BMAA Inhibits Nitrogen Fixation in the Cyanobacterium Nostoc sp. PCC 7120

Abstract
:1. Introduction
2. Results
2.1. 14C-BMAA Uptake

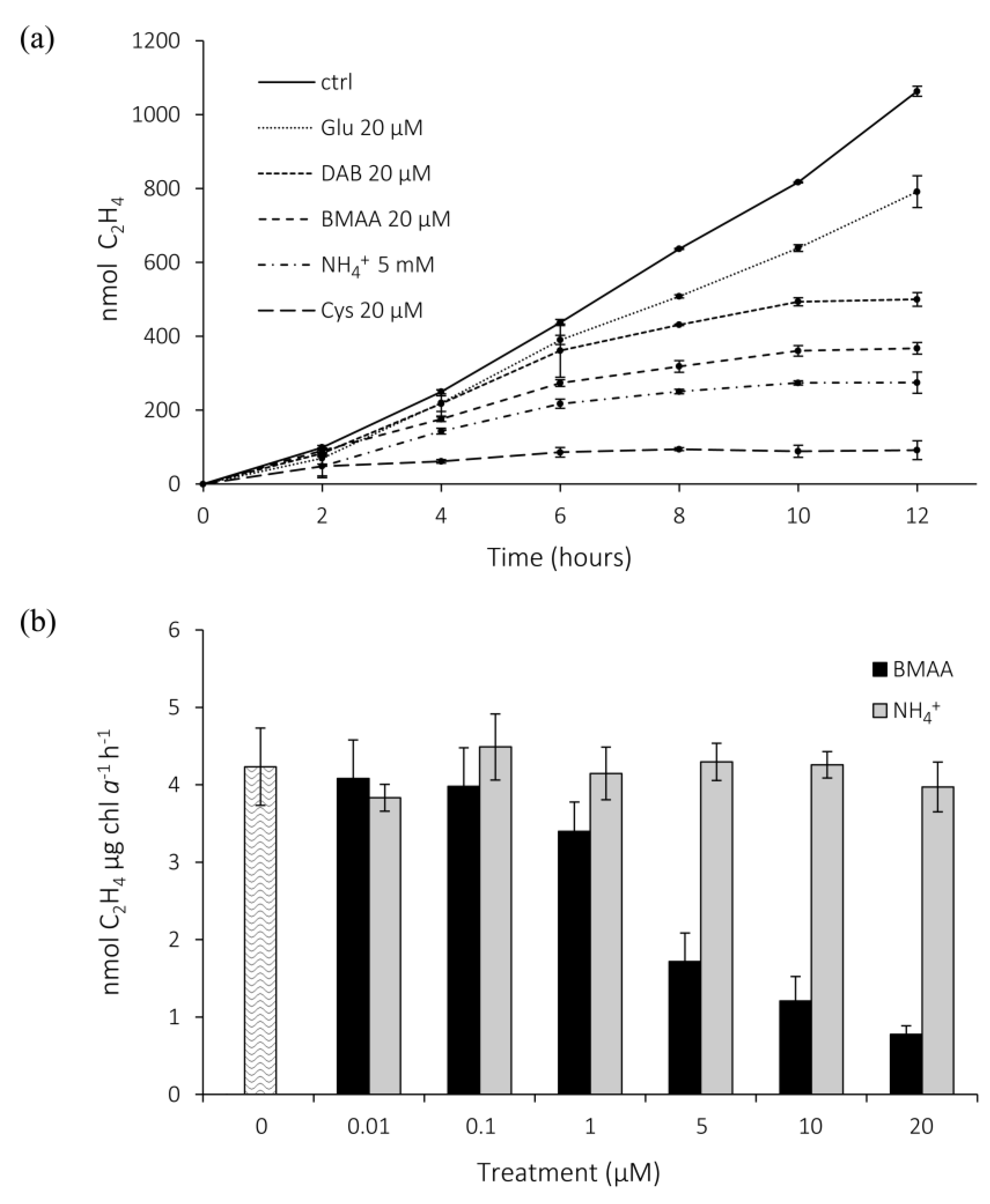
2.2. Effects of BMAA on Nitrogenase Activity


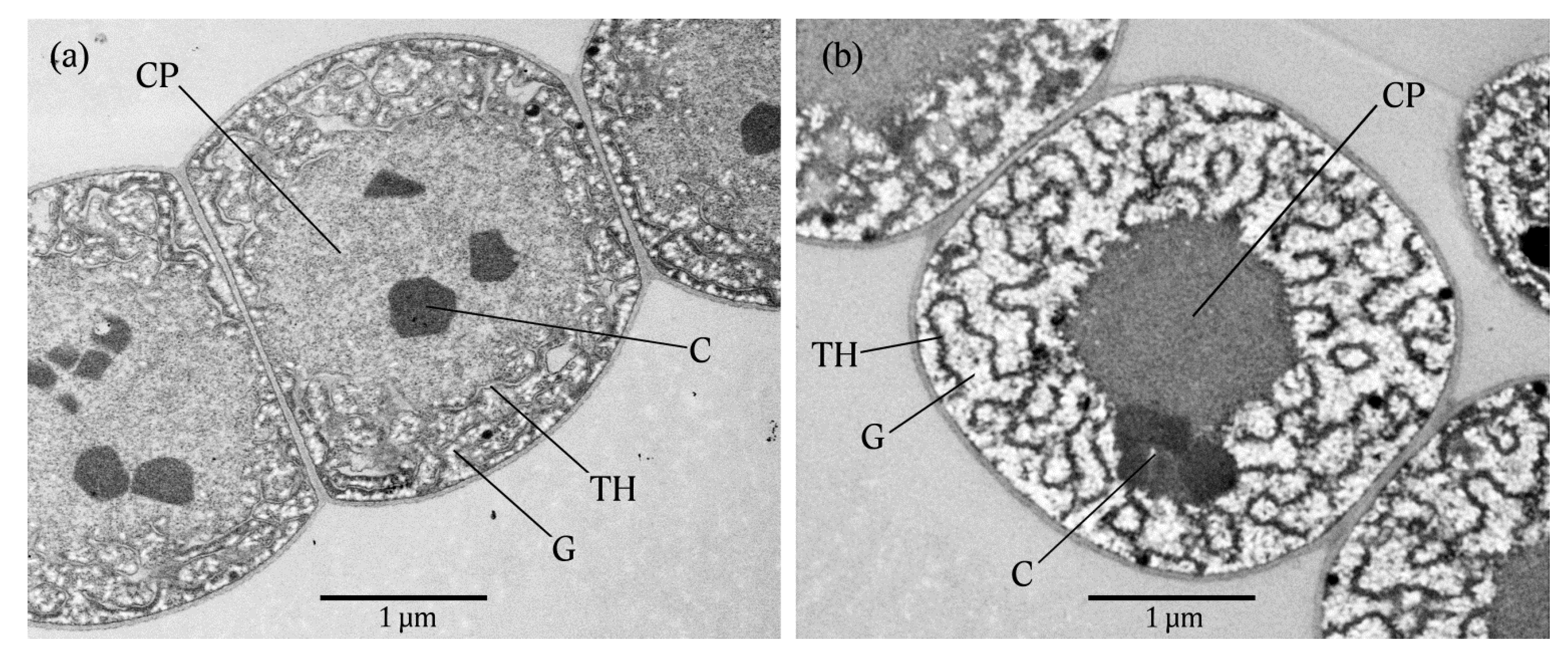
2.3. Transmission Electron Microscopy (TEM)
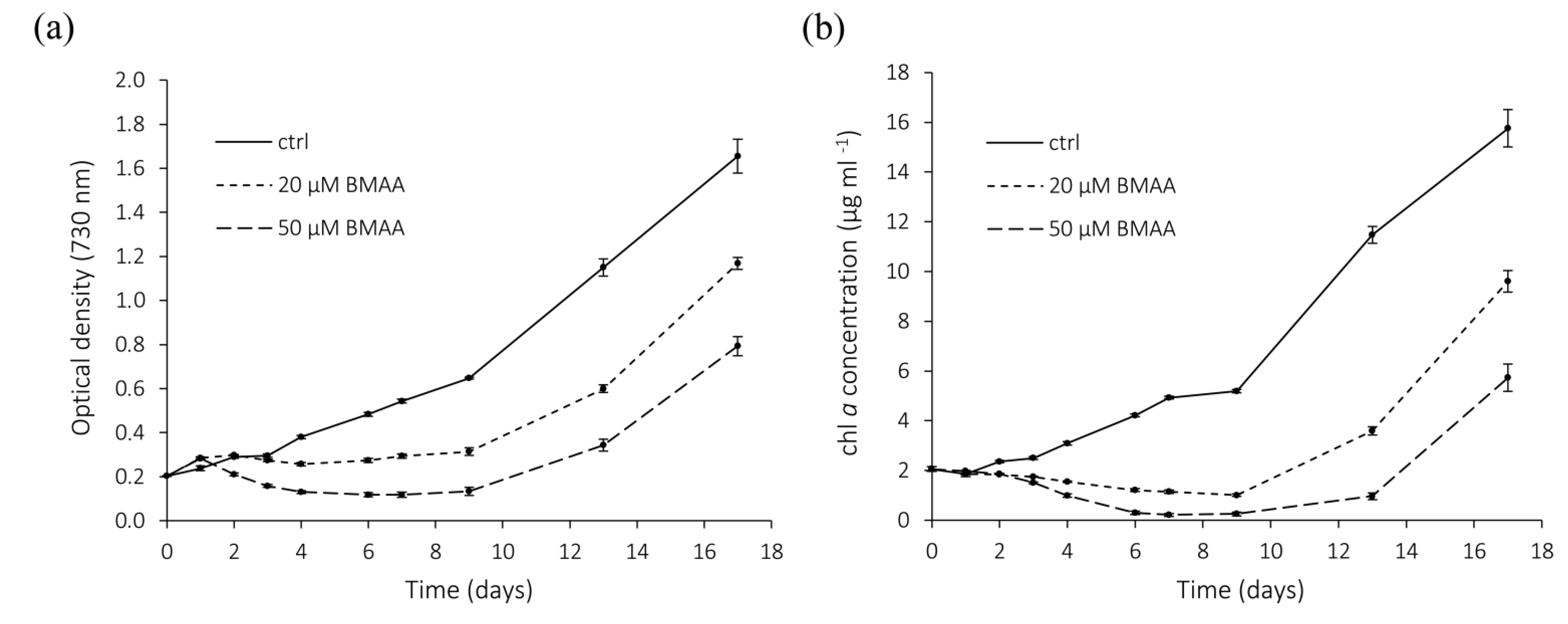
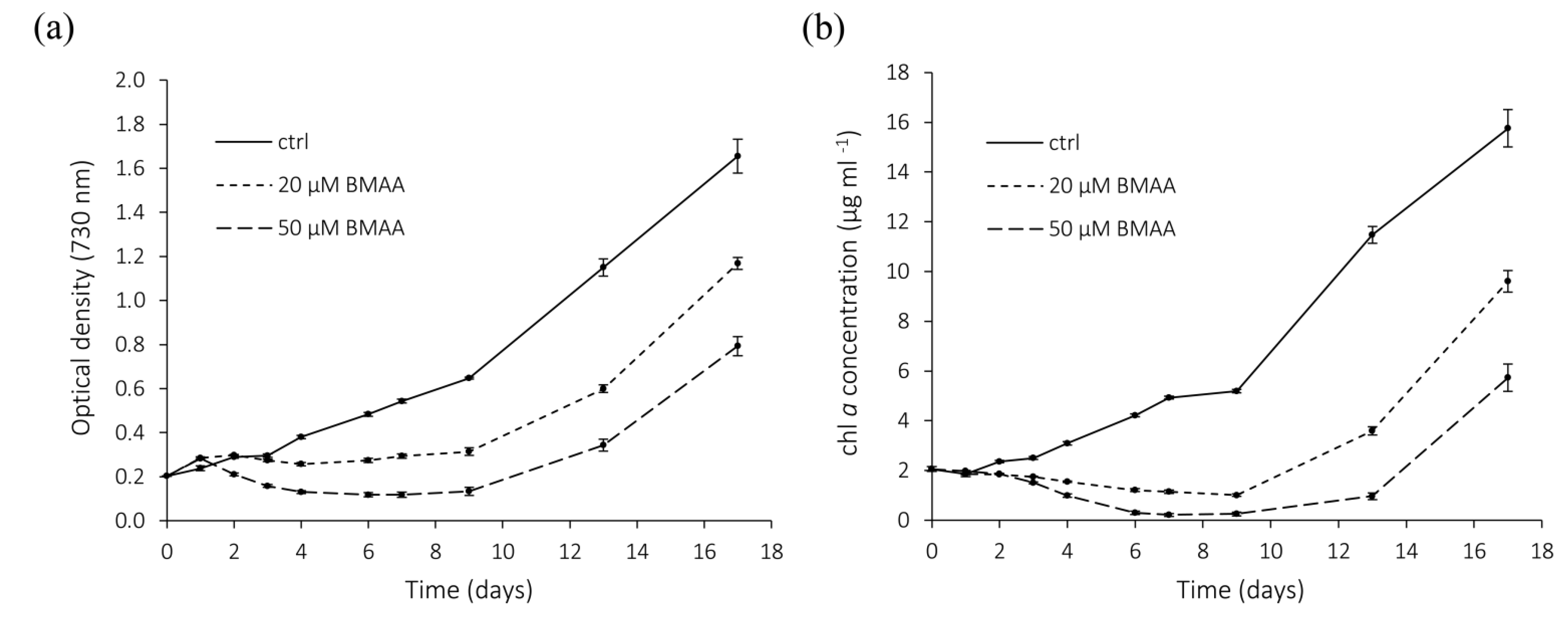
2.4. BMAA—Long Term Effects

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BMAA treatment | Nitrogenase activity (nmol ethylene µg chl a−1 h−1) | ±SE (n = 3) |
|---|---|---|
| No BMAA added | 5.9 | 0.240 |
| Fresh BMAA added | 0.2 * | 0.033 |
| 1 week | 0.2 | 0.082 |
| 2 weeks | 0.4 | 0.038 |
| 3 weeks | 0.4 | 0.032 |
| 4 weeks | 0.2 (ns) | 0.029 |
3. Discussion
4. Experimental Section
4.1. Chemicals
4.2. Organisms and Cultivation
4.3. Growth
4.4. 14C-BMAA Uptake
4.5. Nitrogenase Activity
4.5.1. Accumulated Nitrogenase Activity
4.5.2. Concentration-Dependent Effects of BMAA and NH4+ on Nitrogenase Activity
4.5.3. Effect of BMAA and Other Amino Acids and Nitrogen Sources on Nitrogenase Activity
4.5.4. Capacity of BMAA to Reduce Nitrogenase Activity in Nostoc 7120 Following Maintenance in Cell-Free BG110 Medium
4.6. Transmission Electron Microscopy (TEM)
4.7. Statistical Analysis
5. Conclusions
Acknowledgments
Conflicts of Interest
References
- Metcalf, J.S.; Codd, G.A. Cyanotoxins. In Ecology of Cyanobacteria II: Their Diversity in Space and Time; Whitton, B.A., Ed.; Springer: Dordrecht, The Netherlands, 2012; pp. 651–675. [Google Scholar]
- Dittmann, E.; Fewer, D.P.; Neilan, B.A. Cyanobacterial toxins: Biosynthetic routes and evolutionary roots. FEMS Microbiol. Rev. 2012. [Google Scholar] [CrossRef]
