
Estimating Invasion Success by Non-Native Trees in a National Park Combining WorldView-2 Very High Resolution Satellite Data and Species Distribution Models

 ,
,  ,
,  and
and
Abstract
:1. Introduction
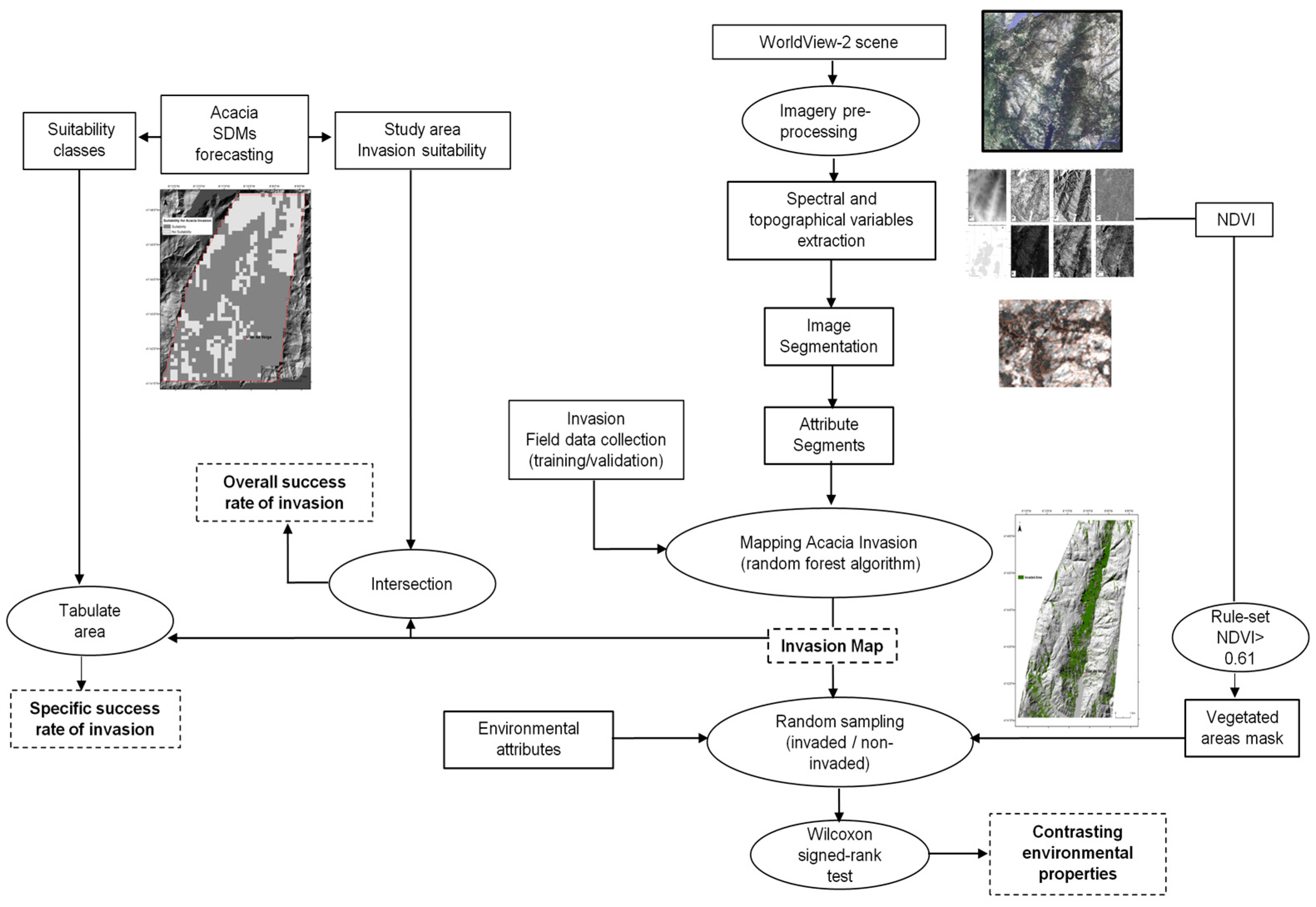
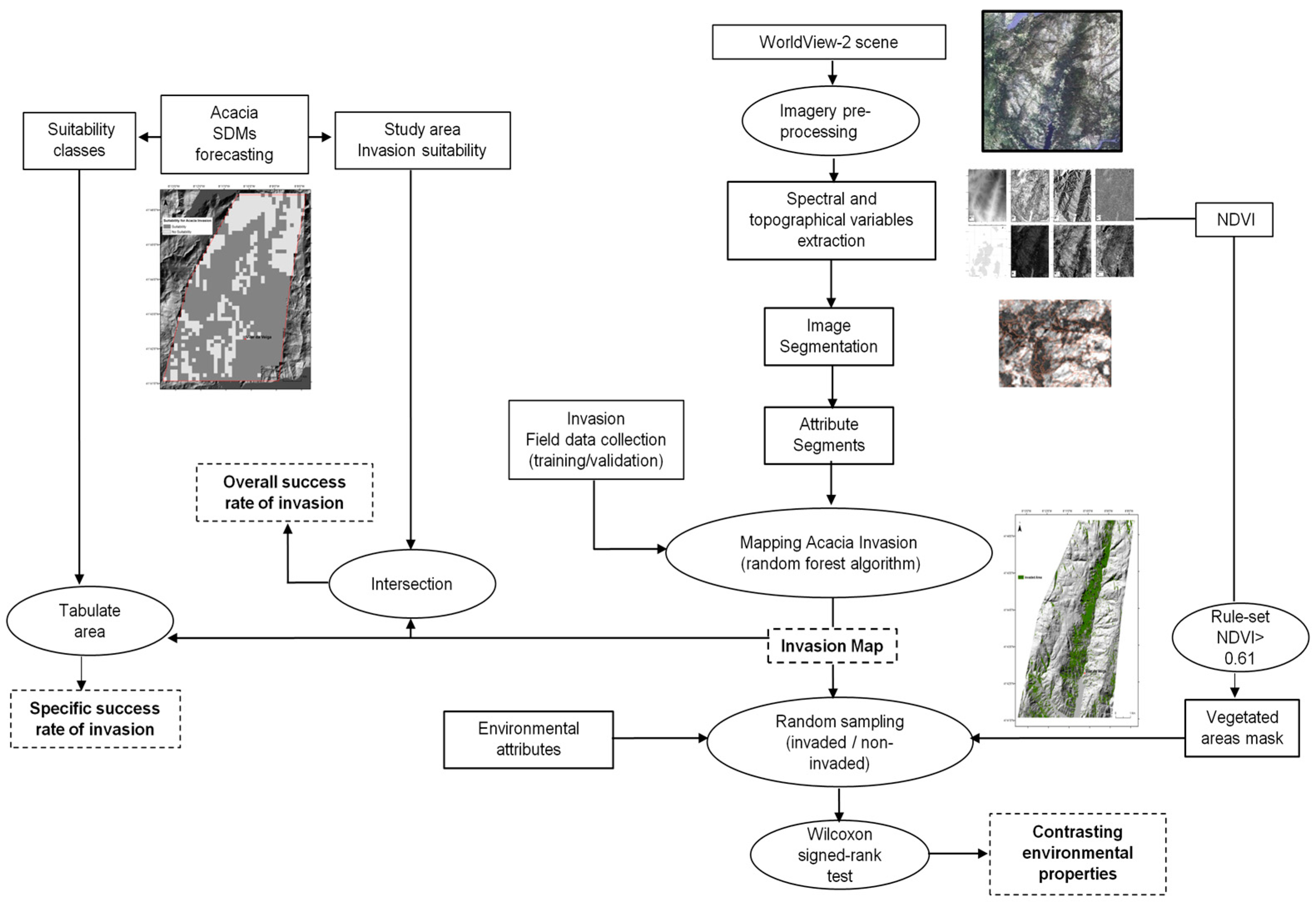
2. Materials and Methods
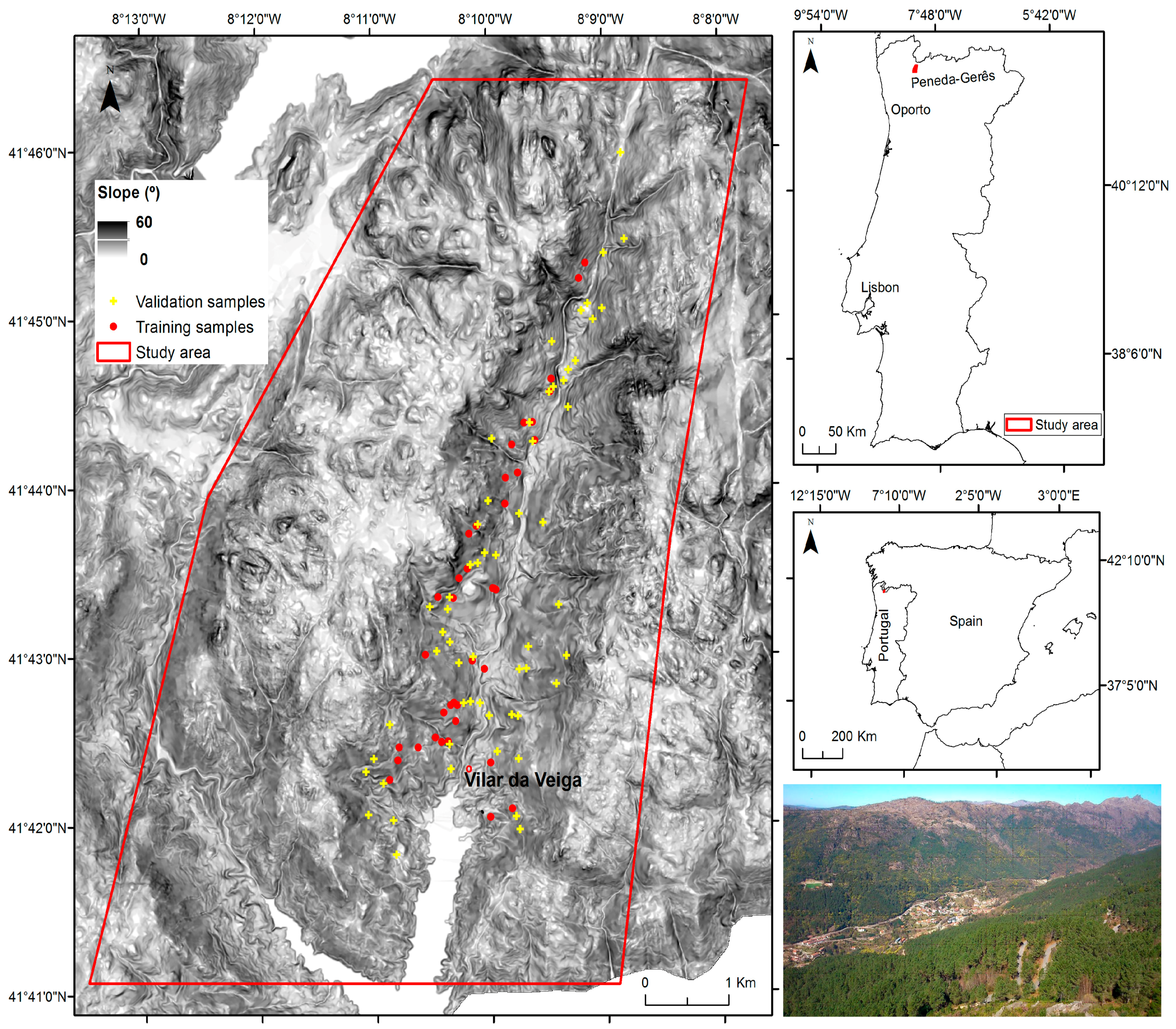
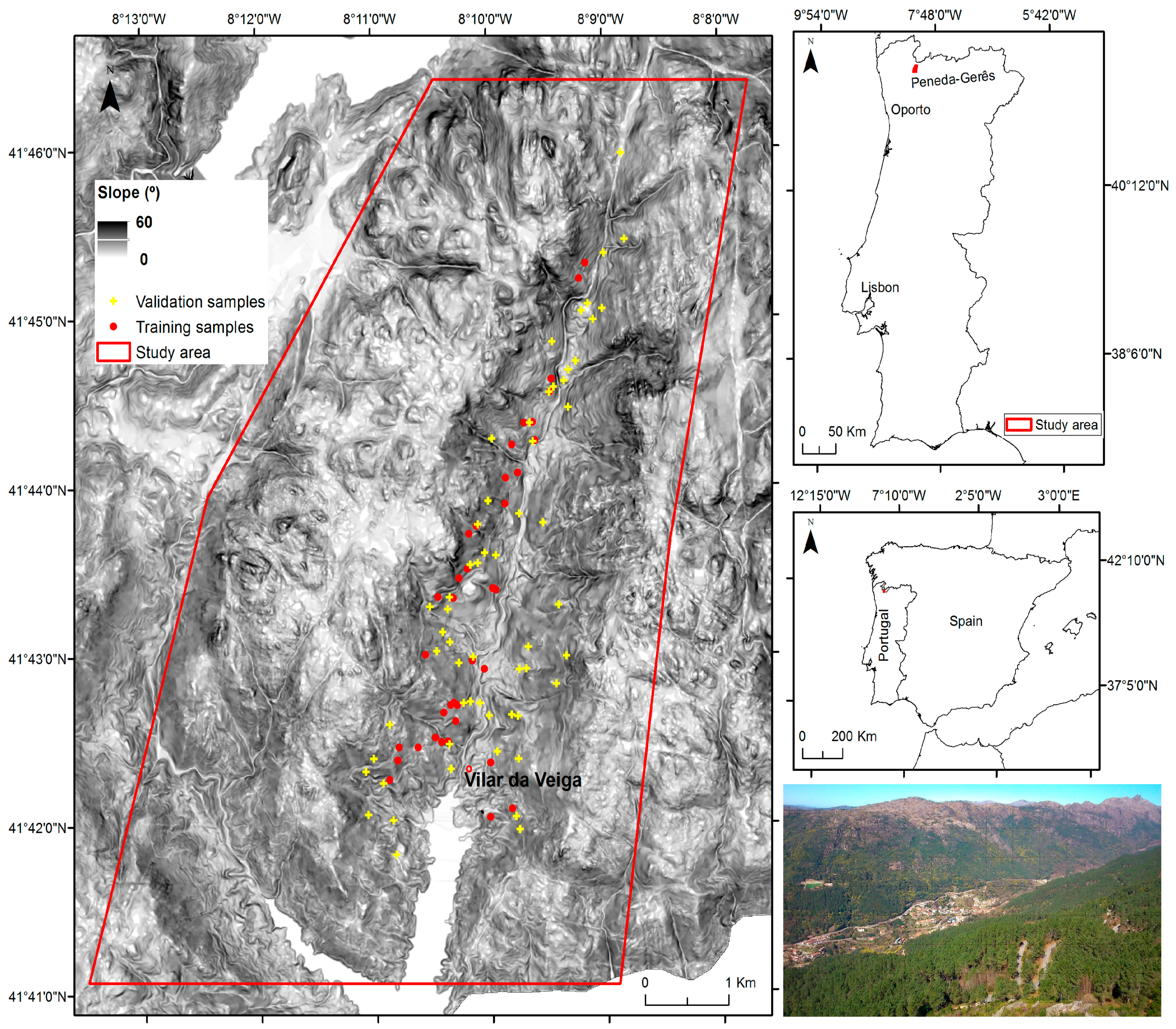
2.1. Study Area
2.2. Input Data Collection
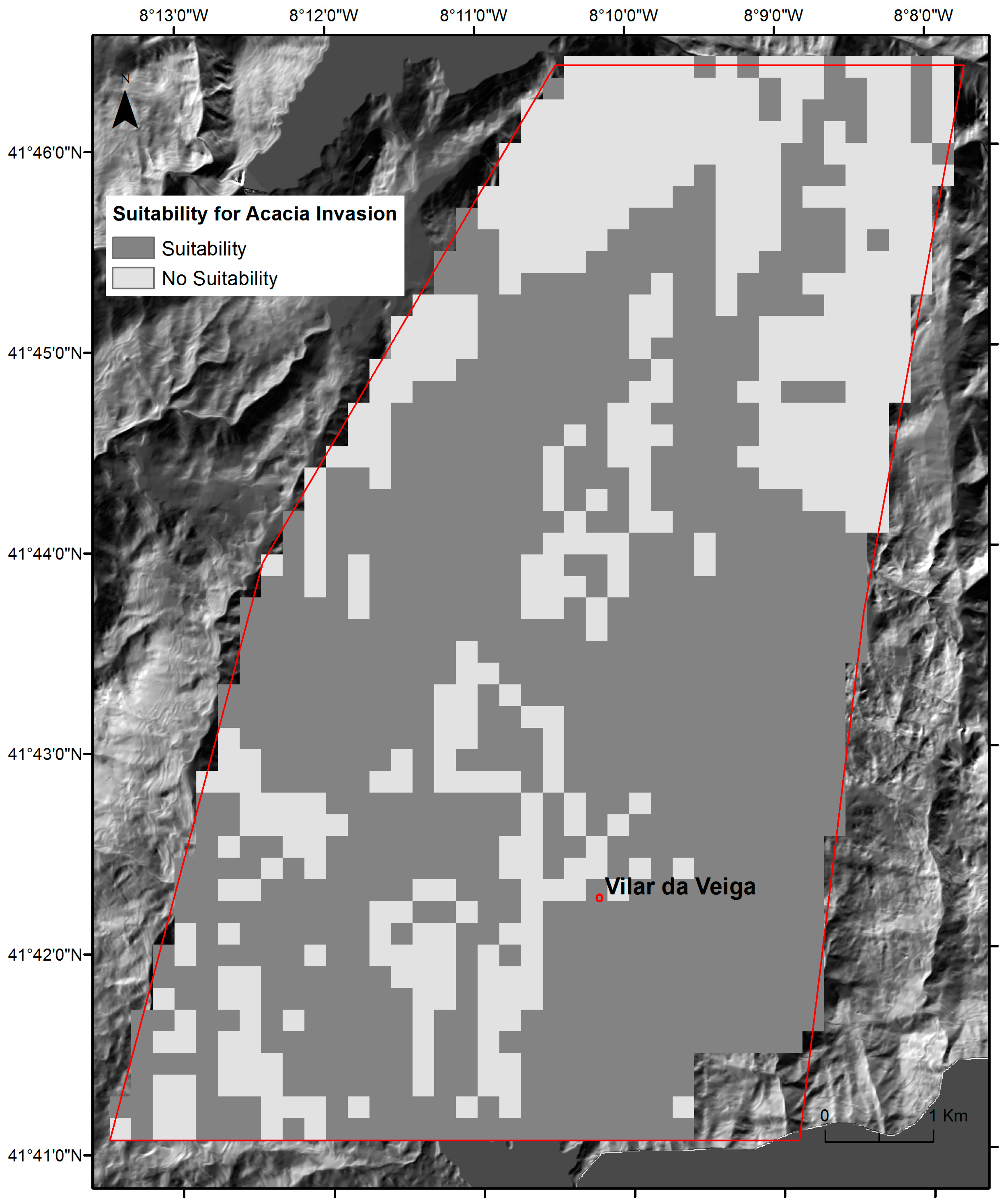
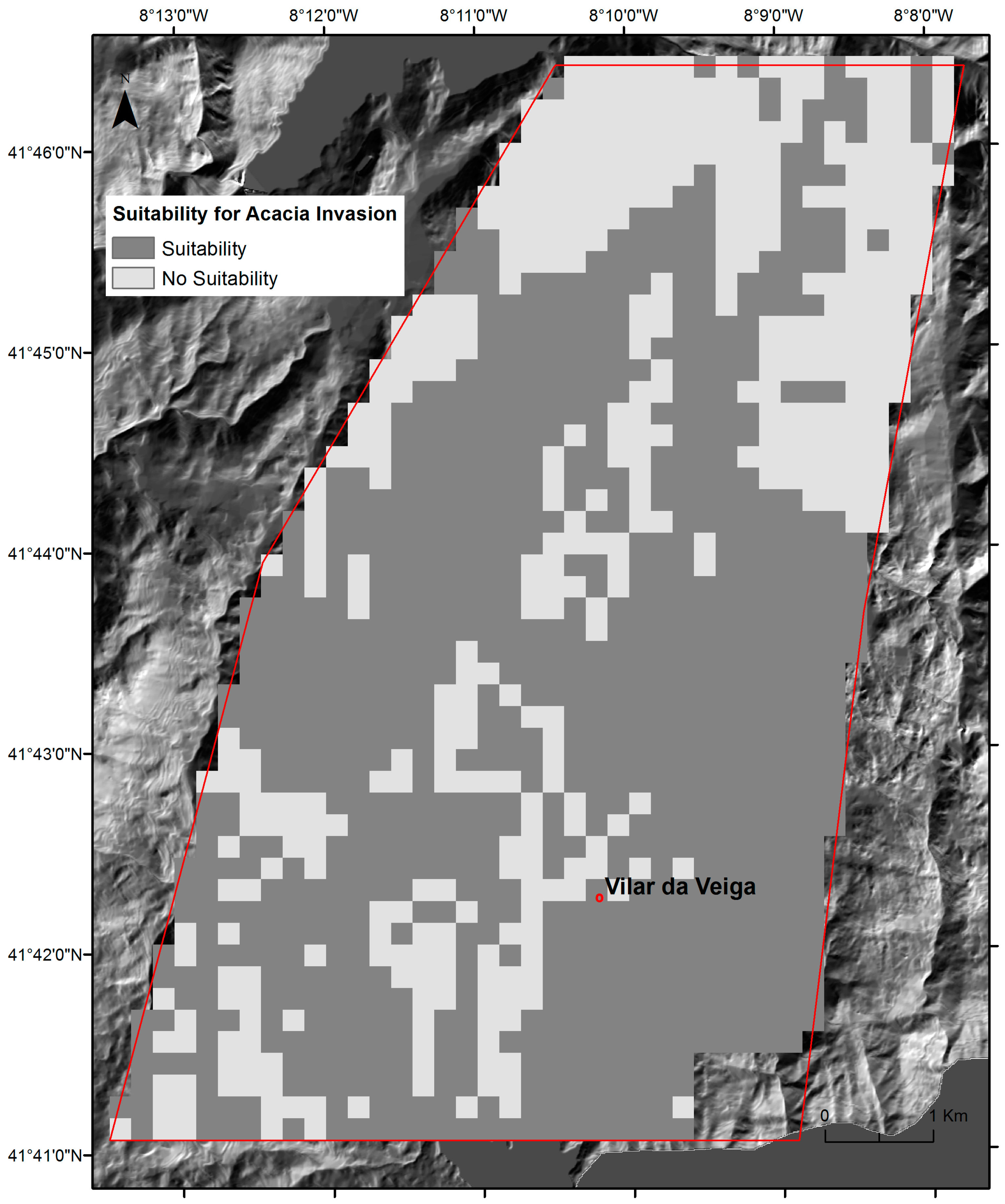
2.3. Regional Spatial-Explicit Suitability for A. dealbata Invasion
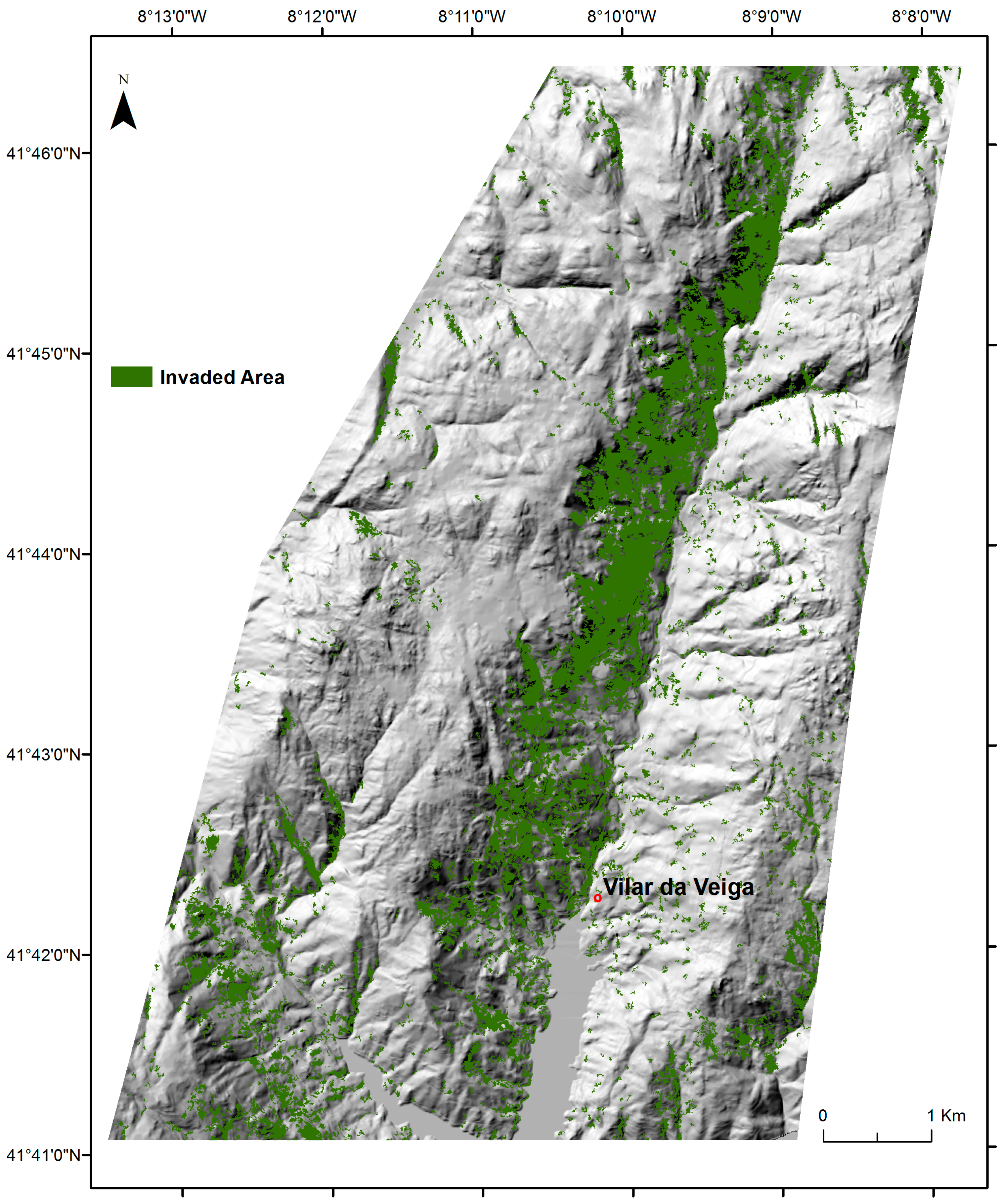
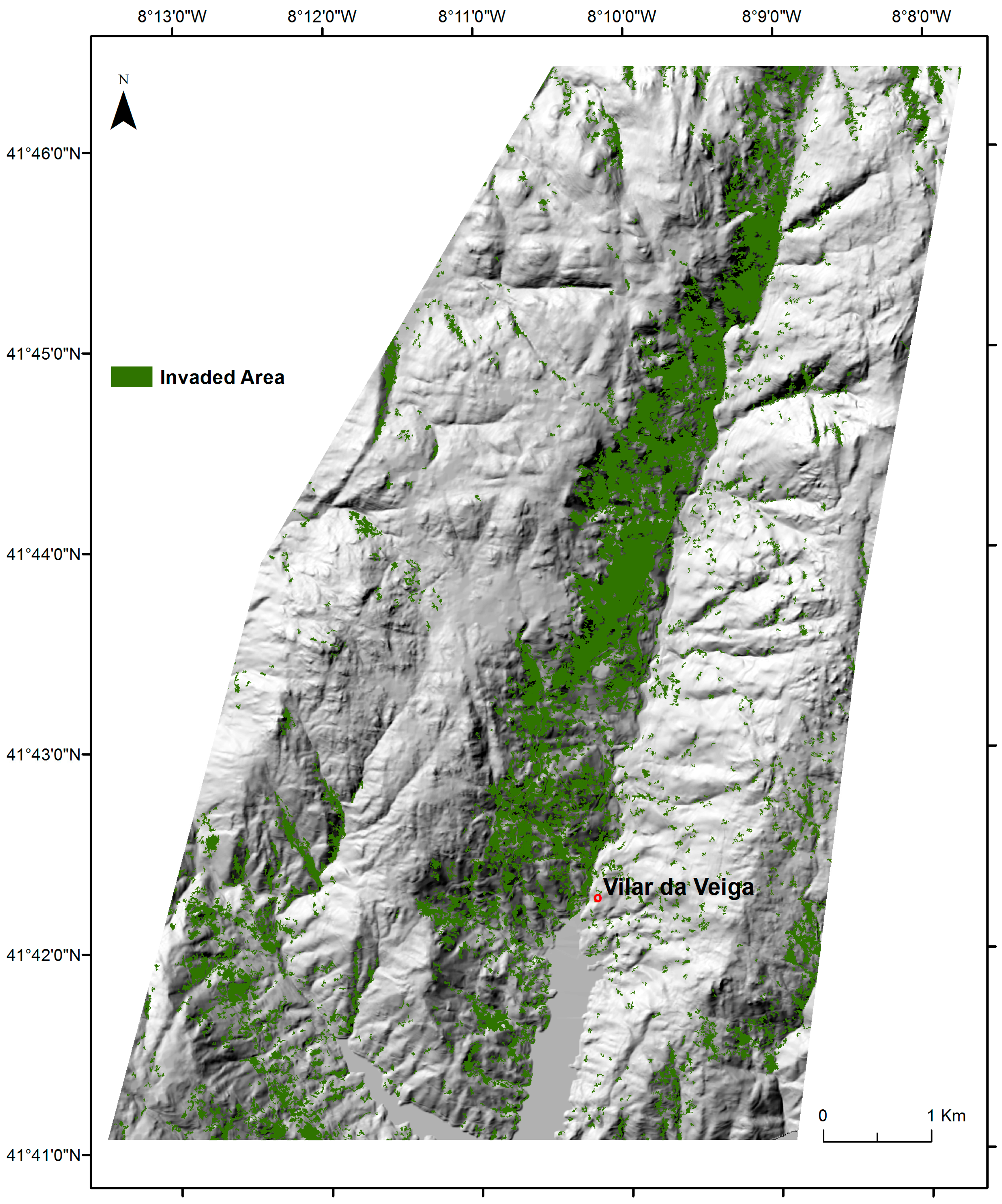
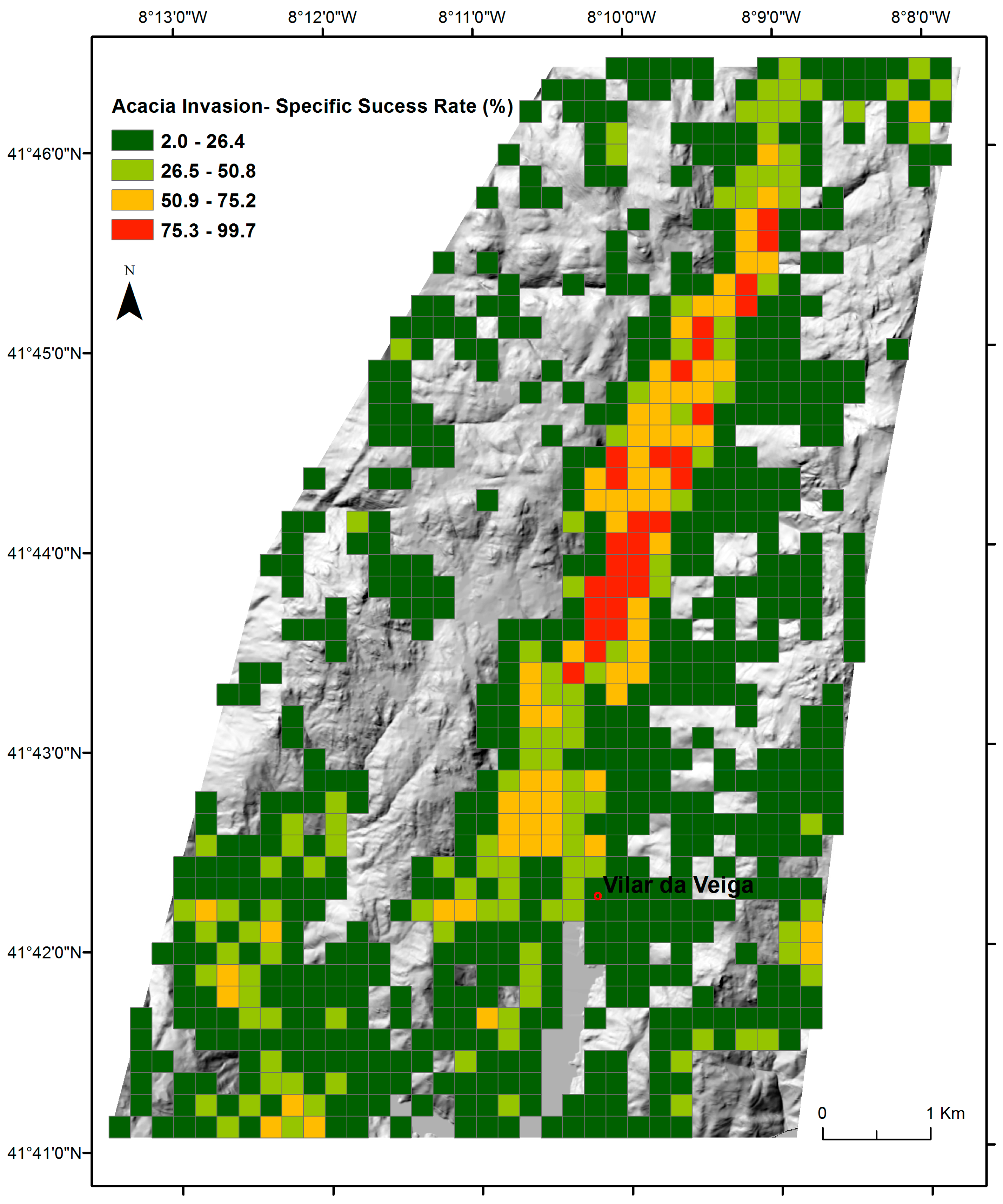
2.4. Mapping of A. dealbata Invasion and Estimation of Success Rate Of Invasion
2.5. Comparison of Environmental Attributes between Invaded and Non-Invaded Vegetated Areas
3. Results
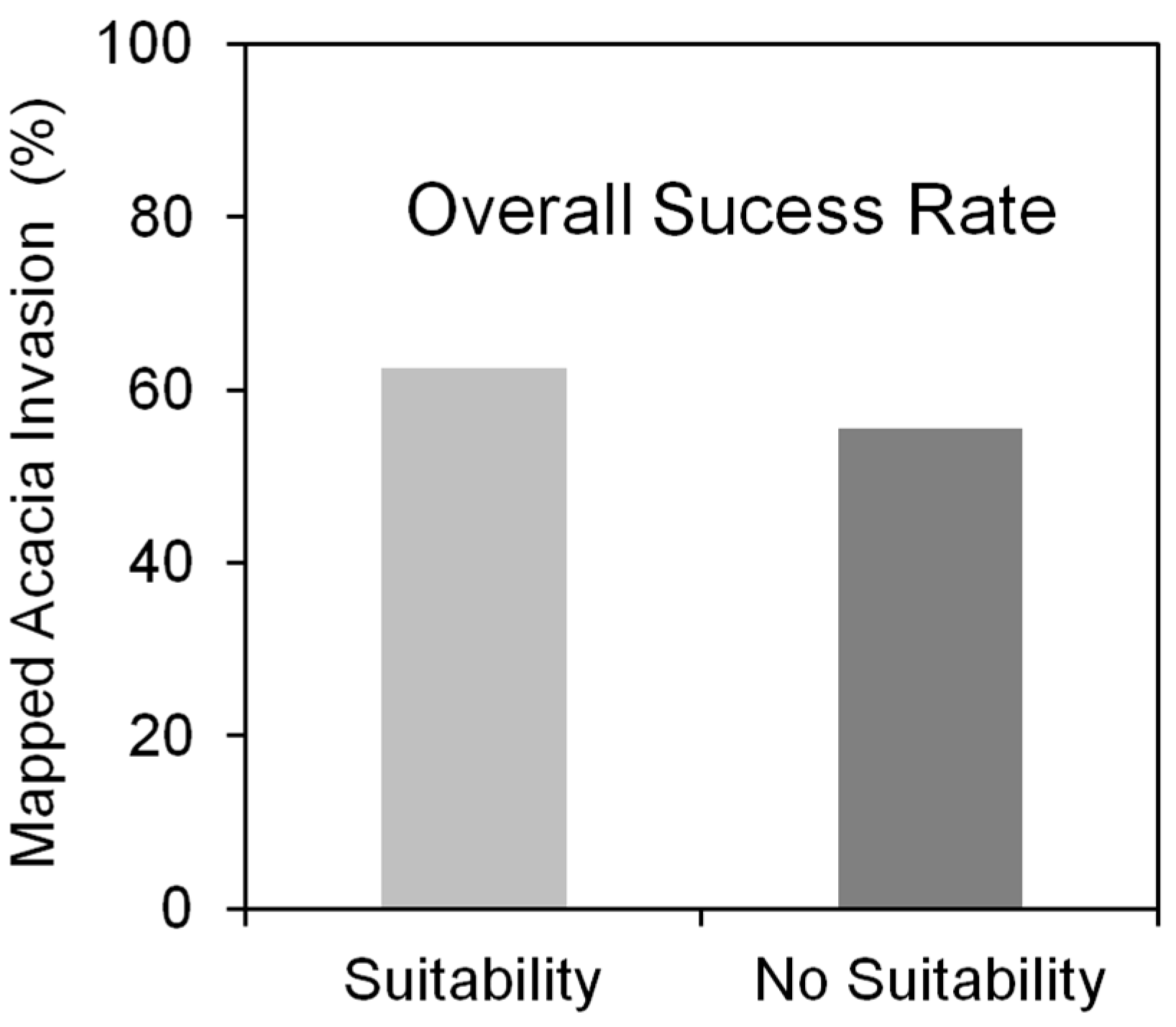
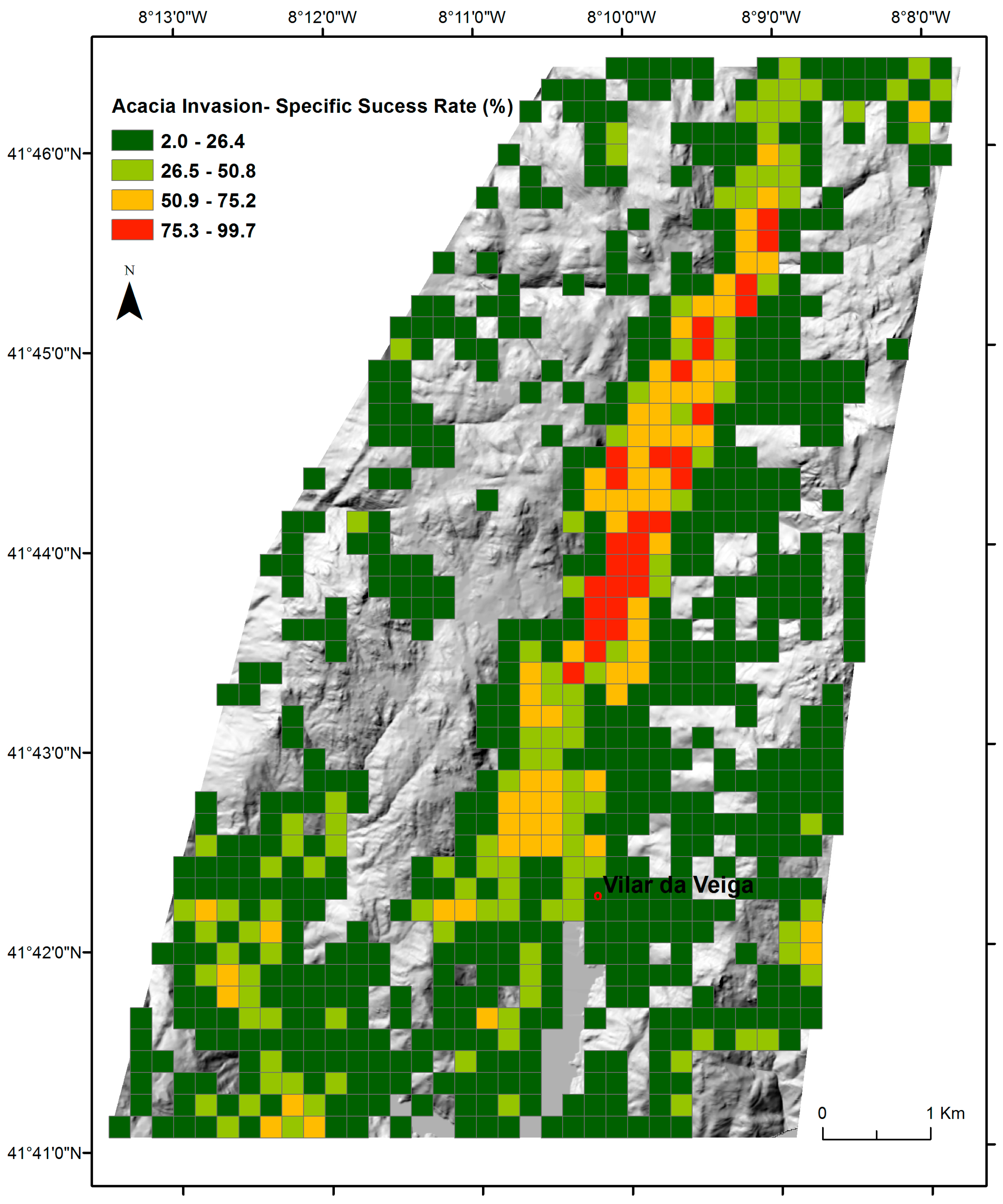
3.1. The Success Rate of A. dealbata Invasion in the Study Area: Mapped and SDMs Forecasted Spatial Patterns
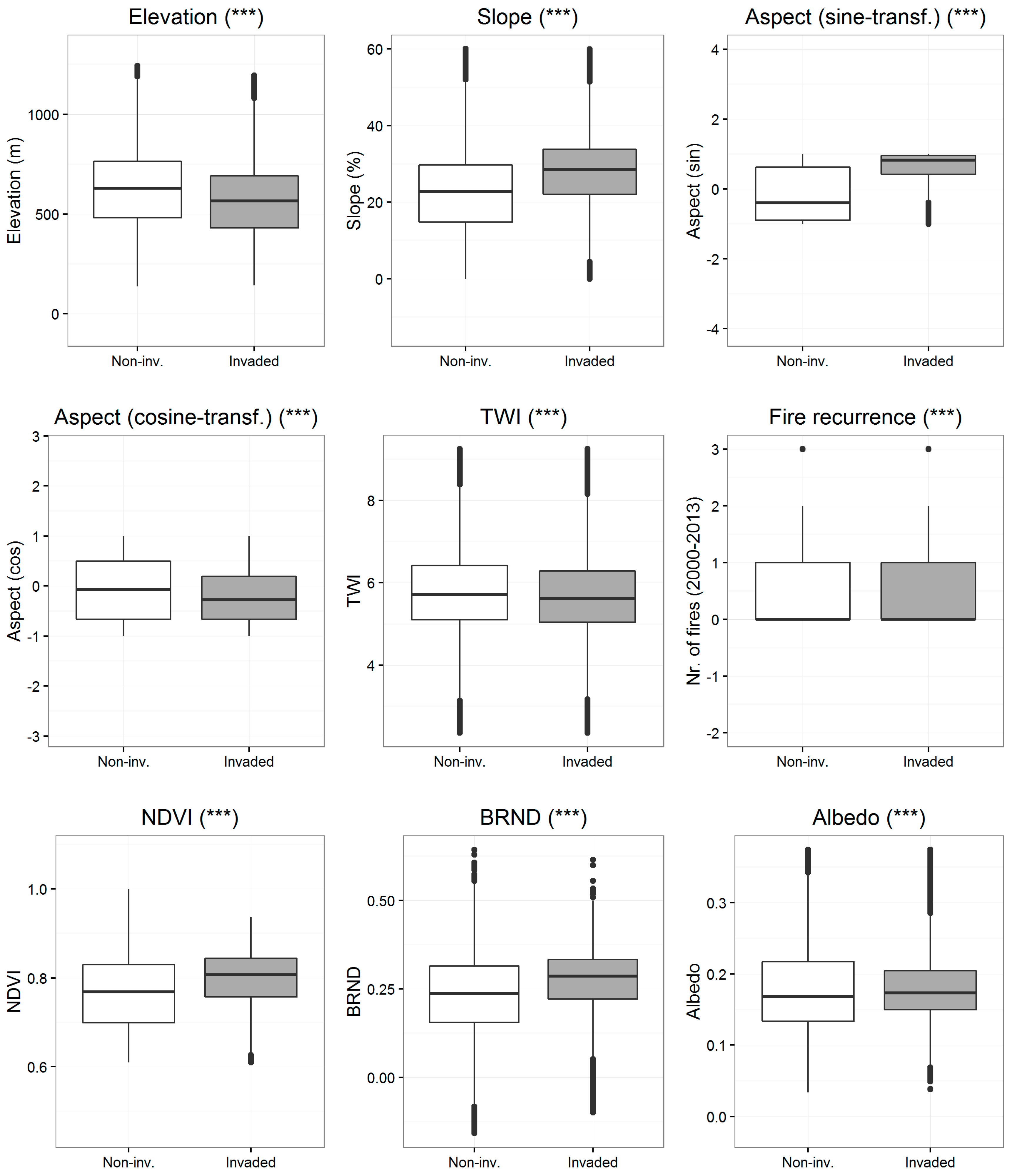
3.2. Variation of Environmental Attributes between Invaded and Non-Invaded Vegetated Areas
4. Discussion
4.1. The Combining of VHR Remote Sensing and SDMs for Invasion Success Assessment
4.2. The A. dealbata Invasion in the Study Area and the Variation of Environmental Attributes between Invaded and Non-Invaded Vegetated Areas: Implications for Management and Landscape Attributes
Acknowledgments
Author Contributions
Conflicts of Interest
References
- United Nations Environment Programme. Report of the Seventh Meeting of the Conference of the Parties to the Convention on Biological Diversity (unep/cbd/cop/35/4); UNEP: Kuala Lumpur, Malasia, 2010. [Google Scholar]
- European Commission. Communication from the Commission to the European Parliament, the Council, the Economic and Social Committee and the Committee of the Regions—Our Life Insurance, our Natural Capital: An eu Biodiversity Strategy to 2020; EC: Brussels, Belgium, 2011. [Google Scholar]
- Kueffer, C.; McDougall, K.; Alexander, J.; Daehler, C.; Edwards, P.; Haider, S.; Milbau, A.; Parks, C.; Pauchard, A.; Reshi, Z.A.; et al. Plant invasions into mountain protected areas: Assessment, prevention and control at multiple spatial scales. In Plant Invasions in Protected Areas: Patterns, Problems and Challenges; Foxcroft, C.L., Pyšek, P., Richardson, M.D., Genovesi, P., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 89–113. [Google Scholar]
- Fernandes, R.F.; Vicente, J.R.; Georges, D.; Alves, P.; Thuiller, W.; Honrado, J.P. A novel downscaling approach to predict plant invasions and improve local conservation actions. Biol. Invasions 2014, 16, 2577–2590. [Google Scholar] [CrossRef]
