Structural Characterization and Anti-HSV-1 and HSV-2 Activity of Glycolipids from the Marine Algae Osmundaria obtusiloba Isolated from Southeastern Brazilian Coast

Abstract
:
1. Introduction
2. Results and Discussion
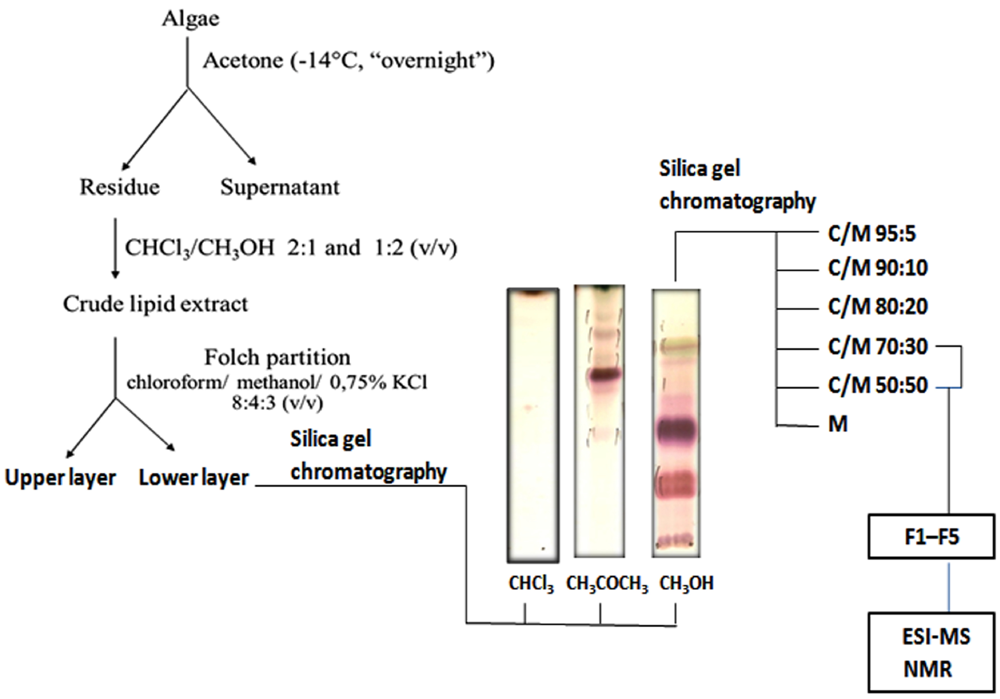
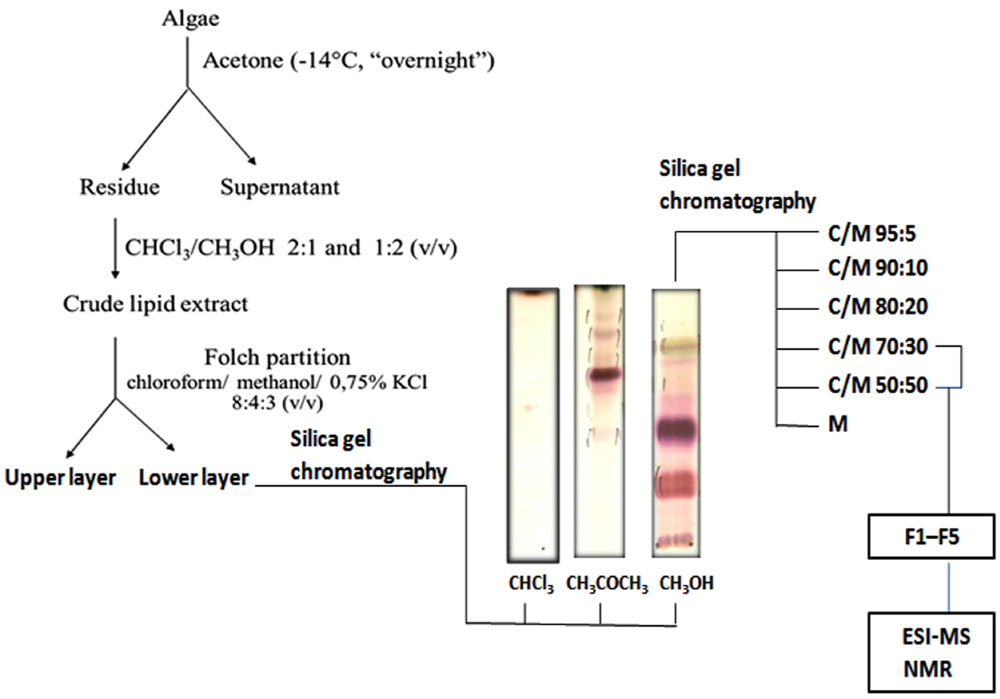