- Stucken, K.; John, U.; Cembella, A.; Murillo, A.A.; Soto-Liebe, K.; Fuentes-Valdes, J.J.; Friedel, M.; Plominsky, A.M.; Vaquez, A.; Glockner, G. The smallest known genomes of multicellular and toxic cyanobacteria: Comparison, minimal gene sets for linked traits and the evolutionary implications. PLoS One 2010, 5, e9235. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Sivonen, K.; Rouhiainen, L.; Fewer, D.P.; Lyra, C.; Rantala-Ylinen, A.; Vestola, J.; Jokela, J.; Rantasärkkä, K.; Li, Z.; et al. Genome-derived insights into the biology of the hepatotoxic bloom-forming cyanobacterium Anabaena sp. strain 90. BMC Genomics 2012, 13, 613. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Georgia, T.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce β-N-methylamino-l-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef]
- Esterhuizen, M.; Downing, T.G. β-N-methylamino-l-alanine (BMAA) in novel South African cyanobacterial isolates. Ecotoxicol. Environ. Saf. 2008, 71, 309–313. [Google Scholar] [CrossRef]
- Spencer, P.S.; Kisby, G.E.; Ludolph, A.C. Slow toxins, biologic markers, and long-latency neurodegenerative disease in the western Pacific region. Neurology 1991, 41, 62–66. [Google Scholar] [CrossRef]
- Murch, S.J.; Cox, P.A.; Banack, S.A. A mechanism for slow release of biomagnified cyanobacterial neurotoxins and neurodegenerative disease in Guam. Proc. Natl. Acad. Sci. USA 2004, 101, 12228–12231. [Google Scholar] [CrossRef]
- Carmichael, W.W.; Jones, C.L.A.; Mahmood, N.A.; Theiss, W.C.; Krogh, P. Algal toxins and water-based diseases. Crit. Rev. Environ. Control 1985, 15, 275–313. [Google Scholar] [CrossRef]
- Spencer, P.S.; Nunn, P.B.; Hugon, J.; Ludolph, A.; Roy, D.N. Motorneurone disease on Guam: Possible role of a food neurotoxin. Lancet 1986, 1, 965. [Google Scholar]
- Pablo, J.; Banack, S.A.; Cox, P.A.; Johnson, T.E.; Papapetropoulos, S.; Bradley, W.G.; Buck, A.; Mash, D.C. Cyanobacterial neurotoxin BMAA in ALS and Alzheimer’s disease. Acta Neurol. Scand. 2009, 120, 216–225. [Google Scholar] [CrossRef]
