Strategies for the Development of Conotoxins as New Therapeutic Leads


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Peptidomimetics
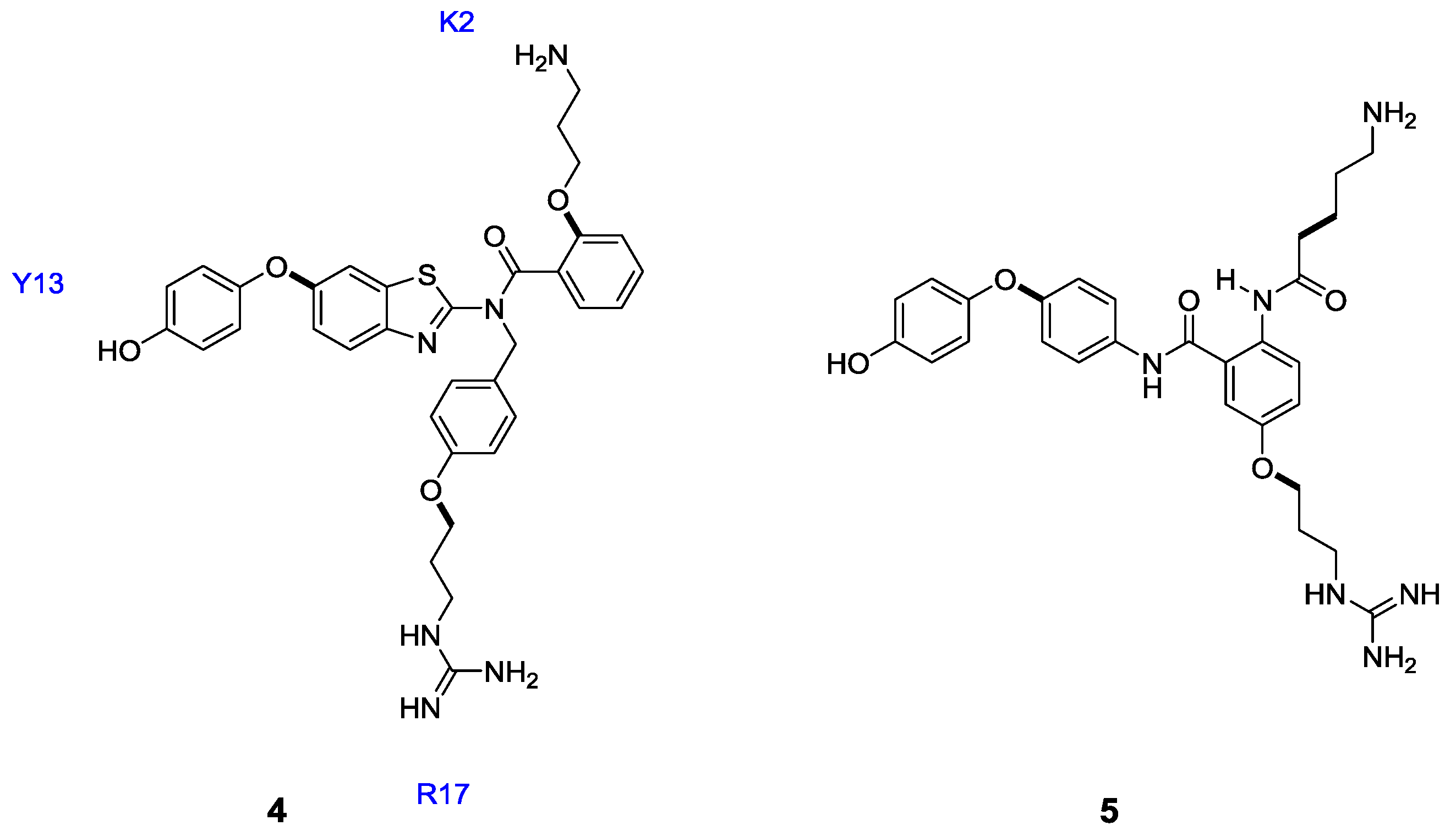
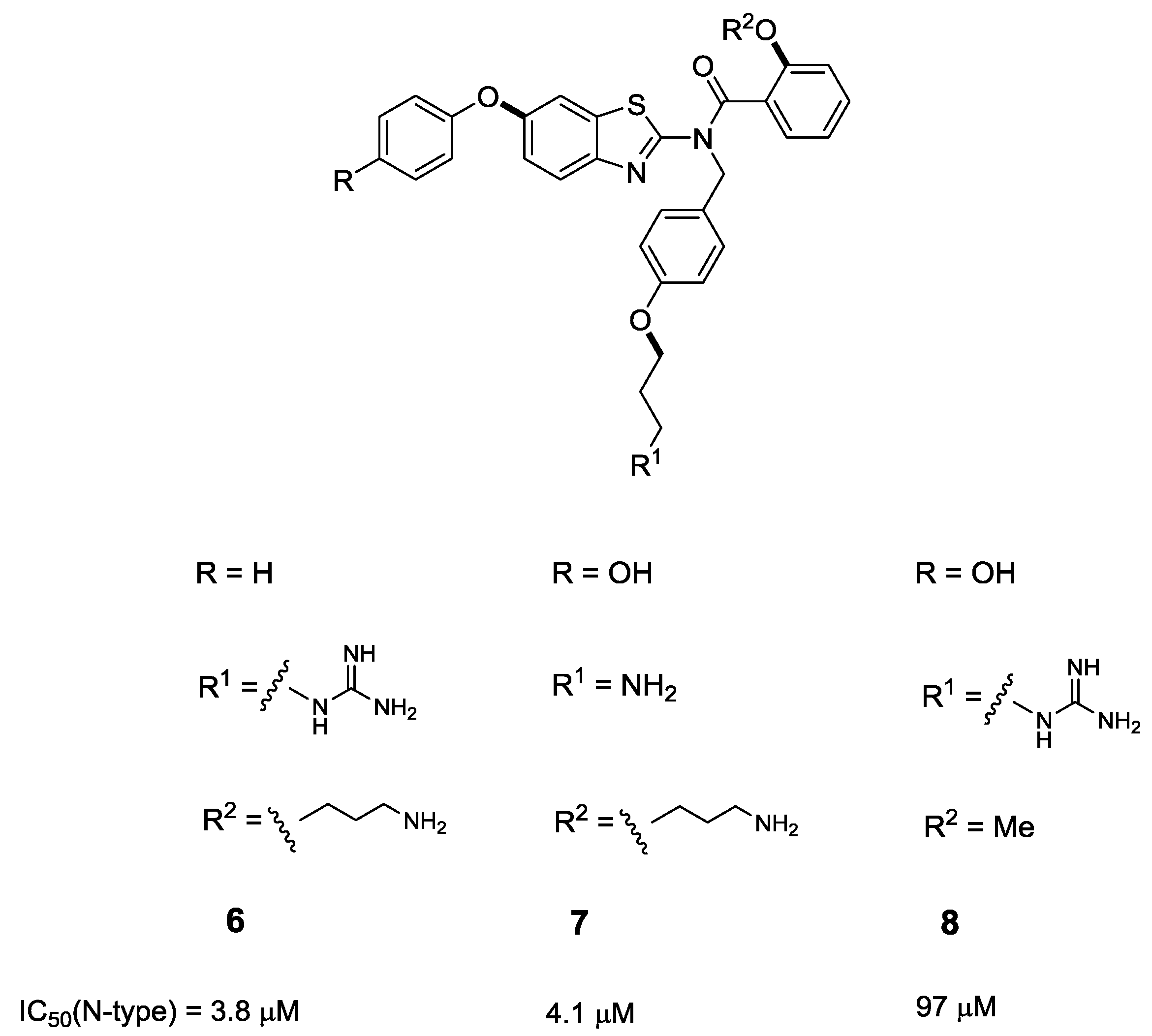

3. Modified Peptides
3.1. Truncated Conotoxins
3.2. Disulfide Isosteres
3.3. Cyclization Strategies
3.4. Backbone Prosthesis
4. Conclusions
Acknowledgments
Conflict of Interest
References
- Lewis, R.J.; Dutertre, S.; Vetter, I.; Christie, M.J. Conus venom peptide pharmacology. Pharm. Rev. 2012, 64, 259–298. [Google Scholar] [CrossRef]
- Olivera, B.M.; Hillyard, D.R.; Marsh, M.; Yoshikami, D. Combinatorial peptide libraries in drug design—Lessons from venomous cone snails. Trends Biotechnol. 1995, 13, 422–426. [Google Scholar] [CrossRef]
- Livett, B.G.; Gayler, K.R.; Khalil, Z. Drugs from the sea: Conopeptides as potential therapeutics. Curr. Med. Chem. 2004, 11, 1715–1723. [Google Scholar] [CrossRef]
- French, R.J.; Terlau, H. Sodium channel toxins—Receptor targeting and therapeutic potential. Curr. Med. Chem. 2004, 11, 3053–3064. [Google Scholar] [CrossRef]
- Norton, R.S.; Olivera, B.M. Conotoxins down under. Toxicon 2006, 48, 780–798. [Google Scholar] [CrossRef]
- Olivera, B.M.; Rivier, J.; Clark, C.; Ramilo, C.A.; Corpuz, G.P.; Abogadie, F.C.; Mena, E.E.; Woodward, S.R.; Hillyard, D.R.; Cruz, L.J. Diversity of Conus neuropeptides. Science 1990, 249, 257–263. [Google Scholar]
- Terlau, H.; Olivera, B.M. Conus venoms: A rich source of novel ion channel-targeted peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef]
- Layer, R.T.; McIntosh, J.M. Conotoxins: Therapeutic potential and application. Mar. Drugs 2006, 4, 119–142. [Google Scholar] [CrossRef]
- Olivera, B.M.; Cruz, L.J. Conotoxins, in retrospect. Toxicon 2001, 39, 7–14. [Google Scholar] [CrossRef]
- Norton, R.S. μ-Conotoxins as leads in the development of new analgesics. Molecules 2010, 15, 2825–2844. [Google Scholar] [CrossRef]
- Alewood, P.; Hopping, G.; Armishaw, C. Marine toxins as sources of drug leads. Aust. J. Chem. 2003, 56, 769–774. [Google Scholar] [CrossRef]
- Kaas, Q.; Yu, R.L.; Jin, A.H.; Dutertre, S.; Craik, D.J. Conoserver: Updated content, knowledge, and discovery tools in the conopeptide database. Nucl. Acids. Res. 2012, 40, D325–D330. [Google Scholar] [CrossRef]
- Violette, A.; Biass, D.; Dutertre, S.; Koua, D.; Piquemal, D.; Pierrat, F.; Stocklin, R.; Favreau, P. Large-Scale discovery of conopeptides and conoproteins in the injectable venom of a fish-hunting cone snail using a combined proteomic and transcriptomic approach. J. Proteomics 2012, 75, 5215–5225. [Google Scholar] [CrossRef]
- Hu, H.; Bandyopadhyay, P.K.; Olivera, B.M.; Yandell, M. Elucidation of the molecular envenomation strategy of the cone snail Conus geographus through transcriptome sequencing of its venom duct. BMC Genomics 2012, 13, 284. [Google Scholar]
- Brust, A.; Palant, E.; Croker, D.E.; Colless, B.; Drinkwater, R.; Patterson, B.; Schroeder, C.I.; Wilson, D.; Nielsen, C.K.; Smith, M.T.; et al. χ-Conopeptide pharmacophore development: Toward a novel class of norepinephrine transporter inhibitor (xen2174) for pain. J. Med. Chem. 2009, 52, 6991–7002. [Google Scholar] [CrossRef]
- Carstens, B.B.; Clark, R.J.; Daly, N.L.; Harvey, P.J.; Kaas, Q.; Craik, D.J. Engineering of Conotoxins for the treatment of pain. Curr. Pharm. Des. 2011, 17, 4242–4253. [Google Scholar] [CrossRef]
- Daly, N.L.; Craik, D.J. Conopeptides as novel options for pain management. Drugs Future 2011, 36, 25–32. [Google Scholar]