- Guisan, A.; Tingley, R.; Baumgartner, J.B.; Naujokaitis-Lewis, I.; Sutcliffe, P.R.; Tulloch, A.I.T.; Regan, T.J.; Brotons, L.; McDonald-Madden, E.; Mantyka-Pringle, C.; et al. Predicting species distributions for conservation decisions. Ecol. Lett. 2013, 16, 1424–1435. [Google Scholar] [CrossRef] [PubMed]
- He, K.S.; Bradley, B.A.; Cord, A.F.; Rocchini, D.; Tuanmu, M.-N.; Schmidtlein, S.; Turner, W.; Wegmann, M.; Pettorelli, N. Will remote sensing shape the next generation of species distribution models? Remote Sens. Ecol. Conserv. 2015, 1, 4–18. [Google Scholar] [CrossRef]
- Hernández, L.; Martínez- Fernández, J.; Cañellas, I.; de la Cueva, A.V. Assessing spatio-temporal rates, patterns and determinants of biological invasions in forest ecosystems. The case of acacia species in NW spain. For. Ecol. Manag. 2014, 329, 206–213. [Google Scholar]
- Richardson, D.M.; Daehler, C.C.; Leishman, M.R.; Pauchard, A.; Pyšek, P. Plant invasions: Theoretical and practical challenges. Biol. Invasions 2010, 12, 3907–3911. [Google Scholar] [CrossRef]
- Bricher, P.K.; Lucieer, A.; Shaw, J.; Terauds, A.; Bergstrom, D.M. Mapping sub-antarctic cushion plants using random forests to combine very high resolution satellite imagery and terrain modelling. PLoS ONE 2013, 8, e72093. [Google Scholar] [CrossRef] [PubMed]
- Lu, D.; Weng, Q. A survey of image classification methods and techniques for improving classification performance. Int. J. Remote Sens. 2007, 28, 823–870. [Google Scholar] [CrossRef]
- Xie, Y.; Sha, Z.; Yu, M. Remote sensing imagery in vegetation mapping: A review. J. Plant Ecol. 2008, 1, 9–23. [Google Scholar] [CrossRef]
- Lucas, R.; Blonda, P.; Bunting, P.; Jones, G.; Inglada, J.; Arias, M.; Kosmidou, V.; Petrou, Z.I.; Manakos, I.; Adamo, M.; et al. The earth observation data for habitat monitoring (eodham) system. Int. J. Appl. Earth Observ. Geoinf. 2015, 37, 17–28. [Google Scholar] [CrossRef]
- Blaschke, T. Object based image analysis for remote sensing. ISPRS J. Photogramm. Remote Sens. 2010, 65, 2–16. [Google Scholar] [CrossRef]
- Fava, F.; Pulighe, G.; Monteiro, A.T. Mapping changes in land cover composition and pattern for comparing mediterranean rangeland restoration alternatives. Land Degrad. Dev. 2016, 27, 671–681. [Google Scholar] [CrossRef]
- Teodoro, A.C.; Araujo, R. Comparison of performance of object-based image analysis techniques available in open source software (spring and orfeo toolbox/monteverdi) considering very high spatial resolution data. J. Appl. Remote Sens. 2016, 10, 016011. [Google Scholar] [CrossRef]
- Dirnböck, T.; Dullinger, S.; Gottfried, M.; Ginzler, C.; Grabherr, G. Mapping alpine vegetation based on image analysis, topographic variables and canonical correspondence analysis. Appl. Veg. Sci. 2003, 6, 85–96. [Google Scholar] [CrossRef]