- Purdie, E.L.; Metcalf, J.S.; Kashmiri, S.; Codd, G.A. Toxicity of the cyanobacterial neurotoxin β-N-methylamino-l-alanine to three aquatic animal species. Amyotroph. Lateral Scler. Front. Degener. 2009, 10, 67–70. [Google Scholar]
- Purdie, E.L.; Samsudin, S.; Eddy, F.B.; Codd, G.A. Effects of the cyanobacterial neurotoxin β-N-methylamino-l-alanine on the early-life stage development of zebrafish (Danio rerio). Aqua Toxicol. 2009, 95, 279–284. [Google Scholar] [CrossRef]
- Lürling, M.; Faassen, E.J.; van Eenennaam, J.S. Effects of the cyanobacterial neurotoxin beta-N-methylamino-l-alanine (BMAA) on the survival, mobility and reproduction of Daphnia magna. J. Plankton Res. 2011, 33, 333–342. [Google Scholar] [CrossRef]
- Esterhuizen-Londt, M.; Pflugmacher, S.; Downing, T.G. The effect of β-N-methylamino-l-alanine (BMAA) on oxidative stress response enzymes of the macrophyte Ceratophyllum demersum. Toxicon 2011, 57, 803–810. [Google Scholar] [CrossRef]
- Contardo-Jara, V.; Funke, M.S.; Peuthert, A.; Pflugmacher, S. β-N-methylamino-l-alanine exposure alters defense against oxidative stress in aquatic plants Lomariopsis lineata, Fontinalis antipyretica, Riccia fluitans and Taxiphyllum barbieri. Ecotoxicol. Environ. Saf. 2013, 88, 72–78. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Banack, S.A.; Lindsay, J.; Morrison, L.F.; Cox, P.A.; Codd, G.A. Co-occurrence of β-N-methylamino-l-alanine, a neurotoxic amino acid with other cyanobacterial toxins in British waterbodies, 1990–2004. Environ. Microbiol. 2008, 10, 702–708. [Google Scholar] [CrossRef]
- Jonasson, S.; Eriksson, J.; Berntzon, L.; Spáčil, Z.; Ilag, L.L.; Ronnevi, L.-O.; Rasmussen, U.; Bergman, B. Transfer of a cyanobacterial neurotoxin within a temperate aquatic ecosystem suggests pathways for human exposure. Proc. Nat. Acad. Sci. USA 2010, 107, 9252–9257. [Google Scholar] [CrossRef]
- Krüger, T.; Mönch, B.; Oppenhäuser, S.; Luckas, B. LC-MS/MS determination of the isomeric neurotoxins BMAA (β-N-methylamino-l-alanine) and DAB (2,4-diaminobutyric acid) in cyanobacteria and seeds of Cycas revoluta and Lathyrus latifolius. Toxicon 2010, 55, 547–557. [Google Scholar] [CrossRef]
