Red Algal Bromophenols as Glucose 6-Phosphate Dehydrogenase Inhibitors

Abstract
:1. Introduction
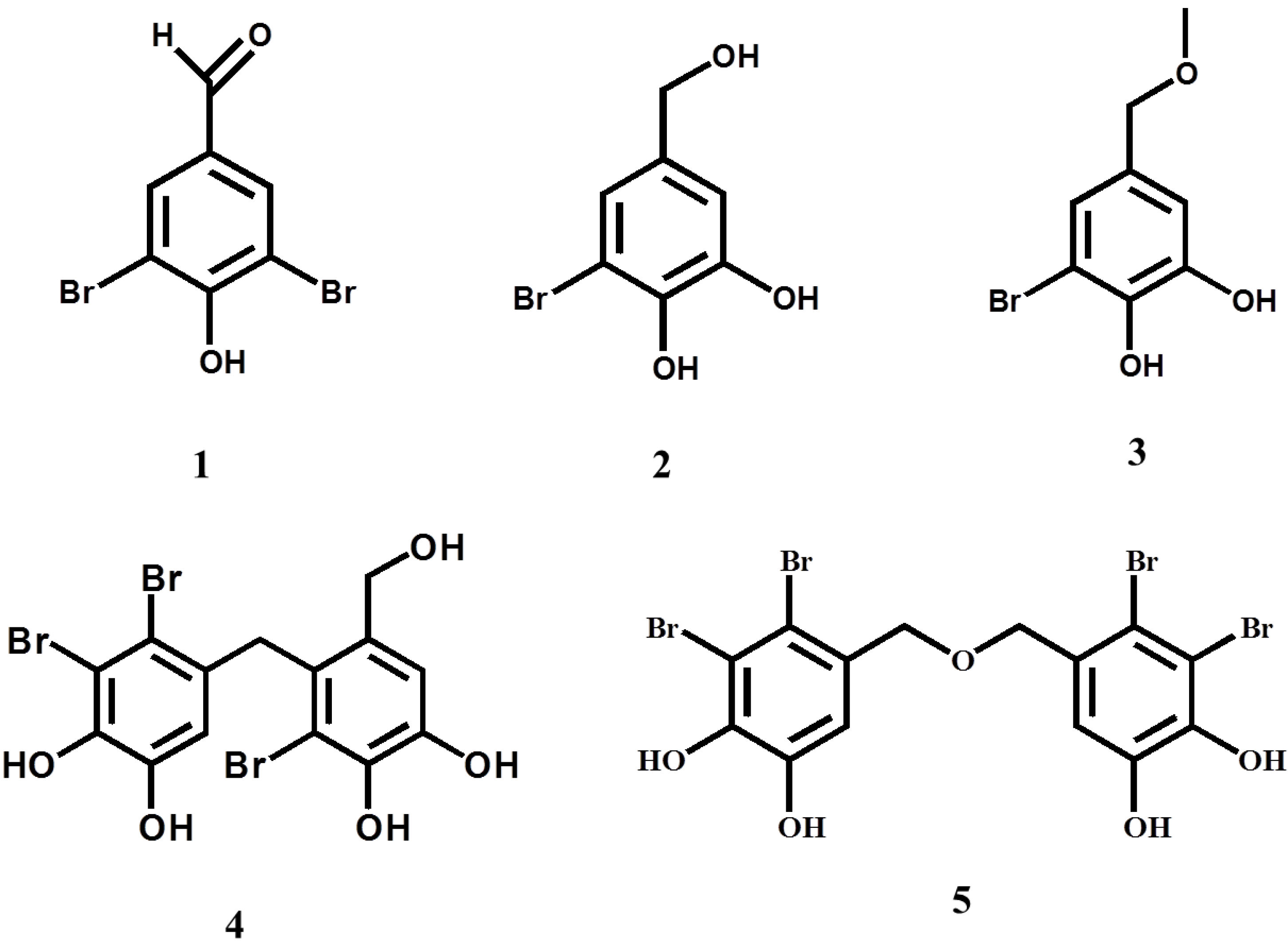
2. Results and Discussion

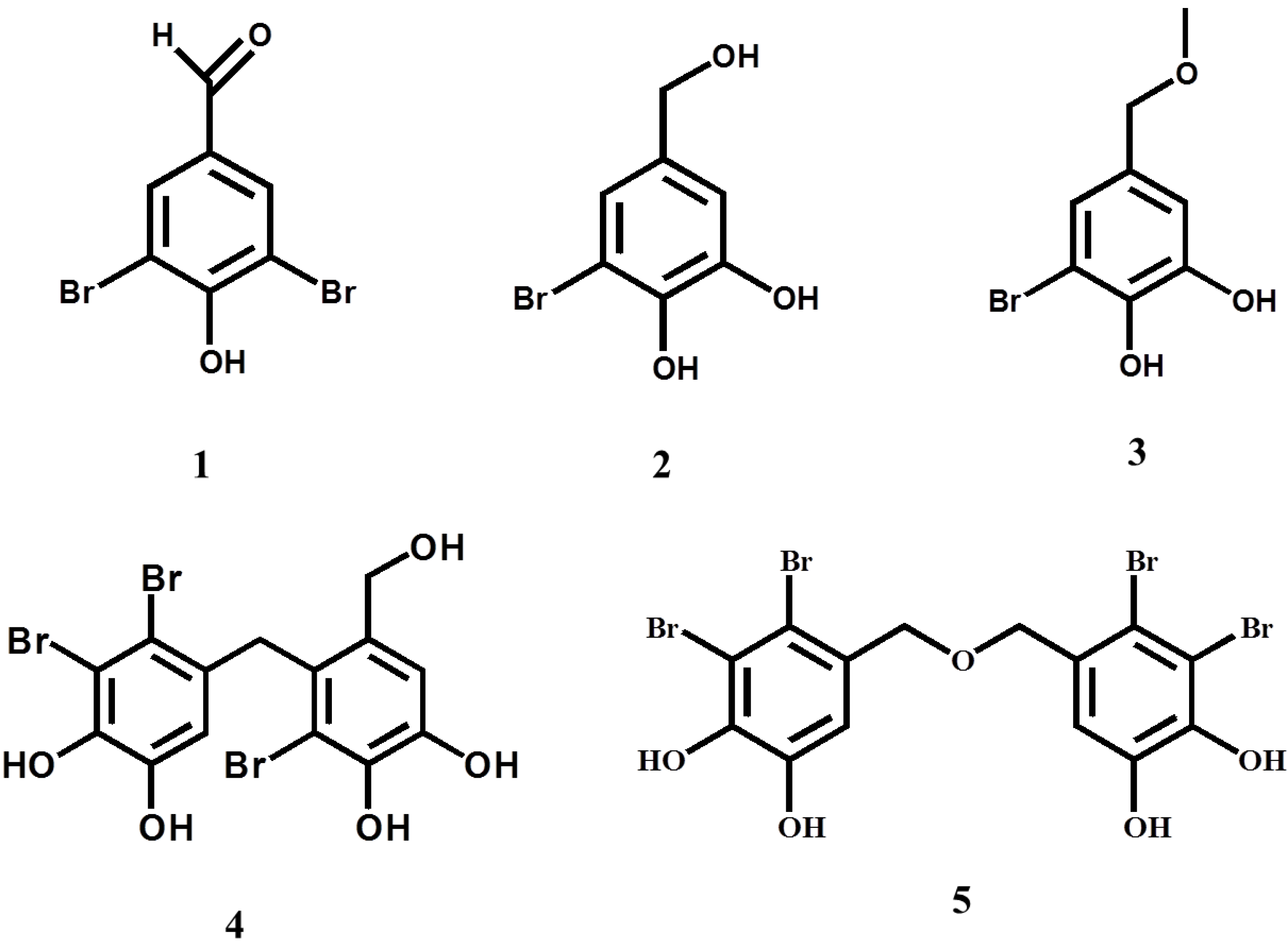{kind=link}
| Compound | IC50a (μM) |
|---|---|
| 1 | 76.6 ± 1.1 |
| 2 | >370 (37.6 ± 1.6%) b |
| 3 | >340 (35.7 ± 4.4%) c |
| 4 | 4.01 ± 0.30 |
| 5 | 0.85 ± 0.10 |
| Epigallocatechin gallate | 7.70 ± 0.14 |
3. Experimental Section
3.1. General
3.2. Algal Material
3.3. Enzyme Assay
3.4. Extraction and Purification of G6PD Inhibitors
4. Conclusions
Acknowledgments
Conflicts of Interest
References
- Ochi, M.; Watanabe, M.; Miura, I.; Taniguchi, M.; Tokoroyama, T. Amijiol, isoamijiol, and 14-deoxyamijiol, three new diterpenoids from the brown seaweed Dictyota linearis. Chem. Lett. 1980, 9, 1229–1232. [Google Scholar]
- Glombitza, W.K.; Hauperich, S. Phlorotannins from the alga Cystophora torulosa. Phytochemistry 1997, 46, 735–740. [Google Scholar] [CrossRef]
- Fuller, W.R.; Cardellina, H.J.; Kato, Y.; Brinen, S.L.; Clardy, J.; Snader, M.K.; Boyd, R.M. A pentahalogenated monoterpene from the red alga Portieria hornemannii produces a novel cytotoxicity profile against a diverse panel of human tumor cell lines. J. Med. Chem. 1992, 35, 3007–3011. [Google Scholar] [CrossRef]
- Lin, S.A.; Stout, P.E.; Prudhomme, J.; Roch, K.L.; Fairchild, R.C.; Franzblau, G.S.; Aalbersberg, W.; Hay, E.M.; Kubanek, J. Bioactive bromophycolides R–U from the Fijian red alga Callophycus serratus. J. Nat. Prod. 2010, 73, 275–278. [Google Scholar] [CrossRef]
- Lhullier, C.; Falkenberg, M.; Ioannou, E.; Quesada, A.; Papazafiri, P.; Horta, A.P.; Schenkel, P.E.; Vagias, C.; Roussis, V. Cytotoxic halogenated metabolites from the Brazilian red alga Laurencia catarinensis. J. Nat. Prod. 2010, 73, 27–32. [Google Scholar] [CrossRef]
