Therapeutic Potential of Non-Psychotropic Cannabidiol in Ischemic Stroke

{kind=link}
{kind=link}
Abstract
:1. Introduction
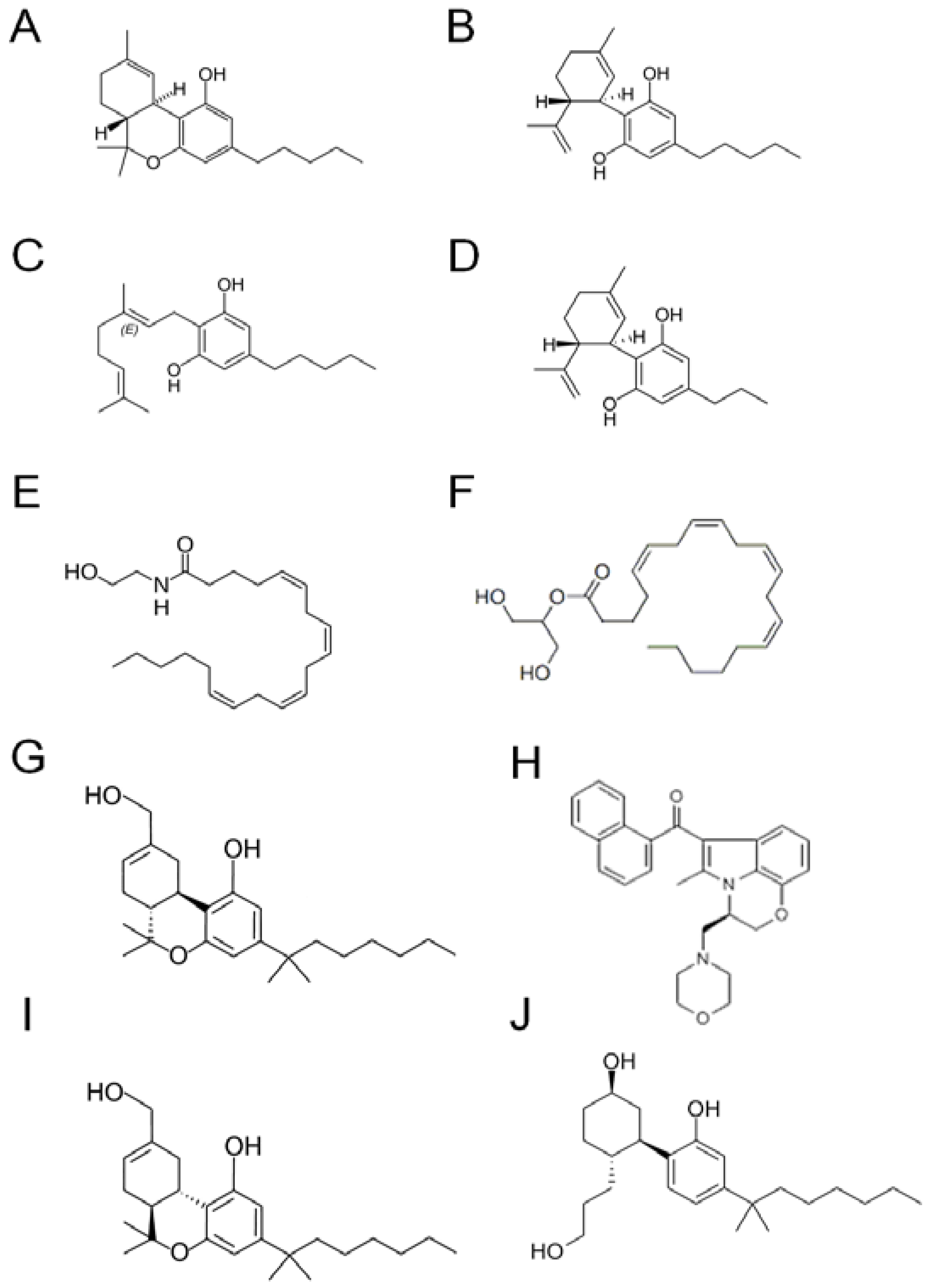
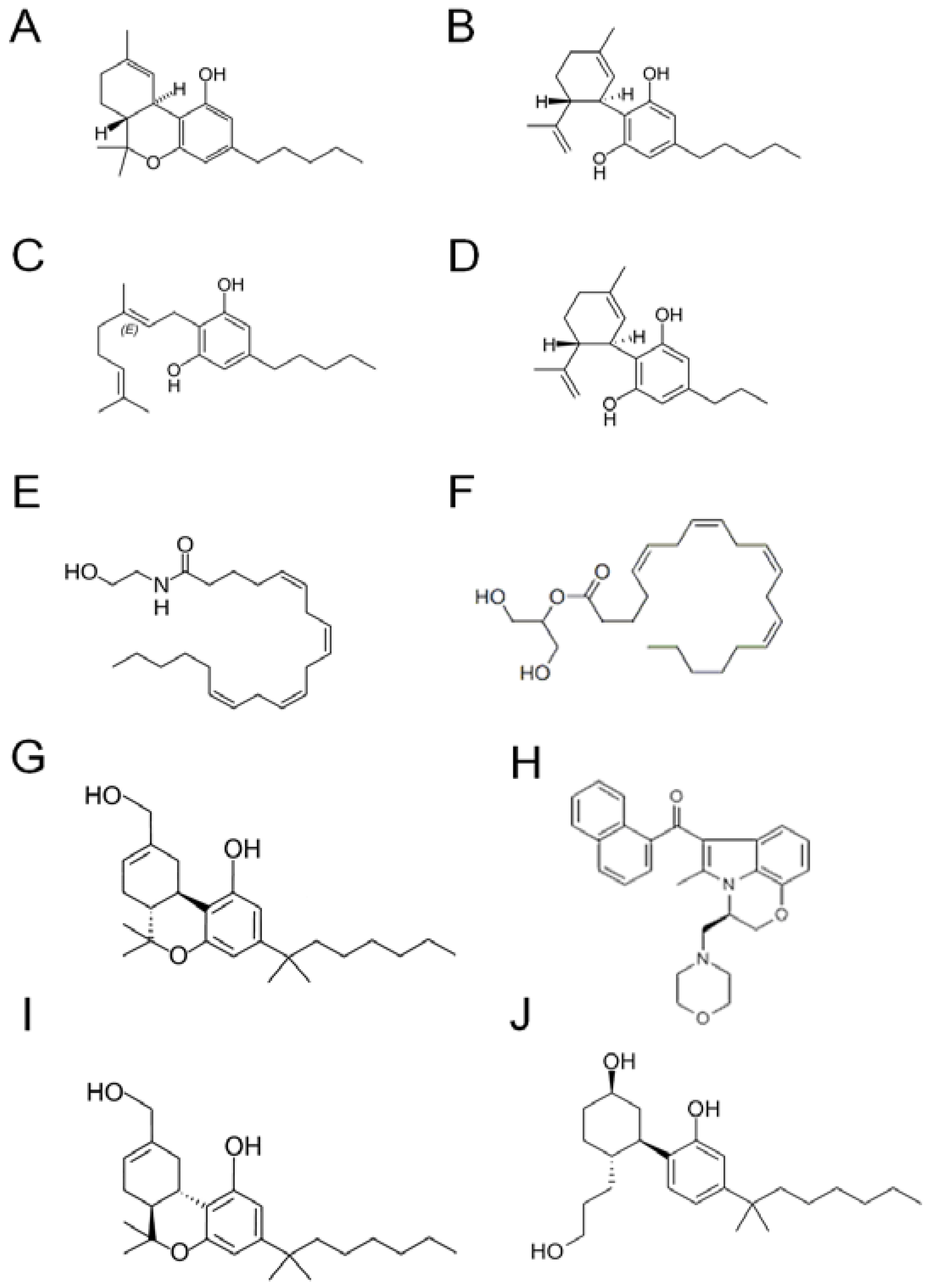
2. Pharmacology of CBD in Ischemic Stroke
2.1. Cannabinoid Receptors and Endocannabinoid System
2.2. Serotonin 5-HT1A Receptor-Dependent Mechanism
2.3. Potent Anti-Oxidant Mechanism
2.4. Cerebroprotective Effect without the Development of Tolerance
2.5. Potent Anti-Inflammatory Effect
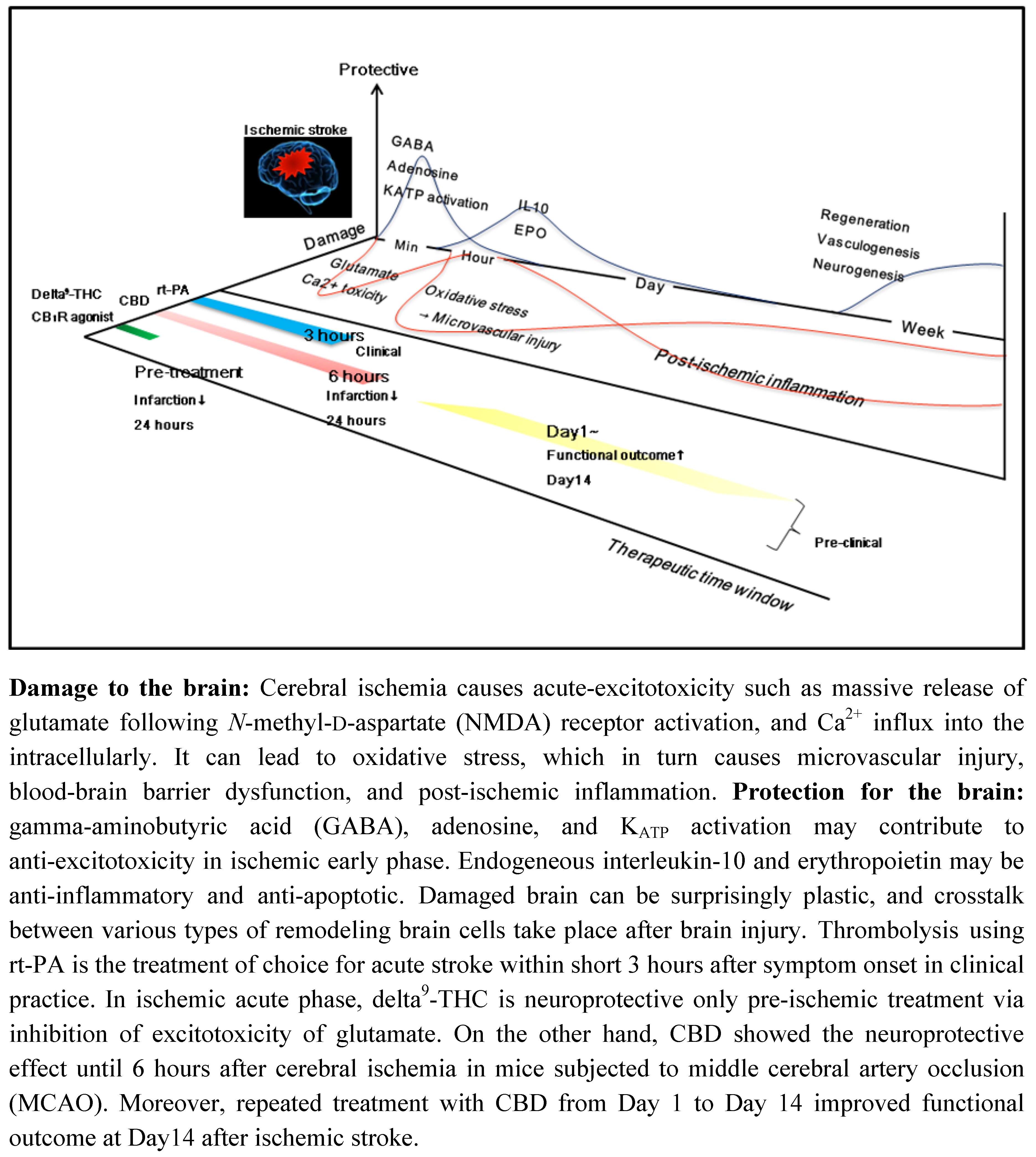
3. Therapeutic Time Window of CBD and Other Cannabinoids in Ischemic Stroke
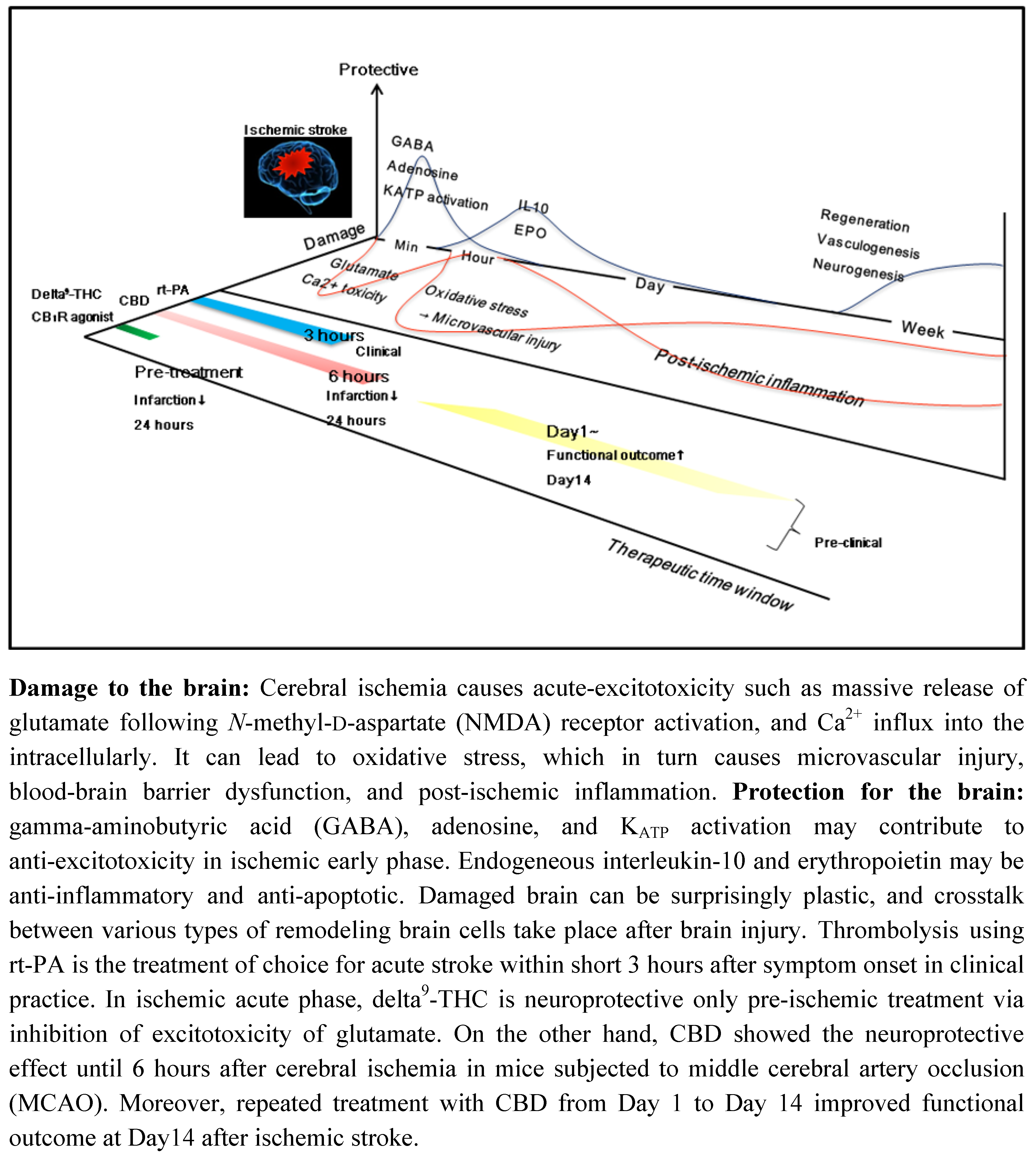
3.1. CBD and Delta9-THC
3.2. Synthetic CB1 Receptor Agonist, HU210 and R(+)-WIN55,212-2
3.3. A Synthetic Non-Competitive NMDA Antagonist, HU-211, Dexanabinol
4. Therapeutic Possibility of CBD
5. Conclusions
Acknowledgements
References
- Wiley, J.L.; Martin, B.R. Cannabinoid pharmacology: implications for additional cannabinoid receptor subtypes. Chem. Phys. Lipids 2002, 121, 57–63. [Google Scholar]
- Martin, B.R.; Compton, D.R.; Thomas, B.F.; Prescott, W.R.; Little, P.J.; Razdan, R.K.; Johnson, M.R.; Melvin, L.S.; Mechoulam, R.; Ward, S.J. Behavioral, biochemical, and molecular modeling evaluations of cannabinoid analogs. Pharmacol. Biochem. Behav. 1991, 40, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Fujiwara, M. Characteristics of abnormal behavior induced by delta9-tetrahydrocannabinol in rats. Nippon. Yakurigaku. Zasshi. 2001, 117, 35–41. [Google Scholar]
- Mishima, K.; Egashira, N.; Hirosawa, N.; Fujii, M.; Matsumoto, Y.; Iwasaki, K.; Fujiwara, M. Characteristics of learning and memory impairment induced by delta9-tetrahydrocannabinol in rats. Jpn. J. Pharmacol. 2001, 87, 297–308. [Google Scholar]
- Nagai, H.; Egashira, N.; Sano, K.; Ogata, A.; Mizuki, A.; Mishima, K.; Iwasaki, K.; Shoyama, Y.; Nishimura, R.; Fujiwara, M. Antipsychotics improve delta9-tetrahydrocannabinol-induced impairment of the prepulse inhibition of the startle reflex in mice. Pharmacol. Biochem. Behav. 2006, 84, 330–336. [Google Scholar]
- Egashira, N.; Matsuda, T.; Koushi, E.; Mishima, K.; Iwasaki, K.; Shoyama, Y.; Fujiwara, M. Involvement of 5-hydroxytriptamine1A receptors in delta9-tetrahydrocannabinol-induced catalepsy-like immobilization in mice. Eur. J. Pharmacol. 2006, 550, 117–122. [Google Scholar]
- Klein, T.W.; Neuton, C.A.; Friedman, H. Cannabinoids and the immune system. Pain. Res. Manag. 2001, 6, 95–101. [Google Scholar]
- Maresz, K.; Carrier, E.J.; Ponomarev, E.D.; Hilard, C.J.; Dittel, B.N. Modulation of the cannabinoid CB2 receptor in microglial cells in response to inflammatory stimuli. J. Neurochem. 2005, 95, 437–445. [Google Scholar]
- Pertwee, R.G. Reseptors and channels targeted by synthetic cannabinoid receptor agonists and antagonists. Curr. Med. Chem. 2010, 17, 1360–1381. [Google Scholar]
