Soil Nematodes and Their Prokaryotic Prey Along an Elevation Gradient in The Mojave Desert (Death Valley National Park, California, USA)

Abstract
:1. Introduction
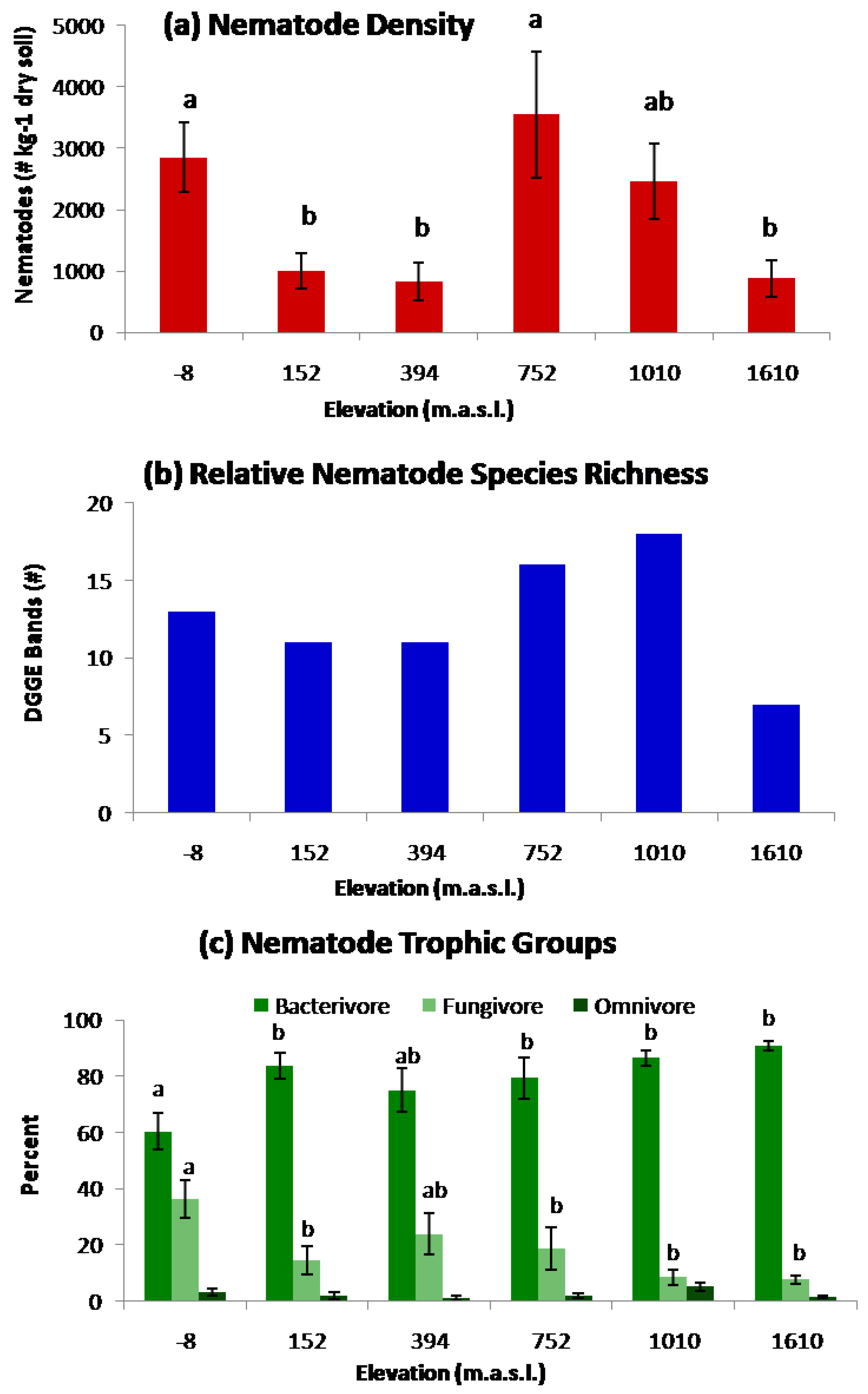
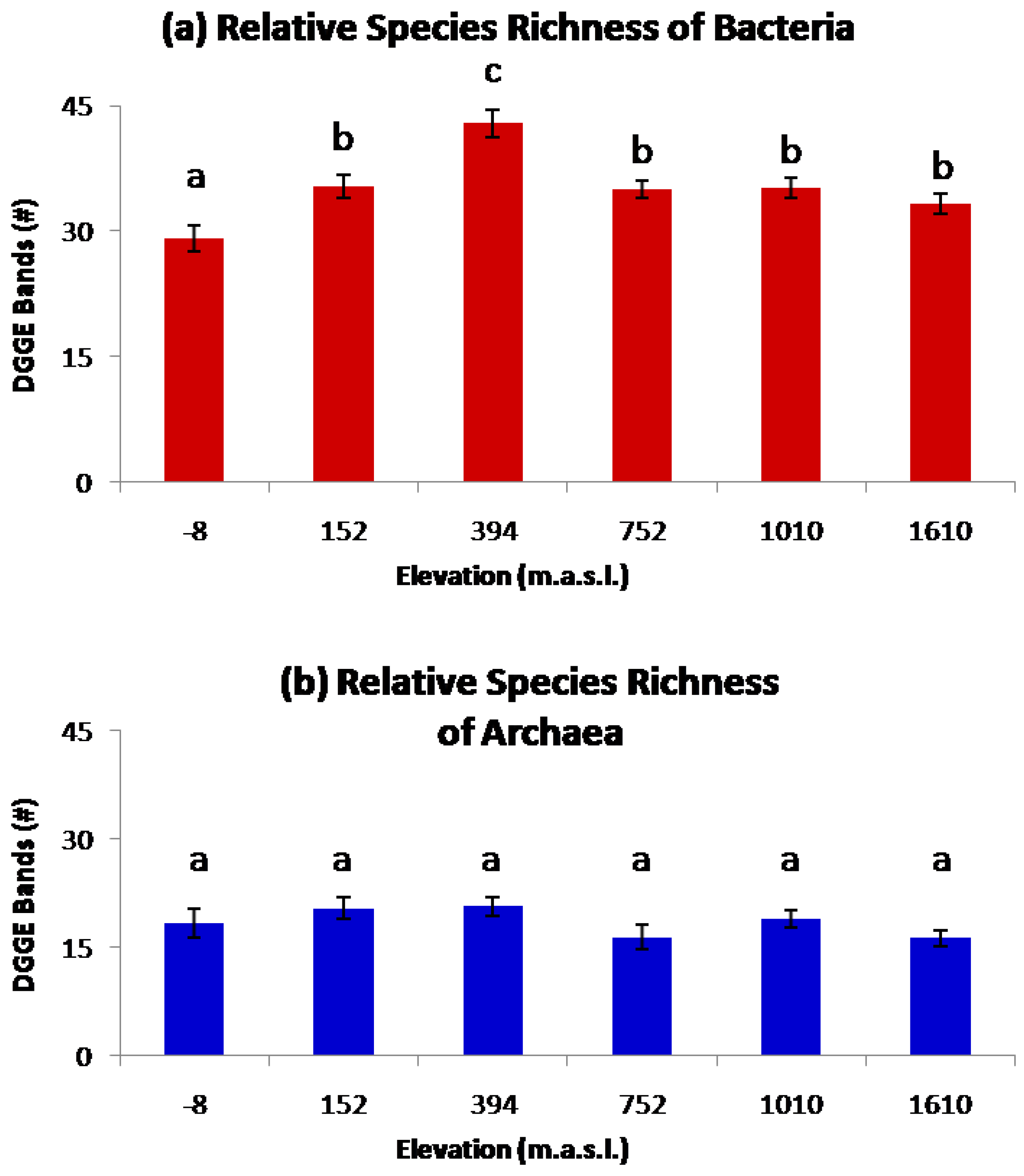
2. Results and Discussion

{kind=link}
{kind=link}
| Site | Elevation (m.a.s.l.) ** | Organic Matter (%, LOI) *** | Gravimetric Soil Moisture (%) *** | pH *** | EC (μS) *** | Respiration (CO2μmol g−1d−1) **** |
|---|---|---|---|---|---|---|
| Sand Dunes | −8 | 0.62 ± 0.07 a | 0.18 ± 0.006 a | 8.66 ± 0.05 a | 183.88 ± 18.76 a | 0.12 ± 0.02 a |
| Unnamed Site | 152 | 1.40 ± 0.10 b | 0.48 ± 0.029 ab | 8.40 ± 0.04 ab | 189.63 ±18.55 a | 0.13 ± 0.01 a |
| Jubilee Pass | 394 | 1.97 ± 0.16 bc | 0.89 ± 0.080 bc | 8.12 ± 0.08 bcd | 176.25 ± 15.72 a | 0.10 ± 0.01 a |
| Darwin Falls | 752 | 2.27 ± 0.38 c | 0.97 ± 0.18 bc | 8.03 ± 0.20 cd | 667.00 ± 169.10 b | 0.16 ± 0.03 a |
| SalsberryPass | 1010 | 3.11 ± 0.18 d | 2.71 ± 0.53 d | 8.22 + 0.08 bc | 241.63 ± 31.85 a | 0.13 ± 0.03 a |
| Dantes View | 1610 | 3.71 ± 0.22 d | 1.50 ± 0.12 c | 7.84 ± 0.07 d | 355.13 ± 44.12 a | 0.12 ± 0.02 a |


| Soil Property | Nematode Density | Bacteria Diversity | Archaea Diversity |
|---|---|---|---|
| Moisture | 0.43 | 0.26 | 0.45 |
| Organic Matter | 0.38 | 0.29 | 0.29 |
| Electrical Conductivity | 0.0028** | 0.36 | 0.023*** |
| pH | 0.21 | 0.097 | 0.407 |
3. Experimental Section
4. Conclusions
Acknowledgments
References
- Bardgett, R.D.; Yeates, G.W.; Anderson, J.M. Patterns and determinants of soil biological diversity. In Biological Diversity and Function in Soil; Bardgett, R.D., Usher, M.B., Eds.; Cambridge University Press: Cambridge, UK, 2005; pp. 100–118. [Google Scholar]
- Fierer, N.; Jackson, R.B. The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. USA 2006, 103, 626–631. [Google Scholar] [CrossRef]
- Waide, R.B.; Willig, M.R.; Steiner, C.F.; Mittelbach, G.; Gough, L.; Dodson, S.I.; Juday, G.P.; Parmenter, R. The relationship between productivity and species richness. Ann. Rev. Ecol. Syst. 1999, 30, 257–300. [Google Scholar] [CrossRef]
- Rosenzweig, M.L. Net primary productivity of terrestrial communities: predictions from climatological data. Am. Nat. 1968, 102, 67–74. [Google Scholar]