- Hodgkin, H.J.; Craige, S.J.; Mcinnes, G.A. The occurrence of 2,3-dibromobenzyl alcohol 4,5-disulfate, dipotassium salt, in Polysiphonia lanosa. Can. J. Chem. 1966, 44, 74–78. [Google Scholar] [CrossRef]
- Wang, B.G.; Gloer, J.B.; Ji, N.Y.; Zhao, J.C. Halogenated organic molecules of Rhodomeraceae origin: Chemistry and biology. Chem. Rev. 2013, 113, 3632–3685. [Google Scholar] [CrossRef]
- Li, K.; Li, X.M.; Gloer, B.J.; Wang, B.G. Isolation, characterization, and antioxidant activity of bromophenols of the marine red alga Rhodomela confervoides. J. Agric. Food Chem. 2011, 59, 9916–9921. [Google Scholar] [CrossRef]
- Liu, M.; Zhang, W.; Wei, J.; Qiu, L.; Lin, X. Marine bromophenol bis(2,3-dibromo-4,5-dihydroxybenzyl)ether, induces mitochondrial apoptosis in K552 cells and inhibits topoisomerase I in vitro. Toxicol. Lett. 2012, 211, 126–134. [Google Scholar] [CrossRef]
- Wiemer, F.D.; Idler, D.D.; Fenical, W. Vidalols A and B, new anti-inflammatory bromophenols from the Caribbean marine red alga Vidalia obtusaloba. Experientia 1991, 47, 851–853. [Google Scholar] [CrossRef]
- Xu, N.; Fan, X.; Yan, X.; Li, X.; Niu, R.; Tseng, C.K. Antibacterial bromophenols from the marine red algae Rhodomela confervoides. Phytochemistry 2003, 62, 1221–1224. [Google Scholar] [CrossRef]
- Kurihara, H.; Mitani, T.; Kawabata, J.; Takahashi, K. Two new bromophenols from the red alga Odonthalia corymbifera. J. Nat. Prod. 1999, 62, 882–884. [Google Scholar] [CrossRef]
- Marks, A.P.; Banks, J. Inhibition of mammarian glucose-6-phosphate dehydrogenase by steroids. Proc. Natl. Acad. Sci. USA 1960, 46, 447–452. [Google Scholar] [CrossRef]
- Yen, T.T.; Allan, A.J.; Pearson, V.D.; Acton, M.J.; Greenberg, M.M. Prevention of obesity in Avy/a mice by dehydroepiandrosterone. Lipids 1977, 12, 409–413. [Google Scholar] [CrossRef]