- Gonçalves, J.; Henriques, R.; Alves, P.; Sousa-Silva, R.; Monteiro, A.T.; Lomba, Â.; Marcos, B.; Honrado, J. Evaluating an unmanned aerial vehicle-based approach for assessing habitat extent and condition in fine-scale early successional mountain mosaics. Appl. Veg. Sci. 2016, 19, 132–146. [Google Scholar] [CrossRef]
- Mallinis, G.; Koutsias, N.; Tsakiri-Strati, M.; Karteris, M. Object-based classification using quickbird imagery for delineating forest vegetation polygons in a mediterranean test site. ISPRS J. Photogramm. Remote Sens. 2008, 63, 237–250. [Google Scholar] [CrossRef]
- Breiman, L. Random forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef]
- Cutler, D.R.; Edwards, T.C.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random forests for classification in ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef] [PubMed]
- Mountrakis, G.; Im, J.; Ogole, C. Support vector machines in remote sensing: A review. ISPRS J. Photogramm. Remote Sens. 2011, 66, 247–259. [Google Scholar] [CrossRef]
- Bastos, R.; Monteiro, A.T.; Carvalho, D.; Gomes, C.; Travassos, P.; Honrado, J.P.; Santos, M.; Cabral, J.A. Integrating land cover structure and functioning to predict biodiversity patterns: A hierarchical modelling framework designed for ecosystem management. Landsc. Ecol. 2016, 31, 701–710. [Google Scholar] [CrossRef]
- Deblauwe, V.; Droissart, V.; Bose, R.; Sonké, B.; Blach-Overgaard, A.; Svenning, J.C.; Wieringa, J.J.; Ramesh, B.R.; Stévart, T.; Couvreur, T.L.P. Remotely sensed temperature and precipitation data improve species distribution modelling in the tropics. Glob. Ecol. Biogeogr. 2016, 25, 443–454. [Google Scholar] [CrossRef]
- Pottier, J.; Malenovský, Z.; Psomas, A.; Homolová, L.; Schaepman, M.E.; Choler, P.; Thuiller, W.; Guisan, A.; Zimmermann, N.E. Modelling plant species distribution in alpine grasslands using airborne imaging spectroscopy. Biol. Lett. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Rocchini, D.; Andreo, V.; Förster, M.; Garzon-Lopez, C.X.; Gutierrez, A.P.; Gillespie, T.W.; Hauffe, H.C.; He, K.S.; Kleinschmit, B.; Mairota, P.; et al. Potential of remote sensing to predict species invasions: A modelling perspective. Prog. Phys. Geogr. 2015, 39, 283–309. [Google Scholar] [CrossRef]
- Plummer, S.E. Perspectives on combining ecological process models and remotely sensed data. Ecol. Model. 2000, 129, 169–186. [Google Scholar] [CrossRef]
- Albert, C.H.; Thuiller, W.; Lavorel, S.; Davies, I.D.; Garbolino, E. Land-use change and subalpine tree dynamics: Colonization of larix decidua in french subalpine grasslands. J. Appl. Ecol. 2008, 45, 659–669. [Google Scholar] [CrossRef]