- Faassen, E.J.; Gillissen, F.; Lürling, M. A comparative study on three analytical methods for the determination of the neurotoxin BMAA in cyanobacteria. PLoS One 2012, 7, e36667. [Google Scholar]
- Downing, S.; Banack, S.A.; Metcalf, J.S.; Cox, P.A.; Downing, T.G. Nitrogen starvation of cyanobacteria results in the production of β-N-methylamino-l-alanine. Toxicon 2011, 58, 187–194. [Google Scholar] [CrossRef]
- Esterhuizen-Londt, M.; Downing, T.G. Solid phase extraction of β-N-methylamino-l-alanine (BMAA) from South African water supplies. Water SA 2011, 37, 523–528. [Google Scholar]
- Lehtimäki, J.; Moisander, P.; Sivonen, K.; Kononen, K. Growth, nitrogen fixation, and nodularin production by two Baltic Sea cyanobacteria. Appl. Environ. Microbiol. 1997, 63, 1647–1656. [Google Scholar]
- Rapala, J.; Sivonen, K.; Lyra, C.; Niemelä, S.I. Variation of microcystins, cyanobacterial hepatotoxins, in Anabaena spp. as a function of growth stimuli. Appl. Environ. Microbiol. 1997, 63, 2206–2212. [Google Scholar]
- Downing, S.; van de Venter, M.; Downing, T.G. The effect of exogenous β-N-methylamino-l-alanine on the growth of Synechocystis PCC6803. Microb. Ecol. 2012, 63, 149–156. [Google Scholar] [CrossRef]
- Larsson, J.; Nylander, J.A.A.; Bergman, B. Genome fluctuations in cyanobacteria reflect evolutionary, developmental and adaptive traits. BMC Evol. Biol. 2011, 11, 187. [Google Scholar] [CrossRef]
- Rippka, R.; Deruelles, J.; Waterbury, J.B.; Herdman, M.; Stanier, R.Y. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J. Gen. Microbiol. 1979, 111, 1–61. [Google Scholar] [CrossRef]
- Sivonen, K.; Halinen, K.; Sihvonen, L.M.; Koskenniemi, K.; Sinkko, H.; Rantasärkkä, K.; Moisander, P.H.; Lyra, C. Bacterial diversity and function in the Baltic Sea with an emphasis on cyanobacteria. AMBIO 2007, 36, 180–185. [Google Scholar] [CrossRef]