- Bowersox, S.S.; Luther, R. Pharmacotherapeutic potential of ω-conotoxin MVIIA (snx-111), an N-type neuronal calcium channel blocker found in the venom of Conus magus. Toxicon 1998, 36, 1651–1658. [Google Scholar] [CrossRef]
- Miljanich, G.P. Ziconotide: Neuronal calcium channel blocker for treating severe chronic pain. Curr. Med. Chem. 2004, 11, 3029–3040. [Google Scholar] [CrossRef]
- Kumar, T.R.S.; Soppimath, K.; Nachaegari, S.K. Novel delivery technologies for protein and peptide therapeutics. Curr. Pharm. Biotechnol. 2006, 7, 261–276. [Google Scholar] [CrossRef]
- Hamman, J.H.; Enslin, G.M.; Kotze, A.F. Oral delivery of peptide drugs—Barriers and developments. Biodrugs 2005, 19, 165–177. [Google Scholar] [CrossRef]
- Adessi, C.; Soto, C. Converting a peptide into a drug: Strategies to improve stability and bioavailability. Curr. Med. Chem. 2002, 9, 963–978. [Google Scholar] [CrossRef]
- Ripka, A.S.; Rich, D.H. Peptidomimetic design. Curr. Opin. Chem. Biol. 1998, 2, 441–452. [Google Scholar] [CrossRef]
- Croft, N.P.; Purcell, A.W. Peptidomimetics: Modifying peptides in the pursuit of better vaccines. Expert Rev. Vaccines 2011, 10, 211–226. [Google Scholar] [CrossRef]
- Smith, A.B.; Charnley, A.K.; Hirschmann, R. Pyrrolinone-Based peptidomimetics. “Let the enzyme or receptor be the judge”. Acc. Chem. Res. 2011, 44, 180–193. [Google Scholar] [CrossRef]
- Vagner, J.; Qu, H.C.; Hruby, V.J. Peptidomimetics, a synthetic tool of drug discovery. Curr. Opin. Chem. Biol. 2008, 12, 292–296. [Google Scholar] [CrossRef]
- Norton, R.S.; Pallaghy, P.K.; Baell, J.B.; Wright, C.E.; Lew, M.J.; Angus, J.A. Polypeptide ω-conotoxin GVIA as a basis for new analgesic and neuroprotective agents. Drug Dev. Res. 1999, 46, 206–218. [Google Scholar] [CrossRef]
- Norton, R.S.; McDonough, S.I. Peptides targeting voltage-gated calcium channels. Curr. Pharm. Des. 2008, 14, 2480–2491. [Google Scholar] [CrossRef]
- Menzler, S.; Bikker, J.A.; Suman-Chauhan, N.; Horwell, D.C. Design and biological evaluation of non-peptide analogues of ω-conotoxin MVIIA. Bioorg. Med. Chem. Lett. 2000, 10, 345–347. [Google Scholar]
- Olivera, B.M.; Gray, W.R.; Zeikus, R.; McIntosh, J.M.; Varga, J.; Rivier, J.; Desantos, V.; Cruz, L.J. Peptide neurotoxins from fish-hunting cone snails. Science 1985, 230, 1338–1343. [Google Scholar]
- Olivera, B.M.; Rivier, J.; Scott, J.K.; Hillyard, D.R.; Cruz, L.J. Conotoxins. J. Biol. Chem. 1991, 266, 22067–22070. [Google Scholar]
- Guo, Z.X.; Cammidge, A.N.; Horwell, D.C. Dendroid peptide structural mimetics of ω-conotoxin MVIIA based on a 2(1H)-quinolinone core. Tetrahedron 2000, 56, 5169–5175. [Google Scholar] [CrossRef]
- Menzler, S.; Bikker, J.A.; Horwell, D.C. Synthesis of a non-peptide analogue of ω-conotoxin MVIIA. Tetrahedron Lett. 1998, 39, 7619–7622. [Google Scholar] [CrossRef]
- McCleskey, E.W.; Fox, A.P.; Feldman, D.H.; Cruz, L.J.; Olivera, B.M.; Tsien, R.W.; Yoshikami, D. ω-Conotoxin—Direct and persistent blockade of specific types of calcium channels in neurons but not muscle. Proc. Natl. Acad. Sci. USA 1987, 84, 4327–4331. [Google Scholar] [CrossRef]
- Olivera, B.M.; McIntosh, J.M.; Cruz, L.J.; Luque, F.A.; Gray, W.R. Purification and sequence of A presynaptic peptide toxin from Conus geographus venom. Biochemistry 1984, 23, 5087–5090. [Google Scholar] [CrossRef]
