Marine Bioactives and Potential Application in Sports

Abstract
:1. Introduction
2. Marine Carotenoids as Antioxidant and Thermogenic Agents


{kind=link}
{kind=link}
| Group | Bioactives | Source | Activity | Health Application |
|---|---|---|---|---|
| Pigments | β-Carotene | Dunaliella salina | Pro-vitamin A; antioxidant | Food supplement [15,16] |
| Astaxanthin | Haematococcus pluvialis | Anti-inflammatory; antioxidant | CTS and muscle soreness [5,15,17,18,19,20,21,22,23] | |
| Fucoxanthin | Undaria pinnatifida | β-oxidation and UCP upregulation | Increase in energy release from fat [24,25,26,27,28] | |
| Lutein, zeaxanthin | Chlorella pyrenoidosa | Antioxidant | Food supplement [15,29] | |
| Phycocyanin | Spirulina | Anti-inflammatory; antioxidant | Muscle soreness [15,29] | |
| Sterols | Stigmasterol | Chaetoceros | Hypocholesterolemic | Dyslipidemia [15,30] |
| Contignasterol | Petrosia contignata | Anti-inflammatory; antiallergic | Asthma/inflammatory diseases [31,32,33,34,35,36,37] | |
| Xestobergsterol | Xestospongia bergquistia | Anti-inflammatory; antiallergic | Asthma/inflammatory diseases [38,39] | |
| Clathriols | Clathria lissosclera | Anti-inflammatory | Inflammatory diseases [40,41,42,43] | |
| Vitamins | C, K, B12, A, E | Arthrospira | Antioxidant; blood cell formation | Immune system reinforcement [15,44] |
| Pavlova | and blood clotting mechanisms | |||
| Proteins | Dunaliella | Anti-catabolic | Muscular status and | |
| Arthrospira platensis | performance improvement [15] | |||
| Enzymes | SOD | Anabaena | Antioxidant; anti-inflammatory | Food supplement [15,44] |
| Carbonic Anhydrase | I. galbana | CO2 conversion into H2CO3/HCO3 | Muscular performance improvement [15] | |
| PUFA | EPA | Odontella, Pavlova | Antimicrobial; | Immune system improvement [45,46,47,48,49,50,51,52,53,54,55] |
| γ-Linolenic acid | Arthrospira | Strongly anti-inflammatory | Tissues integrity; delay of aging | |
| Others | Pacifenols | Laurencia claviformis and tasmanica | COX-inhibitor | Anti-inflammatory [56,57,58,59,60] |
| Epitaondiol | Stypopodium flabelliforme | COX-inhibitor; negative inotropism | Anti-inflammatory; reduced cardiac stress [61,62,63,64,65,66] | |
| Cycloprodigiosine | Serratia marcescens | NF-κB inhibitor; NO stimulator | Anti-inflammatory/anti-arthritic [67,68,69,70,71] | |
| Macrolides | Stylocheilus longicauda | NF-κB inhibitor; antibiotical effect | Immunomodulation; Anti-inflammatory [72,73] | |
| GABA | Porphyridium | Neurotransmitter; antioxidant [15,74] | CNS regulation; immune improvement | |
| Hymenialdisine | Stylissa massa | Inhibition of proteoglycan degradation | Decrease in joint injuries risk [75,76,77,78] | |
| Stypotriol | Stypopodium flabelliforme | PA2 and elastase release inhibition | Anti-inflammatory; less cartilage damage [63,79] |
2.1. Astaxanthin
2.2. Fucoxanthin
3. Marine Natural Products: A Potential Immunomodulating and Anti-Inflammatory Strategy?
3.1. COX Inhibitors: Pacifenol and Epitaondiol
3.2. Marine Steroids: Contignasterol, Xestobergsterol and Clathriols
3.3. Molecules Interfering with NF-κB and Immunomodulation: Cycloprodigiosin and Marine Macrolides
4. Negative Effects of Exercise on Articular Cartilage and the Protective Role of Sea Food: Stypotriol and Hymenialdisine
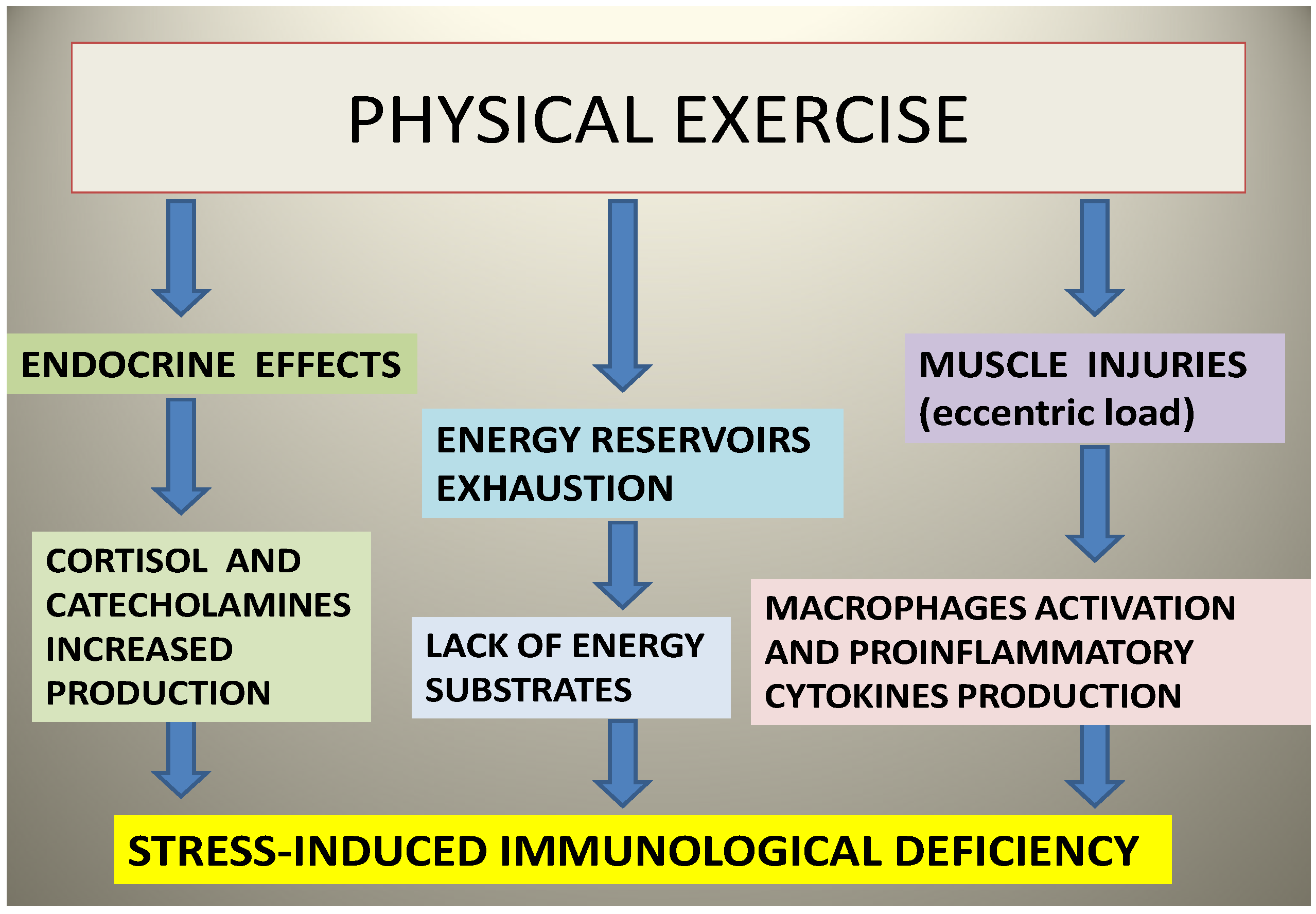
5. Sports and Immune System: A Valuable Ally from the Sea
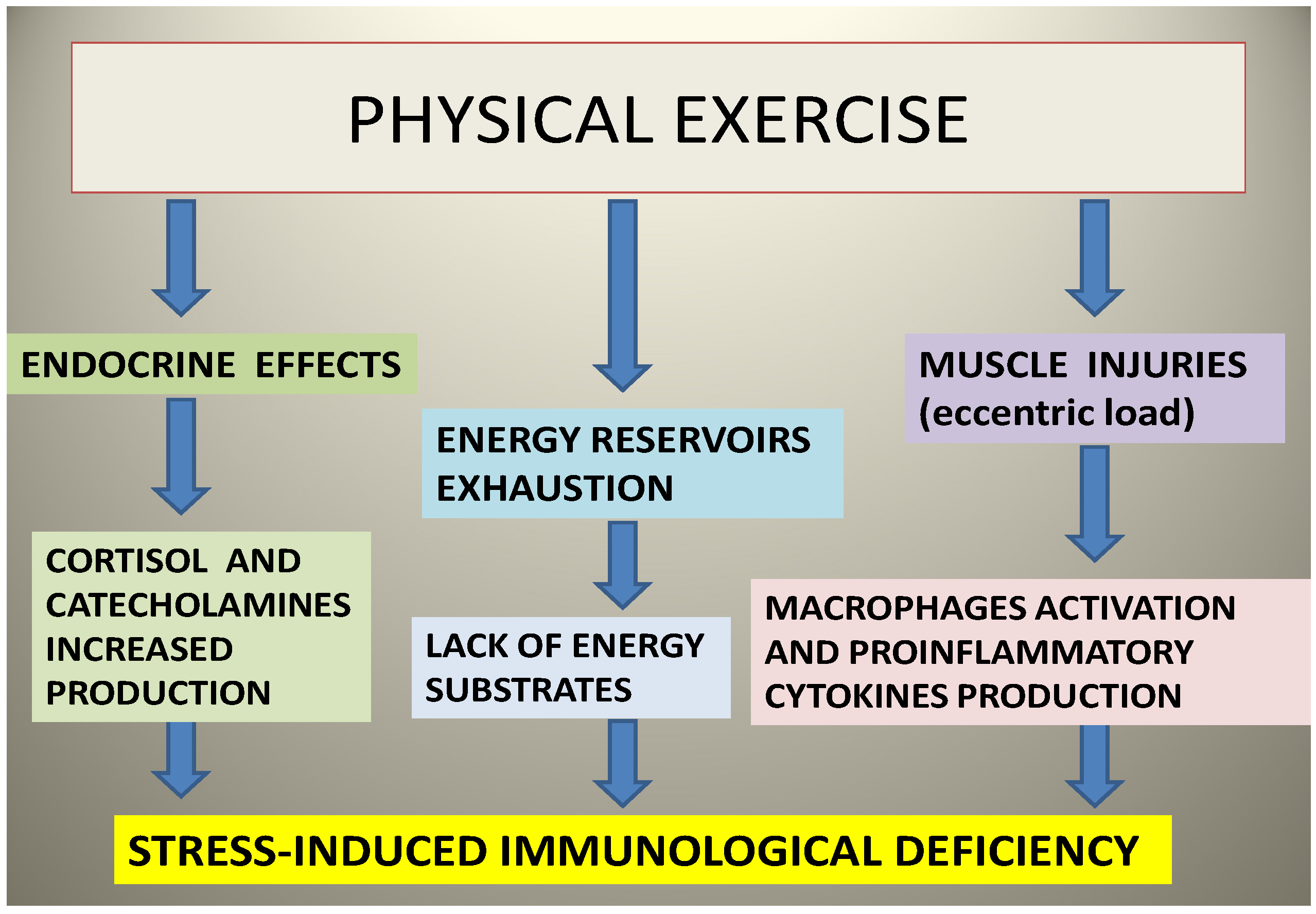
6. PUFAs as the Ideal Sports Supplement
| Common Name | Lipid Name | Chemical Name |
|---|---|---|
| Hexadecatrienoic acid (HTA) | 16:3 (n-3) | all-cis-7,10,13-hexadecatrienoic acid |
| α-linolenic acid (ALA) | 18:3 (n-3) | all-cis-9,12,15-octadecatrienoic acid |
| Stearidonic acid (SDA) | 18:4 (n-3) | all-cis-6,9,12,15,-octadecatetraenoic acid |
| Eicosatrienoic acid (ETE) | 20:3 (n-3) | all-cis-11,14,17-eicosatrienoic acid |
| Eicosatetraenoic acid (ETA) | 20:4 (n-3) | all-cis-8,11,14,17-eicosatetraenoic acid |
| Eicosapentaenoic acid (EPA, timnodonic acid) | 20:5 (n-3) | all-cis-5,8,11,14,17-eicosapentaenoic acid |
| Heneicosapentaenoic acid (HPA) | 21:5 (n-3) | all-cis-6,9,12,15,18-heneicosapentaenoic acid |