- Izzo, A.A.; Borrelli, F.; Capasso, R.; Di Marzo, V.; Mechoulam, R. Non-psychotropic plant cannabinoids: new therapeutic opportunities from an ancient herb. Trends. Pharmacol. Sci. 2009, 30, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Parker, L.A.; Gallily, R. Cannabidiol: an overview of some pharmacological aspects. J. Clin. Phamacol. 2002, 42, 11–19. [Google Scholar]
- Iuvone, T.; Esposito, G.; Esposito, R.; Santamaria, R.; Di Rosa, M.; Izzo, A.A. Neuroprotective effect of cannabidiol, a non-psychoactive component from Cannabis sativa, on beta-amyloid-induced toxicity in PC12 cells. J. Neurochem. 2004, 89, 134–141. [Google Scholar] [CrossRef] [PubMed]
- Braida, D.; Pegorini, S.; Arcidiacono, M.V.; Consalez, G.G.; Croci, L.; Sala, M. Post-ischemic treatment with cannabidiol prevents electroencephalographic flattening, hyperlocomotion and neuronal injury in gerbils. Neurosci. Lett. 2003, 346, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Hampson, A.J.; Grimaldi, M.; Lolic, M.; Wink, D.; Rosenthal, R.; Axelrod, J. Neuroprotective antioxidants from marijuana. Ann. N.Y. Acad. Sci. 2000, 899, 274–282. [Google Scholar]
- Carrier, E.J.; Auchampach, J.A.; Hillard, C.J. Inhibition of an equilibrative nucleoside transporter by cannabidiol: a mechanism of cannabinoid immunosuppression. Proc. Natl. Acad. Sci. USA 2006, 103, 7895–7900. [Google Scholar]
- Ryberg, E.; Larsson, N.; Sjögren, S.; Hjorth, S.; Hermansson, N.O.; Leonova, J.; Elebring, T.; Nilsson, K.; Drmota, T.; Greasley, P.J. The orphan receptor GPR55 is a novel cannabinoid receptor. Br. J. Pharmacol. 2007, 152, 1092–1101. [Google Scholar]
- Molina-Holgado, E.; Vela, J.M.; Arevalo-Martin, A.; Almazan, G.; Molina-Holgado, F.; Borrell, J.; Guaza, C. Cannabinoids promote oligodendrocyte progenitor survival: involvement of cannabinoid receptors and phosphotidylinositol-3 kinase/Akt signaling. J. Neurosci. 2002, 22, 9742–9753. [Google Scholar]
- Leker, R.R.; Gai, N.; Mechoulam, R.; Ovadia, H. Drug-induced hypothermia reduces ischemic damage: effects of the cannabinoid HU-210. Stroke. 2003, 34, 2000–2006. [Google Scholar]
- Pertwee, R.G.; Ross, R.A. Cannabinoid receptors and their liganads. Prostaglandins. Leukot. Essent. Fatty. Acids 2002, 66, 101–121. [Google Scholar]
- Thomas, A.; Baillie, G.L.; Phillips, A.M.; Razdan, R.K.; Ross, R.A.; Pertwee, R.G. Cannabidiol displays unexpectedly high potency as an antagonist of CB1 and CB2 receptor agonist. Br. J. Pharmacol. 2007, 150, 613–623. [Google Scholar]
- Castillo, A.; Tolón, M.R.; Fernández-Ruiz, J.; Romero, J.; Martinez-Orgado, J. The neuroprotective effect of cannabidiol in an in vitro model of newborn hypoxic-ischemic brain damage in mice is mediated by CB(2) and adenosine receptors. Neurobiol. Dis. 2010, 37, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Mishima, K.; Hayakawa, K.; Abe, K.; Ikeda, T.; Egashira, N.; Iwasaki, K.; Fujiwara, M. Cannabidiol prevents cerebral infarction via a serotonergic 5-hydroxytriptamine1A receptor-dependent mechanism. Stroke. 2005, 36, 1077–1082. [Google Scholar]
- Watanabe, K.; Kayano, Y.; Yamamoto, I.; Yoshimura, H. Inhibition of anandamide amidase activity in mouse brain microsomes by cannabinoids. Biol. Pharm. Bull. 1996, 19, 1109–1111. [Google Scholar]