- Dessi, S.; Batteta, B.; Spano, O.; Pulisci, D.; Mulas, M.F.; Muntoni, S.; Armeni, M.; Sanna, C.; Antonucci, R.; Pani, P. Serum lipoprotein pattern as modified in G6PD-deficient children during haemolytic anaemia induced by fava bean ingestion. Int. J. Exp. Pathol. 1992, 73, 157–160. [Google Scholar]
- Kuo, W.Y.; Lin, J.Y.; Tang, K. Human glucose-6-phophate dehydrogenase (G6PD). Gene transforms NIH 3T3 cells and induces tumors in nude mice. Int. J. Cancer 2000, 85, 857–864. [Google Scholar] [CrossRef]
- Li, D.; Zhu, Y.; Tang, Q.; Lu, H.; Li, H.; Yang, Y.; Li, Z.; Tong, S. A new G6PD knockdown tumore-cell line with reduced proliferation and increased susceptibility to oxidative stress. Cancer Biother. Radiopharm. 2009, 24, 81–90. [Google Scholar] [CrossRef]
- Gupta, S.; Cordeiro, T.A.; Michels, P.A.M. Glucose-6-phosphate dehydrogenase is the target for the trypanocidal action of human steroids. Mol. Biochem. Parasitol. 2011, 176, 112–115. [Google Scholar] [CrossRef]
- Hamilton, M.N.; Dawson, M.; Fairweather, E.E.; Hamilton, S.N.; Hitchin, R.J.; James, I.D.; Jones, D.S.; Jordan, M.A.; Lyones, J.A.; Small, F.H.; et al. Novel steroid inhibitors of glucose 6-phosphate dehydrogenase. J. Med. Chem. 2012, 55, 4431–4445. [Google Scholar] [CrossRef]
- Shin, E.S.; Park, J.; Shin, J.M.; Cho, D.; Cho, S.Y.; Shin, D.W.; Ham, M.; Kim, J.B.; Lee, T.R. Cathchin gallates are NADP+-competitive inhibitors of glucose-6-phosphate dehydrogenase and other enzymes that employ NADP+ as a coenzyme. Bioorg. Med. Chem. 2008, 16, 3580–3586. [Google Scholar] [CrossRef]
- Kurata, K.; Amiya, T. A new bromophenol from the red alga Polysiphonia urceolata. Bull. Chem. Soc. Jpn. 1980, 53, 2020–2022. [Google Scholar] [CrossRef]
- Fan, X.; Xu, N.J.; Shi, J.G. Bromophenols from the red algae Rhodomela confervoides. J. Nat. Prod. 2003, 66, 455–458. [Google Scholar] [CrossRef]
