A Therapeutic Potential for Marine Skeletal Proteins in Bone Regeneration


 ,
,  and
and 
Abstract
:1. Introduction
Key Issues in Regenerative Orthopaedics
2. The Earliest Proteins and Genes Have Been Safeguarded throughout Animal Evolution
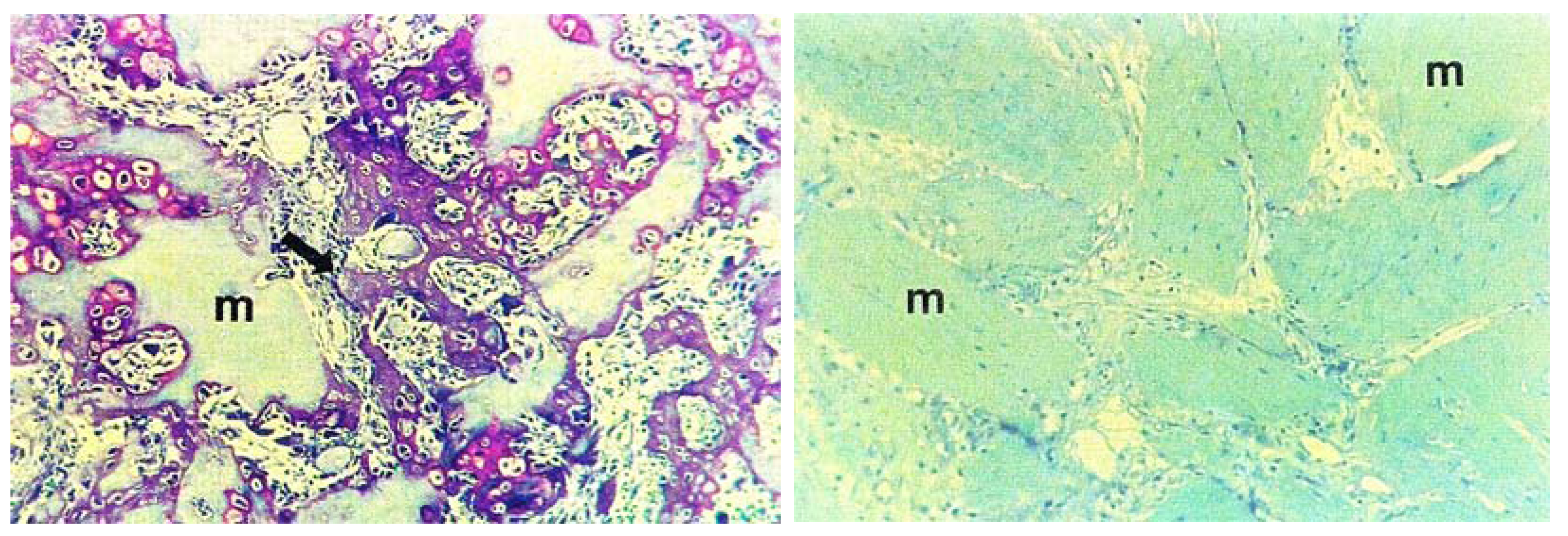
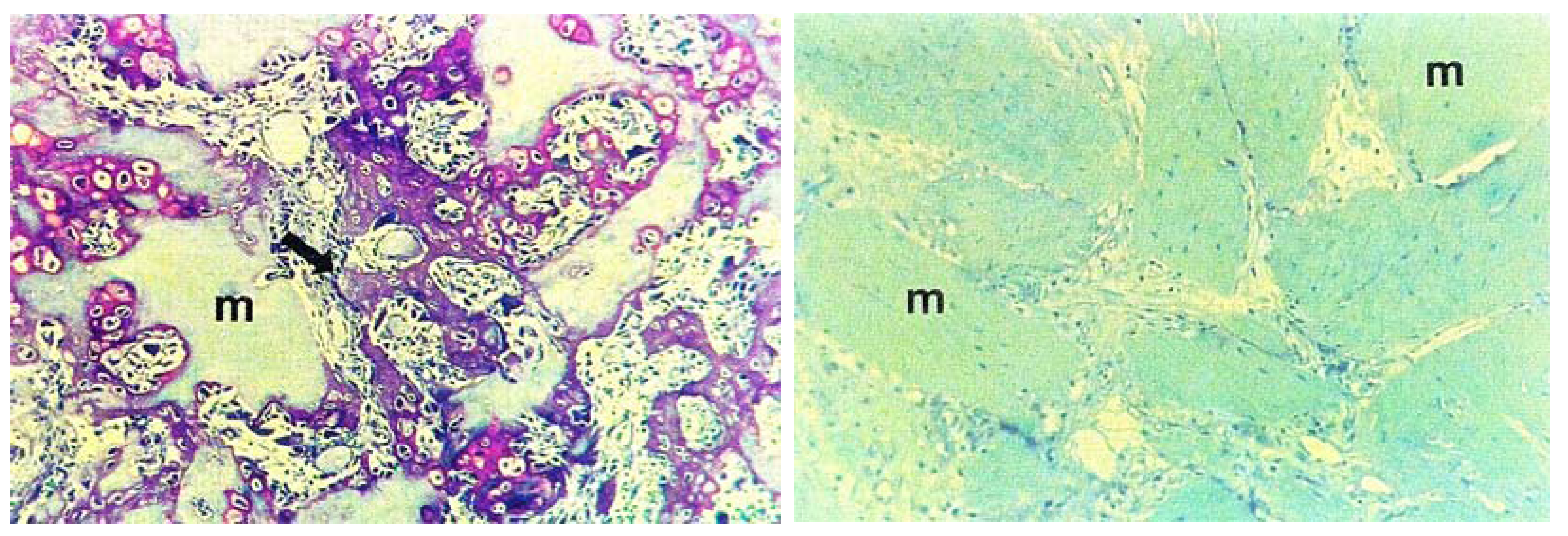
3. Preservation of Proteins during the Evolutionary History of Biomineralization
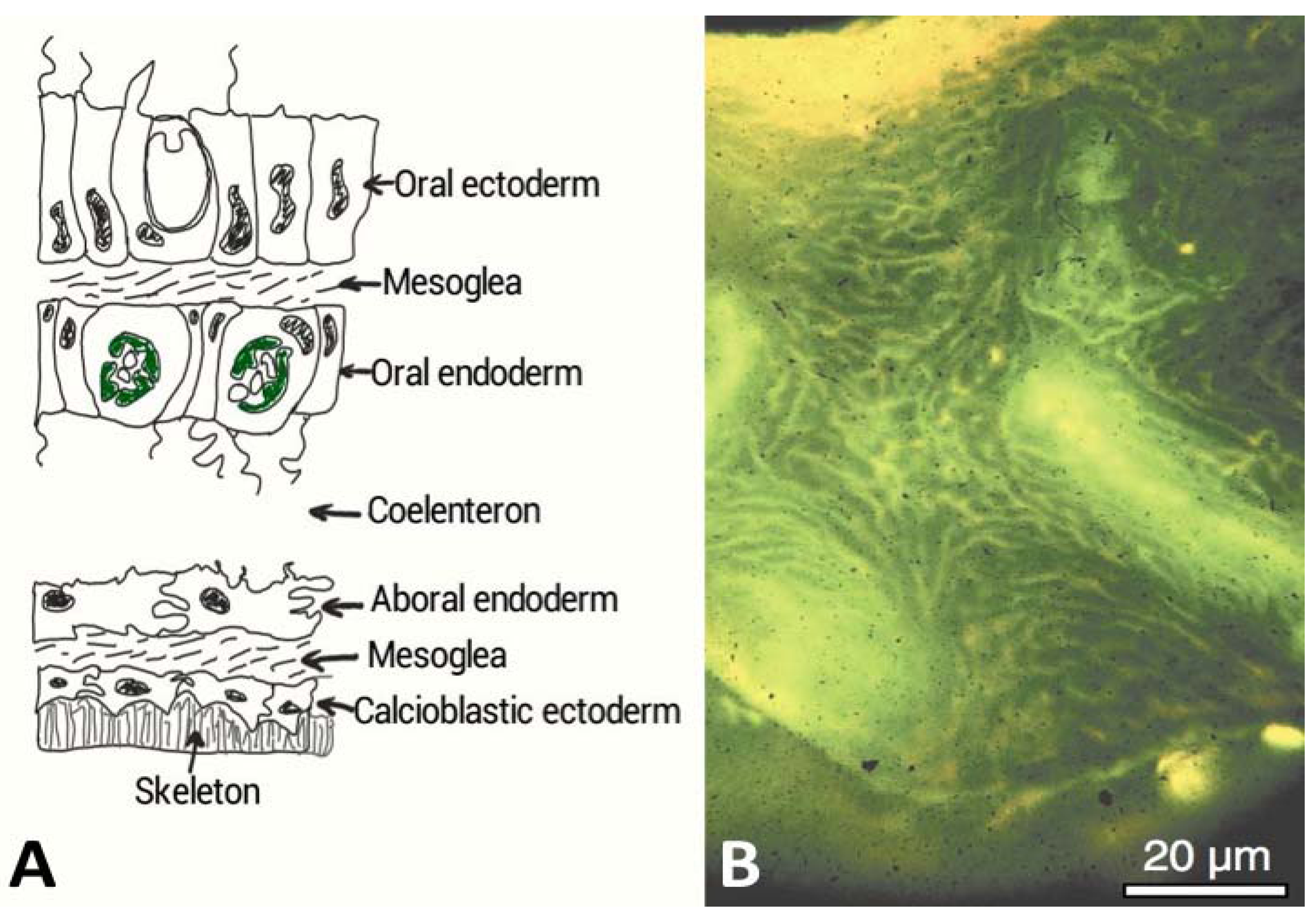
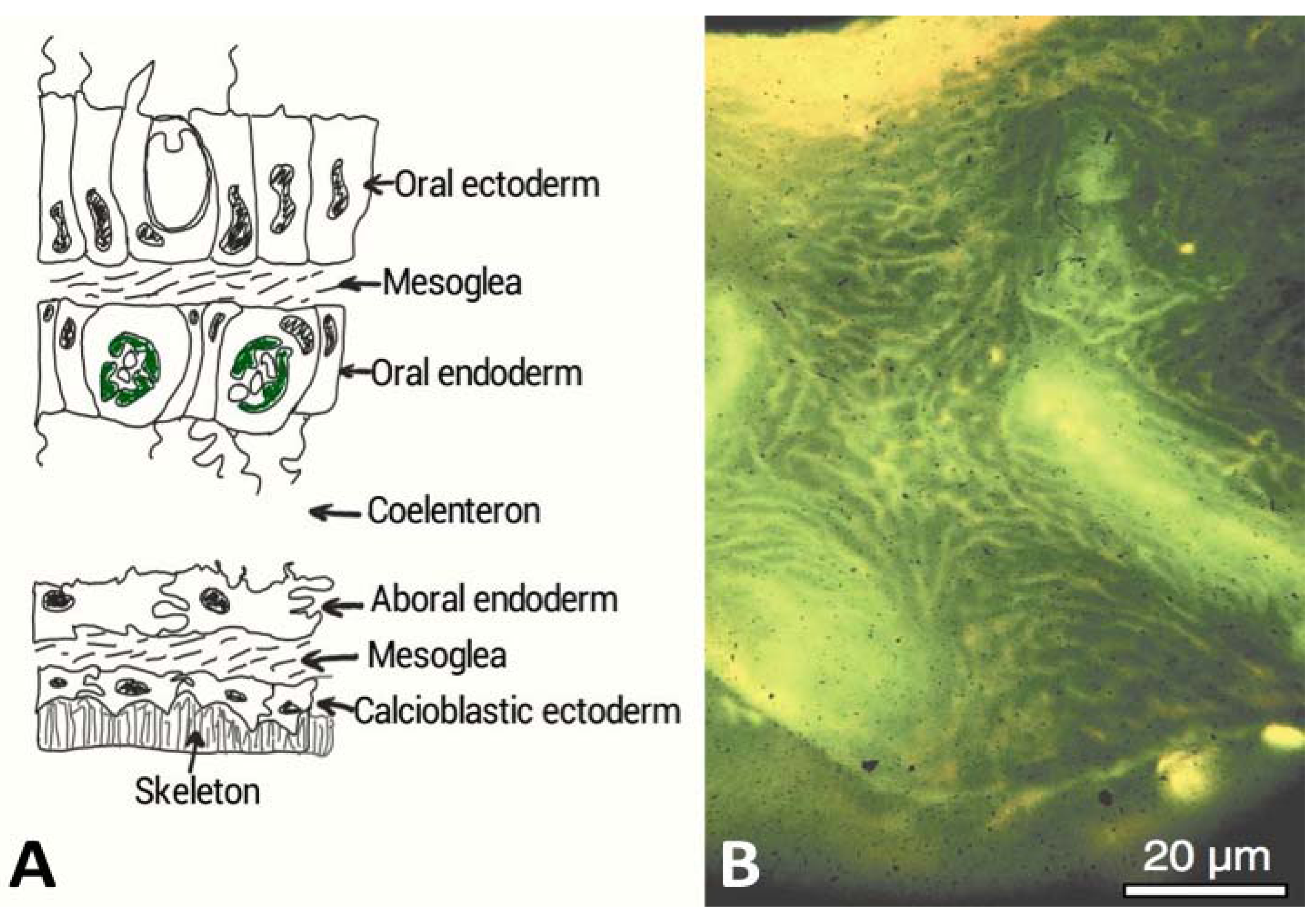
4.Marine Bioprospecting for Skeletal Matrix Proteins
5. Marine Invertebrates Are Sources of Proteins for Regenerative Orthopaedics

{kind=link}
{kind=link}
| Growth factor in bone matrix | Normal role in bone | Marine invertebrate species with growth factor analogue | Normal role in selected marine invertebrate |
|---|---|---|---|
| Bone morphogenic protein (BMP-2, BMP-4) | A keystone regulator of embryonic pattern formation. A key regulator of bone induction, maintenance and repair [33]. | Corals: Turbinaria reniformis, Acropora sp., Pavona cactus, Galaxea fascicularis, Hydnophora pilosa, Stylophora pistillata, Lobophyllia sp. [21,34]. | Secreted by calcifying epithelium during adult skeletogenesis [35]. Larva and polyp axial patterning in embryogenesis [35]. |
| Transforming growth factor (TGF-β)-like class | A vital regulator of embryonic pattern formation. A key regulator of bone induction, maintenance and repair [33]. | Marine sponge [36]: Amphimedon queenslandica Ctenophore [37]: Mnemiopsis leidyi Mollusca [38]: Planorbarius corneus, Viviparus ater, Viviparus contectus, Lymneae stagnalis, Mytilus galloprovincialis. | TGF-β ligands and TGF signal pathway components (e.g., SMAD) in early development and embryo patterning [39]. |
| Vascular endothelial growth factor (VEGF) | A cell signaling protein that induces vasculogenesis and angiogenesis [39]. | Cnidaria: Podocoryne carnea [20]. | VEGF signal pathway elements involved in morphogenesis of tentacle and gastrovascular canals. |
| Fibroblast growth factor (FGF) | ECM signaling protein that activates important pathways in skeletal development and regulate chondrogenesis and osteogenesis. A powerful angiogenic factor [40]. | Cnidaria: Nematostella vectensis [41]. | FGF ligands and receptors present where they function in gastrulation and development of chemosensory apical organ of ciliary larvae [42]. |
| Insulin like growth factor (IGF1-IGF2) | Linchpin in stimulating bone formation and remodeling. Regulates chondrocyte growth and metabolism. Stimulates collagen synthesis. Site-directed recruitment of osteoblasts [39]. | Nacre seashell: Haliotis laevigata [43]. | Nacre perlustrin has homology with N-terminal domain of mammalian insulin-like growth factor binding proteins (IGFBPs) [43]. |