- Bisogno, T.; Hanus, L.; De Petrocellis, L.; Tchilibon, S.; Ponde, D.E.; Brandi, I.; Moriello, A.S.; Davis, J.B.; Mechoulam, R.; Di Marzo, V. Molecular targets for cannabidiol and its synthetic analogues: effect on vanilloid VR1 receptors and on the cellular uptake and enzymatic hydrolysis of anandamide. Br. J. Pharmacol. 2001, 134, 845–852. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.C.; Lo, S.W.; Hsu, K.S. Presynaptic mechanisms underlying cannabinoid inhibition of excitatory synaptic transmission in rat striatal neurons. J. Physiol. 2001, 532, 731–748. [Google Scholar]
- Lévénés, C.; Daniel, H.; Soubrié, P.; Crépel, F. Cannabinoids decrease excitatory synaptic transmission and impair long-term depression in rat cerebellar Purkinje cells. J. Physiol. 1998, 510, 867–879. [Google Scholar]
- Shen, M.; Piser, T.M.; Sybold, V.S.; Thayer, S.A. Cannabinoid receptor agonists inhibit glutamaterigic synaptic transmission in rat hippocampal cultures. J. Neurosci. 1996, 16, 4322–4334. [Google Scholar]
- Köfalvi, A.; Vizi, E.S.; Ledent, C.; Sperlágh, B. Cannabinoids inhibit release of [3H]glutamate from rodent hippocampal synaptosomes via a novel CB1 receptor-independent action. Eur. J. Neurosci. 2003, 18, 1973–1978. [Google Scholar]
- Fowler, C.J. Plant-derived, synthetic and endogenous cannabinoids as neuroprotective agents. Non-psychoactive cannabinoids ‘entourage’ compounds and inhibitors N-acyl ethanolamine breakdown as therapeutic strategies to avoid psychotropic effects. Brain. Res. Brain. Res. Rev. 2003, 41, 26–43. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Mishima, K.; Nozako, M.; Hazekawa, M.; Irie, K.; Fujioka, M.; Orito, K.; Abe, K.; Hasebe, N.; Egashira, N.; Iwasaki, K.; Fujiwara, M. Delayed treatment with cannabidiol has a cerebroprotective action via a cannabinoid receptor-independent myeloperoxidase-inhibiting mechanism. J. Neurochem. 2007, 102, 1488–1496. [Google Scholar]
- Karnoil, I.G.; Shirakawa, I.; Kasinski, N.; Pfeferman, A.; Carlini, E.A. Cannabidiol interferes with the effects of delta9-tetrahydrocannabinol in man. Eur. J. Pharmacol. 1974, 28, 172–177. [Google Scholar]
- Zuardi, A.W. Cannabidiol: from an inactive cannabinoid to a drug with wide spectrum of action. Rev. Bras. Psiquiatr. 2008, 30, 271–280. [Google Scholar]
- Moreira, F.A.; Aguiar, D.C.; Campos, A.C.; Lisboa, S.F.; Terzian, A.L.; Resstel, L.B.; Guimarães, F.S. Antiaversive effects of cannabinoids: is the periaqueductal gray involved? Neural. Plast. 2009. [Google Scholar]
- Campos, A.C.; Guimarães, F.S. Involvement of 5HT1A receptors in the anxiolytic-like effects of cannabidiol injected into the dorsolateral periaqueductal gray of rats. Psychopharmacology (Berl). 2008, 199, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Gardner, C.R. Resent developments in 5HT-related pharmacology of animal models of anxiety. Pharmacol. Biochem. Behav. 1986, 24, 1479–1485. [Google Scholar]
