Pathogen Phytosensing: Plants to Report Plant Pathogens


Abstract
:1. Introduction
2. Results and Discussion
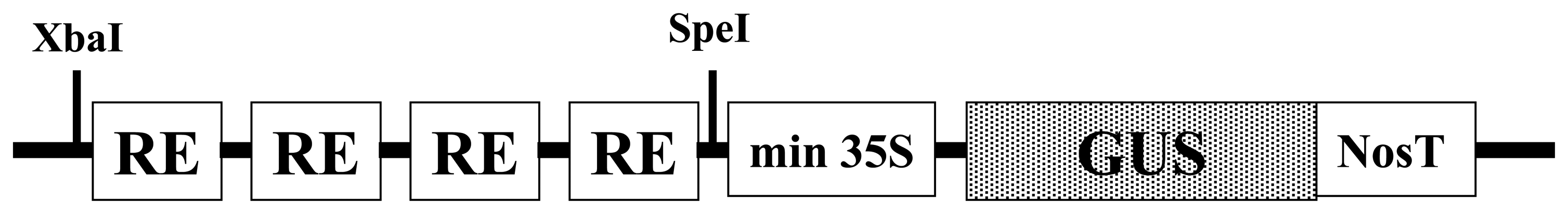
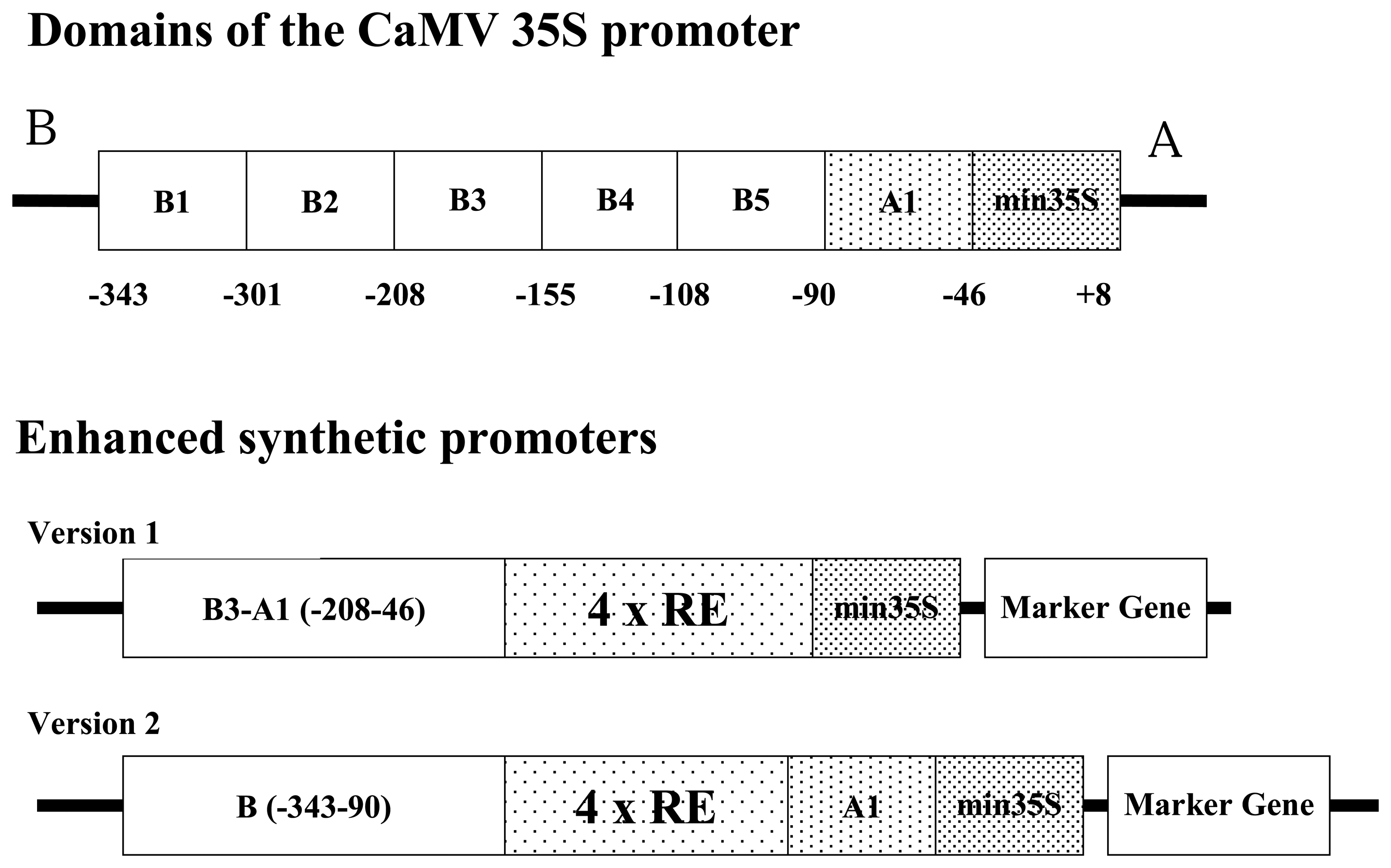
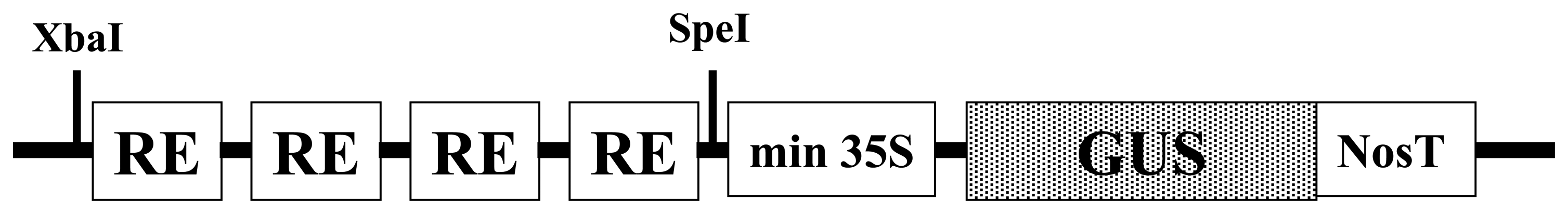
2.1 Construction of synthetic promoters for pathogen phytosensing
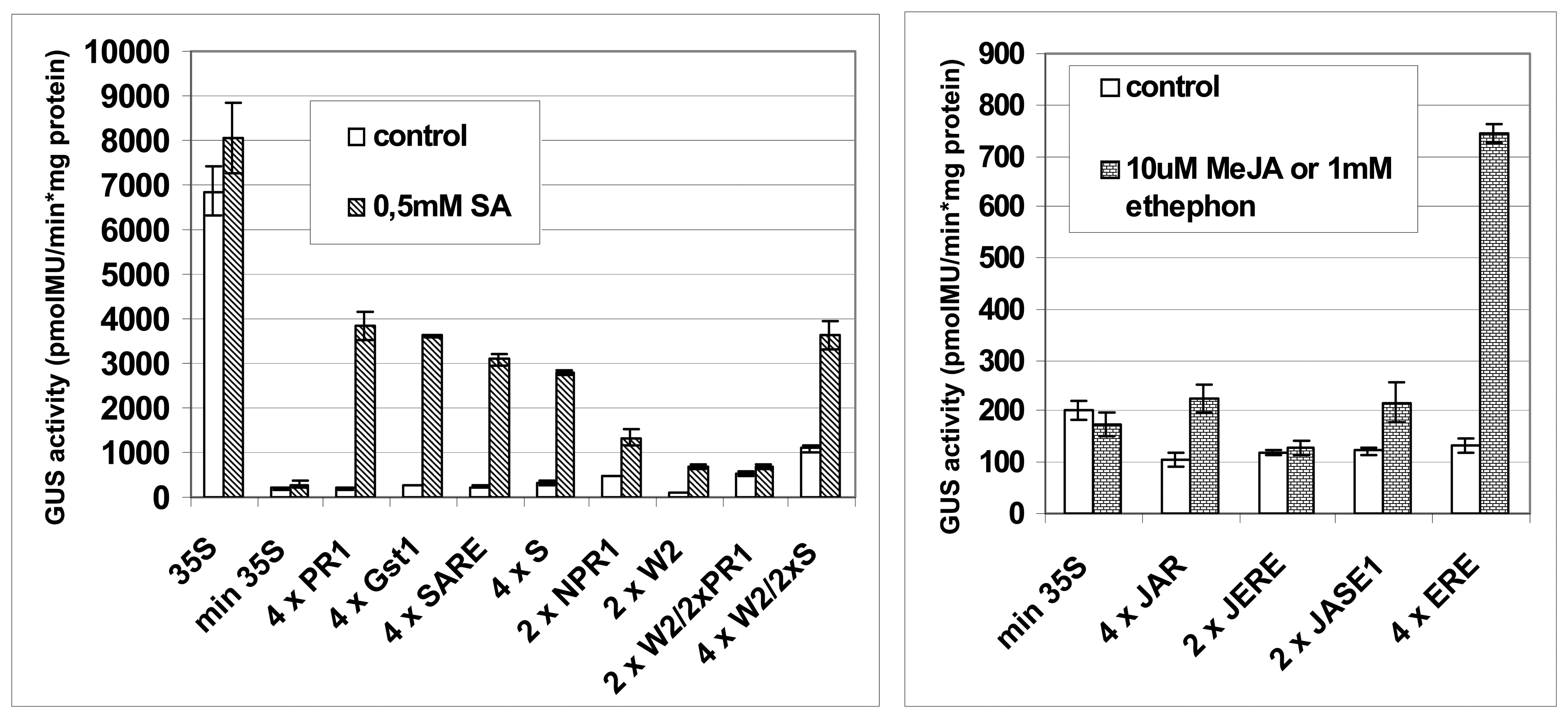
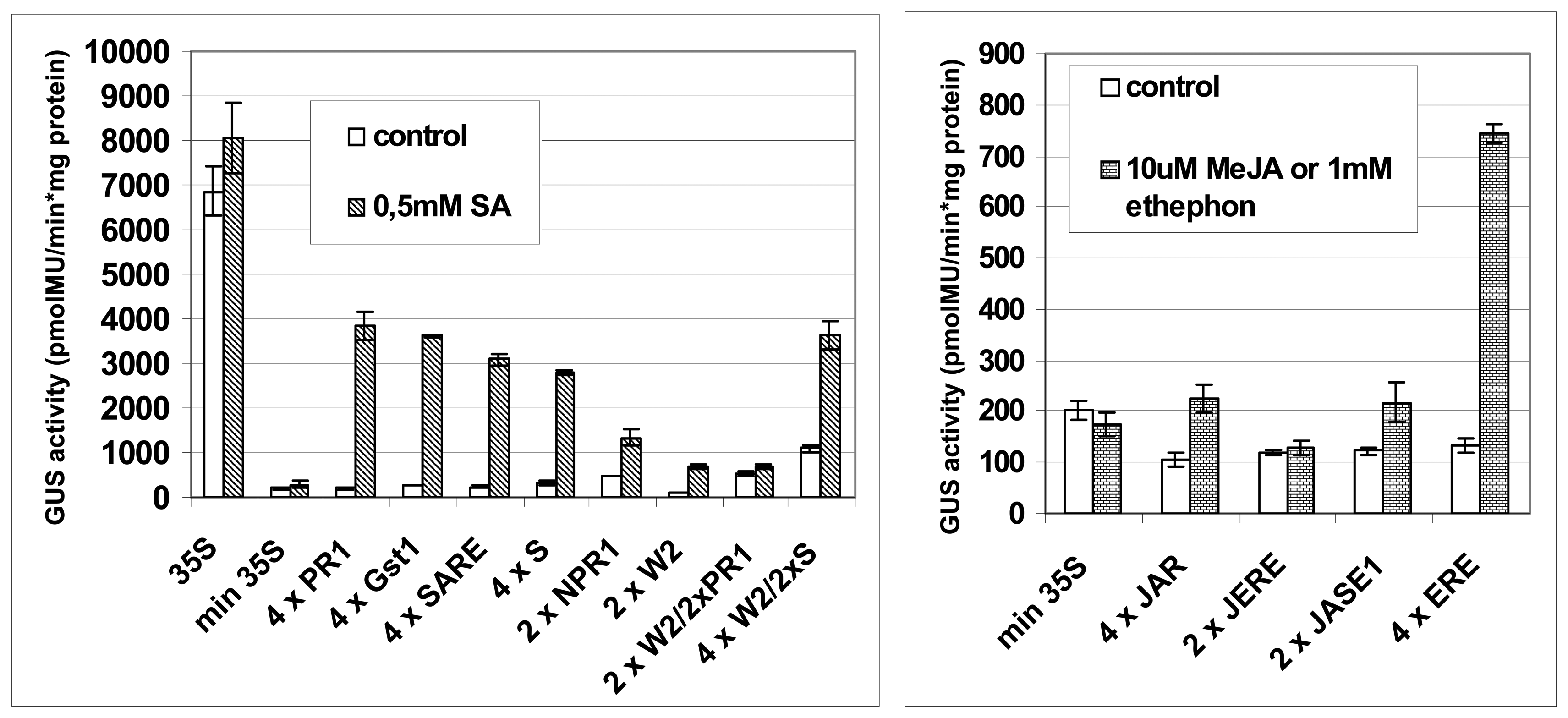
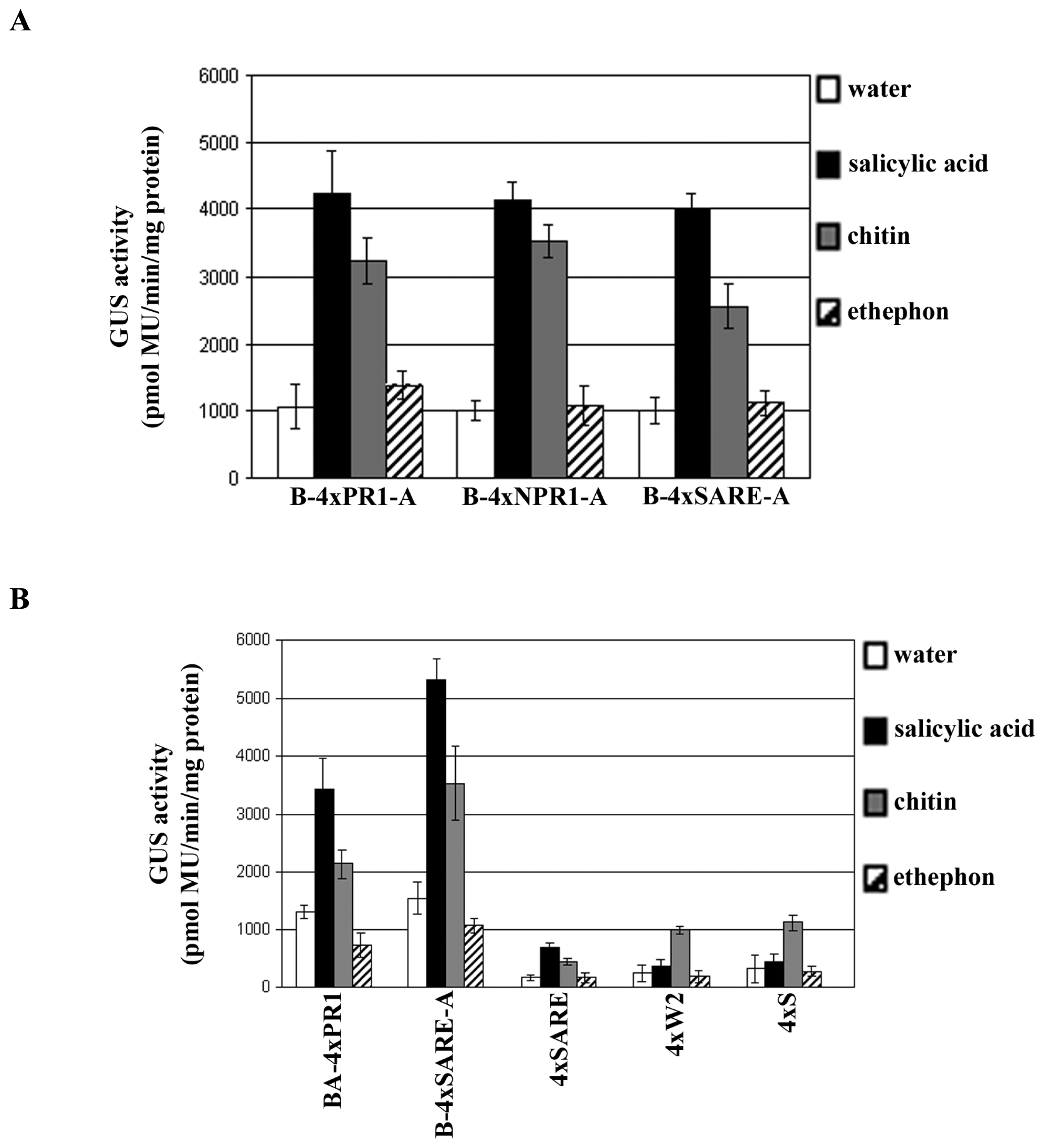
2.2 Assessment of synthetic promoters in transient expression assays
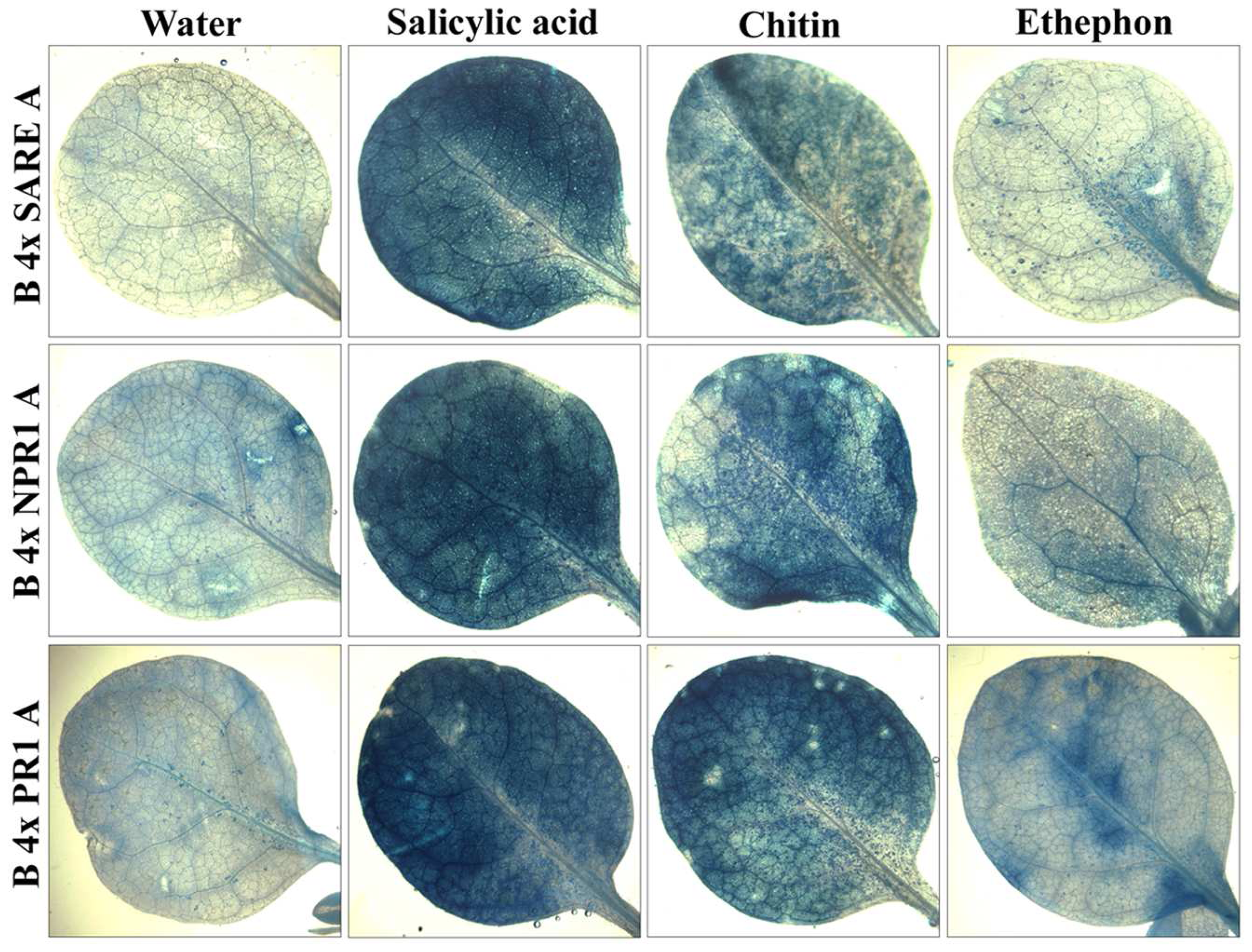
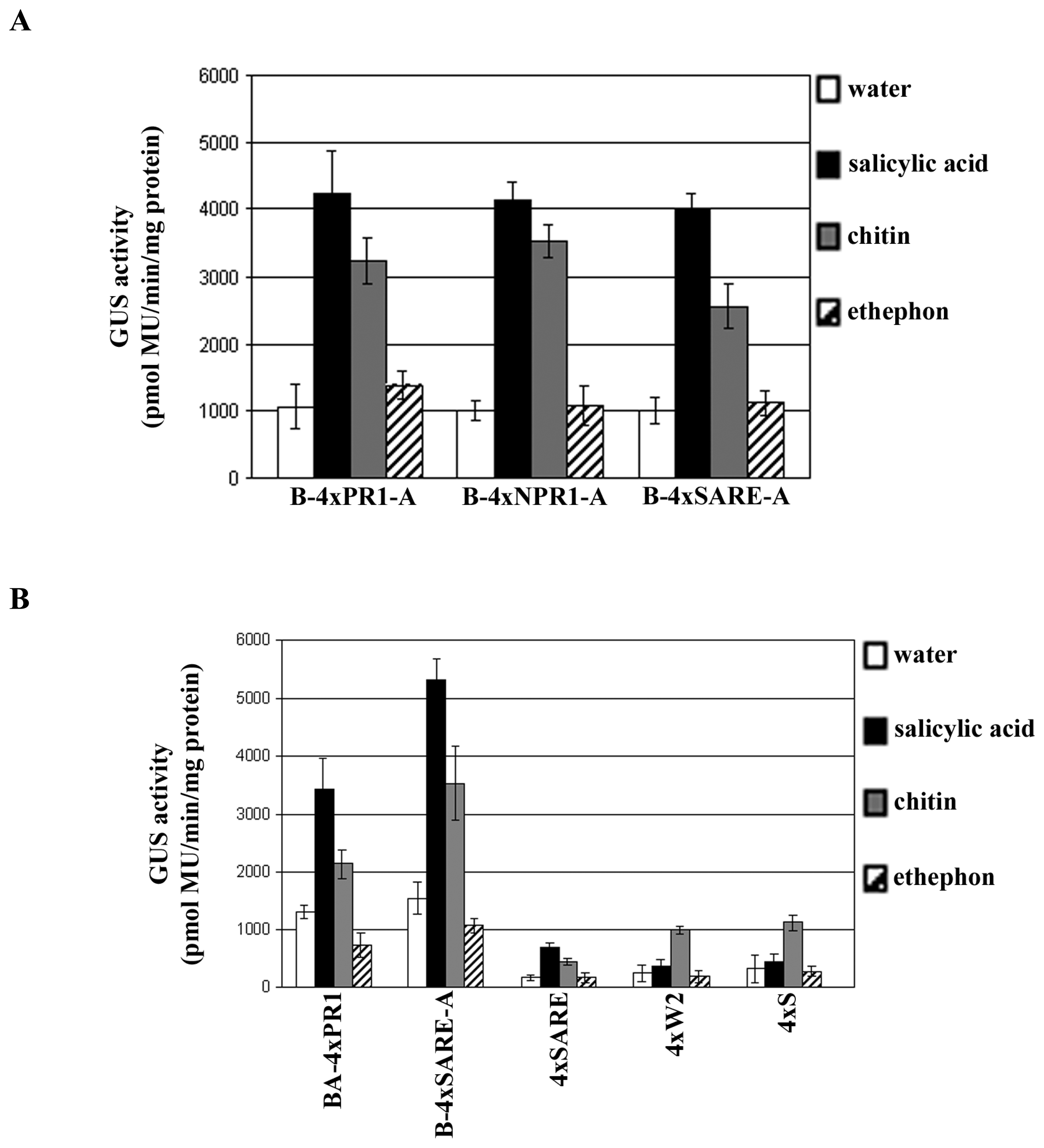
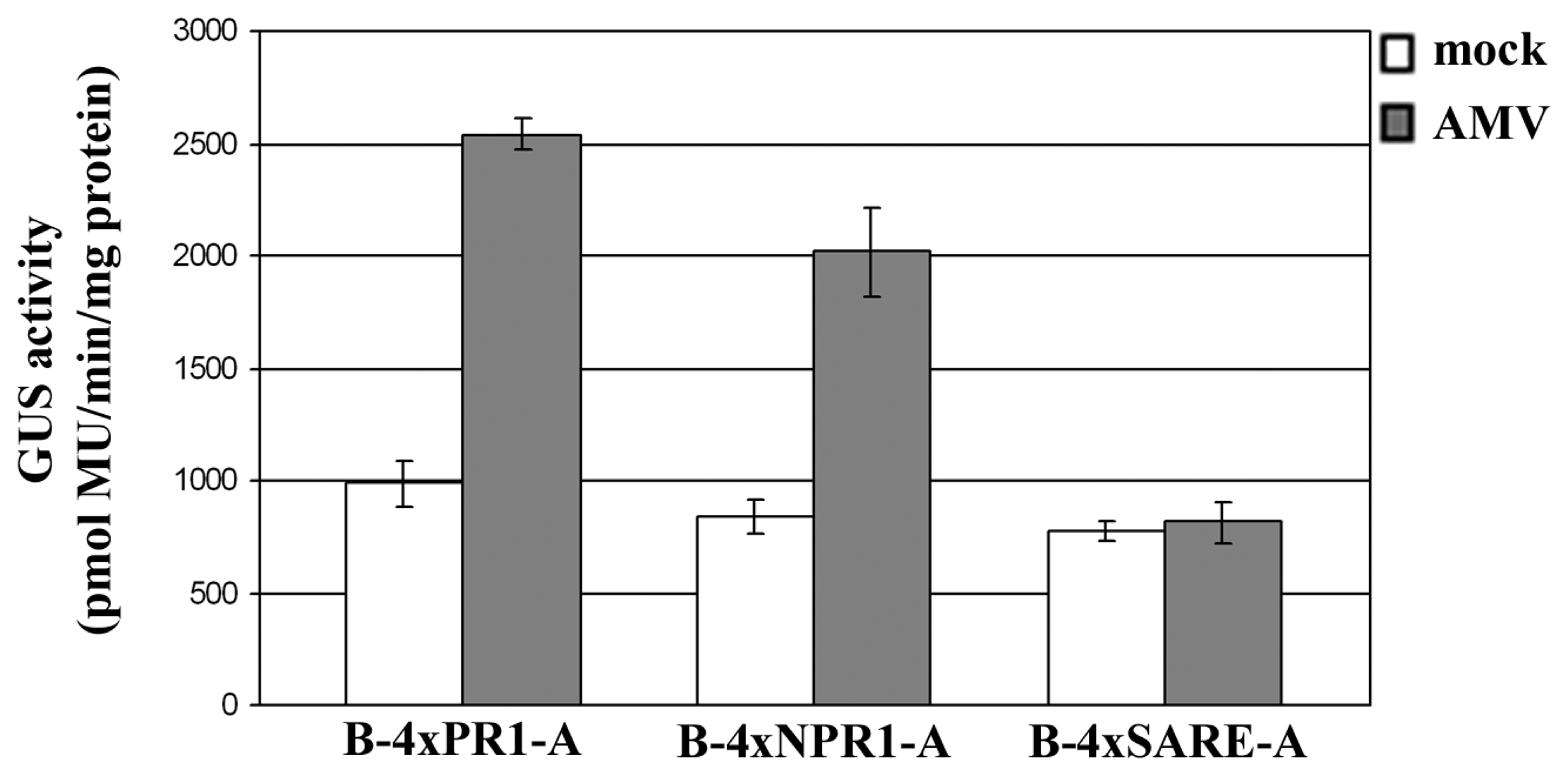
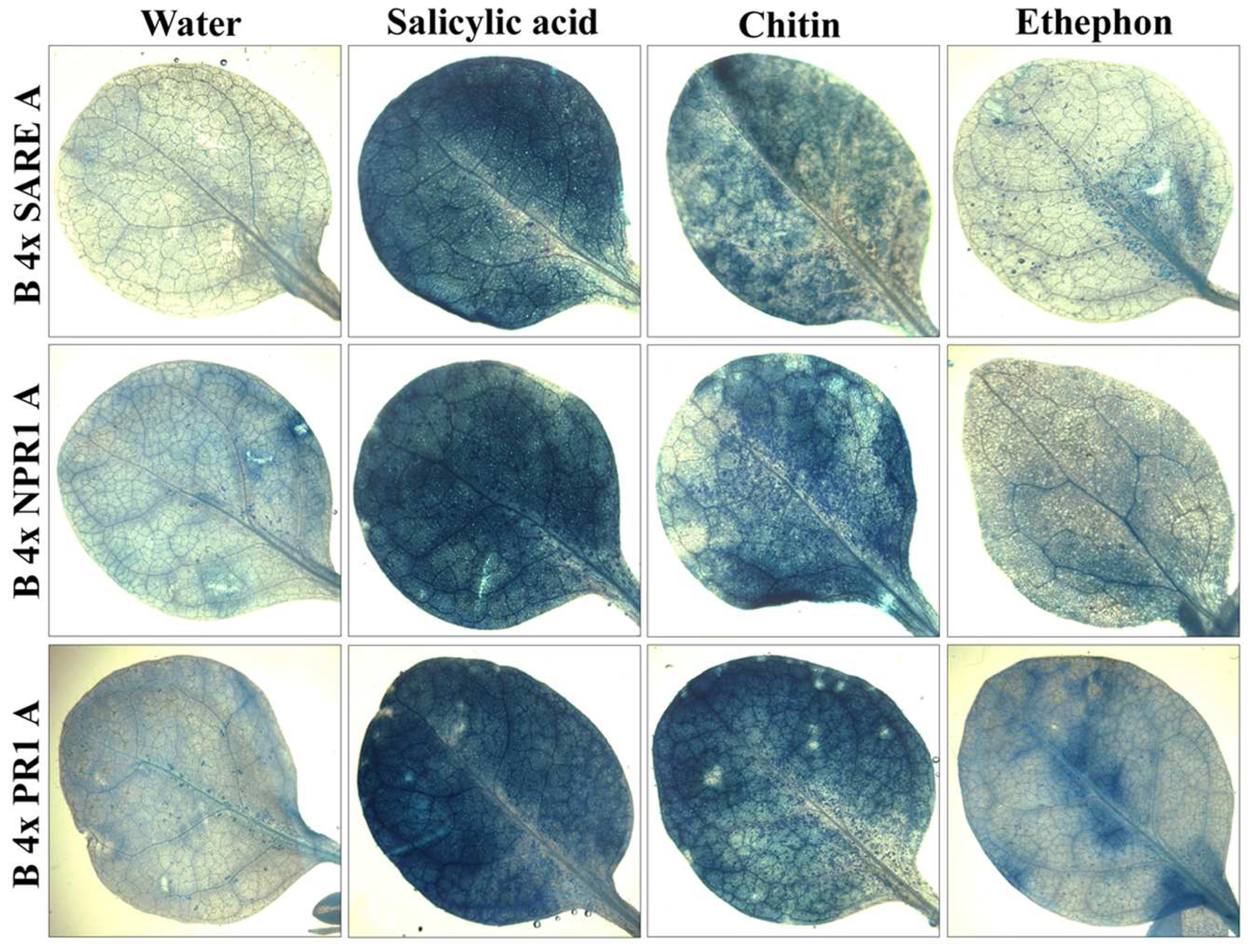
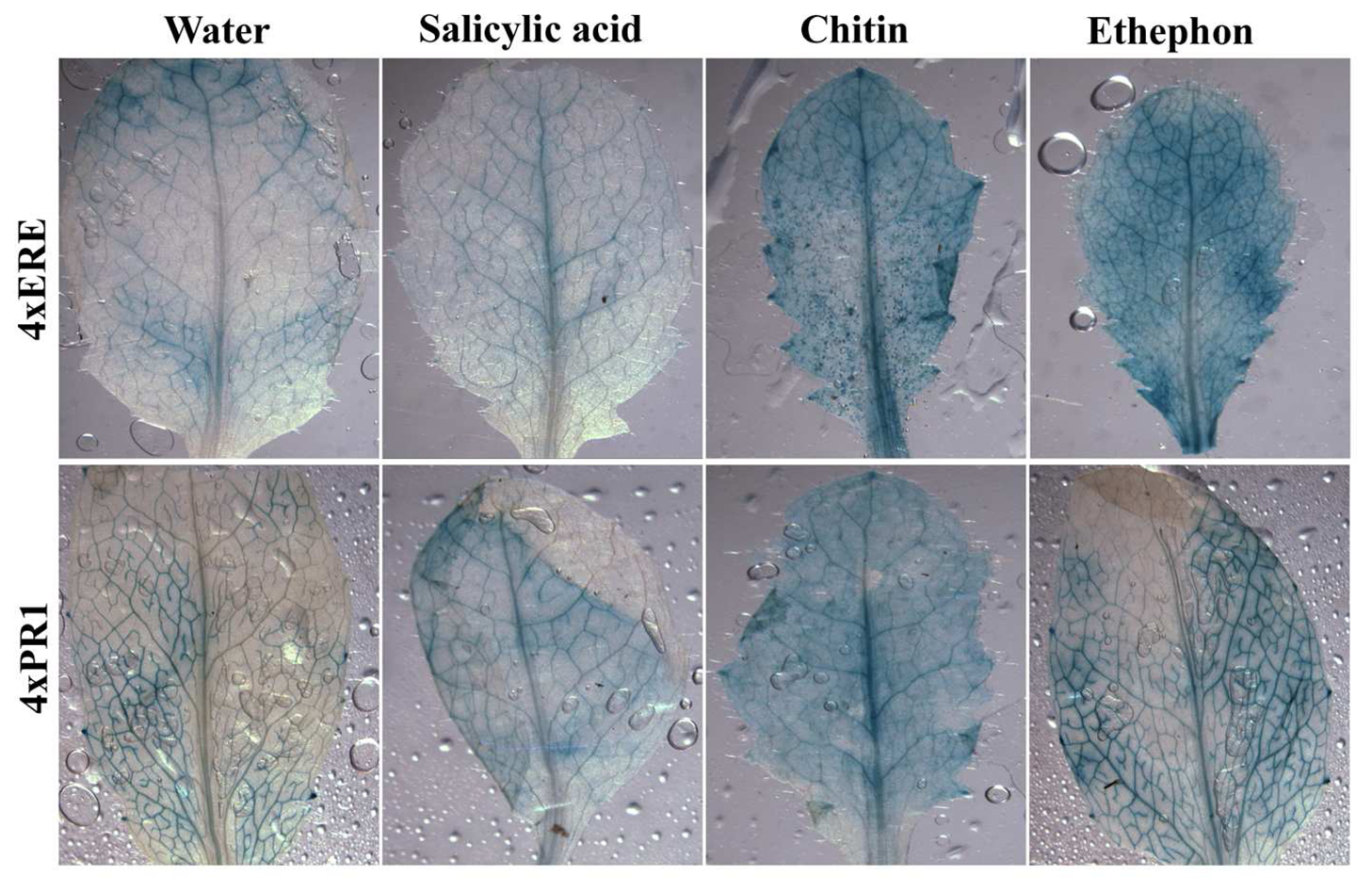
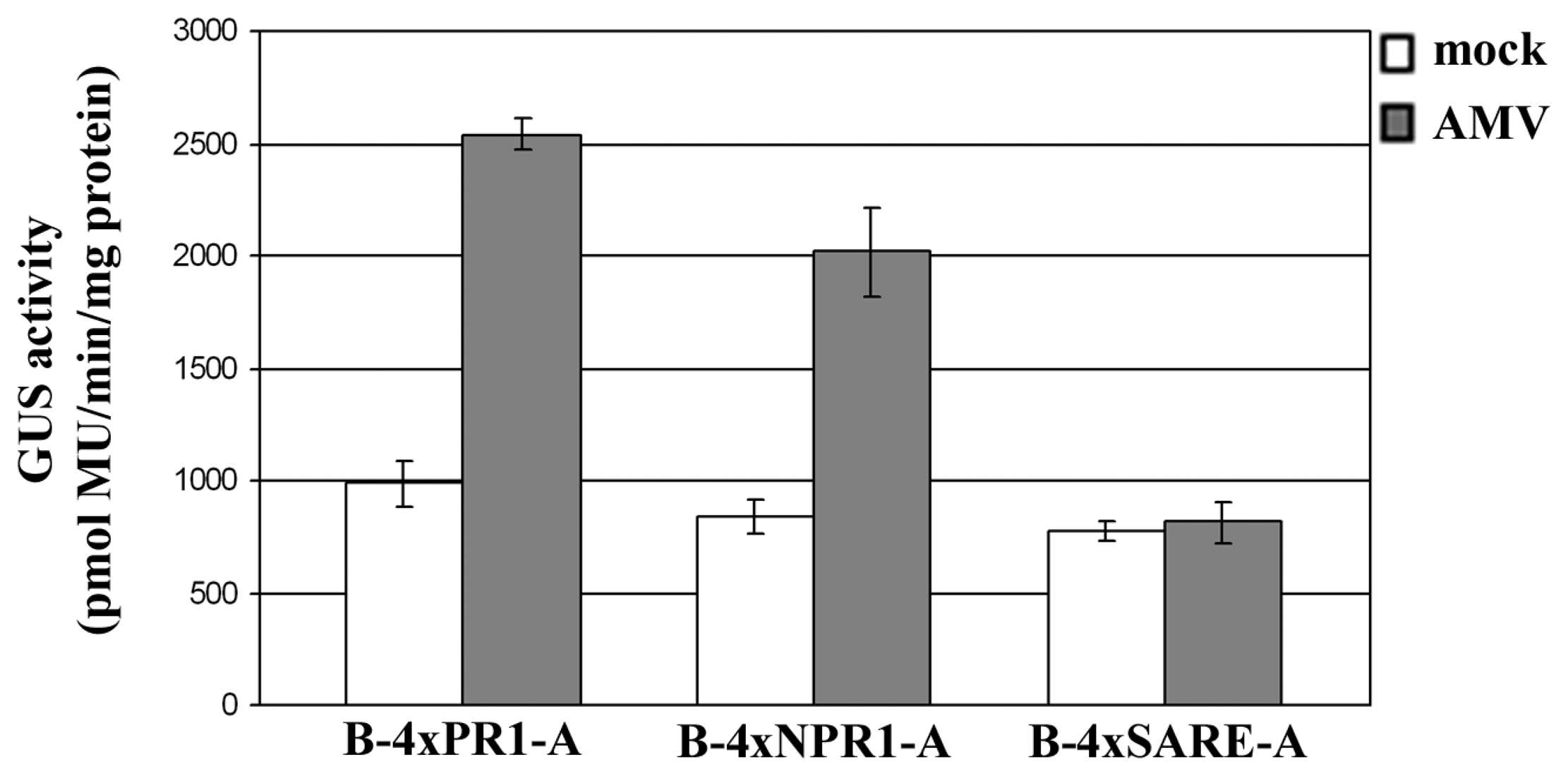
2.3 Examining the synthetic promoters in transgenic plants
3. Concluding remarks
4. Experimental section
4.1 Construction of synthetic promoters
4.2 Protoplasts transient expression assays
4.3 Transgenic plants expression assays
4.4 GUS analysis
4.4.1 Fluorometric assays
4.4.2 Histochemical assays
4.5 Virus inoculation
Acknowledgments
References
- Strange, R.N.; Scott, P.R. Plant disease: a threat to global food security. Annu. Rev. Phytopathol. 2005, 43, 85–117. [Google Scholar]
- Stewart, C.N., Jr. Monitoring the presence and expression of transgenes in living plants. Trends Plant Sci. 2005, 10, 390–396. [Google Scholar]