| Tissue necrosis factor (TNF) | Inflammation cytokine and regulator of immune cells. One role is as a potent actor in bone re-modeling. It carries out this function through the MAPK pathway which, controls differentiation and proliferation through JNK mediator. It is also prominent regulator of osteoclastogenesis [39]. | TNF-α present in Mytilus edulis [44,45]. Their immunocytes respond to TNF in same manner as human granulocytes. | Cytokine analogues present in Mytilus edulis neural tissue [44,45]. |
| Epithelium growth factor (EGF) | A vital regulator of bone cell metabolism in formation and resorption [46]. | Marine sponge: Lubomirskia baicalensis [47]. | Genes coding for EGF-like molecules expressed for patterning silica structural modules |
| Platelet derived growth factor (PDGF) | A keystone regulator of cell division and growth. It is vital inductive signal in tissue remodeling morphogenesis and cell differentiation. It is a potent mitogen for bone cells. It also functions as a chemotactic factor for MSC and osteoblasts [48]. | Mollusca [48] Planorbarius corneus, Viviparus ater, Viviparus contectus, Lymneae stagnalis, Mytilus galloprovincialis. | Stimulate chemotaxis and phagocytic activity of wound repairing Molluscan immunocytes [48]. |
| Coral species | MW banding in kDa | Reference | Amino acid compositions | Protein gel chromatography |
|---|---|---|---|---|
| Synularia polydactyla (spicules) | 109, 83, 70, 63, 41, 30, 22 | Rehman et al. 2005 [49]. | Sequencing of 70 kDa and 63 kDa proteins. Enriched acidic amino acids. 70: glutamate; 63: glycine. |  Reproduced with permission from Wiley-VCH Verlag GmbH, 2005 [49]. |
| Galaxea fascicularis | 53, 45 | Fukuda et al. 2003 [50]. | Rich in cysteine (dicysteine repeat pattern) Aspartate and asparagine. |  Reproduced with permission from Elsevier Inc., 2003 [50]. |
| Balanophylli a europaea | IOM and SOM: 14.4, 13.9, 66 | Goffredo et al. 2011 [51]. | SOM: high acidic amino acids. IOM: high hydrophobic residues. | 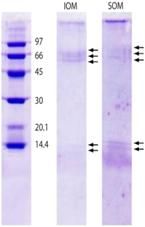 Reproduced with permission from PLoS One [51]. |
| Stylophora Pistillata Pavona Cactus | 55, 37, 47, 68, 50, 47, 37, 33 (from column 1: silver staining) | Puverel et al. 2005 [52]. | SOM: Soluble organic matrix. |  Reproduced with permission from Elsevier Inc., 2005 [52]. |
| C. rubrum | (A): 6 bands at 81, 55, 47, 44, 12 and 10 kDa; | Deubreil et al. [53]. |  Reproduced with permission from Springer 2011 [53]. | |
| C. konojoi | (B): 6 bands at 53, 45, 43, 31, 11.5 and 10.5 kDa; | |||
| C. secundum | (C): 5 bands at 44, 42, 31, 11 and 10.5 kDa; | |||
| C. elatius | (D): 4 bands at 43, 33, 11.5 and 10 kDa. |