- Fernandes, M.J.D.S.M. Recuperação ecológica de áreas invadidas por acacia dealbata link no vale do rio gerês: Um trabalho de sísifo? UTAD (Universidade de Trás-os-Montes e Alto Douro): Vila Real, Portugal, 2008. [Google Scholar]
- European Economic Area. Invasive alien species indicators in europe a review of streamlining european biodiversity (sebi) indicator 10. Eea Tech. Rep. 2012. [Google Scholar] [CrossRef]
- Vicente, J.R.; Fernandes, R.F.; Randin, C.F.; Broennimann, O.; Gonçalves, J.; Marcos, B.; Pôças, I.; Alves, P.; Guisan, A.; Honrado, J.P. Will climate change drive alien invasive plants into areas of high protection value? An improved model-based regional assessment to prioritise the management of invasions. J. Environ. Manag. 2013, 131, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Inventory, P.N.F. National cartography of burnt areas. National Institute for Nature and Forests Conservation: Lisbon, Portugal, 2014. Available online: http://www.icnf.pt/portal/florestas/dfci/inc/info-geo (accessed on 18 January 2017).
- Tucker, C. Red and photographic infrared linear combinations for monitoring vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Navulur, K. Multispectral Image Analysis Using the Object-Oriented Paradigm. CRC Press, Inc.: Boca Raton, FL, USA, 2006. [Google Scholar]
- Nouri, H.; Beecham, S.; Anderson, S.; Nagler, P. High spatial resolution worldview-2 imagery for mapping ndvi and its relationship to temporal urban landscape evapotranspiration factors. Remote Sens. 2014, 6, 580. [Google Scholar] [CrossRef]
- Wolf, A.F. Using worldview-2 vis-nir multispectral imagery to support land mapping and feature extraction using normalized difference index ratios. In Proceedings of the SPIE 8390, Algorithms and Technologies for Multispectral, Hyperspectral, and Ultraspectral Imagery XVIII, Baltimore, Maryland, USA, 23–27 April 2012.
- Cho, M.A.; Skidmore, A.K. A new technique for extracting the red edge position from hyperspectral data: The linear extrapolation method. Remote Sens. Environ. 2006, 101, 181–193. [Google Scholar] [CrossRef]
- Oumar, Z.; Mutanga, O. Using WorldView-2 bands and indices to predict bronze bug thaumastocoris peregrinus damage in plantation forests. Int. J. Remote Sens. 2013, 34, 2236–2249. [Google Scholar] [CrossRef]
- Vaz, A.S.; Marcos, B.; Gonçalves, J.; Monteiro, A.; Alves, P.; Civantos, E.; Lucas, R.; Mairota, P.; Garcia-Robles, J.; Alonso, J.; et al. Can we predict habitat quality from space? A multi-indicator assessment based on an automated knowledge-driven system. Int. J. Appl. Earth Observ. Geoinf. 2015, 37, 106–113. [Google Scholar] [CrossRef]