2.1. Lipid Fractionation
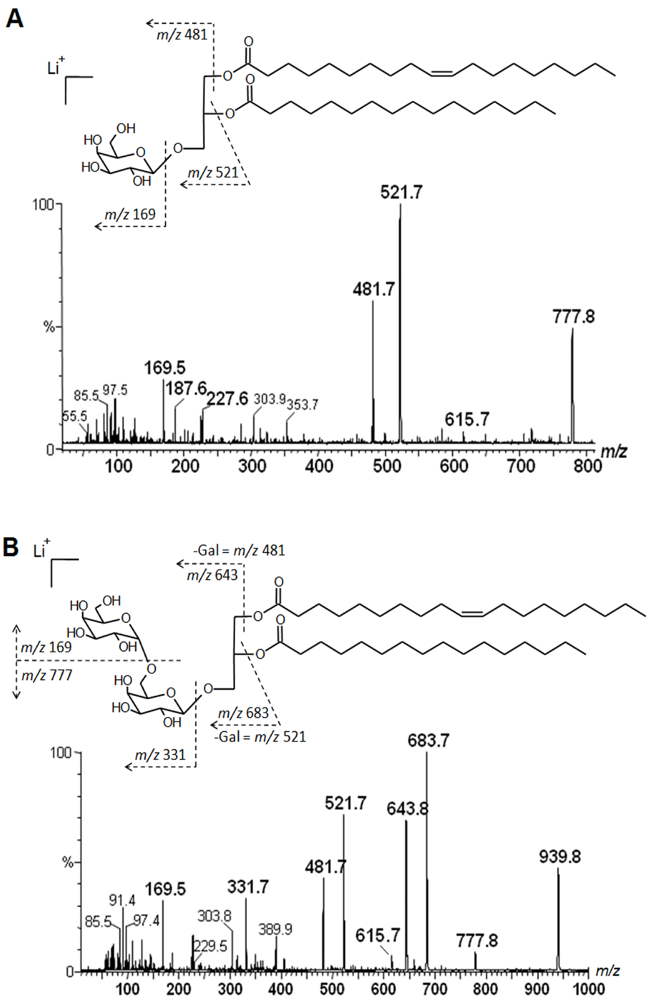
2.2. Mass Spectrometry of Neutral Glycolipids

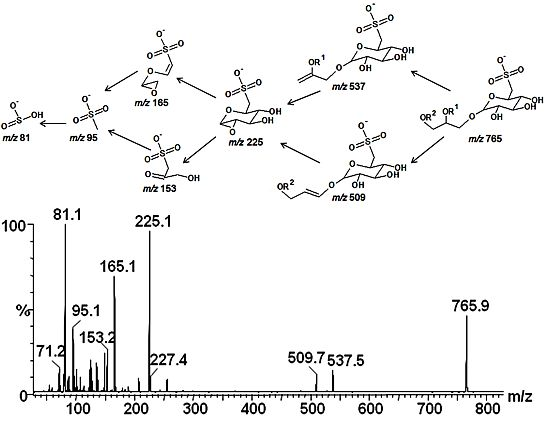
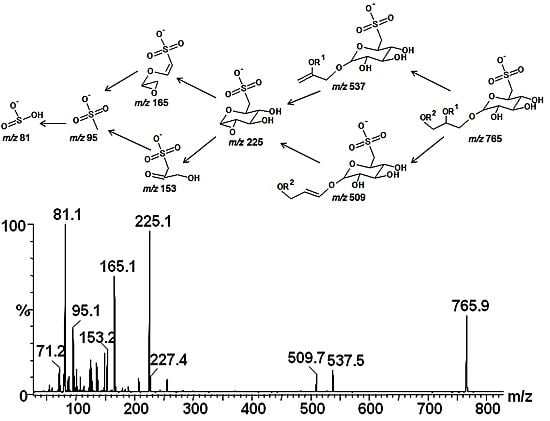
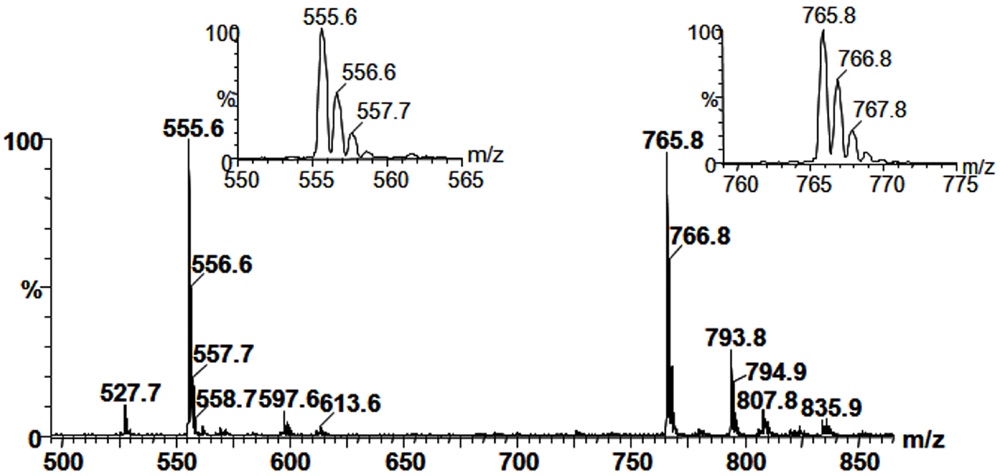
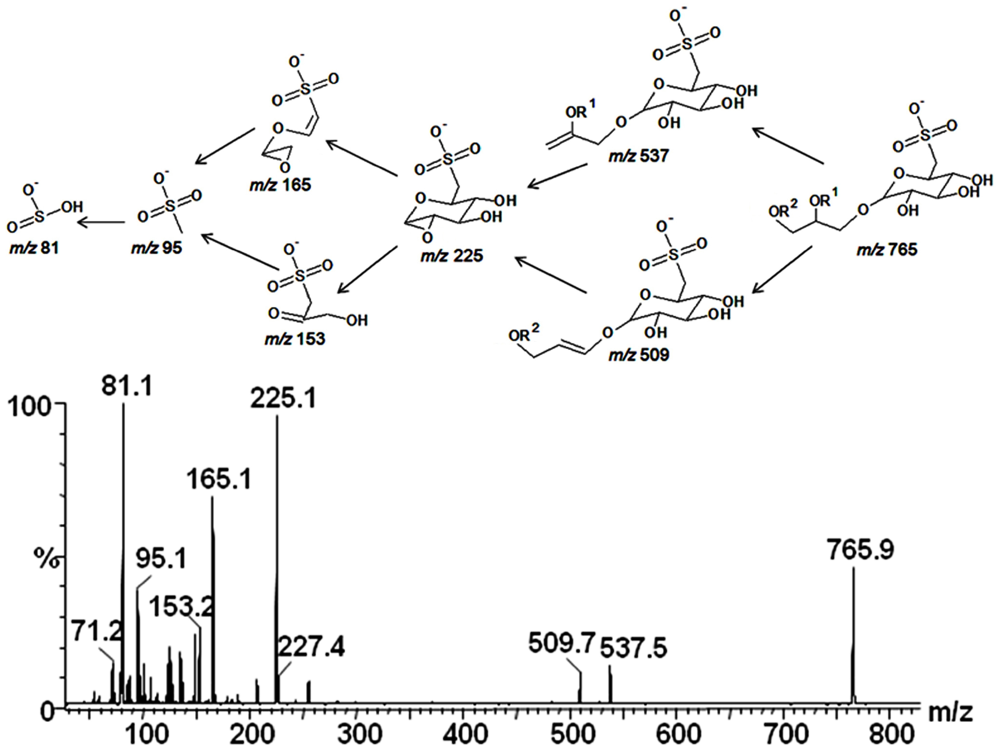