| Docosapentaenoic acid (DPA, clupanodonic acid) | 22:5 (n-3) | all-cis-7,10,13,16,19-docosapentaenoic acid |
| Docosahexaenoic acid (DHA, cervonic acid) | 22:6 (n-3) | all-cis-4,7,10,13,16,19-docosahexaenoic acid |
| Tetracosapentaenoic acid | 24:5 (n-3) | all-cis-9,12,15,18,21-tetracosapentaenoic acid |
| Tetracosahexaenoic acid (nisinic acid) | 24:6 (n-3) | all-cis-6,9,12,15,18,21-tetracosahexaenoic acid |
7. Other Compounds
7.1. Proteins
7.2. Enzymes and Vitamins
7.3. GABA
8. Conclusions
Acknowledgments
Author Contribution
Conflicts of Interest
References
- Sen, C.K. Oxidants and antioxidants in exercise. J. Appl. Physiol. 1995, 79, 675–686. [Google Scholar]
- Powers, S.K.; Jackson, M.J. Exercise-induced oxidative stress: Cellular mechanisms and impact on muscle force production. Physiol. Rev. 2008, 88, 1243–1276. [Google Scholar] [CrossRef]
- Freitas, M.; Gomes, A.; Porto, G.; Fernandes, E. Nickel induces oxidative burst, NF-κB activation and interleukin-8 production in human neutrophils. J. Biol. Inorg. Chem. 2010, 15, 1275–1283. [Google Scholar] [CrossRef]
- Fisher-Wellman, K.; Bloomer, R.J. Acute exercise and oxidative stress: A 30 year history. Dyn. Med. 2009, 8, 1–25. [Google Scholar] [CrossRef]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- Reid, M.B. Nitric oxide, reactive oxygen species, and skeletal muscle contraction. Med. Sci. Sports Exerc. 2001, 33, 371–376. [Google Scholar] [CrossRef]
- Niess, A.M.; Simon, P. Response and adaptation of skeletal muscle to exercise: The role of reactive oxygen species. Front. Biosci. 2007, 12, 4826–4838. [Google Scholar] [CrossRef]
- Sobal, J.; Marquart, L.F. Vitamin/Mineral supplement use among athletes: A review of the literature. Int. J. Sport Nutr. 1994, 4, 320–334. [Google Scholar]
- Barros, M.P.; Marin, D.P.; Bolin, A.P.; de Cássia-Santos-Macedo, R.; Campoio, T.R.; Fineto, C.; Guerra, B.A.; Polotow, T.G.; Vardaris, C.; Mattei, R.; et al. Combined astaxanthin and fish oil supplementation improves glutathione-based redox balance in rat plasma and neutrophils. Chem. Biol. Interact. 2012, 197, 58–67. [Google Scholar] [CrossRef]
- Westerterp, K.R.; Wilson, S.A.J.; Rolland, V. Diet induced thermogenesis measured over 24 h in a respiration chamber: Effect of diet composition. Int. J. Obes. 1999, 23, 287–292. [Google Scholar]
- Pattison, D.J.; Symmons, D.P.M.; Lunt, M.; Welch, A.; Bingham, S.A.; Day, N.E.; Silman, A.J. Dietary beta-cryptoxanthin and inflammatory polyarthritis: Results from a population-based prospective study. Am. J. Clin. Nutr. 2005, 82, 451–455. [Google Scholar]
- Allen, R.G.; Tresini, M. Oxidative stress and gene regulation. Free Radic. Biol. Med. 2000, 28, 463–499. [Google Scholar] [CrossRef]
- Zaccaroni, A.; Perugini, M.; D’Orazio, N.; Manera, M.; Giannella, B.; Zucchini, M.; Giammarino, A.; Riccioni, G.; Ficoneri, C.; Naccari, C.; et al. Investigation of totalarsenic in fish from the central adriatic sea (Italy) in relation to levels found in fishermen’s hair. J. Vet. Pharmacol. Ther. 2006, 29, 178–179. [Google Scholar]
- Miyashita, K. Function of marine carotenoids. Forum Nutr. 2009, 61, 136–146. [Google Scholar] [CrossRef]
- DeJesus-Raposo, M.F.; DeMorais, R.M.S.C.; DeMorais, A.M.M.B. Health application of bioactive compounds from marine microalgae. Life Sci. 2013, 93, 479–486. [Google Scholar] [CrossRef]
- Plaza, M.; Herrero, M.; Cifuentes, A.; Ibañez, E. Innovative natural functional ingredients from microalgae. J. Agric. Food Chem. 2009, 57, 7159–7170. [Google Scholar] [CrossRef]
- Shimidzu, N. Carotenoids as singlet oxygen quenchers in marine organisms. Fish. Sci. 1996, 62, 134–137. [Google Scholar] [CrossRef]
- Bennedsen, M.; Wang, X.; Willén, R.; Wadström, T.; Andersen, L.P. Treatment of H. pylori infected mice with antioxidant astaxanthin reduces gastric inflammation, bacterial load and modulates cytokine release by splenocytes. Immunol. Lett. 1999, 70, 185–189. [Google Scholar]
- Bartelt, S.; Timm, M.; Damsgaard, C.T.; Hansen, E.W.; Hansen, H.S.; Lauritzen, L. The effect of dietary fish oil-supplementation to healthy young men on oxidative burst measured by whole blood chemiluminescence. Br. J. Nutr. 2008, 99, 1230–1238. [Google Scholar]