- Lew, M.J.; Flinn, J.P.; Pallaghy, P.K.; Murphy, R.; Whorlow, S.L.; Wright, C.E.; Norton, R.S.; Angus, J.A. Structure-function relationships of ω-conotoxin GVIA—Synthesis, structure, calcium channel binding, and functional assay of alanine-substituted analogues. J. Biol. Chem. 1997, 272, 12014–12023. [Google Scholar] [CrossRef]
- Flinn, J.P.; Pallaghy, P.K.; Lew, M.J.; Murphy, R.; Angus, J.A.; Norton, R.S. Roles of key functional groups in ω-conotoxin GVIA—Synthesis, structure and functional assay of selected peptide analogues. Eur. J. Biochem. 1999, 262, 447–455. [Google Scholar] [CrossRef]
- Baell, J.B.; Forsyth, S.A.; Gable, R.W.; Norton, R.S.; Mulder, R.J. Design and synthesis of type-III mimetics of omega-conotoxin GVIA. J. Comput. Aided Mol. Des. 2001, 15, 1119–1136. [Google Scholar] [CrossRef]
- Baell, J.B.; Duggan, P.J.; Forsyth, S.A.; Lewis, R.J.; Lok, Y.P.; Schroeder, C.I. Synthesis and biological evaluation of nonpeptide mimetics of ω-conotoxin GVIA. Bioorg. Med. Chem. 2004, 12, 4025–4037. [Google Scholar] [CrossRef]
- Duggan, P.J.; Lewis, R.J.; Lok, Y.P.; Lumsden, N.G.; Tuck, K.L.; Yang, A.J. Low molecular weight non-peptide mimics of ω-conotoxin GVIA. Bioorg. Med. Chem. Lett. 2009, 19, 2763–2765. [Google Scholar] [CrossRef]
- Baell, J.B.; Duggan, P.J.; Forsyth, S.A.; Lewis, R.J.; Lok, Y.P.; Schroeder, C.I.; Shepherd, N.E. Synthesis and biological evaluation of anthranilamide-based non-peptide mimetics of ω-conotoxin GVIA. Tetrahedron 2006, 62, 7284–7292. [Google Scholar]
- Andersson, A.; Baell, J.B.; Duggan, P.J.; Graham, J.E.; Lewis, R.J.; Lumsden, N.G.; Tranberg, C.E.; Tuck, K.L.; Yang, A.J. ω-Conotoxin GVIA mimetics based on an anthranilamide core: Effect of variation in ammonium side chain lengths and incorporation of fluorine. Bioorg. Med. Chem. 2009, 17, 6659–6670. [Google Scholar] [CrossRef]
- Duggan, P.J.; Faber, J.M.; Graham, J.E.; Lewis, R.J.; Lumsden, N.G.; Tuck, K.L. Synthesis and Ca(v)2.2 binding data for non-peptide mimetics of ω-conotoxin GVIA based on a 5-amino-anthranilamide core. Aust. J. Chem. 2008, 61, 11–15. [Google Scholar] [CrossRef]
- Tranberg, C.E.; Yang, A.J.; Vetter, I.; McArthur, J.R.; Baell, J.B.; Lewis, R.J.; Tuck, K.L.; Duggan, P.J. ω-Conotoxin GVIA mimetics that bind and inhibit neuronal Ca(v)2.2 ion channels. Mar. Drugs 2012, 10, 2349–2368. [Google Scholar] [CrossRef]
- Zamponi, G.W.; Feng, Z.P.; Zhang, L.Y.; Pajouhesh, H.; Ding, Y.B.; Belardetti, F.; Dolphin, D.; Mitscher, L.A.; Snutch, T.P. Scaffold-based design and synthesis of potent N-type calcium channel blockers. Bioorg. Med. Chem. Lett. 2009, 19, 6467–6472. [Google Scholar] [CrossRef]
- Pajouhesh, H.; Feng, Z.P.; Ding, Y.B.; Zhang, L.Y.; Morrison, J.L.; Belardetti, F.; Tringham, E.; Simonson, E.; Vanderah, T.W.; Porreca, F.; et al. Structure-activity relationships of diphenylpiperazine N-type calcium channel inhibitors. Bioorg. Med. Chem. Lett. 2010, 20, 1378–1383. [Google Scholar] [CrossRef]
- Scott, V.E.; Vortherms, T.A.; Niforatos, W.; Swensen, A.M.; Neelands, T.; Milicic, I.; Banfor, P.N.; King, A.; Zhong, C.M.; Simler, G.; et al. A-1048400 is a novel, orally active, state-dependent neuronal calcium channel blocker that produces dose-dependent antinociception without altering hemodynamic function in rats. Biochem. Pharmacol. 2012, 83, 406–418. [Google Scholar]
- Doherty, G.A.; Bhatia, P.; Vortherms, T.A.; Marsh, K.C.; Wetter, J.M.; Mack, H.; Scott, V.E.; Jarvis, M.F.; Stewart, A.O. Discovery of diphenyl lactam derivatives as N-type calcium channel blockers. Bioorg. Med. Chem. Lett. 2012, 22, 1716–1718. [Google Scholar] [CrossRef]