2.3. Mass Spectrometry of Sulfolipids
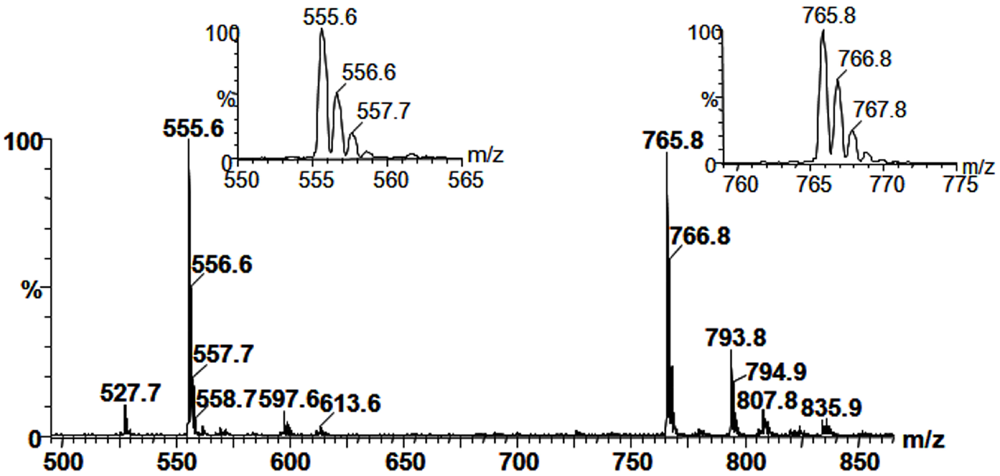
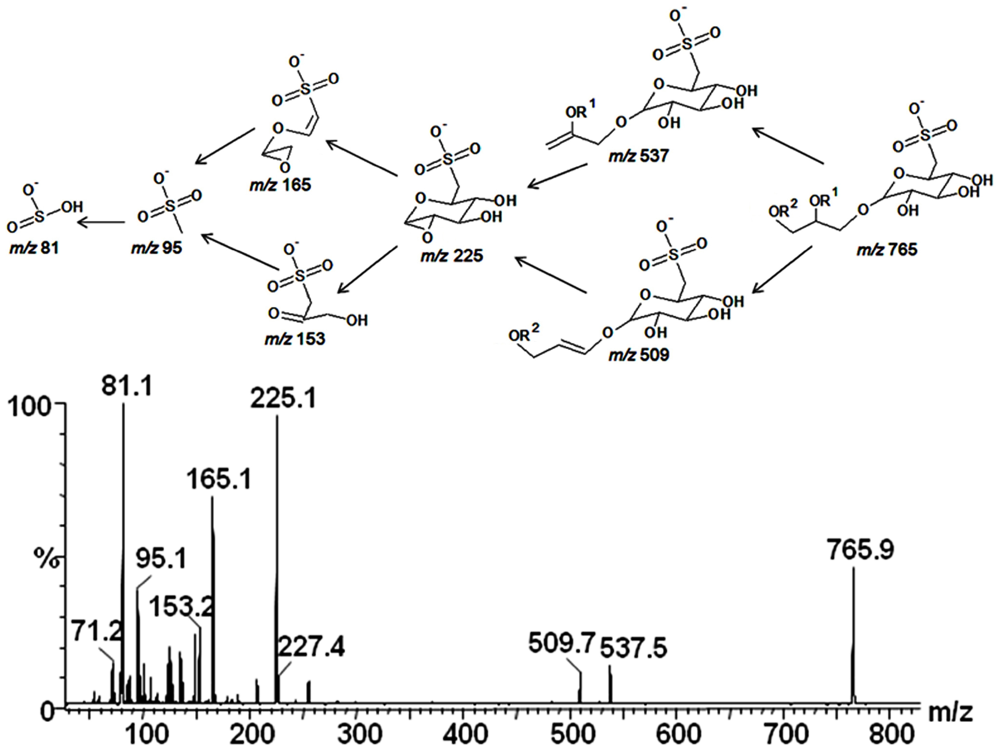
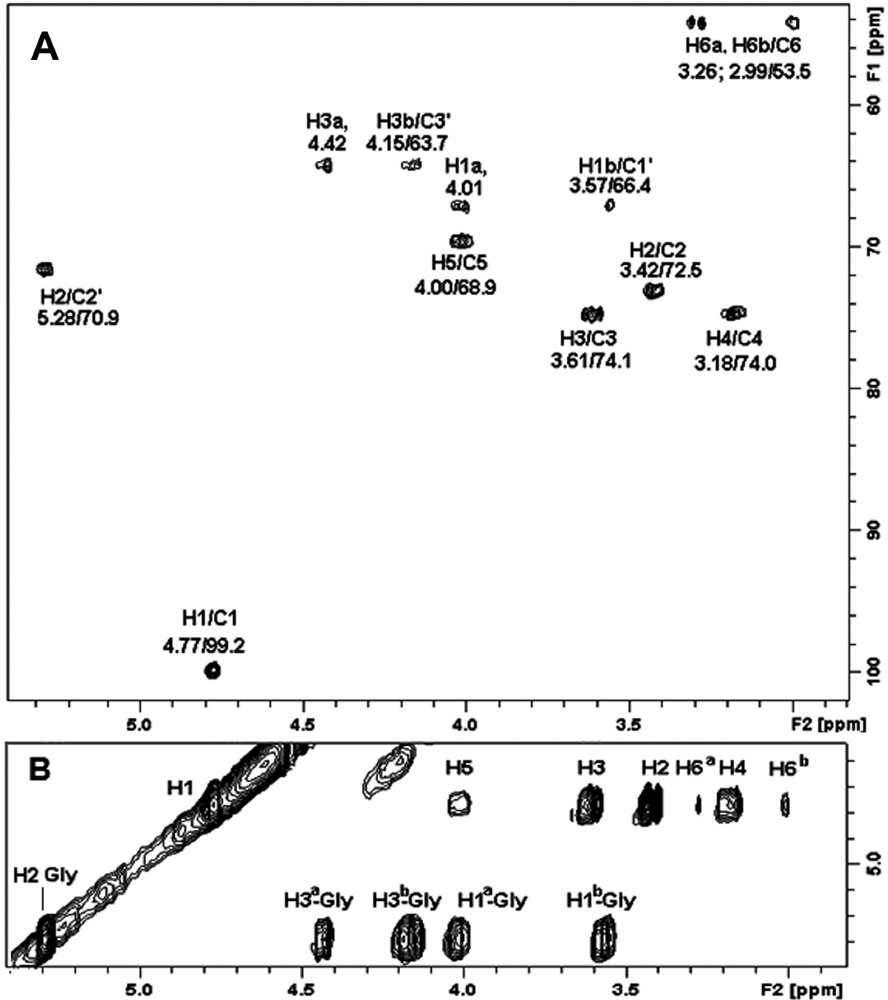
2.4. NMR Spectroscopy of Sulfolipids

2.5. Antiviral Activity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HSV-1 | HSV-2 | |||||||
|---|---|---|---|---|---|---|---|---|
| Compounds | MNTC (µg/mL) | CC50 (µg/mL) | PI | EC50 (µg/mL) | SI | PI | EC50 (µg/mL) | SI |
| Fraction 2 O. obtusiloba | 50 | 72 | 75 | 42 | 1.7 | 96 | 12 | 6.0 |
| Acyclovir | 200 | >200 | 99 | 0.8 | >250 | 99 | 1.38 | >145 |
3. Experimental Section
3.1. Biological Material
3.2. Extraction and Fractionation of Lipids
3.3. Mass Spectrometry
3.4. Nuclear Magnetic Resonance
3.5. Cells and Viruses
3.6. Cytotoxicity Assay
4. Conclusions
Acknowledgments
- Samples Availability: Available from the authors.