- Caughey, G.E.; Mantzioris, E.; Gibson, R.A.; Cleland, L.G.; James, M.J. The effect on human tumor necrosis factor alpha and interleukin 1 beta production of diets enriched in n-3 fatty acids from vegetable oil or fish oil. Am. J. Clin. Nutr. 1996, 63, 116–122. [Google Scholar]
- Lara, J.J.; Economou, M.; Wallace, A.M.; Rumley, A.; Lowe, G.; Slater, C.; Caslake, M.; Sattar, N.; Lean, M.E. Benefits of salmon eating on traditional and novel vascular risk factors in young, non-obese healthy subjects. Atherosclerosis 2007, 193, 213–221. [Google Scholar] [CrossRef]
- Riccioni, G.; D’Orazio, N.; Franceschelli, S.; Speranza, L. Marine carotenoids cardiovascular risk markers. Mar. Drugs 2011, 9, 1166–1175. [Google Scholar] [CrossRef]
- Pashkow, F.J.; Watumull, D.G.; Campbell, C.L. Astaxanthin: A novel potential treatment for oxidative stress and inflammation in cardiovascular disease. Am. J. Cardiol. 2008, 101, 58–68. [Google Scholar] [CrossRef]
- Nomura, T.; Kikuchi, M.; Kubodera, A.; Kawakami, Y. Proton-donative antioxidant activity of fucoxanthin with 1,1-diphenyl-2-picrylhydrazyl (DPPH). Biochem. Mol. Biol. Int. 1997, 42, 361–370. [Google Scholar]
- Woo, M.N.; Jeon, S.M.; Shin, Y.C.; Lee, M.K.; Kang, M.A.; Choi, M.S. Anti-obese property of fucoxanthin is partly mediated by altering lipid-regulating enzymes and uncoupling proteins of visceral adipose tissue in mice. Mol. Nutr. Food Res. 2009, 53, 1603–1611. [Google Scholar] [CrossRef]
- Stock, M.J. Gluttony and thermogenesis revisited. Int. J. Obes. Relat. Metab. Disord. 1999, 23, 1105–1117. [Google Scholar]
- Gesta, S.; Tseng, Y.H.; Kahn, C.R. Developmental origin of fat: Tracking obesity to its source. Cell 2007, 131, 242–256. [Google Scholar]
- Lowell, B.B.; Susullc, V.S.; Hamann, A.; Lawitts, J.A. Development of obesity in transgenic mice after genetic ablation of brown adipose tissue. Nature 1993, 366, 740–742. [Google Scholar] [CrossRef]
- Pulz, O.; Gross, W. Valuable products from biotechnology of microalgae. Appl. Microbiol. Biotechnol. 2004, 65, 635–648. [Google Scholar] [CrossRef]
- Bandarra, N.M.; Pereira, P.A.; Batista, I.; Vilela, M. Fatty acids, sterols and α-tocopherol in Isochrysis galbana. J. Food Lipids 2003, 10, 25–34. [Google Scholar] [CrossRef]
- Terracciano, S.; Aquino, M.; Rodriguez, M.; Monti, M.C.; Casapullo, A.; Riccio, R.; Gomez-Paloma, L. Chemistry and biology of anti-inflammatory marine natural products: Molecule interfering with cyclooxygenase, NF-κB and other unidentified targets. Curr. Med. Chem. 2006, 13, 1947–1969. [Google Scholar] [CrossRef]
- Izzo, I.; Pironti, V.; Della Monica, C.; Sodano, G.; de Riccardis, F. Stereocontrolled synthesis of contignasterol’s side chain. Tetrahedron Lett. 2001, 42, 8977–8980. [Google Scholar] [CrossRef]
- Burgoyne, D.L.; Andersen, R.J.; Alle, T.M. Contignasterol, a highly oxygenated steroid with the unnatural 14-beta configuration from the marine sponge Petrosia contignata Thiele, 1899. J. Org. Chem. 1992, 57, 525–528. [Google Scholar] [CrossRef]
- Takei, M.; Burgoyne, D.L.; Andersen, R.J. Effect of contignasterol on histamine release induced by anti-immunoglobulin E from rat peritoneal mast cells. J. Pharm. Sci. 1994, 83, 1234–1235. [Google Scholar] [CrossRef]
- Parsons, J.P.; Hallstrand, T.S.; Mastronarde, J.G.; Kaminsky, D.A.; Rundell, K.W.; Hull, J.H. An official American Thoracic Society clinical practice guideline: Exercise-induced bronchoconstriction. Am. J. Respir. Crit. Care Med. 2013, 187, 1016–1027. [Google Scholar] [CrossRef]
- Milan, S.J.; Hart, A.; Wilkinson, M. Vitamin C for asthma and exercise-induced bronchoconstriction. Cochrane Libr. 2013, 23, 10–11. [Google Scholar]
- Gross, H.; Conig, G.M. Terpenoids from marine organisms: Unique structures and their pharmacological potential. Phytochem. Rev. 2006, 5, 115–141. [Google Scholar] [CrossRef]
- Kobayashi, J.; Shinonaga, H.; Shigemori, H. Xestobergsterol C, a New Pentacyclic Steroid from the Okinawan Marine Sponge Ircinia sp. and Absolute Stereochemistry of Xestobergsterol A. J. Nat. Prod. 1995, 58, 312–318. [Google Scholar] [CrossRef]