- Bergman, B.; Sandh, G.; Lin, S.; Larsson, J.; Carpenter, E.J. Trichodesmium—A widespread marine cyanobacterium with unusual nitrogen fixation properties. FEMS Microbiol. Rev. 2013, 37, 286–302. [Google Scholar] [CrossRef]
- Flores, E.; Herrero, A. Assimilatory Nitrogen Metabolism and Its Regulation. In The Molecular Biology of Cyanobacteria; Bryant, D.A., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994; pp. 487–517. [Google Scholar]
- Rawson, D.M. The effects of exogenous amino-acids on growth and nitrogenase activity in the cyanobacterium Anabaena cylindrica PCC 7122. J. Gen. Microbiol. 1985, 131, 2549–2554. [Google Scholar]
- Allen, M.M.; Smith, A.J. Nitrogen chlorosis in blue-green algae. Arch. Microbiol. 1969, 69, 114–120. [Google Scholar]
- Montesinos, M.L.; Herrero, A.; Flores, E. Amino acid transport systems required for diazotrophic growth in the cyanobacterium Anabaena sp. strain PCC 7120. J. Bacteriol. 1995, 177, 3150–7157. [Google Scholar]
- Pernil, R.; Picossi, S.; Mariscal, V.; Herrero, A.; Flores, E. ABC-type amino acid uptake transporters Bgt and N-II of Anabaena sp. strain PCC 7120 share an ATPase subunit and are expressed in vegetative cells and heterocysts. Mol. Microbiol. 2008, 67, 1067–1080. [Google Scholar] [CrossRef]
- Nunn, P.B.; Ponnusamy, M. β-N-methylaminoalanine (BMAA): Metabolism and metabolic effects in model systems and in neural and other tissues of the rat in vitro. Toxicon 2009, 54, 85–94. [Google Scholar] [CrossRef]
- Weiss, J.H.; Choi, D.W. Beta-N-methylamino-l-alanine neurotoxicity: Requirement for bicarbonate as a cofactor. Science 1988, 241, 973–975. [Google Scholar]
- Nunn, P.B.; O’Brien, P. The interaction of β-N-methylamino-l-alanine with bicarbonate: An 1H-NMR study. FEBS Lett. 1989, 251, 31–35. [Google Scholar] [CrossRef]