- Bulaj, G.; West, P.J.; Garrett, J.E.; Marsh, M.; Zhang, M.M.; Norton, R.S.; Smith, B.J.; Yoshikami, D.; Olivera, B.M. Novel conotoxins from Conus striatus and Conus kinoshitai selectively block TTX-resistant sodium channels. Biochemistry 2005, 44, 7259–7265. [Google Scholar] [CrossRef]
- Knapp, O.; McArthur, J.R.; Adams, D.J. Conotoxins targeting neuronal voltage-gated sodium channel subtypes: Potential analgesics? Toxins (Basel) 2012, 4, 1236–1260. [Google Scholar] [CrossRef]
- Zhang, M.-M.; Green, B.R.; Catlin, P.; Fiedler, B.; Azam, L.; Chadwick, A.; Terlau, H.; McArthur, J.R.; French, R.J.; Gulyas, J.; et al. Structure/function characterization of μ-conotoxin KIIIA, an analgesic, nearly irreversible blocker of mammalian neuronal sodium channels. J. Biol. Chem. 2007, 282, 30699–30706. [Google Scholar] [CrossRef]
- McArthur, J.R.; Singh, G.; McMaster, D.; Winkfein, R.; Tieleman, D.P.; French, R.J. Interactions of key charged residues contributing to selective block of neuronal sodium channels by μ-conotoxin KIIIA. Mol. Pharmacol. 2011, 80, 573–584. [Google Scholar] [CrossRef]
- Khoo, K.K.; Feng, Z.-P.; Smith, B.J.; Zhang, M.-M.; Yoshikami, D.; Olivera, B.M.; Bulaj, G.; Norton, R.S. Structure of the analgesic μ-conotoxin KIIIA and effects on the structure and function of disulfide deletion. Biochemistry 2009, 48, 1210–1219. [Google Scholar] [CrossRef]
- Khoo, K.K.; Gupta, K.; Green, B.R.; Zhang, M.M.; Watkins, M.; Olivera, B.M.; Balaram, P.; Yoshikami, D.; Bulaj, G.; Norton, R.S. Distinct disulfide isomers of μ-conotoxins KIIIA and KIIIB block voltage-gated sodium channels. Biochemistry 2012, 51, 9826–9835. [Google Scholar] [CrossRef]
- Brady, R.M.; Zhang, M.M.; Gable, R.B.; Norton, R.S.; Baell, J.B. De novo design and synthesis of a μ-KIIIA peptidomimetic. Bioorg. Med. Chem. Lett. 2013, in press. [Google Scholar]
- Lessene, G.; Smith, B.J.; Gable, R.W.; Baell, J.B. Characterization of the two fundamental conformations of benzoylureas and elucidation of the factors that facilitate their conformational interchange. J. Org. Chem. 2009, 74, 6511–6525. [Google Scholar] [CrossRef]
- Herbert, R.H.; Kelleher, F. The solution conformation of 1-(3,5-dimethylphenyl)methyl-3(S)-(1H-indol-3-yl)methyl-6(S)-phenylmethyl-2,5-piperazinedione(1)—An NMR and molecular modeling study. Tetrahedron Lett. 1994, 35, 5497–5500. [Google Scholar] [CrossRef]
- Kopple, K.D.; Marr, D.H. Conformations of cyclic peptides. Folding of cyclic dipeptides containing an aromatic side chain. J. Am. Chem. Soc. 1967, 89, 6193–6200. [Google Scholar] [CrossRef]
- Babine, R.E.; Bender, S.L. Molecular recognition of protein-ligand complexes: Applications to drug design. Chem. Rev. 1997, 97, 1359–1472. [Google Scholar] [CrossRef]
- Brady, R.M.; Tae, H.; Adams, D.J.; Zhang, M.M.; Norton, R.S.; Baell, J.B. Monash Institute of Pharmaceutical Science. Monash University: Parkville 3052, Australia, Unpublished work. 2013. [Google Scholar]
- Jin, A.H.; Daly, N.L.; Nevin, S.T.; Wang, C.I.A.; Dutertre, S.; Lewis, R.J.; Adams, D.J.; Craik, D.J.; Alewood, P.F. Molecular engineering of conotoxins: The importance of loop size to α-conotoxin structure and function. J. Med. Chem. 2008, 51, 5575–5584. [Google Scholar] [CrossRef]
- Sine, S.M.; Kreienkamp, H.J.; Bren, N.; Maeda, R.; Taylor, P. Molecular dissection of subunit interfaces in the acetylcholine-receptor—Identification of determinants of α-conotoxin M1 selectivity. Neuron 1995, 15, 205–211. [Google Scholar]