- Nakamura, A.; Kaji, Y.; Saida, K.; Ito, M.; Nagatoshi, Y.; Hara, N.; Fujimoto, Y. Synthesis ofxestobergsterol A from dehydroepiandrosterone. Tetrahedron Lett. 2005, 46, 6373–6376. [Google Scholar] [CrossRef]
- Jung, M.E.; Johnson, T.W. First total synthesis of xestobergsterol A and active structural analogues of the xestobergsterols. Tetrahedron 2001, 57, 1449–1481. [Google Scholar] [CrossRef]
- Keyzers, R.A.; Northcote, P.T.; Berridge, M.V. Clathriol B, a new 14 beta marine sterol from the New Zealand sponge Clathria lissosclera. Aust. J. Chem. 2003, 56, 279–282. [Google Scholar] [CrossRef]
- Mayer, A.M.S.; Lehmann, V.K.B. Marine pharmacology in 1998: Marine compounds with antibacterial, anticoagulant, antifungal, anti-inflammatory, anthelmintic, antiplatelet, antiprotozoal and antiviral activities; with actions on the cardiovascular, endocrine, immune, and nervous systems and other miscellaneous mechanisms of action. Pharmacologist 2000, 42, 62–69. [Google Scholar]
- Joseph, B.; Sujatha, S. Pharmacologically important natural products from marine sponges. J. Nat. Prod. 2011, 4, 5–12. [Google Scholar]
- Yingying, S.; Changai, W. The optimal growth conditions for the biomass production of Isochrysis galbana. Biotechnol. Bioprocess Eng. 2009, 14, 225–231. [Google Scholar] [CrossRef]
- Schmitz, G.; Ecker, J. The opposing effects of n-3 and n-6 fatty acids. Prog. Lipid Res. 2008, 47, 147–155. [Google Scholar] [CrossRef]
- Adarme-Vega, T.C.; Lim, D.K.; Timmins, M.; Vernen, F.; Li, Y.; Schenk, P.M. Microalgal biofactories: A promising approach towards sustainable omega-3 fatty acid production. Microb. Cell Fact. 2012, 25, 11–96. [Google Scholar]
- Bocanegra, A.; Bastida, S.; Benedí, J.; Ródenas, S.; Sánchez-Muniz, F.J. Characteristics and nutritional and cardiovascular-health properties of seaweeds. J. Med. Food 2009, 12, 236–258. [Google Scholar] [CrossRef]
- Lunn, J.; Theobald, H. The health effects of dietary unsaturated fatty acids. Nutr. Bull. 2006, 31, 178–224. [Google Scholar] [CrossRef]
- Vaughan, R.A.; Garcia-Smith, R.; Bisoffi, M.; Conn, C.A.; Trujillo, K.A. Conjugated linoleic acid or omega 3 fatty acids increase mitochondrial biosynthesis and metabolism in skeletal muscle cells. Lipids Health Dis. 2012, 11, 142–143. [Google Scholar] [CrossRef]
- Tartibian, B.; Maleki, B.H.; Abbasi, A. Omega-3 fatty acid supplementation attenuates oxidative stress and inflammatory markers after eccentric exercise in untrained men. Clin. J. Sport Med. 2011, 21, 131–137. [Google Scholar] [CrossRef]
- Walser, B.; Stebbins, C.L. Omega-3 fatty acid supplementation enhances stroke volume and cardiac output during dynamic exercise. Eur. J. Appl. Physiol. 2008, 104, 455–461. [Google Scholar] [CrossRef]
- Peoples, G.E.; McLennan, P.L.; Howe, P.R.; Groeller, H. Fish oil reduces heart rate and oxygen consumption during exercise. J. Cardiovasc. Pharmacol. 2008, 52, 504–507. [Google Scholar]
- Huffman, D.M; Altena, T.S.; Mawhinney, T.P.; Thomas, T.R. Effects of n-3 fatty acid on free tryptophan and exercise fatigue. Eur. J. Appl. Physiol. 2004, 92, 584–591. [Google Scholar]
- Fontani, G.; Lodi, L.; Migliorini, S.; Corradeschi, F. Effects of omega-3 and policosanol supplementation on attention and reactivity in athletes. J. Am. Coll. Nutr. 2009, 28, 473s–481s. [Google Scholar] [CrossRef]
- Tartibian, B.; Maleki, B.H.; Abbasi, A. Effects of ingestion of omega-3 on perceived pain and external symptoms of delayed onset muscle soreness also in untrained men. Clin. J. Sport Med. 2009, 19, 115–119. [Google Scholar] [CrossRef]
- Clifford, W.; Chang, J. Marine natural products other than pigments. J. Chem. Educ. 1973, 50, 260–262. [Google Scholar] [CrossRef]
- Baker, J.T. Some metabolites from australian marine organisms. Pure Appl. Chem. 1976, 48, 35–44. [Google Scholar] [CrossRef]
- Dembitsky, V.M.; Tolstkov, G.A. Natural Halogenated sesquiterpenes from marine organisms. Chem. Sustain. Dev. 2004, 12, 1–12. [Google Scholar] [CrossRef]
- San Martín, A.; Rovirosa, J.; Astudillo, L.; Sepúlveda, B.; Ruiz, D.; San Martín, C. Biotransformation of the marine sesquiterpene pacifenol by a facultative marine fungus. Nat. Prod. Res. 2008, 22, 1627–1632. [Google Scholar] [CrossRef]