- Waser, L.T.; Kuechler, M.; Juette, K.; Stampfer, T. Evaluating the potential of worldview-2 data to classify tree species and different levels of ash mortality. Remote Sens. 2014, 6, 4515–4545. [Google Scholar] [CrossRef]
- Lyons, E.A.; Jin, Y.; Randerson, J.T. Changes in surface albedo after fire in boreal forest ecosystems of interior alaska assessed using modis satellite observations. J. Geophys. Res. Biogeosci. 2008, 113, 912. [Google Scholar] [CrossRef]
- Bunting, P.; Clewley, D.; Lucas, R.M.; Gillingham, S. The remote sensing and gis software library (rsgislib). Comput. Geosci. 2014, 62, 216–226. [Google Scholar] [CrossRef]
- Delaplace, K.L.W.; Van Coillie, F.M.B.; De Wulf, R.R.; Gabriels, D.; De Smet, K.; Ouessar, M.; Belgacem, A.O.; Houcine, T. Object-based assessment of tree attributes of acacia tortilis in bou-hedma, tunisia. In Proceedings of the Third International Conference of Geographic Object-Based Image Analysis (GEOBIA 2010), Ghent, Belgium, 29 June–2 July 2010.
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. Biomod—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Clewley, D.; Bunting, P.; Shepherd, J.; Gillingham, S.; Flood, N.; Dymond, J.; Lucas, R.; Armston, J.; Moghaddam, M. A python-based open source system for geographic object-based image analysis (geobia) utilizing raster attribute tables. Remote Sens. 2014, 6, 6111–6135. [Google Scholar] [CrossRef]
- Congalton, R.G. A review of assessing the accuracy of classifications of remotely sensed data. Remote Sens. Environ. 1991, 37, 35–46. [Google Scholar] [CrossRef]
- Fabian, P.; Gaël, E.V.; Gramfort, A.; Michel, V.; Thirion, B.; Grisel, O.; Blondel, M.; Prettenhofer, P.; Weiss, R.; Dubourg, V.; et al. Scikit-learn: Machine learning in python. J. Mach. Learn. Res. 2011, 12, 2825–2830. [Google Scholar]
- Buchanan, B.P.; Fleming, M.; Schneider, R.L.; Richards, B.K.; Archibald, J.; Qiu, Z.; Walter, M.T. Evaluating topographic wetness indices across central new york agricultural landscapes. Hydrol. Earth Syst. Sci. 2014, 18, 3279–3299. [Google Scholar] [CrossRef]
- Castro-Díez, P.; Godoy, O.; Saldaña, A.; Richardson, D.M. Predicting invasiveness of australian acacias on the basis of their native climatic affinities, life history traits and human use. Divers. Distrib. 2011, 17, 934–945. [Google Scholar] [CrossRef]
- Ferreras, A.E.; Funes, G.; Galetto, L. The role of seed germination in the invasion process of honey locust (gleditsia triacanthos l., fabaceae): Comparison with a native confamilial. Plant Species Biol. 2015, 30, 126–136. [Google Scholar] [CrossRef]
- Western, A.W.; Grayson, R.B.; Blöschl, G.; Willgoose, G.R.; McMahon, T.A. Observed spatial organization of soil moisture and its relation to terrain indices. Water Resour. Res. 1999, 35, 797–810. [Google Scholar] [CrossRef]