References
- Sanina, N.M.; Goncharova, S.N.; Kostetsky, E.Y. Fatty acid composition of individual polar lipid classes from marine macrophytes. Phytochemistry 2004, 65, 721–730. [Google Scholar]
- Kim, Y.H.; Kim, E.H.; Lee, C.; Kim, M.H.; Rho, J.R. Two new monogalactosyl diacylglycerols from brown alga Sargassum thunbergii. Lipids 2007, 42, 395–399. [Google Scholar]
- Illijas, M.I.; Indy, J.R.; Yasui, H.; Itabashi, Y. Lipid class and fatty acid composition of a little-known and rarely collected alga Exophyllum wentii Weber-van Bosse from Bali Island, Indonesia. J. Oleo Sci. 2009, 58, 103–110. [Google Scholar]
- Al-Fadhli, A.; Wahidulla, S.; D’Souza, L. Glycolipids from the red alga Chondria armata (Kutz.) Okamura. Glycobiology 2006, 16, 902–915. [Google Scholar] [CrossRef]
- Sassaki, G.L.; Gorin, P.A.J.; Tischer, C.A.; Iacomini, M. Sulfonoglycolipids from the lichenized basidiomycete Dictyonema glabratum: Isolation, NMR, and ESI-MS approaches. Glycobiology 2001, 11, 345–351. [Google Scholar]
- Souza, L.M.; Iacomini, M.; Gorin, P.A.J.; Sari, R.S.; Haddad, M.A.; Sassaki, G.L. Glyco- and sphingophosphonolipids from the medusa Phyllorhiza punctata: NMR and ESI-MS/MS fingerprints. Chem. Phys. Lipids 2007, 145, 85–96. [Google Scholar]
- Gustafson, K.R.; Cardellina, J.H.; Fuller, R.W.; Weislow, O.W.; Kiser, R.F.; Snader, K.M.; Patterson, G.M.L.; Boyd, M.R. AIDS-antiviral sulfolipids from cyanobacteria (blue-green algae). J. Nat. Cancer Inst. 1989, 81, 1254–1258. [Google Scholar]
- Reshef, V.; Mizrachi, E.; Maretzki, T.; Silberstein, C.; Loya, S.; Hizi, A.; Carmeli, S. New acylated sulfoglycolipids and digalactolipids and related known glycolipids from cyanobacteria with a potential to inhibit the reverse transcriptase of HIV-1. J. Nat. Prod. 1997, 60, 1251–1260. [Google Scholar]
- Morimoto, T.; Nagatsu, A.; Murakami, N.; Sakakibara, J.; Tokuda, H.; Nishimo, H.; Iwashima, A. Antitumor promoting glyceroglycolipids from the green alga Chlorella vulgaris. Phytochemistry 1995, 40, 1433–1437. [Google Scholar]
- Ohta, K.; Mizushina, Y.; Hirata, N.; Takemura, M.; Sugawara, F.; Matsukage, A.; Yoshida, S.; Sakaguchi, K. Sulfoquinovosyldiacylglycerol, KM043, a new potent inhibitor of eukaryotic DNA polymerases and HIV-reverse transcriptase type 1 from a marine red alga, Gigartina tenella. Chem. Pharm. Bull. (Tokyo) 1998, 46, 684–686. [Google Scholar] [CrossRef]
- Naumann, I.; Darsow, K.H.; Walter, C.; Lange, H.A.; Buchholz, R. Identification of sulfoglycolipids from the alga Porphyridium purpureum by matrix-assisted laser desorption/ionisation quadrupole ion trap time of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2007, 21, 3185–3192. [Google Scholar]
- Damonte, E.B.; Matulewicz, M.C.; Cerezo, A.S.; Coto, C.E. Herpes simplex virus-inhibitory sulfated xylogalactans from the red seaweed Nothogenia fastigiata. Exp. Chemoter. 1996, 42, 57–64. [Google Scholar]
- Witvrouw, M.; de Clercq, E. Sulfated polysaccharides extracted from sea algae as potential antiviral drugs. Gen. Pharm. 1997, 29, 497–511. [Google Scholar]
- Carlucci, M.J.; Ciancia, M.; Matulewicz, M.C.; Cerezo, A.S.; Damonte, E.B. Antiherpetic activity and mode of action of natural carrageenans of diverse structural types. Antiviral Res. 1999, 43, 93–102. [Google Scholar]
- Duarte, M.E.R.; Noseda, D.G.; Noseda, M.D.; Tulio, S.; Pujol, C.A.; Damonte, E.B. Inhibitory effect of sulfated galactans from the marine alga Bostrychia montagnei on herpes simplex virus replication in vitro. Phytomedicine 2001, 8, 53–58. [Google Scholar] [CrossRef]