| Marine sponge species | MW banding in kDa | Reference | Amino acid compositions |
|---|---|---|---|
| Astroscleria willeyana (A coralline demosponge) | 33–41 | Jackson et al. 2006 [54] | Not characterised |
Nacre Organic Matrix Proteins Can Promote Bone Formation in Humans
6. Use of Proteomic Methods to Retrieve Clinically Relevant Matrix Proteins
7. Conclusions
References
- Dimitriou, R.; Jones, E.; McGonagle, D.; Giannoudis, P.V. Bone regeneration: Current concepts and future directions. BMC Med. 2011, 9, 66. [Google Scholar] [CrossRef]
- Romano, N.H.; Sengupta, D.; Chung, C.; Heilshorn, S.C. Protein-engineered biomaterials: Nanoscale mimics of the extracellular matrix. Biochim. Biophys. Acta 2011, 1810, 339–349. [Google Scholar] [CrossRef]
- Lee, K.; Silva, E.A.; Mooney, D.J. Growth factor delivery-based tissue engineering: General approaches and a review of recent developments. J. R. Soc. Interface 2011, 8, 153–170. [Google Scholar] [CrossRef]
- Lutolf, M.P.; Hubbell, J.A. Synthetic biomaterials as instructive extracellular microenvironments for morphogenesis in tissue engineering. Nat. Biotechnol. 2005, 23, 47–55. [Google Scholar] [CrossRef]
- Gillette, B.M.; Jensen, J.A.; Tang, B.; Yang, G.J.; Bazargan-Lari, A.; Zhong, M.; Sia, S.K. In situ collagen assembly for integrating microfabricated three-dimensional cell-seeded matrices. Nat. Mater. 2008, 7, 636–640. [Google Scholar] [CrossRef]
- Geckil, H.; Xu, F.; Zhang, X.; Moon, S.; Demirci, U. Engineering hydrogels as extracellular matrix mimics. Nanomedicine 2010, 5, 469–484. [Google Scholar] [CrossRef]
- Powell, K. It’s the ecology, stupid! Nature 2005, 435, 268–270. [Google Scholar] [CrossRef]
- Kaplan, D.L.; Moon, R.T.; Vunjak-Novakovic, G. It takes a village to grow a tissue. Nat. Biotechnol. 2005, 10, 1237–1239. [Google Scholar]
- Jha, R.K.; Xu, Z.-R. Biomedical compounds from marine organisms. Mar. Drugs 2004, 2, 123–146. [Google Scholar] [CrossRef]
- Senni, K.; Pereira, J.; Gueniche, F.; Delbarre-Ladrat, C.; Sinquin, C.; Ratiskol, J.; Godeau, G.; Fischer, A.M.; Helley, D.; Colliec-Jouault, D. Marine polysaccharides: A source of bioactive molecules for cell therapy and tissue engineering. Mar. Drugs 2011, 9, 1664–1681. [Google Scholar] [CrossRef]
- Farre, B.; Dauphin, Y. Lipids from the nacreous and prismatic layers of two Pteriomorpha Mollusc shells. Comp. Biochem. Physiol. B 2009, 152, 103–109. [Google Scholar] [CrossRef]
- Halvey, S. Microbiology: Applications in Food Biotechnology; Nga, B.H., Lu, Y.K., Eds.; Elsevier Applied Science Press: New York, NY, USA, 1990; pp. 123–134. [Google Scholar]