- Eltze, M.; Boer, R.; Sanders, K.H.; Kolassa, N. Vasodilatation elicited by 5-HT1A receptor agonists in constant-pressure-perfused rat kidney is mediated by blockade of alpha 1A-adrenoceptors. Eur. J. Pharmacol. 1991, 202, 33–44. [Google Scholar]
- Verbeuren, T.J. Vasodilator effect of tertatolol in isolated perfused rat kidneys: involvement of endothelial 5-HT1A receptors. Cardiology. 1993, 1, 5–9. [Google Scholar]
- Semkova, I.; Wolz, P.; Krieglstein, J. Neuroprotective effect of 5-HT1A receptor agonist, BAY X 3702 demonstrated in vitro and in vivo. Eur. J. Pharmacol. 1998, 359, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.H.; Svensson, E.; Dewael, Y.; Grillner, S. 5-HT inhibits N-type but not L-type Ca (2+) channels via 5-HT1A receptors in lamprey spinal neurons. Eur. J. Neurosci. 2003, 18, 2919–2924. [Google Scholar]
- Madhavan, L.; Freed, W.J.; Anantharam, V.; Kanthasamy, A.G. 5-hydroxytryptamine 1A receptor activation protects against N-methyl-D-aspartate-induced apoptotic cell death in striatal and mesencephalic cultures. J. Pharmacol. Exp. Ther. 2003, 304, 913–923. [Google Scholar]
- Russo, E.B.; Burnett, A.; Hall, B.; Parker, K.K. Agonistic properties of cannabidiol at 5-HT1a receptors. Neurochem. Res. 2005, 30, 1037–1043. [Google Scholar]
- Magen, I.; Avraham, Y.; Ackerman, Z.; Vorobiev, L.; Mechoulam, R.; Berry, E.M. Cannabidiol ameliorates cognitive and motor impairments in bile-duct ligated mice via 5-HT1A receptor activation. Br. J. Pharmacol. 2010, 159, 950–957. [Google Scholar]
- Hampson, A.J.; Grimaldi, M.; Axelrod, J.; Wink, D. Cannabidiol and (-) Delta9-tetrahydrocannabinol are neuroprotective antioxidants. Proc. Natl. Acad. Sci. USA 1998, 95, 8268–8273. [Google Scholar]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. Lebnsm-Wiss. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Hayakawa, K.; Mishima, K.; Nozako, M.; Ogata, A.; Hazekawa, M.; Liu, A.X.; Fujioka, M.; Abe, K.; Hasebe, N.; Egashira, N.; Iwasaki, K.; Fujiwara, M. Repeated treatment with cannabidiol but not Delta9-tetrahydrocannabinol has a neuroprotective effect without the development of tolerance. Neuropharmacology 2007, 52, 1079–1087. [Google Scholar]
- Hamelink, C.; Hampson, A.; Wink, D.A.; Eiden, L.E.; Eskay, R.L. Comparison of cannabidiol, antioxidants, and diuretics in reversing binge ethanol-induced neurotoxicity. J. Pharmacol. Exp. Ther. 2005, 314, 780–788. [Google Scholar] [CrossRef] [PubMed]
- Abood, M.E.; Martin, B.R. Neurobiology of marijuana abuse. Trend. Pharmacol. Sci. 1992, 13, 201–206. [Google Scholar]
- Sim, L.J.; Hampson, R.E.; Deadwyler, S.A.; Childers, S.R. Effects of chronic treatment with D9-tetrahydrocannabinol on cannabinoid-stimulated [35S]GTPgS autoradiography in rat brain. J. Neurosci. 1996, 1, 8057–8066. [Google Scholar]
- Paquett, J.J.; Wang, H.Y.; Bakshi, K.; Olmstead, M.C. Cannabinoid-induced tolerance is associated with a CB1 receptor G protein coupling switch that is prevented by ultra-low dose rimonabant. Behav. Pharmacol. 2007, 18, 767–776. [Google Scholar] [CrossRef] [PubMed]