- Kodama, S.; Okada, K.; Inui, H.; Ohkawa, H. Aryl hydrocarbon receptor (AhR)-mediated reporter gene expression systems in transgenic tobacco plants. Planta 2007, 227, 37–45. [Google Scholar]
- Kovalchuk, I.; Kovalchuk, O. Transgenic plants as sensors of environmental pollution genotoxicity. Sensors 2008, 8, 1539–1558. [Google Scholar]
- Gurr, S.J.; Rushton, P.J. Engineering plants with increased disease resistance: how are we going to express it? Trends Biotechnol. 2005, 10, 390–396. [Google Scholar]
- Metraux, J.P.; Nawrath, C.; Genoud, T. Systemic acquired resistance. Euphytica 2002, 124, 237–243. [Google Scholar]
- Jones, J.D.G.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. [Google Scholar]
- Martin, G.B.; Bogdanove, A.J.; Sessa, G. Understanding the functions of plant disease resistance proteins. Annu. Rev. Plant Biol. 2003, 54, 23–61. [Google Scholar]
- Nürnberger, T.; Brunner, F.; Kemmerling, B.; Piater, L. Innate immunity in plants and animals: striking similarities and obvious differences. Immunol. Rev. 2004, 198, 249–266. [Google Scholar]
- Desikan, R.; Clarke, A.; Atherfold, P.; Hancock, J.T.; Neill, S.J. Harpin induces mitogen-activated protein kinase activity during defense responses in Arabidopsis thaliana suspension cultures. Planta 1999, 210, 97–103. [Google Scholar]
- Grant, M.; Lamb, C. Systemic immunity. Curr. Opin. Plant Biol. 2006, 9, 414–420. [Google Scholar]