- Sampath, T.K.; Rashka, K.E.; Doctor, J.S.; Tucker, R.F.; Hoffman, F.M. Drosophila transforming growth factor β superfamily proteins induce endochondral bone formation in mammals. Proc. Natl. Acad. Sci. USA 1993, 90, 6004–6008. [Google Scholar]
- Harcet, M.; Roller, M.; Cetkovic, H.; Perina, D.; Wiens, M.; Muller, W.E.G.; Vlahovicek, K. Demosponge EST sequencing reveals a complex genetic toolkit of the simplest metazoans. Mol. Biol. Evol. 2010, 27, 2747–2756. [Google Scholar]
- Ostrander, G.K.; Salzberg, S.L.; Downs, C.; Heidelberg, K.; Venter, J.C.; Fraser, C.M. Sequencing the Genome of the Coral, Porites lobata; National Human Genome Research Institute (NIH): Bethesda, MD, USA, 2003. [Google Scholar]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A revolutionary tool for transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Lowenstam, H.A.; Weiner, S. On Biomineralization; Oxford University Press: New York, NY, USA, 1989. [Google Scholar]
- Ozbek, S.; Balasubramanian, P.G.; Chiquet-Ehrismann, R.; Tucker, R.P.; Adams, J.C. The evolution of extracellular matrix. Mol. Cell. Biol. 2010, 21, 4300–4305. [Google Scholar] [CrossRef]
- Bentley, A.A.; Adams, J.C. The evolution of thrombospondins and their ligand binding activities. Mol. Biol. Evol. 2010, 2, 218–221. [Google Scholar]
- Seipel, K.; Eberhardt, M.; Muller, P.; Pescia, E.; Yanze, N.; Schmid, V. Homologs of vascular endothelial growth factor and receptor, VEGF and VEGFR, in the jellyfish Podocoryne carnea. Dev. Dyn. 2004, 231, 303–312. [Google Scholar] [CrossRef]
- Zoccola, D.; Moya, A.; Beranger, G.E.; Tambutte, E.; Allemand, D.; Carle, G.F.; Tambutte, S. Specific expression of BMP2/4 ortholog in biomineralizing tissues of corals and action on mouse BMP receptor. Mar. Biotechnol. 2000, 11, 260–269. [Google Scholar]
- Allemand, D.; Tambutté, E.; Zoccola, D.; Tambutté, S. Coral Calcification, Cells to Reef. In Coral Reefs: An Ecosystem in Transition; Dubinsky, Z., Stambler, N., Eds.; Springer Science and Business Media B.V.: Dordrecht, The Netherlands, 2011; pp. 119–150. [Google Scholar]
- Kramarsky-Winter, E.; Loya, Y. Tissue regeneration in the coral Fungia granulosa: The effect of extrinsic and intrinsic factors. Mar. Biol. 2000, 137, 867–873. [Google Scholar] [CrossRef]
- Nichols, S.A.; Dirks, W.; Pearse, J.S.; King, N. Early evolution of animal cell signaling and adhesion genes. Proc. Natl. Acad. Sci. USA 2006, 103, 12451–12456. [Google Scholar]
- Gautret, P.; Cuif, J.-P.; Stolarski, J. Organic components of the skeleton of Scleractinian corals—Evidence from in situ acridine orange staining. Acta Palaentol. Pol. 2000, 45, 107–118. [Google Scholar]
- Li, J.W.-H.; Vederas, J.C. Drug discovery and natural products: End of an era or an endless frontier? Science 2009, 325, 161. [Google Scholar] [CrossRef]