- Blair, R.E.; Deshpande, L.S.; Sombati, S.; Elphick, M.R.; Martin, B.R.; DeLorenzo, R.J. Prolonged exposure to WIN55, 212-2 causes downregulation of the CB1 receptor and the development of tolerance to its anticonvulsant effects in the hippocampal neuronal culture of acquired epilepsy. Neuropharmacology 2009, 57, 208–218. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Narimatsu, S.; Yamamoto, I.; Yoshimura, H. Difference in tolerance development o hypothermia and pentobarbital-induced sleep prolongating effect of 11-hydroxy-delta 8-tetrahydrocannabinol and 11-oxo-delta 8-tetrahydrocannabinol in mice. Eur. J. Pharmacol. 1982, 77, 53–56. [Google Scholar]
- Watanabe, K.; Yamamoto, I.; Yoshimura, H. Development of tolerance and cross-tolerance to the cataleptogenic effects of delta 8-tetrahydrocannabinol and 11-hydroxy-delta 8-tetrahydrocannabinol in mice. Eur. J. Pharmacol. 1983, 94, 349–351. [Google Scholar]
- Sim-Selly, I.J.; Martin, B.R. Effect of chronic administration of R-(+)-[2,3-dyhydro-5methyl-3-[(morpholinyl)methyl]pyrrolo[1,2,3-de]-1,4-benzoxazinyl]-(1-naphthalenyl-9-methanone mesylate (WIN55,212-2) or delta(9)-tetrahydrocannabinol on cannabinoid receptor adaptation in mice. J. Pharmacol. Exp. Ther. 2002, 303, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Renyi, L.; Moller, K.A.; Ensler, K.; Evenden, J. The non-competitive NMDA receptor antagonist (+)MK801 counteracts the long-lasting attenuation of the hypothermic response induced by acute doses of 8-OH-DPAT in the rat. Neuropharmacology 1992, 31, 1265–1268. [Google Scholar]
- Young, A.H.; McShane, R.; Park, S.B.; Cowen, P.J. Buspirone-induced hypothermia in nomal male volunteers. Biol. Psychiatr. 1993, 34, 665–666. [Google Scholar]
- Esposito, G.; Scuderi, C.; Savani, C.; Steardo, L., Jr.; De Filippis, D.; Cottone, P.; Iuvone, T.; Cuomo, V.; Steardo, L. Cannabidiol in vivo blunts beta-amyloid induced neuroinflammation by suppressing IL-1beta and iNOS expression. Br. J. Pharmacol. 2007, 151, 1272–1279. [Google Scholar] [PubMed]
- McHugh, D.; Tanner, C.; Mechoulam, R.; Pertwee, R.G.; Ross, R.A. Inhibition of human neutrophil chemotaxis by endogenous cannabinoids and psytocannabinoids: evidence for a site distinct from CB1 and CB2. Mol. Pharmacol. 2008, 73, 441–450. [Google Scholar] [PubMed]
- Hayakawa, K.; Mishima, K.; Irie, K.; Hazekawa, M.; Mishima, S.; Fujioka, M.; Orito, K.; Egashira, N.; Katsurabayashi, S.; Takasaki, K.; Iwasaki, K.; Fujiwara, M. Cannabidiol prevents a post-ischemic injury progressively induced by cerebral ischemia via a high-mobility group box1-inhibiting mechanism. Neuropharmacology 2008, 55, 1280–1286. [Google Scholar]
- Bullock, R.; Zauner, A.; Myseros, J.S.; Marmarou, A.; Woodward, J.J.; Young, H.F. Evidence for prolonged release of excitatory amino acids in severe human head trauma. Relationship to clinical events. Ann. N. Y. Acad. Sci. 1995, 765, 290–297. [Google Scholar] [CrossRef] [PubMed]
- Obrenovitch, T.P.; Richards, D.A. Extracellular neurotransmitter changes in cerebral ischaemia. Cerebrovasc. Brain. Metab. Rev. 1995, 7, 1–54. [Google Scholar]