- Hu, S.H.; Gehrmann, J.; Guddat, L.W.; Alewood, P.F.; Craik, D.J.; Martin, J.L. The 1.1 angstrom crystal structure of the neuronal acetylcholine receptor antagonist, α-conotoxin PnIA from Conus pennaceus. Structure 1996, 4, 417–423. [Google Scholar]
- Kirby, D.A.; Britton, K.T.; Aubert, M.L.; Rivier, J.E. Identification of high-potency neuropeptide Y analogues through systematic lactamization. J. Med. Chem. 1997, 40, 210–215. [Google Scholar]
- Yao, S.G.; Smith-White, M.A.; Potter, E.K.; Norton, R.S. Stabilization of the helical structure of Y2-selective analogues of neuropeptide Y by lactam bridges. J. Med. Chem. 2002, 45, 2310–2318. [Google Scholar]
- Yang, B.; Liu, D.X.; Huang, Z.W. Synthesis and helical structure of lactam bridged BH3 peptides derived from pro-apoptotic Bcl-2 family proteins. Bioorg. Med. Chem. Lett. 2004, 14, 1403–1406. [Google Scholar] [CrossRef]
- Shepherd, N.E.; Abbenante, G.; Fairlie, D.P. Consecutive cyclic pentapeptide modules form short α-helices that are very stable to water and denaturants. Angew. Chem. Int. Ed. 2004, 43, 2687–2690. [Google Scholar] [CrossRef]
- Shepherd, N.E.; Hoang, H.N.; Abbenante, G.; Fairlie, D.P. Left- and right-handed α-helical turns in homo- and hetero-chiral helical scaffolds. J. Am. Chem. Soc. 2009, 131, 15877–15886. [Google Scholar]
- Lanigan, M.D.; Pennington, M.W.; Lefievere, Y.; Rauer, H.; Norton, R.S. Designed peptide analogues of the potassium channel blocker ShK toxin. Biochemistry 2001, 40, 15528–15537. [Google Scholar]
- Khoo, K.K.; Wilson, M.J.; Smith, B.J.; Zhang, M.M.; Gulyas, J.; Yoshikami, D.; Rivier, J.E.; Bulaj, G.; Norton, R.S. Lactam-stabilized helical analogues of the analgesic μ-conotoxin KIIIA. J. Med. Chem. 2011, 54, 7558–7566. [Google Scholar]
- Zhang, M.M.; Han, T.S.; Olivera, B.M.; Bulaj, G.; Yoshikami, D. μ-Conotoxin KIIIA derivatives with divergent affinities versus efficacies in blocking voltage-gated sodium channels. Biochemistry 2010, 49, 4804–4812. [Google Scholar]
- Holford, M.; Zhang, M.M.; Gowd, K.H.; Azam, L.; Green, B.R.; Watkins, M.; Ownby, J.P.; Yoshikami, D.; Bulaj, G.; Olivera, B.M. Pruning nature: Biodiversity-derived discovery of novel sodium channel blocking conotoxins from Conus bullatus. Toxicon 2009, 53, 90–98. [Google Scholar]
- Stevens, M.; Peigneur, S.; Dyubankova, N.; Lescrinier, E.; Herdewijn, P.; Tytgat, J. Design of bioactive peptides from naturally occurring μ-conotoxin structures. J. Biol. Chem. 2012, 287, 31382–31392. [Google Scholar]
- Khoo, K.K.; Norton, R.S. Role of Disulfide Bonds in Peptide and Protein Conformation. In Amino Acids, Peptides and Proteins in Organic Chemistry; Wiley-VCH Verlag: Weinheim, Germany; Volume 5, pp. 395–417.
- Muttenthaler, M.; Andersson, A.; de Araujo, A.D.; Dekan, Z.; Lewis, R.J.; Alewood, P.F. Modulating oxytocin activity and plasma stability by disulfide bond engineering. J. Med. Chem. 2010, 53, 8585–8596. [Google Scholar]
- Armishaw, C.J.; Daly, N.L.; Nevin, S.T.; Adams, D.J.; Craik, D.J.; Alewood, P.F. α-Selenoconotoxins, a new class of potent α7 neuronal nicotinic receptor antagonists. J. Biol. Chem. 2006, 281, 14136–14143. [Google Scholar]
- Stymiest, J.L.; Mitchell, B.F.; Wong, S.; Vederas, J.C. Synthesis of biologically active dicarba analogues of the peptide hormone oxytocin using ring-closing metathesis. Org. Lett. 2003, 5, 47–49. [Google Scholar] [CrossRef]
- Bondebjerg, J.; Grunnet, M.; Jespersen, T.; Meldal, M. Solid-Phase synthesis and biological activity of a thioether analogue of conotoxin G1. ChemBioChem 2003, 4, 186–194. [Google Scholar]