- Barrett, J.E.; Virginia, R.A.; Wall, D.H.; Cary, S.C.; Adams, B.J.; Hacker, A.L.; Aislabie, J.M. Co-variation in soil biodiversity and biogeochemistry in Northern and Southern Victoria Land, Antarctica. Antarct. Sci. 2006, 18, 535–548. [Google Scholar] [CrossRef]
- Fierer, N.; McCain, C.M.; Meir, P.; Zimmermann, M.; Rapp, J.M.; Silman, M.R.; Knight, R. Microbes do not follow the elevational diversity patterns of plants and animals. Ecology 2011, 92, 797–804. [Google Scholar] [CrossRef]
- Wu, T.; Ayres, E.; Bardgett, R.D.; Wall, D.H.; Garey, J.R. A molecular study of the worldwide distribution and diversity of soil animals. Proc. Natl. Acad. Sci. USA 2011, 108, 17720–17725. [Google Scholar]
- Death Valley National Park Homepage. Available online: http://www.nps.gov/deva (accessed on 11 August 2012).
- Titus, J.H.; Nowakw, R.S.; Smith, S.D. Soil resource heterogeneity in the Mojave Desert. J. Arid Environ. 2002, 52, 269–292. [Google Scholar] [CrossRef]
- Whitford, W.G. Ecology of Desert Systems; Academic Press: San Diego, CA, USA, 2002. [Google Scholar]
- Freckman, D.W.; Mankau, R. Distribution and trophic structure of nematodes in desert soils. Ecol. Bull. 1977, 25, 511–514. [Google Scholar]
- Treonis, A.M.; Michelle, E.H.; O’Leary, C.A.; Austin, E.E.; Marks, C.B. Identification and localization of food source microbial nucleic acids inside soil nematodes. Soil Biol. Biochem. 2010, 42, 2005–2011. [Google Scholar] [CrossRef]
- Bates, S.; Caporaso, J.G.; Walters, W.A.; Knight, R.; Fierer, N. Examining the global distribution of dominant archaeal populations in soil. ISME J. 2011, 5, 908–917. [Google Scholar] [CrossRef]
- Muyzer, G.; Dewaal, E.C.; Uitterlinden, A.G. Profiling of complex microbial-populations by denaturing gradient gel electrophoresis analysis of polymerase chain reaction-amplified genes coding for 16S rRNA. Appl. Environ. Microb. 1993, 59, 695–700. [Google Scholar]
- vreås, L.; Jensen, S.; Daae, F.L.; Torsvik, V. Microbial community changes in a perturbed agricultural soil investigated by molecular and physiological approaches. Appl. Environ. Microb. 1998, 64, 2739–2742. [Google Scholar]
- Oros-Sichler, M.; Costa, R.; Heuer, H.; Smalla, K. Molecular fingerprinting techniques to analyze soil microbial communities. In Modern Soil Microbiology, 2nd; Elsas, J.D.V., Jansson, J.K., Trevors, J.T., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 355–377. [Google Scholar]