- White, J.D.; Running, S.W. Testing scale dependent assumptions in regional ecosystem simulations. J. Veg. Sci. 1994, 5, 687–702. [Google Scholar] [CrossRef]
- Moore, I.D.; Gessler, P.E.; Nielsen, G.A.; Peterson, G.A. Soil attribute prediction using terrain analysis. Soil Sci. Soc. Am. J. 1993, 57, 443–452. [Google Scholar] [CrossRef]
- Sørensen, R.; Zinko, U.; Seibert, J. On the calculation of the topographic wetness index: Evaluation of different methods based on field observations. Hydrol. Earth Syst. Sci. 2006, 10, 101–112. [Google Scholar] [CrossRef]
- Bellingham, P.J.; Sparrow, A.D. Resprouting as a life history strategy in woody plant communities. Oikos 2000, 89, 409–416. [Google Scholar] [CrossRef]
- Pettorelli, N.; Vik, J.O.; Mysterud, A.; Gaillard, J.-M.; Tucker, C.J.; Stenseth, N.C. Using the satellite-derived ndvi to assess ecological responses to environmental change. Trends Ecol. Evol. 2005, 20, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Graham, A. Remote Sensing of Vegetation Principles, Techniques, and Applications; Oxford University Press: Oxford, UK, 2012; pp. 1653–1654. [Google Scholar]
- Pettorelli, N. The Normalized Difference Vegetation Index; OUP Oxford: Oxford, UK, 2013. [Google Scholar]
- Higgins, S.I.; Richardson, D.M.; Cowling, R.M. Validation of a spatial simulation model of a spreading alien plant population. J. Appl. Ecol. 2001, 38, 571–584. [Google Scholar] [CrossRef]
- Immitzer, M.; Atzberger, C.; Koukal, T. Tree species classification with random forest using very high spatial resolution 8-band WorldView-2 satellite data. Remote Sens. 2012, 4, 2661–2693. [Google Scholar] [CrossRef]
- Peerbhay, K.Y.; Mutanga, O.; Ismail, R. Investigating the capability of few strategically placed worldview-2 multispectral bands to discriminate forest species in kwazulu-natal, south africa. IEEE J. Sel. Top. Appl. Earth Observ. Remote Sens. 2014, 7, 307–316. [Google Scholar] [CrossRef]
- Cho, M.A.; Malahlela, O.; Ramoelo, A. Assessing the utility worldview-2 imagery for tree species mapping in south african subtropical humid forest and the conservation implications: Dukuduku forest patch as case study. Int. J. Appl. Earth Observ. Geoinf. 2015, 38, 349–357. [Google Scholar] [CrossRef]
- Yu, Q.; Gong, P.; Clinton, N.; Biging, G.; Kelly, M.; Schirokauer, D. Object-based detailed vegetation classification with airborne high spatial resolution remote sensing imagery. Photogramm. Eng. Remote Sens. 2006, 72, 799–811. [Google Scholar] [CrossRef]
- Grosse-Stoltenberg, A.; Hellmann, C.; Werner, C.; Oldeland, J.; Thiele, J. Evaluation of continuous VNIR–SWIR spectra versus narrowband hyperspectral indices to discriminate the invasive acacia longifolia within a mediterranean dune ecosystem. Remote Sens. 2016, 8, 334. [Google Scholar] [CrossRef]