- Areche, C.; San Martin, A.; Rovirosa, J.; Munoz, M.A.; Barragan, A.H.; Bucio, M.A.; Nathan, P.J. Stereostructure reassignment and absolute configuration of isoepitaondiol, a meroditerpenoid from Stypopodium flabelliforme. J. Nat. Prod. 2010, 73, 79–82. [Google Scholar] [CrossRef]
- Areche, C.; Vaca, I.; Labbe, P.; Delgado, J.S.; Astudillo, L.; Silva, M.; Rovirosa, J.; San Martin, A. Biotransformation of Stypotriol triacetate by Aspergillus niger. J. Mol. Struct. 2011, 998, 167–170. [Google Scholar] [CrossRef]
- Martinez, J.L.; Sepulveda, S.P.; Rovirosa, J.; San Martin, A. Efectos en aorta y auricula aisladasde rata de diacetil epitaondiol, diterpenoide del alga Stypopodium flabelliforme. An. Asoc. Quim. Argent. 1997, 85, 69–75. [Google Scholar]
- Gil, B.; Ferrandiz, M.L.; Sanz, M.J.; Terencio, M.C.; Ubeda, A.; Rovirosa, J.; San Martin, A.; Alcaraz, M.J.; Payá, M. Inhibition of inflammatory responses by epitaondiol and other marine natural products. Life Sci. 1995, 57, 25–30. [Google Scholar]
- Llanio, M.; Fernández, M.D.; Cabrera, B.; Bermejo, P.; Abad, M.J.; Payá, M.; Alcaraz, M.J. The marine plant thalassia testudinum possesses anti-inflammatory and analgesic properties. Pharmacologyonline 2006, 3, 594–600. [Google Scholar]
- Areche, C.; San Martín, A.; Rovirosa, J.; Sepúlveda, B. Gastroprotective activity of epitaondiol and sargaol. Nat. Prod. Commun. 2011, 6, 1073–1074. [Google Scholar]
- Ziemann, E.; Zembroñ-Lacny, A.; Kasperska, A.; Antosiewicz, A.; Grzywacz, T.; Garsztka, T.; Laskowski, R. Exercise training-induced changes in inflammatory mediators and heat shock proteins in young tennis players. J. Sports Sci. Med. 2013, 12, 282–289. [Google Scholar]
- Mayer, A.M.S.; Rodriguez, A.D.; Berlinck, R.G.S.; Hamann, M.T. Marine pharmacology in 2003–04: Marine compounds with anthelminthic, antibacterial, anticoagulant, antifungal, anti-inflammatory, antimalarial, antiplatelet, antiprotozoal, antituberculosis, and antiviral activities affecting the cardiovascular, immune and nervous systems and other miscellaneous mechanisms of action. Comp. Biochem. Physiol. 2007, 145, 553–581. [Google Scholar]
- Lee, J.S.; Kim, Y.S.; Park, S.; Kim, J.; Kang, S.J.; Lee, M.H.; Ryu, S.; Choi, J.M.; Oh, T.K.; Yoon, J.H. Exceptional production of both prodigiosin and cycloprodigiosin as major metabolic constituents by a novel marine bacterium, Zooshikella rubidus S1-1. Appl. Environ. Microbiol. 2011, 77, 4967–4973. [Google Scholar] [CrossRef]
- Kamata, K.; Okamoto, S.; Oka, S.; Kamata, H.; Yagisawa, H.; Hirata, H. Cycloprodigiosin hydrocloride suppresses tumor necrosis factor (TNF) alpha-induced transcriptional activatio by NF-κB. FEBS Lett. 2001, 507, 74–80. [Google Scholar] [CrossRef]
- Teshima, S.; Nakanishi, H.; Kamata, K.; Kaibori, M.; Kwon, A.H.; Kamiyama, Y.; Nishizawa, M.; Ito, S.; Okumura, T. Cycloprodigiosin up-regulates inducible nitric oxide synthase gene expression in hepatocytes stimulated by interleukin-1β. Nitric Oxide 2004, 11, 9–16. [Google Scholar] [CrossRef]
- Higa, T.; Tanaka, J. Studies in Natural Products Chemistry: Bioactive marine macrolides. Struct. Chem. 1996, 19, 549–626. [Google Scholar]
- Jacobson, P.B.; Jacobs, R.S. Fuscoside: An anti-inflammatory marine natural product which selectively inhibits 5-lipoxigenase. Part I: Physiological and biochemical studies in murine inflammatory models. J. Pharmacol. Exp. Ther. 1992, 262, 866–873. [Google Scholar]
- Morton, J.P.; Kayani, A.C.; McArdle, A.; Drust, B. The exercise-induced stress response of skeletal muscle, with specific emphasis on humans. Sports Med. 2009, 39, 643–662. [Google Scholar] [CrossRef]
- Enna, S.J; McCarson, K.E. The role of GABA in the mediation and perception of pain. Adv. Pharmacol. 2006, 54, 1–27. [Google Scholar] [CrossRef]
- Tasdemir, D.; Mallon, R.; Greenstein, M.; Feldberg, L.; Kim, S.; Collins, K.; Wojciechowicz, D.; Mangalindan, G.; Concepcion, G.; Harper, M.K.; et al. Aldisine alkaloids from the Philippine sponge Stylissa massa are potent inhibitors of mitogen-activated protein kinase-1 (MEK-1). J. Med. Chem. 2002, 45, 529–532. [Google Scholar] [CrossRef]