- Foucher, A.L.J.L.; Bongers, T.; Noble, L.R.; Wilson, M.J. Assessment of nematode biodiversity using DGGE of 18S rDNA following extraction of nematodes from soil. Soil Biol. Biochem. 2004, 36, 2027–2032. [Google Scholar] [CrossRef]
- Takemoto, S.; Niwa, S.; Okada, H. Effect of Storage Temperature on Soil Nematode Community Structures as Revealed by PCR-DGGE. J. Nematol. 2010, 42, 324–331. [Google Scholar]
- Wang, J.; Yang, D.; Zhang, Y.; Shen, J.; van der Gast, C.; Hahn, M.W.; Wu, Q. Do patterns of bacterial diversity along salinity gradients differ from those observed for macroorganisms? PLoS One 2011, 6, 1–8. [Google Scholar]
- Freckman, D.W.; Virginia, R.A. Plant-feeding nematodes in deep-rooting desert ecosystems. Ecology 1989, 70, 1665–1678. [Google Scholar] [CrossRef]
- Madin, K.A.C.; Crowe, J.H. Anhydrobiosis in nematodes: Carbohydrate and lipid metabolism during dehydration. J. Exp. Zool. 1975, 193, 335–342. [Google Scholar] [CrossRef]
- Treonis, A.M.; Wall, D.H.; Virginia, R.A. The use of anhydrobiosis by soil nematodes in the Antarctic Dry Valleys. Funct. Ecol. 2000, 14, 460–467. [Google Scholar] [CrossRef]
- Crowe, J.H.; Hoekstra, F.A.; Crowe, L.M. Anhydrobiosis. Annual Rev. Physiol. 1992, 54, 579–599. [Google Scholar] [CrossRef]
- Treonis, A.M.; Wall, D.H.; Virginia, R.A. Invertebrate biodiversity in Antarctica Dry Valley soils and sediments. Ecosystems 1999, 2, 482–492. [Google Scholar] [CrossRef]
- Adams, B.J.; Wall, D.H.; Gozel, U.; Hogg, I.D. The southernmost worm, Scottnema lindsayae (Nematoda): diversity, dispersal and ecological stability. Polar Biol. 2006, 30, 809–815. [Google Scholar]
- Steinberger, Y.; Liang, W.; Savinka, E.; Meshi, T.; Barness, G. Nematode community composition and diversity associated with a topoclimatic transect in a rain shadow desert. Eur. J. Soil Biol. 2001, 37, 315–320. [Google Scholar] [CrossRef]
- Powers, L.E.; Ho, M.; Freckman, D.W.; Virginia, R.A. Distribution, community structure, and microhabitats of soil invertebrates along an elevational gradient in Taylor Valley, Antarctica. Arctic Alpine Res. 1998, 30, 133–141. [Google Scholar] [CrossRef]
- Bryant, J.; Lamanna, C.; Morlon, H.; Kerkhoff, A.J.; Enquist, B.J.; Green, J.L. Microbes on mountainsides: Contrasting elevational patterns of bacterial and plant diversity. Proc. Natl. Acad. Sci. USA 2008, 105, 11505–11511. [Google Scholar]