- Romanos, M.T.V.; Andrada-Serpa, M.J.; Mourão, P.A.S.; Yoneshigue-Valentin, Y.; Costa, S.S.; Pereira, M.S.; Miranda, M.M.F.S.; Gonçalves, J.L.S.; Wigg, M.D. A sulphated fucan from the Laminaria abyssalis inhibits the human T cell lymphotropic virus type 1-induced syncytium formation in HeLa cells. Antivir. Chem. Chemother. 2002, 13, 219–221. [Google Scholar]
- Duarte, M.E.R.; Cauduro, J.P.; Noseda, D.G.; Noseda, M.D.; Gonçalves, A.G.; Pujol, C.A.; Damonte, E.B.; Cerezo, A.S. The structure of the agaran sulfate from Acanthophora spicifera (Rhodomelaceae, Ceramiales) and its antiviral activity. Relation between structure and antiviral activity in agarans. Carbohydr. Res. 2004, 339, 335–347. [Google Scholar] [CrossRef]
- Talarico, L.B.; Pujol, C.A.; Zibetti, R.G.; Faria, P.C.; Noseda, M.D.; Duarte, M.E.; Damonte, E.B. The antiviral activity of sulfated polysaccharides against dengue virus is dependent on virus serotype and host cell. Antivir. Res. 2005, 66, 103–110. [Google Scholar]
- Wang, H.; Li, Y.L.; Shen, W.Z.; Rui, W.; Ma, X.J.; Cen, Y.Z. Antiviral activity of a sulfoquinovosyldiacylglycerol (SQDG) compound isolated from the green alga Caulerpa racemosa. Bot. Mar. 2007, 50, 185–190. [Google Scholar]
- Chirasuwan, N.; Chaiklahan, R.; Kittakoop, P.; Chanasattru, W.; Ruengjitchatchawalya, M.; Tanticharoen, M.; Bunnag, B. Anti HSV-1 activity of sulphoquinovosyl diacylglycerol isolated from Spirulina platensis. Sci. Asia 2009, 35, 137–141. [Google Scholar]
- El-Baroty, G.S.; El-Baz, F.K.; Abd-Elmoein, A.; Abd El Baky, H.H.; Ali, M.M.; Ibrahim, A.E. Evaluation of glycolipids of some egyptian marine algae as a source of bioactive substances. EJEAFChe 2011, 10, 2114–2128. [Google Scholar]
- Mattos, B.B.; Romanos, M.T.V.; Souza, L.M.; Sassaki, G.L.; Barreto-Bergter, E. Glycolipids from macroalgae: potential biomolecules for marine biotechnology? Braz. J. Pharmacogn. 2011, 21, 244–247. [Google Scholar]
- Christophers, J.; Sutton, R.N. Characterisation of acyclovir-resistant and -sensitive clinical herpes simplex virus isolates from an immunocompromised patient. J. Antimicrob. Chemother. 1987, 20, 389–398. [Google Scholar]
- Barreto-Bergter, E.; Sassaki, G.L.; Souza, L.M. Structural analysis of fungal cerebrosides. Front. Microbiol. 2011, 2, 1–11. [Google Scholar]
- Folch, J.; Lees, M.; Sloane-Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Skipski, V.P. Thin layer chromatography of neutral glycolipids. Methods Enzymol. 1975, 35, 396–425. [Google Scholar]
- Kim, Y.H.; Gil, J.H.; Hong, J.; Yoo, J.S. Tandem mass spectrometric analysis of fatty acyl groups of galactolipid molecular species from wheat flour. Microchem. J. 2001, 68, 143–155. [Google Scholar]
- Scoparo, C.T.; Souza, L.M.; Dartora, N.; Sassaki, G.L.; Gorin, P.A.J.; Iacomini, I. Analysis of Camellia sinensis green and black teas via ultra high performance liquid chromatography assisted by liquid-liquid partition and two-dimensional liquid chromatography (size exclusion × reversed phase). J. Chromatogr. A 2012, 1222, 29–37. [Google Scholar]
- Souza, L.M.; Müller-Santos, M.; Iacomini, M.; Gorin, P.A.J.; Sassaki, G.L. Positive- and negative-tandem mass spectrometric fingerprints of lipids from the halophilic archaea Haloarcula marismortui. J. Lipid Res. 2009, 50, 1363–1373. [Google Scholar]
- Benson, A.A.; Daniel, H.; Wiser, R.A. A sulfolipid in plants. Proc. Natl. Acad. Sci. USA 1959, 45, 1582–1587. [Google Scholar]