- Breton, J.J.; Fletcher, M.C. The natural product hymenialdisine inhibits interleukin-8 production in U937 cells by inhibition of nuclear factor-B. J. Pharmacol. Exp. Ther. 1997, 282, 459–466. [Google Scholar]
- Skropeta, D.; Pastro, N.; Zivanovic, A. Kinase inhibitors from marine sponges. Mar. Drugs 2011, 9, 2131–2154. [Google Scholar] [CrossRef]
- Meijer, L.; Thunnissen, A.M.; White, A.W.; Garnier, M.; Nikolic, M.; Tsai, L.H.; Walter, J.; Cleverley, K.E.; Salinas, P.C.; Wu, Y.Z.; et al. Inhibition of cyclin-dependent kinases, GSK-3beta and CK1 by hymenialdisine, a marine sponge constituent. Chem. Biol. 2000, 7, 51–63. [Google Scholar] [CrossRef]
- Tan, A.S.; Berridge, M.V. Superoxide produced by activated neutrophils efficiently reduces the tetrazolium salt, WST-1 to produce a soluble formazan: a simple colorimetric assay for measuring respiratory burst activation and for screening anti-inflammatory agents. J. Immunol. Methods 2000, 238, 59–68. [Google Scholar] [CrossRef]
- Higuera-Ciapara, I.; Valenzuela, L.F.; Goycoolea, F.M. Astaxanthin: A review of its chemistry and applications. Crit. Rev. Food Sci. Nutr. 2006, 46, 185–196. [Google Scholar] [CrossRef]
- De Bruin, A.P.K.; Oudejans, R.R.D.; Bakker, F.C. Dieting and body image in aesthetic sports: A comparison of Dutch female gymnasts and non-aesthetic sport participants. Psychol. Sport Exerc. 2007, 40, 507–520. [Google Scholar]
- El Ghoch, M.; Soave, F.; Calugi, S.; Dalle Grave, R. Eating disorders, physical fitness and sport performance: A systematic review. Nutrients 2013, 5, 5140–5160. [Google Scholar] [CrossRef]
- Sherman, R.; Thompson, R.A.; Rose, J. Body mass index and athletic performance in elite female gymnasts. J. Sport Behav. 1996, 19, 9–10. [Google Scholar]
- Cederberg, A.; Grønning, L.M.; Ahren, B.; Tasken, K.; Carlsson, P.; Enerbäck, S. FOXC2 is a winged helix gene that counteracts obesity, hypertriglyceridemia, and diet-induced insulin resistance. Cell 2001, 106, 563–573. [Google Scholar] [CrossRef]
- Echtay, K.S. Mitochondrial uncoupling proteins: What is their physiological role? Free Radic. Biol. Med. 2007, 43, 1351–1371. [Google Scholar] [CrossRef]
- Jeon, S.M.; Kim, H.J.; Woo, M.N.; Lee, M.K.; Shin, Y.C.; Park, Y.B.; Choi, M.S. Fucoxanthin-rich seaweed extract suppresses body weight gain and improves lipid metabolism in high-fat-fed C57BL/6J mice. Biotechnol. J. 2010, 5, 961–969. [Google Scholar]
- Nicholls, A.R.; Polman, R.C. Coping in sport: A systematic review. J. Sports Sci. 2007, 25, 11–31. [Google Scholar] [CrossRef]
- Habelt, S.; Hasler, C.C.; Steinbrück, K.; Majewski, M. Sport injuries in adolescents. Orthop. Rev. 2011, 3, 82–86. [Google Scholar]
- Lee, B.A.; Lee, S.H.; Oh, D.J. Effects of peripheral injury in athletes with long-term exercise participation in modern pentathlons. J. Exerc. Rehabil. 2013, 9, 481–488. [Google Scholar] [CrossRef]
- Ferreira, S.H.; Vane, J.R. New aspects of the mode of action of non steroid anti-inflammatory drugs. Ann. Rev. Pharmacol. 1974, 14, 57–73. [Google Scholar] [CrossRef]
- Bjorkman, D.J. The effect of aspirin and non steroidal anti-inflammatory drugs on prostaglandins. Am. J. Med. 1998, 105, 8–12. [Google Scholar] [CrossRef]
- Sims, J.; Fenical, W.; Wing, R.; Radlick, P. Marine natural products, pacifenol, a rare sesquiterpene containing bromine and chlorine from the red alga, Laurencia pacifica. J. Am. Chem. Soc. 1971, 93, 3774–3775. [Google Scholar] [CrossRef]
- Rovirosa, J.; San Martin, A. Antimicrobial activity of the brown alga Stypopodium flabelliforme constituents. Fitoterapia 1997, 68, 473–475. [Google Scholar]
- Soares, A.R.; Abrantes, J.L.; Souza, T.M.L.; Fontes, C.F.L.; Pereira, R.C.; Frugulhetti, I.C.D.P.P.; Teixeira, V.L. In vitro antiviral effect of meroditerpenes isolated from the Brazilian seaweed Stypopodium zonale (Dictyotales). Planta Med. 2007, 73, 1221–1224. [Google Scholar] [CrossRef]
- Pereira, D.M.; Cheel, J.; Areche, C.; San Martin, A.; Rovirosa, J.; Silva, L.R.; Valentao, P.; Andrade, P.B. Anti-proliferative activity of meroditerpenoids isolated from the brown alga Stypopodium flabelliforme against several cancer cell lines. Mar. Drugs 2011, 9, 852–862. [Google Scholar] [CrossRef]