- Proksch, P.; Edrada-Ebel, R. Drugs from the sea—Opportunities and obstacles. Mar. Drugs 2003, 1, 5–17. [Google Scholar] [CrossRef]
- Manning, T.J.; Rhodes, E.; Land, M.; Parkman, R.; Sumner, N.; Lam, T.T.; Marshall, A.G.; Phillips, D. Impact of environmental conditions on the marine natural product bryostatin 1. Nat. Prod. Res. 2006, 20, 611–628. [Google Scholar] [CrossRef]
- Bruckner, A.W. Life-saving products form coral reefs: Coral reefs are storehouses of genetic resources with vast medicinal potential, but they must be properly managed. Issues Sci. Technol. 2002, 18, 39–45. [Google Scholar]
- Helman, Y.; Natale, F.; Sherrell, R.M.; LaVigne, M.; Starovoytov, V.; Gorbunov, M.Y.; Falkowski, P.G. Extracellular matrix production and calcium carbonate precipitation by coral cells in vitro. Proc. Natl. Acad. Sci. USA 2008, 105, 54–58. [Google Scholar]
- Horoszowski-Fridman, Y.B.; Izhaki, I.; Rinkevich, B. Engineering of coral reef larval supply through transplantation of nursery farmed gravid colonies. J. Exp. Mar. Biol. Ecol. 2011, 3, 162–166. [Google Scholar] [CrossRef]
- Gateno, D.; Barki, Y.; Rinkevich, B. Aquarium maintenance of reef octocorals raised from field collected larvae. Aquar. Sci. Conserv. 2000, 2, 227–236. [Google Scholar]
- Sykaras, N.; Opperman, L.A. Bone morphogenetic proteins (BMPs): How do they function and what can they offer the clinician? J. Oral Sci. 2003, 45, 57–73. [Google Scholar] [CrossRef]
- Lelong, C.; Mathieu, M.; Favrel, P. Identification of new bone morphogenetic protein-related members in invertebrates. Biochimie 2001, 83, 423–426. [Google Scholar] [CrossRef]
- Reber-Muller, S.; Streitwolf-Engel, R.; Yanze, N.; Schmid, V.; Stierwald, M.; Erb, M.; Seipel, K. BMP2/4 and BMP5-8 in jellyfish development and transdifferentiation. Int. J. Dev. Biol. 2006, 50, 377–384. [Google Scholar]
- Srivastava, M.; Simakov, O.; Chapman, J.; Fahey, B.; Gauthier, M.E.A.; Mitros, T.; Richards, G.S.; Conaco, C.; Dacre, M.; Hellsten, U.; et al. The Amphimedon queenslandica genome and the evolution of animal complexity. Nature 2010, 466, 720–726. [Google Scholar]
- Pang, K.; Ryan, J.F.; Baxevanis, A.D.; Martindale, M.Q. Evolution of the TGF-β signaling pathway and its potential role in the Ctenophore, Mnemiopsis leidyi. PLoS One 2011, 6, e24152. [Google Scholar]
- Franchini, A.; Ottaviani, E. Repair of molluscan tissue injury: Role of PDGF and TGF-β1. Tissue Cell 2000, 32, 312–321. [Google Scholar] [CrossRef]
- Lind, M. Growth factors: Possible new clinical tools. Acta Orthop. Scand. 1996, 67, 407–417. [Google Scholar] [CrossRef]