- Pieterse, C.M.; van Wees, S.C.; van Pelt, J.A.; Knoester, M.; Laan, R.; Gerrits, H.; Weisbeek, P.J.; van Loon, L.C. A novel signaling pathway controlling induced systemic resistance in Arabidopsis. Plant Cell 1998, 10, 1571–1580. [Google Scholar]
- Ryals, J.A.; Neuenschwander, U.H.; Willits, M.G.; Molina, A.; Steiner, H.Y.; Hunt, M.D. Systemic acquired resistance. Plant Cell 1996, 8, 1809–1819. [Google Scholar]
- Thomma, B.P.H.J.; Eggermont, K.; Penninckx, I.A.M.A.; Mauch-Mani, B.; Vogelsang, R.; Cammue, B.P.A.; Broekaert, W.F. Separate jasmonate-dependent and salicylate-dependent defense-response pathways in Arabidopsis are essential for resistance to distinct microbial pathogens. Proc. Natl. Acad. Sci. USA 1998, 95, 15107–15011. [Google Scholar]
- Kessmann, H.; Staub, T.; Ligon, J.; Oostendorp, M.; Ryals, J. Activation of systemic acquired disease resistance in plants. Euro. J. plant Pathol. 1994, 100, 359–69. [Google Scholar]
- Rushton, P.J.; Reinstedler, A.; Lipka, V.; Lippok, B.; Somssich, I.E. Synthetic plant promoters containing defined regulatory elements provide novel insights into pathogen- and wound-induced signaling. Plant Cell 2002, 14, 749–762. [Google Scholar]
- Venter, M. Synthetic promoters: genetic control through cis engineering. Trends Plant Sci. 2007, 12, 118–124. [Google Scholar]
