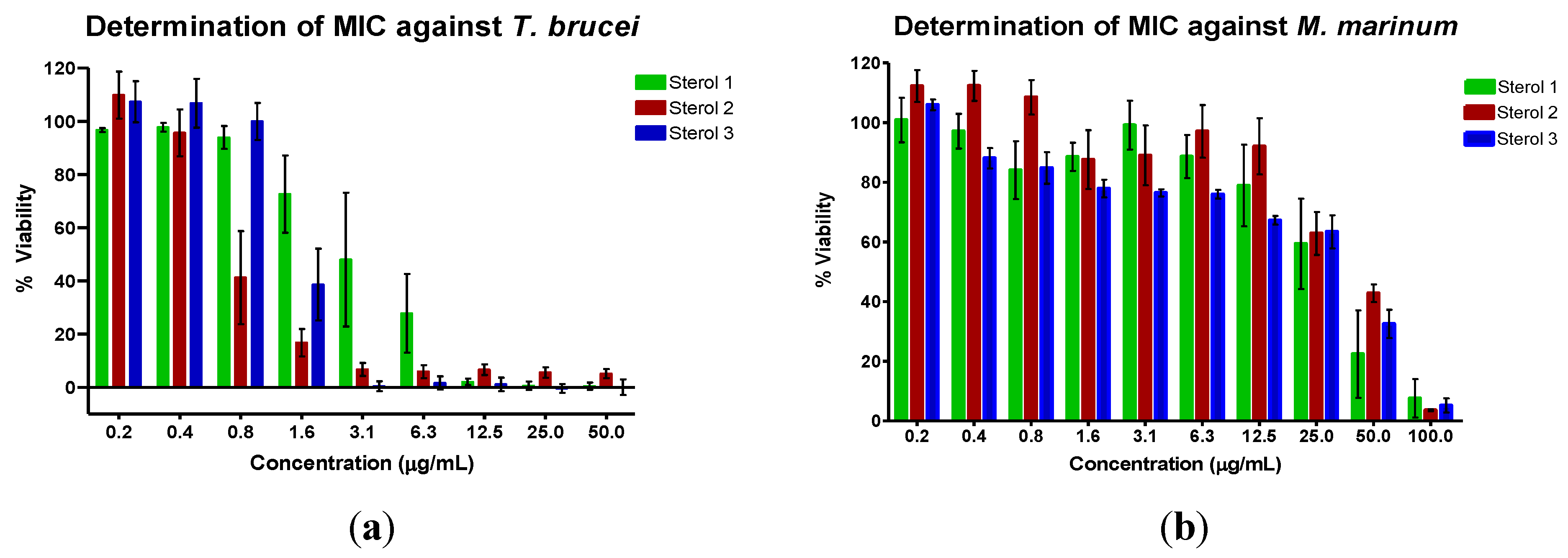

3. Experimental Section
3.1. Acquisition of Sponge Sample
3.2. Extraction and Isolation of Sponge Metabolites
3.3. Analysis of Sterols
3.4. Antitrypanosomal Assay
3.5. Anti-Mycobacterium Assay
3.6. Cell Cytotoxicity Assay
4. Conclusions
Supplementary Files
Acknowledgments
Author Contributions
Conflicts of Interest
References
- WHO. The Global Burden of Disease 2004 Update; World Health Organization: Geneva, Switzerland, 2008. [Google Scholar]
- WHO. Sustaining the Drive to Overcome the Global Impact of Neglected Tropical Diseases: Second Who Report on Neglected Tropical Diseases; World Health Organization: Geneva, Switzerland, 2013. [Google Scholar]
- Legros, D.; Ollivier, G.; Gastellu-Etchegorry, M.; Paquet, C.; Burri, C.; Jannin, J.; Büscher, P. Treatment of human African trypanosomiasis—Present situation and needs for research and development. Lancet Infect. Dis. 2002, 2, 437–440. [Google Scholar] [CrossRef]
- Bern, C.; Montgomery, S.P.; Herwaldt, B.L.; Rassi, A.; Marin-Neto, J.A.; Dantas, R.O.; Maguire, J.H.; Acquatella, H.; Morillo, C.; Kirchhoff, L.V.; et al. Evaluation and treatment of chagas disease in the united states: A systematic review. JAMA 2007, 298, 2171–2181. [Google Scholar] [CrossRef]
- Tiemersma, E.; van der Werf, M.; Borgdorff, M.; Williams, B.; Nagelkerke, N. Natural history of tuberculosis: Duration and fatality of untreated pulmonary tuberculosis in HIV negative patients: A systematic review. PLoS One 2011, 6, e17601. [Google Scholar]
- WHO. Global Tuberculosis Report 2012; World Health Organization: Geneva, Switzerland, 2012. [Google Scholar]
- Cordeiro, A.T.; Thiemann, O.H.; Michels, P.A. Inhibition of Trypanosoma brucei glucose-6-phosphate dehydrogenase by human steroids and their effects on the viability of cultured parasites. Bioorg. Med. Chem. 2009, 17, 2483–2489. [Google Scholar] [CrossRef]
- Gupta, S.; Cordeiro, A.T.; Michels, P.A. Glucose-6-phosphate dehydrogenase is the target for the trypanocidal action of human steroids. Mol. Biochem. Parasitol. 2011, 176, 112–115. [Google Scholar] [CrossRef]
- Magaraci, F.; Jimenez, C.J.; Rodrigues, C.; Rodrigues, J.C.; Braga, M.V.; Yardley, V.; de Luca-Fradley, K.; Croft, S.L.; de Souza, W.; Ruiz-Perez, L.M.; et al. Azasterols as inhibitors of sterol 24-methyltransferase in Leishmania species and Trypanosoma cruzi. J. Med. Chem. 2003, 46, 4714–4727. [Google Scholar] [CrossRef]
- Gros, L.; Castillo-Acosta, V.M.; Jiménez Jiménez, C.; Sealey-Cardona, M.; Vargas, S.; Manuel Estévez, A.; Yardley, V.; Rattray, L.; Croft, S.L.; Ruiz-Perez, L.M.; et al. New azasterols against Trypanosoma brucei: Role of 24-sterol methyltransferase in inhibitor action. Antimicrob. Agents Chemother. 2006, 50, 2595–2601. [Google Scholar] [CrossRef]
- Lepesheva, G.I.; Ott, R.D.; Hargrove, T.Y.; Kleshchenko, Y.Y.; Schuster, I.; Nes, W.D.; Hill, G.C.; Villalta, F.; Waterman, M.R. Sterol 14alpha-demethylase as a potential target for antitrypanosomal therapy: Enzyme inhibition and parasite cell growth. Chem. Biol. 2007, 14, 1283–1293. [Google Scholar] [CrossRef]