- Ornitz, D.M.; Marie, P.J. FGF signaling pathways in endochondral and intramembranous bone development and human genetic disease. Genes Dev. 2002, 15, 1446–1465. [Google Scholar]
- Rentzsch, F.; Fritzenwanker, J.H.; Scholz, C.B.; Technau, U. FGF signaling controls formation of the apical sensory organ in the cnidarian Nematostella vectensis. Development 2008, 135, 1761–1769. [Google Scholar] [CrossRef]
- Matus, D.Q.; Thomsen, G.H.; Martindale, M.Q. FGF signaling in gastrulation and neural development in Nematostella vectensis, an Anthozoan cnidarians. Dev. Genes Evol. 2007, 217, 137–148. [Google Scholar] [CrossRef]
- Weiss, I.M.; Göhring, W.; Fritz, M.; Mann, K. Perlustrin, a Haliotis laevigata (Abalone) Nacre protein, is homologous to the insulin-like growth factor binding protein N-terminal module of vertebrates. Biochem. Biophys. Res. Commun. 2001, 285, 244–249. [Google Scholar] [CrossRef]
- Beschin, A.; Bilej, M.; Torreele, E.; DeBaetselier, P. On the existence of cytokines in invertebrates. Cell. Mol. Life Sci. 2001, 58, 801–814. [Google Scholar] [CrossRef]
- Gerber, S.; Cadet, P.; Sheehan, M.; Stefano, G.B.; Mantione, K.J. Vertebrate interleukins originated in invertebrates? Invertebr. Surviv. J. 2000, 4, 95–100. [Google Scholar]
- Marie, P.J.; Hott, M.; Perheentupa, J. Effects of epidermal growth factor on bone formation and resorption in vivo. Am. J. Physiol. Endocrinol. Metab. 1990, 258, E275–E281. [Google Scholar]
- Wiens, M.; Belikov, S.I.; Kaluzhnaya, O.V.; Adell, T.; Schröder, H.C.; Perovic-Ottstadt, S.; Kaandorp, J.A.; Müller, W.E. Regional and modular expression of morphogenetic factors in the Demosponge, Lubomirskia baicalensis. Micron 2008, 39, 447–460. [Google Scholar] [CrossRef]
- Franchini, A.; Kletsas, D.; Ottaviani, E. Presence of PDGF and TGF-β1 immunoreactive molecules in invertebrate and vertebrate immunocytes: An evolutionary approach. Histochem. J. 1996, 28, 599–605. [Google Scholar] [CrossRef]
- Rehman, M.A.; Isa, Y.; Uehara, T. Proteins of calcified endoskeleton: II Partial amino acid sequences of endoskeletal proteins and the characterisation of proteinaceous organic matrix of spicules from the alcyonarian, Synularia polydactyla. Proteomics 2005, 5, 885–893. [Google Scholar] [CrossRef]
- Fukuda, I.; Ooki, S.; Fulita, T.; Murayama, E.; Nagasawa, H.; Isa, Y.; Watanabe, T. Molecular cloning of cDNA encoding a soluble protein in the coral exoskeleton. Biochem. Biophys. Res. Commun. 2003, 304, 11–17. [Google Scholar] [CrossRef]
- Goffredo, S.; Vergni, P.; Reggi, M.; Caroselli, E.; Sparla, F.; Levy, O.; Dubinsky, Z.; Falini, G. The skeletal organic matrix from Mediterranean coral, Balanophyllia europaea influences calcium carbonate precipitation. PLoS One 2011, 6, e22338. [Google Scholar]