- Al Sabi, A.; McArthur, J.; Ostroumov, V.; French, R.J. Marine toxins that target voltage-gated sodium channels. Mar. Drugs 2006, 4, 157–192. [Google Scholar] [CrossRef]
- Nahas, R.; Abatis, D.; Anagnostopoulou, M.A.; Kefalas, P.; Vagias, C.; Roussis, V. Radical-scavenging activity of Aegean Sea marine algae. Food Chem. 2007, 102, 577–581. [Google Scholar] [CrossRef]
- Ozkan, C.; Sarpel, Y.; Biçer, O.S. The effects of exercise on articular cartilage. Acta Orthop. Traumatol. Turc. 2007, 41, 13–18. [Google Scholar]
- Buckwalter, J.A.J. Sports, joint injury, and post-traumatic osteoarthritis. Orthop. Sports Phys. Ther. 2003, 33, 578–588. [Google Scholar] [CrossRef]
- Ferrando, F.S.; San Martin, A. Epitaondiol: The first polycyclic meroditerpenoid containing two fused six-membered rings forced into the twist-boat conformation. J. Org. Chem. 1995, 60, 1475–1478. [Google Scholar] [CrossRef]
- Badger, A.M.; Cook, M.N.; Swift, B.A.; Newman-Tarr, T.M.; Gowen, M.; Lark, M. Inhibition of interleukin-1-induced proteoglycan degradation and nitric oxide production in bovine articular cartilage/chondrocyte cultures by the natural product, hymenialdisine. J. Pharmacol. Exp. Ther. 1999, 290, 587–593. [Google Scholar]
- Bruunsgaard, H. Exercise induced increase in serum interleukin 6 in humans is related to muscle damage. J. Physiol. 1997, 499, 833–841. [Google Scholar]
- Baum, M.; Liesen, H. Exhausting interval exercise as a trigger for activation of the immune and coagulation system. Deutsch. Z. Sportmed. 1993, 44, 423–428. [Google Scholar]
- Niemann, D.V.; Johanssen, L.M.; Lee, J.W.; Arabatzis, K. Infectious episodes in runners before and after the Los Angeles marathon. J. Sportmed. Phys. 1990, 30, 316–328. [Google Scholar]
- Jensen, P.R.; Williams, P.G.; Oh, D.C.; Zeigler, L.; Fenical, W. Species specific secondary metabolite production in marine actinomycetes of the Genus Salinispora. Appl. Environ. Microbiol. 2007, 73, 1146–1152. [Google Scholar] [CrossRef]
- Schmitt, E.K.; Riwanto, M.; Sambandamurthy, V.; Roggo, S.; Miault, C.; Zwingelstein, C.; Krastel, P.; Noble, C.; Beer, D.; Rao, S.P.S.; et al. The natural product cyclomarin kills mycobacterium tuberculosis by targeting the Clp-C1 subunit of the caseinolytic protease. Angew. Chem. Int. Ed. 2011, 50, 5889–5891. [Google Scholar] [CrossRef]
- Trischman, J.A.; Tapiolas, D.M.; Jensen, P.R.; Dwight, R.; Fenical, W.; McKee, T.C.; Ireland, C.M.; Stout, T.J.; Clardy, J. Salinamides A and B: Anti-inflammatory depsipeptides from amarine Streptomycetes. J. Am. Chem. Soc. 1994, 116, 757–758. [Google Scholar] [CrossRef]
- Pietra, F. Secondary metabolites from marine microorganisms: Bacteria, protozoa, algae and fung. Achievements and prospects. Nat. Prod. Rep. 1997, 14, 453–464. [Google Scholar] [CrossRef]
- Mickleborough, T.D. Omega-3 polyunsaturated fatty acids in physical performance optimization. Int. J. Sport Nutr. Exerc. Metab. 2013, 23, 83–96. [Google Scholar]
- Phillips, S.M. Dietary protein requirements and adaptive advantages in athletes. Br. J. Nutr. 2012, 108, 158–167. [Google Scholar] [CrossRef]
- Waly, M.I.; Kilani, H.A.; Al-Busafi, M.S. Nutritional practise of atlete in Oman: A descriptive study? Oman Med. J. 2013, 28, 360–364. [Google Scholar] [CrossRef]
- Harnedy, P.A.; FitzGerald, R.J. Bioactive peptides from marine processing waste and shellfish: A review. J. Funct. Foods 2012, 4, 6–24. [Google Scholar] [CrossRef]
- Ursu, A.V.; Marcati, A.; Sayd, T.; Sante-Lhoutellier, V.; Djelveh, G.; Michaud, P. Extraction, fractionation and functional properties of proteins from the microalgae Chlorella vulgaris. Bioresour. Technol. 2014, 157, 134–139. [Google Scholar] [CrossRef]
- Rodriguez, N.R.; Di Marco, N.M.; Langley, S. American college of sports medicine position stand. Nutrition and athletic performance. Med. Sci. Sports Exerc. 2009, 41, 709–731. [Google Scholar] [CrossRef]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef]
- Honkanen, P. Consumer acceptance of (marine) functional food. Mari. Funct. Food 2009, 1, 141–154. [Google Scholar]