- Obrenovitch, T.P. The ischaemic penumbra: twenty years on. Cerebrovasc. Brain. Metab. Rev. 1995, 7, 297–323. [Google Scholar]
- Martinez-Tica, J.F.; Zornow, M.H. Effects of adenosine agonists and an antagonist on excitatory transmitter release from the ischemic rabbit hippocampus. Brain. Res. 2000, 872, 110–115. [Google Scholar]
- Lakhan, S.E.; Kirchgessner, A.; Hofer, M. Inflammatory mechanisms in ischemic stroke: therapeutic approaches. J. Transl. Med. 2009, 7, 97. [Google Scholar]
- Chen, Y.; Swanson, R.A. Astrocytes and brain injury. J. Cereb. Blood. Flow. Metab. 2003, 23, 137–149. [Google Scholar]
- Chopp, M.; Zhang, Z.G.; Jiang, Q. Neurogenesis, angiogenesis, and MRI induces of functional recovery from stroke. Stroke. 2007, 38, 827–831. [Google Scholar] [CrossRef] [PubMed]
- Lo, E.H. A new penumbra: transitioning from injury into repair after stroke. Nat. Med. 2008, 14, 497–500. [Google Scholar]
- Sun, Y.; Jin, K.; Xie, L.; Childs, J.; Mao, X.O.; Logvinova, A.; Greenberg, D.A. VEGF-induced neuroprotection, neurogenesis, and angiogenesis after focal cerebral ischemia. J. Clin. Invest. 2003, 11, 1843–1851. [Google Scholar]
- Zhang, Z.G.; Zhang, L.; Jiang, Q.; Zhang, R.; Davies, K.; Powers, C.; Bruggen, N.; Chopp, M. VEGF enhances angiogenesis and promotes blood-brain barrier leakage in the ischemic brain. J. Clin. Invest. 2000, 106, 829–838. [Google Scholar]
- Minger, S.L.; Ekonomou, A.; Carta, E.M.; Chinoy, A.; Perry, R.H.; Ballard, C.G. Endogeneous neurogenesis in the human brain following cerebral ischemia. Regen. Med. 2007, 2, 69–74. [Google Scholar]
- Pérez de la Ossa, N.; Dávalos, A. Neuroprotection in cerebral infarction: the opportunity of new studies. Cerebrovasc. Dis. 2007, 24, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Hayakawa, K.; Irie, K.; Sano, K.; Watanabe, T.; Higuchi, S.; Enoki, M.; Nakano, T.; Harada, K.; Ishikane, S.; Ikeda, T.; Fujioka, M.; Orito, K.; Iwasaki, K.; Mishima, K.; Fujiwara, M. Therapeutic time window of cannabidiol treatment on delayed ischemic damage via high-mobility group box1-inhibiting mechanism. Biol. Pharm. Bull. 2009, 32, 1538–1544. [Google Scholar]
- Nagayama, T.; Sinor, A.D.; Simon, R.P.; Chen, J.; Graham, S.H.; Jin, K.; Greenberg, D.A. Cannabinoids and neuroprotection in global and focal cerebral ischemia and in neuronal cultures. J. Neurosci. 1999, 19, 2987–2995. [Google Scholar]
- Shohami, E.; Novikov, M.; Mechoulam, R. A nonpsychotropic cannabinoid, HU-211, has cerebroprotective effects after closed head injury in the rat. J. Neurotrauma 1993, 10, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Shohami, E.; Novikov, M.; Bass, R. Long-term effect of HU-211, a novel non-competitive NMDA antagonist, on motor and memory functions after closed head injury in the rat. Brain. Res. 1995, 674, 55–62. [Google Scholar] [CrossRef] [PubMed]
- Belayev, L.; Busto, R.; Watson, B.D.; Ginsberg, M.D. Post-ischemic administration of HU211, a novel non-competitive NMDA antagonist, protects against blood-brain barrier disruption in photochemical cortical infarction in rats: a quantitative study. Brain. Res. 1995, 702, 266–270. [Google Scholar] [CrossRef] [PubMed]