- Galande, A.K.; Bramlett, K.S.; Burris, T.P.; Wittliff, J.L.; Spatola, A.F. Thioether side chain cyclization for helical peptide formation: inhibitors of estrogen receptor-coactivator interactions. J. Peptide Res. 2004, 63, 297–302. [Google Scholar]
- Rew, Y.; Malkmus, S.; Svensson, C.; Yaksh, T.L.; Chung, N.N.; Schiller, P.W.; Cassel, J.A.; DeHaven, R.N.; Goodman, M. Synthesis and biological activities of cyclic lanthionine enkephalin analogues: δ-Opioid receptor selective ligands. J. Med. Chem. 2002, 45, 3746–3754. [Google Scholar]
- Blackwell, H.E.; Sadowsky, J.D.; Howard, R.J.; Sampson, J.N.; Chao, J.A.; Steinmetz, W.E.; O’Leary, D.J.; Grubbs, R.H. Ring-closing metathesis of olefinic peptides: Design, synthesis, and structural characterization of macrocyclic helical peptides. J. Org. Chem. 2001, 66, 5291–5302. [Google Scholar] [CrossRef]
- Khan, S.N.; Kim, A.; Grubbs, R.H.; Kwon, Y.U. Ring-Closing metathesis approaches for the solid-phase synthesis of cyclic peptoids. Org. Lett. 2011, 13, 1582–1585. [Google Scholar] [CrossRef]
- Miller, S.J.; Blackwell, H.E.; Grubbs, R.H. Application of ring-closing metathesis to the synthesis of rigidified amino acids and peptides. J. Am. Chem. Soc. 1996, 118, 9606–9614. [Google Scholar]
- Miller, S.J.; Grubbs, R.H. Synthesis of conformationally restricted amino-acids and peptides employing olefin metathesis. J. Am. Chem. Soc. 1995, 117, 5855–5856. [Google Scholar] [CrossRef]
- Robinson, A.J.; Elaridi, J.; van Lierop, B.J.; Mujcinovic, S.; Jackson, W.R. Microwave-assisted RCM for the synthesis of carbocyclic peptides. J. Peptide Sci. 2007, 13, 280–285. [Google Scholar]
- Robinson, A.J.; van Lierop, B.J.; Garland, R.D.; Teoh, E.; Elaridi, J.; Illesinghe, J.P.; Jackson, W.R. Regioselective formation of interlocked dicarba bridges in naturally occurring cyclic peptide toxins using olefin metathesis. Chem. Commun. 2009, 28, 4293–4295. [Google Scholar]
- Van Lierop, B.J.; Bornschein, C.; Jackson, W.R.; Robinson, A.J. Ring-closing metathesis in peptides—The sting is in the tail! Aust. J. Chem. 2011, 64, 806–811. [Google Scholar]
- Richardson, J.S. The anatomy and taxonomy of protein structure. Adv. Protein Chem. 1981, 34, 167–339. [Google Scholar]
- Hase, S.; Morikawa, T.; Sakakiba, S. Synthesis of A biologically active analog of deamino-8-arginine-vasopressin which does not contain a disulfide bond. Experientia 1969, 25, 1239–1240. [Google Scholar] [CrossRef]
- Kambayashi, Y.; Nakajima, S.; Ueda, M.; Inouye, K. A dicarba analog of β-atrial natriuretic peptide (β-anp) inhibits guanosine 3′,5′-cyclic-monophosphate production induced by α-anp in cultured rat vascular smooth-muscle cells. FEBS Lett. 1989, 248, 28–34. [Google Scholar] [CrossRef]
- Oka, T.; Nakanishi, A.; Okada, T. Studies on pharmacological and biochemical properties of deamino-dicarba-[Gly7]-oxytocin (Y-5350). Jpn. J. Pharmacol. 1975, 25, 15–24. [Google Scholar] [CrossRef]
- Stymiest, J.L.; Mitchell, B.F.; Wong, S.; Vederas, J.C. Synthesis of oxytocin analogues with replacement of sulfur by carbon gives potent antagonists with increased stability. J. Org. Chem. 2005, 70, 7799–7809. [Google Scholar] [CrossRef]
- Knerr, P.J.; Tzekou, A.; Ricklin, D.; Qu, H.C.; Chen, H.; van der Donk, W.A.; Lambris, J.D. Synthesis and activity of thioether-containing analogues of the complement inhibitor compstatin. ACS Chem. Biol. 2011, 6, 753–760. [Google Scholar]