- Singh, D.; Takahashi, K.; Kim, M.; Chun, J.; Adams, J.M. A hump-backed trend in bacterial diversity with elevation on Mount Fuji, Japan. Microb. Ecol. 2012, 63, 429–437. [Google Scholar] [CrossRef]
- Kushner, D.J. The Halobacteriaceae. In The Bacteria; Woese, C.R., Wolfe, R.S., Eds.; Academic Press, Inc.: New York, NY, USA, 1985; Volume 8, pp. 171–214. [Google Scholar]
- Fierer, N.; Breitbart, M.; Nulton, J.; Salamon, P.; Lozupone, C.; Jones, R.; Robeson, M.; Edwards, R.A.; Felts, B.; Rayhawk, S.; et al. Metagenomic and small-subunit rRNA analyses reveal the genetic diversity of bacteria, archaea, fungi, and viruses in soil. Appl. Environ. Microb. 2007, 73, 7059–7066. [Google Scholar] [CrossRef]
- Gamfeldt, L.; Hillebrand, H.; Jonsson, P.R. Species richness changes across two trophic levels simultaneously affect prey and consumer biomass. Ecol. Lett. 2005, 8, 696–703. [Google Scholar] [CrossRef]
- Behmer, S.T.; Joern, A. Coexisting generalist herbivores occupy unique nutritional feeding niches. Proc. Natl. Acad. Sci. USA. 2008, 105, 1977–1982. [Google Scholar] [CrossRef]
- Zibilske, L.M. Carbon mineralization. In Methods of Soil Analysis, Part 2: Microbiological and Biochemical Properties; Weaver, R.W., Ed.; Soil Science Society of America: Madison, WI, USA, 1994; pp. 835–863. [Google Scholar]
- Freckman, D.W.; Baldwin, J.G. Nematoda. In Soil Biology Guide; Dindal, D.L., Ed.; John Wiley & Sons: New York, NY, USA, 1990; pp. 155–200. [Google Scholar]
- Yeates, G.W.; Bongers, T.; de Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding-habits in soil nematode families and genera - an outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar]
- Jenkins, W.R. A rapid centrifugal-flotation technique for separating nematodes from soil. Plant Dis. Rep. 1964, 48, 692. [Google Scholar]
- Blaxter, M.L.; de Ley, P.; Garey, J.; Liu, L.X.; Scheldeman, P.; Vierstraete, A.; Vanfleteren, J.R.; Mackey, L.Y.; Dorris, M.; Frisse, L.M.; et al. A molecular evolutionary framework for the phylum Nematoda. Nature 1998, 392, 71–75. [Google Scholar] [CrossRef]
- Okada, H.; Oba, H. Comparison of nematode community similarities assessed by polymerase chain reaction-denaturing gradient gel electrophoresis (PCR-DGGE) and by morphological identification. Nematology 2008, 10, 689–700. [Google Scholar] [CrossRef]