- Puverel, S.; Tambutte, E.; Pereira-Mouries, L.; Zoccola, D.; Allemand, D.; Tambutte, S. Soluble organic matrix of two Scleractinian corals: Partial and comparative analysis. Comp. Biochem. Physiol. B 2005, 141, 480–487. [Google Scholar] [CrossRef]
- Debreuil, J.; Tambutté, S.; Zoccola, D.; Segonds, N.; Techer, N.; Marschal, C.; Allemand, D.; Kosuge, S.; Tambutté, É. Specific organic matrix characteristics in skeletons of Corallium species. Mar. Biol. 2011, 158, 2765–2774. [Google Scholar]
- Jackson, D.J.; Macis, L.; Reitner, J.; Degnan, B.M.; Wörheide, G. Sponge Paleogenomics reveals an ancient role for carbonic anhydrase in skeletogenesis. Science 2007, 316, 1893–1895. [Google Scholar] [CrossRef]
- Atlan, G.; Balmain, N.; Berland, S.; Vidal, B.; Lopez, E. Rconstruction of human maxillary defects with nacre powder: Histological evidence for bone regeneration. C. R. Acad. Sci. Ser. III 1997, 320, 253–258. [Google Scholar] [CrossRef]
- Mouriès, L.P.; Almeida, M.J.; Milet, C.; Lopez, E. Bioactivity of nacre water-soluble organic matrix from the bivalve mollusc Pinctada maxima in three mammalian cell types: Fibroblasts, bone marrow stromal cells and osteoblasts. Comp. Biochem. Physiol. B 2002, 132, 217–229. [Google Scholar] [CrossRef]
- Almeida, M.J.; Milet, C.; Peduzzi, J.; Pereira, L.; Haigle, J.; Barthelemy, M.; Lopez, E. Effect of water-soluble matrix fraction extracted from the nacre of Pinctada maxima on the alkaline phosphatase activity of cultured fibroblasts. J. Exp. Zool. 2000, 15, 327–334. [Google Scholar]
- Milet, C.; Berland, S.; Lamghari, M.; Mouries, L.; Jolly, C.; Borzeix, S.; Doumenc, D.; Lopez, E. Conservation of signal molecules involved in biomineralisation control in calcifying matrices of bone and shell. C. R. Palevol 2004, 3, 493–501. [Google Scholar] [CrossRef]
- Oliveira, D.V.; Silva, T.S.; Cordeiro, O.D.; Cavaco, S.I.; Simes, D.C. Identification of proteins with potential osteogenic activity present in the water-soluble matrix proteins from Crassostrea gigas nacre using a proteomic approach. Sci. World J. 2012, 2012, 765909. [Google Scholar]
- Pennington, S.R.; Wilkins, M.R.; Hochstrasser, D.F.; Dunn, M.J. Proteome analysis: From protein characterization to biological function. Trends Cell Biol. 1997, 7, 168–173. [Google Scholar] [CrossRef]
- Wilkins, M.R.; Sanchez, J.-C.; Gooley, A.A.; Appel, R.D.; Humphrey-Smith, I.; Hochstrasser, D.F.; Williams, K.L. Progress with proteome projects expressed by a genome should be identified and how to do it. Biotechnol. Genet. Eng. Rev. 1996, 13, 19–50. [Google Scholar]
- Molloy, M.P.; Witzmann, F.A. Proteomics: Technologies and applications. Brief. Funct. Genomics Proteomics 2002, 1, 23–39. [Google Scholar]