- Lorenzo, P.; González, L.; Reigosa, M.J. The genus acacia as invader: The characteristic case of acacia dealbata link in europe. Ann. For. Sci. 2010, 67, 101. [Google Scholar] [CrossRef]
- Gassó, N.; Thuiller, W.; Pino, J.; Vilà, M. Potential distribution range of invasive plant species in spain. NeoBiota 2012, 12, 25–40. [Google Scholar]
- Marchante, H.; Freitas, H.; Hoffmann, J.H. Post-clearing recovery of coastal dunes invaded by acacia longifolia: Is duration of invasion relevant for management success? J. Appl. Ecol. 2011, 48, 1295–1304. [Google Scholar] [CrossRef]
- Alba, C.; Skalova, H.; McGregor, K.F.; D’Antonio, C.; Pysek, P. Native and exotic plant species respond differently to wildfire and prescribed fire as revealed by meta-analysis. J. Veg. Sci. 2015, 26, 102–113. [Google Scholar] [CrossRef]
- Dickens, S.J.M.; Allen, E.B. Exotic plant invasion alters chaparral ecosystem resistance and resilience pre- and post-wildfire. Biol. Invasions 2014, 16, 1119–1130. [Google Scholar] [CrossRef]
- Morris, T.L.; Esler, K.J.; Barger, N.N.; Jacobs, S.M.; Cramer, M.D. Ecophysiological traits associated with the competitive ability of invasive australian acacias. Divers. Distrib. 2011, 17, 898–910. [Google Scholar] [CrossRef]
- Rascher, K.G.; Grosse-Stoltenberg, A.; Maguas, C.; Werner, C. Understory invasion by acacia longifolia alters the water balance and carbon gain of a mediterranean pine forest. Ecosystems 2011, 14, 904–919. [Google Scholar] [CrossRef]
- Carvalho-Santos, C.; Nunes, J.P.; Monteiro, A.T.; Hein, L.; Honrado, J.P. Assessing the effects of land cover and future climate conditions on the provision of hydrological services in a medium-sized watershed of portugal. Hydrol. Processes 2016, 30, 720–738. [Google Scholar] [CrossRef]
- Foley, J.A.; Costa, M.H.; Delire, C.; Ramankutty, N.; Snyder, P. Green surprise? How terrestrial ecosystems could affect earth’s climate. Front. Ecol. Environ. 2003, 1, 38–44. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Rationale | Formula | Reference |
|---|---|---|---|
| NDVI | Vegetated areas where non-native invasion occurs can be identified by ratio between red and near-infrared regions. | (NIR − R)/(NIR + R) | [32] |
| TCTw | Enhance the separation between forest/natural vegetation and cultivated vegetated areas. | Uwetness = 0.652 × B + 0.375 × G − 0.639 × R − 0.163 × NIR1 | [33] |
| WBI | Canopy moisture content vary according vegetation and cover types. WBI can help to incorporate this variation and improve non-native invasion mapping. | (B/NIR1) | [12] |
| NDSI | Identify rocky and bare areas, a main feature of landscape. | (G − Y)/(G + Y) | [34,35] |
| REP | Chlorophyll concentration may vary across plants and the REP narrowband index may help to include this feature to better map non-native invasion. | (RE − (NIR2 − R)) | [36,37] |