- Lamb, D.; Kelly, D.; Manning, N.; Kelly, S. A sterol biosynthetic pathway in Mycobacterium. FEBS Lett. 1998, 437, 142–144. [Google Scholar] [CrossRef]
- Bellamine, A.; Mangla, A.; Dennis, A.; Nes, W.; Waterman, M. Structural requirements for substrate recognition of Mycobacterium tuberculosis 14 alpha-demethylase: Implications for sterol biosynthesis. J. Lipid Res. 2001, 42, 128–136. [Google Scholar]
- Taylor, M.; Radax, R.; Steger, D.; Wagner, M. Sponge-associated microorganisms: Evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. Rev. 2007, 71, 295–347. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Keyzers, R.A.; Munro, M.H.; Prinsep, M.R. Marine natural products. Nat. Prod. Microbiol. Rep. 2013, 30, 237–323. [Google Scholar] [CrossRef]
- Blunt, J.W.; Copp, B.R.; Munro, M.H.; Northcote, P.T.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2004, 21, 1–49. [Google Scholar] [CrossRef]
- Sheikh, Y.; Djerassi, C. Steroids from sponges. Tetrahedron 1974, 30, 4095–4103. [Google Scholar] [CrossRef]
- Bergmann, W.; Feeney, R. The isolation of a new thymine pentoside from sponges. J. Am. Chem. Soc. 1950, 72, 2809–2810. [Google Scholar] [CrossRef]
- Bergmann, W.; Feeney, R. Contributions to the study of marine products. 32. The nucleosides of sponges. 1. J. Org. Chem. 1951, 16, 981–987. [Google Scholar] [CrossRef]
- Bergmann, W.; Burke, D. Contributions to the study of marine products. 39. The nucleosides of sponges. 3. Spongothymidine and spongouridine. J. Org. Chem. 1955, 20, 1501–1507. [Google Scholar] [CrossRef]
- Newman, D.; Cragg, G. Marine natural products and related compounds in clinical and advanced preclinical trials. J. Nat. Prod. 2004, 67, 1216–1238. [Google Scholar] [CrossRef]
- Kennedy, J.; Baker, P.; Piper, C.; Cotter, P.; Walsh, M.; Mooij, M.; Bourke, M.; Rea, M.; O’Connor, P.; Ross, R.; et al. Isolation and analysis of bacteria with antimicrobial activities from the marine sponge Haliclona simulans collected from irish waters. Mar. Biotechnol. 2009, 11, 384–396. [Google Scholar] [CrossRef]
- Yu, S.; Deng, Z.; Proksch, P.; Lin, W. Oculatol, oculatolide, and a-nor sterols from the sponge Haliclona oculata. J. Nat. Prod. 2006, 69, 1330–1334. [Google Scholar] [CrossRef]
- Bergmann, W.; Schedl, H.; Low, E. Contributions to the study of marine products. 19. Chalinasterol. J. Org. Chem. 1945, 10, 587–593. [Google Scholar] [CrossRef]
- Giner, J. Biosynthesis of marine sterol side chains. Chem. Rev. 1993, 93, 1735–1752. [Google Scholar] [CrossRef]
- Bazin, M.A.; Loiseau, P.M.; Bories, C.; Letourneux, Y.; Rault, S.; El Kihel, L. Synthesis of oxysterols and nitrogenous sterols with antileishmanial and trypanocidal activities. Eur. J. Med. Chem. 2006, 41, 1109–1116. [Google Scholar] [CrossRef]
- Ikekawa, N.; Tsuda, K.; Morisaki, N. Saringosterol: A new sterol from brown algae. Chem. Ind. 1966, 85, 1179–1180. [Google Scholar]
- Ayyad, S.E.; Sowellim, S.Z.; El-Hosini, M.S.; Abo-Atia, A. The structural determination of a new steroidal metabolite from the brown alga Sargassum asperifolium. Z für Naturforschung. C J. Biosci. 2003, 58C, 333–336. [Google Scholar]