- Myers, T.G.; Nelson, S.D. Neuroactive carbamate adducts of β-N-methylamino-l-alanine and ethylenediamine. Detection and quantitation under physiological conditions by 13C NMR. J. Biol. Chem. 1990, 265, 10193–10195. [Google Scholar]
- García-González, M.; Sivak, M.N.; Guerrero, M.G.; Preiss, J.; Lara, C. Depression of carbon flow to the glycogen pool induced by nitrogen assimilation in intact cells of Anacystis nidulans. Physiol. Plant. 1992, 86, 360–364. [Google Scholar] [CrossRef]
- Ernst, A.; Böger, P. Glycogen accumulation and the induction of nitrogenase activity in the heterocyst-forming cyanobacterium Anabaena variabilis. J. Gen. Microbiol. 1985, 131, 3147–3153. [Google Scholar]
- Liu, X.; Rush, T.; Zapata, J.; Lobner, D. β-N-methylamino-l-alanine induces oxidative stress and glutamate release through action on system Xc−. Exp. Neurol. 2009, 217, 429–433. [Google Scholar] [CrossRef]
- Rao, S.D.; Banack, S.A.; Cox, P.A.; Weiss, J.H. BMAA selectively injures motor neurons via AMPA/kainate receptor activation. Exp. Neurol. 2006, 201, 244–252. [Google Scholar] [CrossRef]
- Ross, S.M.; Seelig, M.; Spencer, P.S. Specific antagonism of excitotoxic action of ‘uncommon’ amino acids assayed in organotypic mouse cortical cultures. Brain Res. 1987, 425, 120–127. [Google Scholar] [CrossRef]
- Weiss, J.H.; Christine, C.W.; Choi, D.W. Bicarbonate dependence of glutamate receptor activation by β-N-methylamino-l-alanine: Channel recording and study with related compounds. Neuron 1989, 3, 321–326. [Google Scholar] [CrossRef]
- Lobner, D.; Piana, P.M.; Salous, A.K.; Peoples, R.W. β-N-methylamino-l-alanine enhances neurotoxicity through multiple mechanisms. Neurobiol. Dis. 2007, 25, 360–366. [Google Scholar] [CrossRef]
- Cucchiaroni, M.L.; Viscomi, M.T.; Bernardi, G.; Molinari, M.; Guatteo, E.; Mercuri, N.B. Metabotropic glutamate receptor 1 mediates the electrophysiological and toxic actions of the cycad derivative β-N-methylamino-l-alanine on substantia nigra pars compacta DAergic neurons. J. Neurosci. 2010, 30, 5176–5188. [Google Scholar] [CrossRef]