- Kooshki, M.; Mentewab, A.; Stewart, C.N., Jr. Pathogen inducible reporting in transgenic tobacco using a GFP construct. Plant Sci. 2003, 165, 213–219. [Google Scholar]
- Lebel, E.; Heifetz, P.; Thorne, L.; Uknes, S.; Ryals, J.; Ward, E. Functional analysis of regulatory sequences controlling PR-1 gene expression in Arabidopsis. Plant J. 1998, 16, 223–233. [Google Scholar]
- Guerineau, F.; Benjdia, M.; Zhou, D.X. A jasmonate-responsive element within the A. thaliana vsp1 promoter. J. Exp. Bot. 2003, 54, 1153–1162. [Google Scholar]
- Shah, J.; Klessig, D.F. Identification of a salicylic acid-responsive element in the promoter of the tobacco pathogenesis-related β-1,3-glucanase gene, PR-2d. Plant J. 1996, 10, 1089–1101. [Google Scholar]
- Ohme-Takagi, M.; Shinshi, H. Ethylene-inducible DNA binding proteins that interact with an ethylene-responsive element. Plant Cell 1995, 7, 173–182. [Google Scholar]
- Brown, R.L.; Kazan, K.; McGrath, K.C.; Maclean, D.J.; Manners, J.M. A role for the GCC-box in jasmonate-mediated activation of the PDF1.2 gene of Arabidopsis. Plant Physiol. 2003, 132, 1020–1032. [Google Scholar]
- Kirsch, C.; Takamiya-Wik, M.; Schmelzer, E.; Hahlbrock, K.; Somssich, I.E. A novel regulatory element involved in rapid activation of parsley ELI7 gene family members by fungal elicitor or pathogen infection. Mol. Plant Pathol. 2000, 1, 243–251. [Google Scholar]