- Lavie, G.; Teichner, A.; Shohami, E.; Ovadia, H.; Leker, R.R. Long term cerebroprotective effects of dexanabinol in a model of focal cerebral ischemia. Brain. Res. 2001, 901, 195–201. [Google Scholar]
- Collin, C.; Davies, P.; Mutiboko, I.K.; Ratcliffe, S. Sativex Spasticity in MS Study Group. Eur. J. Neurol. 2007, 14, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Selvarajah, D.; Gandhi, R.; Emery, C.J.; Tesfaya, S. Randamized placebo-controlled double-blind clinical trial of cannabis-based medicinal product (Sativex) in painful diabetic neuropathy: depression is a major confounding factor. Diabetes Care 2010, 33, 128–130. [Google Scholar]
- Ryan, D.; Drysdale, A.J.; Lafourcade, C.; Pertwee, R.G.; Platt, B. Cannabidiol targets mitochondria to regulate intracellular Ca2+ levels. J. Neurosci. 2009, 29, 2053–2063. [Google Scholar] [PubMed]
- Knoller, N.; Levi, L.; Shoshan, I.; Reichenthal, E.; Razon, N.; Rappaport, Z.H.; Biegon, A. Dexanabinol (HU-211) in the treatment of severe closed head injury: a randomized, placebo-controlled, phase Ⅱ clinical trial. Crit. Care. Med. 2002, 30, 548–554. [Google Scholar] [CrossRef] [PubMed]
- Mechoulam, R.; Hanus, L. The cannabinoids: an overview. Therapeutic implications in vomiting and nausea after cancer chemotherapy, in appetite promotion, in multiple sclerosis and in neuroprotection. Pain. Res. Manag. 2001, 6, 67–73. [Google Scholar] [PubMed]
- Slieker, F.J.; Kompanje, E.J.; Murray, G.D.; Ohman, J.; Stocchetti, N.; Teasdale, S.G.; Maas, A.I. SAPHIR and Pharmos TBI Investigators. Neurosurgery 2008, 62, 1321–1329. [Google Scholar] [PubMed]
- Tomillero, A.; Moral, M.A. Gateways to clinical trials. Methods. Find. Exp. Clin. Pharmacol. 2010, 32, 193–215. [Google Scholar]
- Fujiwara, M.; Egashira, N. New perspectives in the studies on endocannabinoid and cannabis: abnormal behaviors associate with CB1 cannabinoid receptor and development of therapeutic application. J. Pharmacol. Sci. 2004, 96, 362–366. [Google Scholar]
- Egashira, N.; Matsuda, T.; Koushi, E.; Mishima, K.; Iwasaki, K.; Shoyama, Y.; Fujiwara, M. Involvement of 5-hydroxytriptamine1A receptors in Delta9-tetrahydrocannabinol-induced catalepsy-like immobilization in mice. Eur. J. Pharmacol. 2006, 550, 117–122. [Google Scholar]
- Egashira, N.; Ishigami, N.; Mishima, K.; Iwasaki, K.; Oishi, R.; Fujiwara, M. Delta9-Tetrahydrocannabinol-induced cognitive deficits are reversed by olanzapine but not haloperidol in rats. Prog. Neuropsychopharmacol. Biol. Psychiatry. 2008, 32, 499–506. [Google Scholar]
- Egashira, N.; Matsuda, T.; Koushi, E.; Higashihara, F.; Mishima, K.; Chidori, S.; Hasebe, N.; Iwasaki, K.; Nishimura, R.; Oishi, R.; Fujiwara, M. Delta (9)-tetrahydrocannabinol prolongs the immobility time in the mouse forced swim test: involvement of cannabinoid CB (1) receptor and serotonergic system. Eur. J. Pharmacol. 2008, 589, 117–121. [Google Scholar]