- Dauphin, Y. Comparative studies of skeletal soluble matrices from some Scleractinian corals and molluscs. Int. J. Biol. Macromol. 2001, 28, 293–304. [Google Scholar] [CrossRef]
- Watanabe, T.; Fukuda, I.; China, K.; Isa, Y. Molecular analyses of protein components of the organic matrix in the exoskeleton of two Scleractinian coral species. Comp. Biochem. Physiol. B 2003, 136, 767–774. [Google Scholar] [CrossRef]
- Marin, F.; Luquet, G. Molluscan shell proteins. C. R. Palevol 2004, 3, 469–492. [Google Scholar] [CrossRef]
- Sroga, G.E.; Karim, L.; Colón, W.; Vashishth, D. Biochemical characterization of major bone-matrix proteins using nanoscale-size bone samples and proteomics methodology. Mol. Cell. Proteomics 2011, 10, M110.006718. [Google Scholar] [CrossRef]
- Marie, B.; Marin, F.; Marie, A.; Bedouet, L.; Dubost, L.; Alcaraz, G.; Milet, C.; Luquet, G. Evolution of nacre: Biochemistry and proteomics of the shell organic matrix of the Cephalopod Nautilus macromphalus. ChemBioChem 2009, 10, 1495–1506. [Google Scholar]
- Marie, B.; Marie, A.; Jackson, D.J.; Dubost, L.; Degnan, B.M.; Milet, C.; Marin, F. Proteomic analysis of the organic matrix of the abalone Haliotis asinine calcified shell. Proteomic Sci. 2010, 8, 54. [Google Scholar]
- Drake, J.L.; Mass, T.; Haramaty, L.; Zelzion, H.; Bhattacharya, D.; Falkowski, P.G. Proteomic analysis of skeletal organic matrix from the stony coral, Stylophora pistillata. Proc. Natl. Acad. Sci. USA 2013, 110, 3788–3793. [Google Scholar]
- Mann, K.; Poustka, A.J.; Mann, M. In-depth, high accuracy proteomics of sea urchin tooth organic matrix. Proteome Sci. 2008, 6, 33. [Google Scholar] [CrossRef]
- Samples Availability: Available from the authors.
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Green, D.W.; Padula, M.P.; Santos, J.; Chou, J.; Milthorpe, B.; Ben-Nissan, B. A Therapeutic Potential for Marine Skeletal Proteins in Bone Regeneration. Mar. Drugs 2013, 11, 1203-1220. https://doi.org/10.3390/md11041203
Green DW, Padula MP, Santos J, Chou J, Milthorpe B, Ben-Nissan B. A Therapeutic Potential for Marine Skeletal Proteins in Bone Regeneration. Marine Drugs. 2013; 11(4):1203-1220. https://doi.org/10.3390/md11041203
Chicago/Turabian StyleGreen, David W., Matthew P. Padula, Jerran Santos, Joshua Chou, Bruce Milthorpe, and Besim Ben-Nissan. 2013. "A Therapeutic Potential for Marine Skeletal Proteins in Bone Regeneration" Marine Drugs 11, no. 4: 1203-1220. https://doi.org/10.3390/md11041203
APA StyleGreen, D. W., Padula, M. P., Santos, J., Chou, J., Milthorpe, B., & Ben-Nissan, B. (2013). A Therapeutic Potential for Marine Skeletal Proteins in Bone Regeneration. Marine Drugs, 11(4), 1203-1220. https://doi.org/10.3390/md11041203