- Kurihara, H.; Mitani, T.; Kawabata, J.; Takahashi, K. Inhibitory potencies of bromophenols from Rhodomelaceae algae against α-glucosidase activity. Fish. Sci. 1999, 65, 300–303. [Google Scholar] [CrossRef]
- Kim, K.Y.; Choi, K.S.; Kurihara, H.; Kim, S.M. β-Glucuronidase inhibitory activity of bromophenols purified from Grateloupia elliptica. Food Sci. Biotechnol. 2008, 17, 1110–1114. [Google Scholar]
- Shi, D.; Li, J.; Jiang, B.; Guo, S.; Su, H.; Wang, T. Bromophenols as inhibitors of protein tyrosine phosphatase 1B with antidiabetic properties. Bioorg. Med. Chem. Lett. 2012, 22, 2827–2832. [Google Scholar]
- Kim, K.Y.; Nguyen, T.H.; Kurihara, H.; Kim, S.M. α-glucosidase inhibitory activity of bromophenol purified from the red alga Polyopes lancifolia. J. Food Sci. 2010, 75, 145–150. [Google Scholar]
- Olive, C.; Geroch, E.M.; Levy, R.H. Glucose 6-phosphate dehydrogenase from Leuconostoc mesenteroides. J. Biol. Chem. 1971, 246, 2047–2057. [Google Scholar]
Supplementary Files
© 2013 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Mikami, D.; Kurihara, H.; Kim, S.M.; Takahashi, K. Red Algal Bromophenols as Glucose 6-Phosphate Dehydrogenase Inhibitors. Mar. Drugs 2013, 11, 4050-4057. https://doi.org/10.3390/md11104050
Mikami D, Kurihara H, Kim SM, Takahashi K. Red Algal Bromophenols as Glucose 6-Phosphate Dehydrogenase Inhibitors. Marine Drugs. 2013; 11(10):4050-4057. https://doi.org/10.3390/md11104050
Chicago/Turabian StyleMikami, Daisuke, Hideyuki Kurihara, Sang Moo Kim, and Koretaro Takahashi. 2013. "Red Algal Bromophenols as Glucose 6-Phosphate Dehydrogenase Inhibitors" Marine Drugs 11, no. 10: 4050-4057. https://doi.org/10.3390/md11104050
APA StyleMikami, D., Kurihara, H., Kim, S. M., & Takahashi, K. (2013). Red Algal Bromophenols as Glucose 6-Phosphate Dehydrogenase Inhibitors. Marine Drugs, 11(10), 4050-4057. https://doi.org/10.3390/md11104050