- Yu, D.; Chen, C.; Chen, Z. Evidence for an important role of WRKY DNA binding proteins in the regulation of NPR-1 gene expression. Plant Cell 2001, 13, 1527–1539. [Google Scholar]
- Menke, F.L.H.; Champion, A.; Kijne, J.W.; Memelink, J. A novel jasmonate- and elicitor-responsive element in the periwinkle secondary metabolite biosynthetic gene Str interacts with a jasmonate- and elicitor-inducible AP2-domain transcription factor. ORCA2. EMBO J. 1999, 18, 4455–4463. [Google Scholar]
- He, Y.; Gan, S. Identical promoter elements are involved in regulation of the OPR1 gene by senescence and jasmonic acid in Arabidopsis. Plant Mol. Biol. 2001, 47, 495–505. [Google Scholar]
- Stewart, C.N., Jr. Go with the glow: fluorescent proteins to light transgenic organisms. Trends Biotechnol. 2006, 24, 155–162. [Google Scholar]
- Benfey, P.N.; Ren, L.; Chua, N.H. Tissue-specific expression from CaMV 35S enhancer subdomains in early stages of plant development. EMBO J. 1990, 9, 1677–1684. [Google Scholar]
- Fang, R.X.; Nagy, F.; Sivasubramaniam, S.; Chua, N.H. Multiple cis-regulatory elements for maximal expression of the cauliflower mosaic virus 35S promoter in transgenic plants. Plant Cell 1989, 1, 141–150. [Google Scholar]
- Raventos, D.; Jensen, A.B.; Rask, M.B.; Casacuberta, J.M.; Mundy, J.; San Segundo, B. A 20 bp cis-acting element is both necessary and sufficient to mediate elicitor response of a maize PRms gene. Plant J. 1995, 7, 147–155. [Google Scholar]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar]