- McIntosh, J.M.; Yoshikami, D.; Mahe, E.; Nielsen, D.B.; Rivier, J.E.; Gray, W.R.; Olivera, B.M. A nicotinic acetylcholine-receptor ligand of unique specificity, α-conotoxin ImI. J. Biol. Chem. 1994, 269, 16733–16739. [Google Scholar]
- MacRaild, C.A.; Illesinghe, J.; van Lierop, B.J.; Townsend, A.L.; Chebib, M.; Livett, B.G.; Robinson, A.J.; Norton, R.S. Structure and activity of (2,8)-dicarba-(3,12)-cystino α-ImI, an α-conotoxin containing a nonreducible cystine analogue. J. Med. Chem. 2009, 52, 755–762. [Google Scholar] [CrossRef]
- Robinson, A.J. Personal communication, Monash University: Clayton 3800, Australia, 2013.
- Van Lierop, B.J.; Robinson, S.D.; Kompella, S.N.; McArthur, J.R.; Hung, A.; MacRaild, C.A.; Adams, D.J.; Norton, R.S.; Robinson, A.J. Dicarba α-conotoxin Vc1.1 analogues with differential selectivity for nicotinic acetylcholine and GABAB receptors. ACS Chem. Biol. 2013, in press. [Google Scholar]
- Dekan, Z.; Vetter, I.; Daly, N.L.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. α-Conotoxin ImI incorporating stable cystathionine bridges maintains full potency and identical three-dimensional structure. J. Am. Chem. Soc. 2011, 133, 15866–15869. [Google Scholar] [CrossRef]
- Bock, J.E.; Gavenonis, J.; Kritzer, J.A. Getting in shape: Controlling peptide bioactivity and bioavailability using conformational constraints. ACS Chem. Biol. 2013, 8, 488–499. [Google Scholar]
- Adessi, C.; Soto, C. Strategies to improve stability and bioavailability of peptide drugs. Front. Med. Chem.Online 2004, 1, 513–528. [Google Scholar] [CrossRef]
- Craik, D.J.; Adams, D.J. Chemical modification of conotoxins to improve stability and activity. ACS Chem. Biol. 2007, 2, 457–468. [Google Scholar]
- Boger, D.L. Vancomycin, teicoplanin, and ramoplanin: Synthetic and mechanistic studies. Med. Res. Rev. 2001, 21, 356–381. [Google Scholar] [CrossRef]
- Boger, D.L.; Kim, S.H.; Mori, Y.; Weng, J.H.; Rogel, O.; Castle, S.L.; McAtee, J.J. First and second generation total synthesis of the teicoplanin aglycon. J. Am. Chem. Soc. 2001, 123, 1862–1871. [Google Scholar]
- Borel, J.F. Mechanism of action of cyclosporine-A and rationale for use in nephrotic syndrome. Clin. Nephrol. 1991, 35, S23–S30. [Google Scholar]
- Allison, S.J. Basic research: An oral cyclic peptide drug to reverse kidney fibrosis? Nat. Rev. Nephrol. 2012, 8, 193–193. [Google Scholar]
- Clark, R.J.; Akcan, M.; Kaas, Q.; Daly, N.L.; Craik, D.J. Cyclization of conotoxins to improve their biopharmaceutical properties. Toxicon 2012, 59, 446–455. [Google Scholar] [CrossRef]
- Clark, R.J.; Jensen, J.; Nevin, S.T.; Callaghan, B.P.; Adams, D.J.; Craik, D.J. The engineering of an orally active conotoxin for the treatment of neuropathic pain. Angew. Chem. Int. Ed. 2010, 49, 6545–6548. [Google Scholar]
- Clark, R.J.; Fischer, H.; Nevin, S.T.; Adams, D.J.; Craik, D.J. The synthesis, structural characterization, and receptor specificity of the α-conotoxin Vc1.1. J. Biol. Chem. 2006, 281, 23254–23263. [Google Scholar] [CrossRef]
- Satkunanathan, N.; Livett, B.; Gayler, K.; Sandall, D.; Down, J.; Khalil, Z. α-Conotoxin Vc1.1 alleviates neuropathic pain and accelerates functional recovery of injured neurones. Brain Res. 2005, 1059, 149–158. [Google Scholar] [CrossRef]
- Vincler, M.; Wittenauer, S.; Parker, R.; Ellison, M.; Olivera, B.M.; McIntosh, J.M. Molecular mechanism for analgesia involving specific antagonism of α9, α10 nicotinic acetylcholine receptors. Proc. Natl. Acad. Sci. USA 2006, 103, 17880–17884. [Google Scholar]