- Muyzer, G.; Teske, A.; Wirsen, C.O.; Jannasch, H.W. Phylogenetic relationships of Thiomicrospira species and their identification in deep-sea hydrothermal vent samples by denaturing gradient gel-electrophoresis of 16S rDNA fragments. Arch. Microbiol. 1995, 164, 165–172. [Google Scholar] [CrossRef]
- Manganelli, M.; Malfatti, F.; Samo, T.J.; Mitchell, B.G.; Wang, H.; Azam, F. Major role of microbes in carbon fluxes during austral winter in the southern Drake Passage. PLoS One 2009, 4, 1–11. [Google Scholar]
- DeLong, E.F. Archaea in coastal marine environments. Proc. Natl. Acad. Sci. USA 1992, 89, 5685–5689. [Google Scholar] [CrossRef]
- Raskin, L.; Stromley, J.M.; Rittmann, B.E.; Stahl, D.A. Group-specific 16S rRNA hybridization probes to describe natural communities of methanogens. Appl. Environ. Microbiol. 1994, 60, 1232–1240. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Treonis, A.; Sutton, K.; Kavanaugh, B.; Narla, A.; McLlarky, T.; Felder, J.; O’Leary, C.; Riley, M.; Pikus, A.; Thomas, S. Soil Nematodes and Their Prokaryotic Prey Along an Elevation Gradient in The Mojave Desert (Death Valley National Park, California, USA). Diversity 2012, 4, 363-374. https://doi.org/10.3390/d4040363
Treonis A, Sutton K, Kavanaugh B, Narla A, McLlarky T, Felder J, O’Leary C, Riley M, Pikus A, Thomas S. Soil Nematodes and Their Prokaryotic Prey Along an Elevation Gradient in The Mojave Desert (Death Valley National Park, California, USA). Diversity. 2012; 4(4):363-374. https://doi.org/10.3390/d4040363
Chicago/Turabian StyleTreonis, Amy, Kelsey Sutton, Brendan Kavanaugh, Archana Narla, Timothy McLlarky, Jasmine Felder, Cecilia O’Leary, Megan Riley, Alyxandra Pikus, and Sarah Thomas. 2012. "Soil Nematodes and Their Prokaryotic Prey Along an Elevation Gradient in The Mojave Desert (Death Valley National Park, California, USA)" Diversity 4, no. 4: 363-374. https://doi.org/10.3390/d4040363
APA StyleTreonis, A., Sutton, K., Kavanaugh, B., Narla, A., McLlarky, T., Felder, J., O’Leary, C., Riley, M., Pikus, A., & Thomas, S. (2012). Soil Nematodes and Their Prokaryotic Prey Along an Elevation Gradient in The Mojave Desert (Death Valley National Park, California, USA). Diversity, 4(4), 363-374. https://doi.org/10.3390/d4040363