- Gauvin, A.; Smadja, J.; Aknin, M.; Faure, R.; Gaydou, E. Isolation of bioactive 5 alpha,8 alpha-epidioxy sterols from the marine sponge Luffariella cf. variabilis. Can. J. Chem. 2000, 78, 986–992. [Google Scholar]
- Wei, X.; Rodríguez, A.D.; Wang, Y.; Franzblau, S.G. Synthesis and in vitro biological evaluation of ring b abeo-sterols as novel inhibitors of mycobacterium tuberculosis. Bioorg. Med. Chem. Lett. 2008, 18, 5448–5450. [Google Scholar] [CrossRef]
- Hoet, S.; Pieters, L.; Muccioli, G.G.; Habib-Jiwan, J.L.; Opperdoes, F.R.; Quetin-Leclercq, J. Antitrypanosomal activity of triterpenoids and sterols from the leaves of Strychnos spinosa and related compounds. J. Nat. Prod. 2007, 70, 1360–1363. [Google Scholar] [CrossRef]
- Kitagawa, I.; Kobayashi, M.; Sugawara, T.; Yosioka, I. Thornasterol a and b, two genuine sapogenols from the starfish Acanthaster planci. Tetrahedron Lett. 1975, 16, 967–970. [Google Scholar] [CrossRef]
- Kitagawa, I.; Kobayashi, M.; Sugawara, T. Saponin and sapogenol. 25. Steroidal saponins from the starfish Acanthaster planci l. (crown of thorns). 1. Structures of two genuine sapogenols, thornasterol a and thornasterol b, and their sulfates. Chem. Pharm. Bull. 1978, 26, 1852–1863. [Google Scholar] [CrossRef]
- ApSimon, J.; Buccini, J.; Badriper, S. Marine organic chemistry. 1. Isolation of 3beta,6alpha-dihydroxy-5alpha-pregn-9(11)-en-20-one from saponins of starfish Asterias forbesi. A rapid method for extracting starfish saponins. Can. J. Chem. 1973, 51, 850–855. [Google Scholar] [CrossRef]
- ApSimon, J.; Badripersaud, S.; Buccini, J.; Eenkhoorn, J. Marine organic chemistry. 4. Isolation of (20r)-5-alpha-pregn-9(11)-ene-3-beta,6-alpha,20-triol from the saponins of the starfish, Asterias forbesi and Asterias vulgaris, and its synthesis. Can. J. Chem. 1980, 58, 2703–2708. [Google Scholar] [CrossRef]
- Goad, L.; Garneau, F.; Simard, J.; ApSimon, J.; Girard, M. Isolation of delta-9(11)-sterols from the sea cucumber Psolus fabricii. implications for holothurin biosynthesis. Tetrahedron Lett. 1985, 26, 3513–3516. [Google Scholar] [CrossRef]
- Cordeiro, M.; Djerassi, C. Biosynthetic studies of marine lipids. 25. Biosynthesis of delta-9(11)-sterols and delta-7-sterols and saponins in sea cucumbers. J. Org. Chem. 1990, 55, 2806–2813. [Google Scholar] [CrossRef]
- Ballantine, J.; Williams, K.; Burke, B. Marine sterols. 4. C21 sterols from marine sources. Identification of pregnane derivatives in extracts of sponge Haliclona rubens. Tetrahedron Lett. 1977, 18, 1547–1550. [Google Scholar] [CrossRef]
- Wachter, G.; Franzblau, S.; Montenegro, G.; Hoffmann, J.; Maiese, W.; Timmermann, B. Inhibition of Mycobacterium tuberculosis growth by saringosterol from Lessonia nigrescens. J. Nat. Prod. 2001, 64, 1463–1464. [Google Scholar] [CrossRef]
- Elenkov, I.; Milkova, T.; Andreev, S.; Popov, S. Sterol composition and biosynthesis in the black sea sponge Dysidea fragilis. Comp. Biochem. Physiol. B 1994, 107, 547–551. [Google Scholar]