- Vincler, M.; McIntosh, J.M. Targeting the α9, α10 nicotinic acetylcholine receptor to treat severe pain. Expert Opin. Ther. 2007, 11, 891–897. [Google Scholar]
- Callaghan, B.; Haythornthwaite, A.; Berecki, G.; Clark, R.J.; Craik, D.J.; Adams, D.J. Analgesic α-conotoxins Vc1.1 and RgIA inhibit N-type calcium channels in rat sensory neurons via GABAB receptor activation. J. Neurosci. 2008, 28, 10943–10951. [Google Scholar] [CrossRef]
- Callaghan, B.; Adams, D.J. Analgesic α-conotoxins Vc1.1 and RgIA inhibit N-type calcium channels in sensory neurons of α9 nicotinic receptor knockout mice. Channels (Austin) 2010, 4, 51–54. [Google Scholar]
- Cuny, H.; de Faoite, A.; Huynh, T.G.; Yasuda, T.; Berecki, G.; Adams, D.J. γ-Aminobutyric acid type B (GABAB) receptor expression is needed for inhibition of N-type (Cav2.2) calcium channels by analgesic α-conotoxins. J. Biol. Chem. 2012, 287, 23948–23957. [Google Scholar]
- Green, B.R.; Catlin, P.; Zhang, M.M.; Fiedler, B.; Bayudan, W.; Morrison, A.; Norton, R.S.; Smith, B.J.; Yoshikami, D.; Olivera, B.M.; et al. Conotoxins containing nonnatural backbone spacers: Cladistic-based design, chemical synthesis, and improved analgesic activity. Chem. Biol. 2007, 14, 399–407. [Google Scholar]
- Veronese, F.M.; Pasut, G. PEGylation, successful approach to drug delivery. Drug Discov. Today 2005, 10, 1451–1458. [Google Scholar] [CrossRef]
- Doyle, M.E.; Greig, N.H.; Holloway, H.W.; Betkey, J.A.; Bernier, M.; Egan, J.M. Insertion of an N-terminal 6-aminohexanoic acid after the 7 amino acid position of glucagon-like peptide-1 produces a long-acting hypoglycemic agent. Endocrinology 2001, 142, 4462–4468. [Google Scholar] [CrossRef]
- Langer, M.; la Bella, R.; Garcia-Garayoa, E.; Beck-Sickinger, A.G. Tc-99m-labeled neuropeptide Y analogues as potential tumor imaging agents. Bioconjugate Chem. 2001, 12, 1028–1034. [Google Scholar] [CrossRef]
- Rist, B.; Wieland, H.A.; Willim, K.D.; Beck-Sickinger, A.G. A rational approach for the development of reduced-size analogues of neuropeptide Y with high affinity to the Y-1 receptor. J. Peptide Sci. 1995, 1, 341–348. [Google Scholar]
- Wang, C.Z.; Zhang, H.; Jiang, H.; Lu, W.Y.; Zhao, Z.Q.; Chi, C.W. A novel conotoxin from Conus striatus, μ-SIIIA, selectively blocking rat tetrodotoxin-resistant sodium channels. Toxicon 2006, 47, 122–132. [Google Scholar] [CrossRef]
- Hunskaar, S.; Hole, K. The formalin test in mice—Dissociation between inflammatory and noninflammatory pain. Pain 1987, 30, 103–114. [Google Scholar] [CrossRef]
- Han, T.S.; Zhang, M.M.; Walewska, A.; Gruszczynski, P.; Robertson, C.R.; Cheatham, T.E.; Yoshikami, D.; Olivera, B.M.; Bulaj, G. Structurally minimized μ-conotoxin analogues as sodium channel blockers: Implications for designing conopeptide-based therapeutics. ChemMedChem 2009, 4, 406–414. [Google Scholar] [CrossRef]
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Brady, R.M.; Baell, J.B.; Norton, R.S. Strategies for the Development of Conotoxins as New Therapeutic Leads. Mar. Drugs 2013, 11, 2293-2313. https://doi.org/10.3390/md11072293
Brady RM, Baell JB, Norton RS. Strategies for the Development of Conotoxins as New Therapeutic Leads. Marine Drugs. 2013; 11(7):2293-2313. https://doi.org/10.3390/md11072293
Chicago/Turabian StyleBrady, Ryan M., Jonathan B. Baell, and Raymond S. Norton. 2013. "Strategies for the Development of Conotoxins as New Therapeutic Leads" Marine Drugs 11, no. 7: 2293-2313. https://doi.org/10.3390/md11072293