- Higuchi, S.; Irie, K.; Mishima, S.; Araki, M.; Ohji, M.; Shirakawa, A.; Akitake, Y.; Matsuyama, K.; Mishima, K.; Mishima, K.; Iwasaki, K.; Fujiwara, M. The cannabinoid 1-receptor silent antagonist-2050 attenuates preference for high-fat diet and activated astrocytes in mice. J. Pharmacol. Sci. 2010, 112, 369–372. [Google Scholar]
- Hayakawa, K.; Mishima, K.; Abe, K.; Hasebe, N.; Takamatsu, F.; Yasuda, H.; Ikeda, T.; Inui, K.; Egashira, N.; Iwasaki, K.; Fujiwara, M. Cannabidiol prevents infarction via the non-CB1 cannabinoid receptor mechanism. Neuroreport 2004, 15, 2381–2385. [Google Scholar]
- Mechoulam, R.; Feigenbaum, J.J.; Lander, N.; Segal, M.; Järbe, T.U.; Hiltunen, A.J.; Consroe, P. Enantiomeric cannabinoids: stereospecificity of psychotropic activity. Experientia. 1988, 44, 762–764. [Google Scholar]
- Feigenbaum, J.J.; Bergmann, F.; Richmond, S.A.; Mechoulam, R.; Nadler, V.; Kloog, Y.; Sokolovsky, M. Proc. Natl. Acad. Sci. USA 1989, 86, 9584–9587.
- Eshhar, N.; Striem, S.; Kohen, R.; Tirosh, O.; Biegon, A. Neuroprotective and antioxidant activities of HU-211, a novel NMDA receptor antagonist. Eur. J. Pharmacol. 1995, 283, 19–29. [Google Scholar]
- Shohami, E.; Gallily, R.; Mechoulam, R.; Bass, R.; Ben-Hur, T. Cytokine production in the brain following closed head injury: dexanabinol (HU-211) is a novel TNF-alpha inhibitor and an effective neuroprotectant. J. Neuroimmunol. 1997, 72, 169–177. [Google Scholar]
- Yoles, E.; Belkin, M.; Schwartz, M. HU-211, a nonpsychotropic cannabinoid, produces short- and long-term neuroprotection after optic nerve axotomy. J. Neurotrauma 1996, 13, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Constam, D.B.; Philipp, J.; Malipiero, U.V.; ten Dijke, P.; Schachner, M.; Fontana, A. Differential expression of transforming growth factor-beta 1, -beta 2, and -beta 3 by glioblastoma cells, astrocytes, and microglia. J. Immunology 1992, 148, 1404–1410. [Google Scholar]
- Strauss, S.; Otten, U.; Joggerst, B.; Piuss, K.; Volk, B. Increased levels of nerve growth factor (NGF) protein and mRNA and reactive gliosis following kainic acid injection into the rat striatum. Neurosci. Lett. 1994, 168, 193–196. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an Open Access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Hayakawa, K.; Mishima, K.; Fujiwara, M. Therapeutic Potential of Non-Psychotropic Cannabidiol in Ischemic Stroke. Pharmaceuticals 2010, 3, 2197-2212. https://doi.org/10.3390/ph3072197
Hayakawa K, Mishima K, Fujiwara M. Therapeutic Potential of Non-Psychotropic Cannabidiol in Ischemic Stroke. Pharmaceuticals. 2010; 3(7):2197-2212. https://doi.org/10.3390/ph3072197
Chicago/Turabian StyleHayakawa, Kazuhide, Kenichi Mishima, and Michihiro Fujiwara. 2010. "Therapeutic Potential of Non-Psychotropic Cannabidiol in Ischemic Stroke" Pharmaceuticals 3, no. 7: 2197-2212. https://doi.org/10.3390/ph3072197
APA StyleHayakawa, K., Mishima, K., & Fujiwara, M. (2010). Therapeutic Potential of Non-Psychotropic Cannabidiol in Ischemic Stroke. Pharmaceuticals, 3(7), 2197-2212. https://doi.org/10.3390/ph3072197