| BGND | Separate tree and shrubs can help to better map non-native invasion. The normalized blue/green and blue/red ratios enhanced tree dominated areas and crown structure. | (B − G)/(B + G) | [38,39] |
| LSA | Surface albedo vary within vegetation types and stand age. This feature at fine resolution may help to better separate non-native invasion within woody/forest types. | ((Y + R) × 0.35)/2 + (0.7 × (NIR1 + NIR2))/2 − 0.69 | [39,40] |
| FDI | Large reflection of shadow in FDI may help to distinguish better woody from forest types. | NIR1 − (RE + B) | [41,42] |
| Aspect | Exposure influence light interception and distribution of functional types. | ||
| Slope | Slope influences antrophic activities as farmland management options and therefore vegetation distribution patterns. | ||
| Note: For the invasion mapping with random forest, the predictor refers to the average value of each variable in each segment. | |||
| Reference Data | Classified As | |||
|---|---|---|---|---|
| Invasion | Non Invasion | Σ | Producer’s Accuracy | |
| Invasion | 543 | 63 | 606 | 86.8 |
| Non Invasion | 82 | 984 | 1066 | 93.9 |
| Total | 625 | 1047 | 1672 | |
| User’s accuracy | 89.6 | 92.3 | ||
| Overall Acuracy (%) | 91.3 | Kappa Coefficient | 0.81 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monteiro, A.T.; Gonçalves, J.; Fernandes, R.F.; Alves, S.; Marcos, B.; Lucas, R.; Teodoro, A.C.; Honrado, J.P. Estimating Invasion Success by Non-Native Trees in a National Park Combining WorldView-2 Very High Resolution Satellite Data and Species Distribution Models. Diversity 2017, 9, 6. https://doi.org/10.3390/d9010006
Monteiro AT, Gonçalves J, Fernandes RF, Alves S, Marcos B, Lucas R, Teodoro AC, Honrado JP. Estimating Invasion Success by Non-Native Trees in a National Park Combining WorldView-2 Very High Resolution Satellite Data and Species Distribution Models. Diversity. 2017; 9(1):6. https://doi.org/10.3390/d9010006
Chicago/Turabian StyleMonteiro, Antonio T., João Gonçalves, Rui F. Fernandes, Susana Alves, Bruno Marcos, Richard Lucas, Ana Claúdia Teodoro, and João P. Honrado. 2017. "Estimating Invasion Success by Non-Native Trees in a National Park Combining WorldView-2 Very High Resolution Satellite Data and Species Distribution Models" Diversity 9, no. 1: 6. https://doi.org/10.3390/d9010006
APA StyleMonteiro, A. T., Gonçalves, J., Fernandes, R. F., Alves, S., Marcos, B., Lucas, R., Teodoro, A. C., & Honrado, J. P. (2017). Estimating Invasion Success by Non-Native Trees in a National Park Combining WorldView-2 Very High Resolution Satellite Data and Species Distribution Models. Diversity, 9(1), 6. https://doi.org/10.3390/d9010006