- Chiu, A.S.; Gehringer, M.M.; Welch, J.H.; Neilan, B.A. Does α-amino-β-methylaminopropionic acid (BMAA) play a role in neurodegeneration? Int. J. Environ. Res. Public Health 2011, 8, 3728–3746. [Google Scholar] [CrossRef]
- Gallon, J.R. Reconciling the incompatible: N2 fixation and O2. New Phyt. 1992, 122, 571–609. [Google Scholar] [CrossRef]
- Chen, G.Q.; Cui, C.; Mayer, M.L.; Gouaux, E. Functional characterization of a potassium-selective prokaryotic glutamate receptor. Nature 1999, 402, 817–821. [Google Scholar] [CrossRef]
- Cameron, J.C.; Pakrasi, H.B. Essential role of glutathione in acclimation to environmental and redox perturbations in the cyanobacterium Synechocystis sp. PCC 6803. Plant Physiol. 2010, 154, 1672–1685. [Google Scholar] [CrossRef]
- Rodgers, K.; Dunlop, R. The cyanobacteria-derived neurotoxin BMAA can be incorporated into cell proteins and could thus be an environmental trigger for ALS and other neurological diseases associated with protein misfolding. Amyotroph. Lateral Scler. Front. Degener. 2011, 12, 22. [Google Scholar]
- Olney, J.W.; Zorumski, C.; Price, M.T.; Labruyere, J. l-cysteine, a bicarbonate-sensitive endogenous excitotoxin. Science 1990, 248, 596–599. [Google Scholar]
- Chen, C.H.; Flory, W.; Koeppe, R.E. Variation of neurotoxicity of l- and d-2,4-diaminobutyric acid with route of administration. Toxicol. Appl. Pharm. 1972, 23, 334–338. [Google Scholar] [CrossRef]
- Berman-Frank, I.; Bidle, K.D.; Haramaty, L.; Falkowski, P.G. The demise of the marine cyanobacterium, Trichodesmium spp. via an autocatalyzed cell death pathway. Limnol. Oceanogr. 2004, 49, 997–1005. [Google Scholar] [CrossRef]
- Bidle, K.D.; Falkowski, P.G. Cell death in planktonic, photosynthetic microorganisms. Nat. Rev. Microbiol. 2004, 2, 643–655. [Google Scholar] [CrossRef]