- Luo, K.; Duan, H.; Zhao, D.; Zheng, X.; Deng, W.; Chen, Y.; Stewart, C.N., Jr.; Wu, Y.; Jiang, X.; He, A.; McAvoy, R.; Pei, Y.; Li, Y. GM-gene-deletor: fused loxP-FRT recognition sequences dramatically improves efficiency of FLP or Cre recombinase on transgene excision from pollen and seed of tobacco plants. Plant Biotechnol. J. 2007, 5, 263–274. [Google Scholar]
- Curtis, M.D.; Grossniklaus, U. A Gateway cloning vector set for high-throughout functional analysis of genes in planta. Plant Physiol. 2003, 133, 462–469. [Google Scholar]
- Abel, S.; Theologis, A. Transient transformation of Arabidopsis leaf protoplasts: a versatile experimental system to study gene expression. Plant J. 1994, 5, 421–427. [Google Scholar]
- Horsch, R.B.; Fry, J.E.; Hoffman, N.L.; Eichholtz, D.; Rogers, S.G.; Fraley, R.T. A simple and general method for transferring genes into plants. Science 1985, 227, 1229–1231. [Google Scholar]
- Clough, S.J.; Bent, A.F. Floral dip: a simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar]
- Jefferson, R.A.; Kavanagh, T.A.; Bevan, M.W. GUS fusions: β-glucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J. 1987, 99, 3901–3907. [Google Scholar]