- Pinheiro, A.; Dethoup, T.; Bessa, J.; Silva, A.; Kijjoa, A. A new bicyclic sesquiterpene from the marine sponge associated fungus Emericellopsis minima. Phytochem. Lett. 2012, 5, 68–70. [Google Scholar] [CrossRef]
- Ramos-Ligonio, A.; López-Monteon, A.; Trigos, A. Trypanocidal activity of ergosterol peroxide from pleurotus ostreatus. Phytother. Res. 2012, 26, 938–943. [Google Scholar] [CrossRef]
- Truong, N.B.; Pham, C.V.; Doan, H.T.; Nguyen, H.V.; Nguyen, C.M.; Nguyen, H.T.; Zhang, H.J.; Fong, H.H.; Franzblau, S.G.; Soejarto, D.D.; et al. Antituberculosis cycloartane triterpenoids from Radermachera boniana. J. Nat. Prod. 2011, 74, 1318–1322. [Google Scholar] [CrossRef]
- Palermo, J.; Brasco, M.; Hughes, E.; Seldes, A.; Balzaretti, V.; Cabezas, E. Short side chain sterols from the tunicate Polizoa opuntia. Steroids 1996, 61, 2–6. [Google Scholar] [CrossRef]
- Carlson, R.; Popov, S.; Massey, I.; Delseth, C.; Ayanoglu, E.; Varkony, T.; Djerassi, C. Minor and trace sterols in marine invertebrates. 6. Occurrence and possible origins of sterols possessing unusually short hydrocarbon side chains. Bioorg. Chem. 1978, 7, 453–479. [Google Scholar] [CrossRef]
- Lorente, S.O.; Rodrigues, J.C.; Jiménez Jiménez, C.; Joyce-Menekse, M.; Rodrigues, C.; Croft, S.L.; Yardley, V.; de Luca-Fradley, K.; Ruiz-Pérez, L.M.; Urbina, J.; et al. Novel azasterols as potential agents for treatment of leishmaniasis and trypanosomiasis. Antimicrob. Agents Chemother. 2004, 48, 2937–2950. [Google Scholar] [CrossRef]
- Song, Z.; Nes, W.D. Sterol biosynthesis inhibitors: Potential for transition state analogs and mechanism-based inactivators targeted at sterol methyltransferase. Lipids 2007, 42, 15–33. [Google Scholar] [CrossRef]
- Reeves, E.K.; Hoffman, E.P.; Nagaraju, K.; Damsker, J.M.; McCall, J.M. Vbp15: Preclinical characterization of a novel anti-inflammatory delta 9,11 steroid. Bioorg. Med. Chem. 2013, 21, 2241–2249. [Google Scholar] [CrossRef]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Viegelmann, C.; Parker, J.; Ooi, T.; Clements, C.; Abbott, G.; Young, L.; Kennedy, J.; Dobson, A.D.W.; Edrada-Ebel, R. Isolation and Identification of Antitrypanosomal and Antimycobacterial Active Steroids from the Sponge Haliclona simulans. Mar. Drugs 2014, 12, 2937-2952. https://doi.org/10.3390/md12052937
Viegelmann C, Parker J, Ooi T, Clements C, Abbott G, Young L, Kennedy J, Dobson ADW, Edrada-Ebel R. Isolation and Identification of Antitrypanosomal and Antimycobacterial Active Steroids from the Sponge Haliclona simulans. Marine Drugs. 2014; 12(5):2937-2952. https://doi.org/10.3390/md12052937
Chicago/Turabian StyleViegelmann, Christina, Jennifer Parker, Thengtheng Ooi, Carol Clements, Gráinne Abbott, Louise Young, Jonathan Kennedy, Alan D. W. Dobson, and RuAngelie Edrada-Ebel. 2014. "Isolation and Identification of Antitrypanosomal and Antimycobacterial Active Steroids from the Sponge Haliclona simulans" Marine Drugs 12, no. 5: 2937-2952. https://doi.org/10.3390/md12052937