- Benson, A.A. The plant sulpholipid. Adv. Lipid Res. 1963, 1, 387–394. [Google Scholar]
- Naumann, I.; Klein, B.C.; Bartel, S.J.; Darsow, K.H.; Buchholz, R.; Lange, H.A. Identification of sulfoquinovosyldiacyglycerides from Phaeodactylum tricornutum by matrix-assisted laser desorption/ionization QTrap time-of-flight hybrid mass spectrometry. Rapid Commun. Mass Spectrom. 2011, 25, 2517–2523. [Google Scholar]
- Zhang, X.; Fhaner, C.J.; Ferguson-Miller, S.M.; Reid, G.E. Evaluation of ion activation strategies and mechanisms for the gas-phase fragmentation of sulfoquinovosyldiacylglycerol lipids from Rhodobacter sphaeroides. Int. J. Mass Spectrom. 2012, in press.. [Google Scholar]
- Sassaki, G.L.; Machado, M.J.; Tischer, C.A.; Gorin, P.A.J.; Iacomini, M. Glycosyldiacylglycerolipids from the lichen Dictyonema glabratum. J. Nat. Prod. 1999, 62, 844–847. [Google Scholar]
- Siddantha, A.K.; Ramvat, B.K.; Chauvan, V.D.; Achari, B.; Dutta, P.K.; Pakrashi, S.C. Sulphoglycolipid from the green alga Enteromorpha flexuosa (Wulf). J. Agric. Bot. Mar. 1991, 34, 365–367. [Google Scholar]
- Siddhanta, A.K.; Mody, K.H.; Ramavat, B.K.; Chauan, V.D.; Sharma, M.; Garg, S.H. Characterization of Sulphonoglycolipid from the Red Alga Laurencia pedicularoides. Bot. Mar. 1995, 38, 329–331. [Google Scholar]
- Sassaki, G.L.; Riter, D.S.; Filho, A.S.P.; Guerrini, M.; Lima, M.A.; Cosentino, C.; Souza, L.M.; Cipriani, T.R.; Rudd, T.R.; Nader, H.B.; et al. A robust method to quantify low molecular weight contaminants in heparin: detection of tris(2-n-butoxyethyl) phosphate. Analyst 2011, 136, 2330–2338. [Google Scholar]
- Markoulatos, P.; Georgopoulou, A.; Siafakas, N.; Plakokefalos, E.; Tzanakaki, G.; Kourea-Kremastinou, J. Laboratory diagnosis of common herpesvirus infections of the central nervous system by a multiplex PCR assay. J. Clin. Microbiol. 2001, 39, 4426–4432. [Google Scholar]
- Borenfreund, E.; Puerner, J.A. Toxicity determined in vitro by morphological alterations and neutral red absorption. Toxicol. Lett. 1985, 24, 119–124. [Google Scholar]
- Reed, L.J.; Muench, H. A simple method of estimating 50 per cent end-points. Am. J. Hyg. 1938, 27, 493–497. [Google Scholar]
- Nishimura, T.; Toku, K.; Fukuyasu, H. Antiviral compounds. XII. Antiviral activity of aminohydrazones of alkoxyphenyl substituted carbonyl compounds against influenza virus in eggs and mice. Kitasato Arch. Exp. Med. 1977, 50, 39–46. [Google Scholar]
Supplementary Files
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
De Souza, L.M.; Sassaki, G.L.; Romanos, M.T.V.; Barreto-Bergter, E. Structural Characterization and Anti-HSV-1 and HSV-2 Activity of Glycolipids from the Marine Algae Osmundaria obtusiloba Isolated from Southeastern Brazilian Coast. Mar. Drugs 2012, 10, 918-931. https://doi.org/10.3390/md10040918
De Souza LM, Sassaki GL, Romanos MTV, Barreto-Bergter E. Structural Characterization and Anti-HSV-1 and HSV-2 Activity of Glycolipids from the Marine Algae Osmundaria obtusiloba Isolated from Southeastern Brazilian Coast. Marine Drugs. 2012; 10(4):918-931. https://doi.org/10.3390/md10040918
Chicago/Turabian StyleDe Souza, Lauro M., Guilherme L. Sassaki, Maria Teresa Villela Romanos, and Eliana Barreto-Bergter. 2012. "Structural Characterization and Anti-HSV-1 and HSV-2 Activity of Glycolipids from the Marine Algae Osmundaria obtusiloba Isolated from Southeastern Brazilian Coast" Marine Drugs 10, no. 4: 918-931. https://doi.org/10.3390/md10040918