- Hurst, S.; Zainal, Z.; Caterson, B.; Hughes, C.E.; Harwood, J.L. Dietary fatty acids and arthritis. Prostaglandins Leukot. Essent. Fatty Acids 2010, 82, 315–318. [Google Scholar] [CrossRef]
- Stamp, L.K.; James, M.J.; Cleland, L.G. Diet and rheumatoid arthritis: A review of theliterature. Semin. Arthritis Rheum. 2005, 35, 77–94. [Google Scholar] [CrossRef]
- Dekkers, J.C.; van Doornen, L.J.; Kemper, H.C. The role of antioxidant vitamins and enzymes in the prevention of exercise-induced muscle damage. Sports Med. 1996, 21, 213–238. [Google Scholar]
- Mayer, A.M.S.; Rodríguez, A.D.; Taglialatela-Scafati, O.; Fusetani, N. Marine pharmacology in 2009–2011: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, and antiviral activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar. Drugs 2013, 11, 2510–2573. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.M.; Rajapakse, N.; Kim, S.K. Anti-inflammatory effect of Ishige okamurae ethanolic extract via inhibition of NF-κB transcription factor in RAW264.7 cells. Phytother. Res. 2009, 23, 628–634. [Google Scholar] [CrossRef]
- Mancini-Filho, J.; Novoa, A.V.; González, A.E.; de Andrade-Wartha, E.R.; de Silva, A.M.; Pinto, J.R.; Mancini, D.A. Free phenolic acids from the seaweed Halimeda monile with antioxidant effect protecting against liver injury. Z. Naturforsch C 2009, 64, 657–663. [Google Scholar]
- Grünewald, N.; Alban, S. Optimized and standardized isolation and structural characterization of anti-inflammatory sulfated polysaccharides from the red alga Delesseria sanguinea (Hudson) Lamouroux (Ceramiales, Delesseriaceae). Biomacromolecules 2009, 9, 2998–3008. [Google Scholar] [CrossRef]
- Badiu, D.L; Luque, R.; Dumitrescu, E.; Craciun, A.; Dinca, D. Amino acids from Mytilus galloprovincialis and Rapana venosa molluscs accelerate skin wounds healing via enhancement of dermal and epidermal neoformation. Protein J. 2010, 29, 81–92. [Google Scholar] [CrossRef]
- Newman, D.; Cragg, G. Marine natural products and related compounds in clinical and advanced preclinical trials. J. Nat. Prod. 2004, 67, 1216–1238. [Google Scholar] [CrossRef]
- D’Orazio, N.; Gammone, M.A.; Gemello, E.; DeGirolamo, M.; Cusenza, S.; Riccioni, G. Marine bioactives: Pharmacological properties and potential applications against inflammatory diseases. Mar. Drugs 2012, 10, 812–833. [Google Scholar] [CrossRef]
- D’Orazio, N.; Gemello, E.; Gammone, M.A.; DeGirolamo, M.; Ficoneri, C.; Riccioni, G. Fucoxantin: A treasure from the sea. Mar. Drugs 2012, 10, 604–616. [Google Scholar] [CrossRef]
- Huleihel, M.; Ishanu, V.; Tal, J.; Arad, S.M. Activity of Porphyridium sp. polysaccharide against Herpes simplex viruses in vitro and in vivo. J. Biochem. Biophys. Methods 2002, 50, 189–200. [Google Scholar] [CrossRef]
- Dvir, I.; Stark, A.H.; Chayoth, R.; Madar, Z.; Arad, S.M. Hypocholesterolemic effects of nutraceuticals produced from the red microalga Porphyridium sp. in rats. Nutrients 2009, 1, 156–167. [Google Scholar] [CrossRef]
- Gardeva, E.; Toshkova, R.; Minkova, K.; Gigova, L. Cancer protective action of polysaccharide derived from microalga Porphyridium cruentum, a biological background. Biotechnol. Equip. 2009, 23, 783–787. [Google Scholar]
- Arad, S.M.; Atar, D. Viscosupplementation with algal polysaccharides in the treatment of arthritis. WO/2007/066340; (Ben Gurion University of the Negev Research and Development Authority), 14 June 2007. [Google Scholar]
© 2014 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Gammone, M.A.; Gemello, E.; Riccioni, G.; D'Orazio, N. Marine Bioactives and Potential Application in Sports. Mar. Drugs 2014, 12, 2357-2382. https://doi.org/10.3390/md12052357
Gammone MA, Gemello E, Riccioni G, D'Orazio N. Marine Bioactives and Potential Application in Sports. Marine Drugs. 2014; 12(5):2357-2382. https://doi.org/10.3390/md12052357
Chicago/Turabian StyleGammone, Maria Alessandra, Eugenio Gemello, Graziano Riccioni, and Nicolantonio D'Orazio. 2014. "Marine Bioactives and Potential Application in Sports" Marine Drugs 12, no. 5: 2357-2382. https://doi.org/10.3390/md12052357