- Hajimorad, M.R.; Francki, R.I.B. Alfalfa mosaic virus isolates from luceme in South Australia: biological variability and antigenic similarity. Ann. Appl. Biol. 1998, 113, 45–54. [Google Scholar]
- Hajimorad, M.R.; Hill, J.H. Rsv1-mediated resistance against Soybean mosaic virus-N is hypersensitive response-independent at inoculation site, but has the potential to initiate a hypersensitive response-like mechanism. Mol. Plant-Microbe Interact. 2001, 14, 587–598. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cis-acting regulatory element (RE) | Source and gene Promoter | Stimuli reported to cause induction | Reference |
|---|---|---|---|
| PR1-motif ACGTCATAGATGTGGCGGCATATATTCTTCAGGACTTTTC | Arabidopsis PR1 | Salicylic acid | [19] |
| JAR (jasmonic acid responsive element) CAACGACACGCCAAATTCTAATTTAGCACAGTCTCACGTG | Arabidopsis VSP1 | Jasmonic acid | [20] |
| GST1-box TTCTAGCCACCAGATTTGACCAAAC | Potato GST1 | Phytophthora elicitor, oomycetes, fungi, bacteria | [16] |
| SARE (salicylic acid responsive element) TTCGACCTCCAAAGAGGACCCAGAAT | Tobacco PR2-d | Salicylic acid | [21] |
| ERE (ethylene responsive element) CAGCCGCCAAAGAGGACCCAGAAT | Tobacco chitinase | Ethylene, Phytophthora elicitor, oomycetes, fungi, bacteria | [16,22,23] |
| S-box CAGCCACCAAAGAGGACCCAGAAT | Parsley ELI7 | Phytophthora sojae elicitor, fungal elicitor, oomycetes, fungi, bacteria | [16,24] |