- Franklin, D.J.; Brussaard, C.P.D.; Berges, J.A. What is the role and nature of programmed cell death in phytoplankton ecology? Eur. J. Phycol. 2006, 41, 1–14. [Google Scholar] [CrossRef]
- Orellana, M.V.; Pang, W.L.; Durand, P.M.; Whitehead, K.; Baliga, N.S. A role for programmed cell death in the microbial loop. PLoS One 2013, 8, e62595. [Google Scholar]
- Asplund-Samuelsson, J.; Bergman, B.; Larsson, J. Prokaryotic caspase homologs: Phylogenetic patterns and functional characteristics reveal considerable diversity. PLoS One 2012, 7, e49888. [Google Scholar]
- Talling, J.F.; Driver, D. Some Problems in the Estimation of Chlorophyll-a in Phytoplankton. In Primary Productivity Measurement, Marine and Freshwater; Doty, M.S., Ed.; U.S. Atomic Energy Commision Report No. TID-7633; AEC: Honolulu, HI, USA, 1961; pp. 142–146. [Google Scholar]
- Capone, D.G.; Montoya, J.P. Nitrogen Fixation and Denitrification. In Methods in Microbiology. Marine Microbiology; Paul, J.H., Ed.; Academic Press Ltd.: San Diego, CA, USA, 2001; Volume 30, pp. 501–515. [Google Scholar]
- Lundgren, P.; Söderbäck, E.; Singer, A.; Carpenter, E.J.; Bergman, B. Katagnymene: Characterization of a novel marine diazotroph. J. Phycol. 2001, 37, 1052–1062. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Berntzon, L.; Erasmie, S.; Celepli, N.; Eriksson, J.; Rasmussen, U.; Bergman, B. BMAA Inhibits Nitrogen Fixation in the Cyanobacterium Nostoc sp. PCC 7120. Mar. Drugs 2013, 11, 3091-3108. https://doi.org/10.3390/md11083091
Berntzon L, Erasmie S, Celepli N, Eriksson J, Rasmussen U, Bergman B. BMAA Inhibits Nitrogen Fixation in the Cyanobacterium Nostoc sp. PCC 7120. Marine Drugs. 2013; 11(8):3091-3108. https://doi.org/10.3390/md11083091
Chicago/Turabian StyleBerntzon, Lotta, Sven Erasmie, Narin Celepli, Johan Eriksson, Ulla Rasmussen, and Birgitta Bergman. 2013. "BMAA Inhibits Nitrogen Fixation in the Cyanobacterium Nostoc sp. PCC 7120" Marine Drugs 11, no. 8: 3091-3108. https://doi.org/10.3390/md11083091