| NPR1-motif TTGACTTGACTTGGCTCTGCTCGTCAA | Arabidopsis NPR1 | Salicylic acid, Pseudomonas syringae pv. tomato | [25] |
| JERE (jasmonic acid responsive element) AGACCGCCAAAGAGGACCCAGAAT | Periwinkle Str | Jasmonic acid, yeast-derived elicitors, Phytophthora elicitor, oomycetes, fungi, bacteria | [16,26] |
| JASE1 (jasmonic acid responsive element) CGTCAATGAATACGTCATC | Arabidopsis OPR1 | Jasmonic acid | [27] |
| W-box TTATTCAGCCATCAAAAGTTGACCAA-TAAT | Parsley PR1 | Fungal elicitor, oomycetes, fungi, bacteria | [16] |
© 2008 by MDPI (http://www.mdpi.org). Reproduction is permitted for noncommercial purposes.
Share and Cite
Mazarei, M.; Teplova, I.; Hajimorad, M.R.; Stewart, C.N., Jr. Pathogen Phytosensing: Plants to Report Plant Pathogens. Sensors 2008, 8, 2628-2641. https://doi.org/10.3390/s8042628
Mazarei M, Teplova I, Hajimorad MR, Stewart CN Jr. Pathogen Phytosensing: Plants to Report Plant Pathogens. Sensors. 2008; 8(4):2628-2641. https://doi.org/10.3390/s8042628
Chicago/Turabian StyleMazarei, Mitra, Irina Teplova, M. Reza Hajimorad, and C. Neal Stewart, Jr. 2008. "Pathogen Phytosensing: Plants to Report Plant Pathogens" Sensors 8, no. 4: 2628-2641. https://doi.org/10.3390/s8042628
APA StyleMazarei, M., Teplova, I., Hajimorad, M. R., & Stewart, C. N., Jr. (2008). Pathogen Phytosensing: Plants to Report Plant Pathogens. Sensors, 8(4), 2628-2641. https://doi.org/10.3390/s8042628