Ecological Processes and Contemporary Coral Reef Management

College of Science and Mathematics, University of the Virgin Islands, #2 Brewer’s Bay, St. Thomas, Virgin Islands 00802, 9990, USA
Diversity 2010, 2(5), 717-737; https://doi.org/10.3390/d2050717
Submission received: 8 March 2010
/
Revised: 14 April 2010
/
Accepted: 28 April 2010
/
Published: 11 May 2010
(This article belongs to the Special Issue Spatial and Temporal Benthic Diversity Patterns)
{kind=link}
{kind=link}
{kind=link}
Abstract
:Top-down controls of complex foodwebs maintain the balance among the critical groups of corals, algae, and herbivores, thus allowing the persistence of corals reefs as three-dimensional, biogenic structures with high biodiversity, heterogeneity, resistance, resilience and connectivity, and the delivery of essential goods and services to societies. On contemporary reefs world-wide, however, top-down controls have been weakened due to reduction in herbivory levels (overfishing or disease outbreak) while bottom-up controls have increased due to water quality degradation (increase in sediment and nutrient load) and climate forcing (seawater warming and acidification) leading to algal-dominated alternate benthic states of coral reefs, which are indicative of a trajectory towards ecological extinction. Management to reverse common trajectories of degradation for coral reefs necessitates a shift from optimization in marine resource use and conservation towards building socio-economic resilience into coral reef systems while attending to the most manageable human impacts (fishing and water quality) and the global-scale causes (climate change).
1. Coral Reefs
Coral reef ecosystems are biogenic, massive, three-dimensional structures that develop in well-lit, warm, aragonite-saturated, tropical and subtropical waters (30ºN to 30ºS), either around the coast (fringing reefs), at a distance from the coast (barrier reefs) or on top of volcanic islands (atolls). With a global cover of only 0.1–0.5% of the ocean floor they constitute essential life-support systems by providing essential goods (e.g., fish protein, building materials) and services (e.g., tourism, protection from waves and erosion, land-building) to more than 100 countries and tens of millions of people world-wide [1], which are estimated to be worth $607,500 per km2 per year [2]. These goods and services derive from the interplay of exceptionally high genetic-, species-, and habitat-level diversity, gross productivity, and functional complexity at the surface, only a few cm thick, living veneer of a coral reef [3]. The relatively high gross productivity of coral reefs is based upon complex and efficient biological recycling, resulting in a high retention and exchange of nutrients within the diverse organisms that make up the coral reef community [3].
Inorganic carbon is fixed by primary producers (unicellular, free-living and symbiotic with reef organisms algae; fleshy, filamentous, prostrate and frondose, and crustose coralline algae) through photosynthesis and, then, is partitioned between the bioconstructional (limestone, coral rubble, sediments, skeletal elements of foraminifera, mollusks and echinoderms) and the trophic (producers-consumers-decomposers) pathways [4]. Cnidarian, scleractinian corals are the primary bioconstructors, or foundation species, on reefs, that generate the porous, three-dimensional, wave-resisting, made of limestone, physical framework, which, in turn, facilitates a hierarchy of numerous associated (through shelter or/and nutrition) species [5,6]. Thus, the greatest contribution of corals is in the provision of structure and the accumulation of carbonate for reef growth as either primary (abundant, large size, robust colonies) or secondary framework builders. Another bioconstructor group, the crustose coralline algae form the protective algal ridge/reef crest, cement the dead coral and other carbonate fragments into a stable framework through calcification and, prevent propagules of fouling organisms from colonizing, and induce larval settlement for many benthic organisms [7]. Bioconstruction activity is affected by modifier groups, namely calcification enhancers (symbiotic, unicellular dinoflagellates within coral endodermis that effectively recycle nutrients), bioeroders (filamentous algae, fish and invertebrates that bore into living and dead framework or etch, scrape or nip into its surface) and sediment operators (mollusks, pollychaeta worms, holothurians and fish). Bioeroding fishes, for example, remove dead tissue from corals [8] exposing the hard, reef matrix for settlement of coralline algae and corals. Scrapers directly remove algae and sediment by close cropping, facilitating settlement, growth and survival of coralline algae and corals [9]. Grazers remove seaweed, reducing coral overgrowth and shading by macro-algae [10]. Facilitators are grazing herbivores (e.g., sea urchins, crown-of-thorns starfish, parrotfishes), whose actions directly affect the apportionment of carbon between and within bioconstructional and trophic pathways [4]. Parrotfish have several crucial roles in the dynamics of tropical reefs: they graze fleshy seaweeds that compete with juvenile and adult corals for space; they erode dead coral skeletons and generate reef sediments; and they are an important trophic link between their natural predators and algal primary producers [8]. Fish functional groups are generally synonymous with guilds of species from different trophic levels within a food chain (e.g., predators and herbivores), reflecting their role as a major conduit for the flow of energy from primary producers to consumers on reefs. Herbivores reduce algal standing stock but not all herbivorous fish are the same, i.e., they do not feed on the same algal component (epilithic algal matrix, macroalgae, algal film on sand) in the same way (herbivore, omnivore, detritivore) [11]. Although there is little evidence for direct competitive interactions [10], corals compete with algae for the limiting resources of space and light on reef benthos. Corals while preyed upon by a few omnivorous fishes and specialist invertebrates (e.g., the crown-of-thorns sea star Acanthaster planci, corallivorous gastropods such as Drupella spp.), generally achieve dominance under the controls of high herbivory and low nutrient concentrations [12]. Through direct predation (herbivory and carnivory) or filter feeding, carbon is transferred to detritus, where decomposers recycle nutrients back into the coral reef system. Organic matter may also be imported/exported in the form of detritus, phytoplankton, zooplankton and vertebrates leading to stronger or weaker relationships with adjacent ecosystems, namely mangroves and seagrasses [13,14]. Corals and fishes play multiple functional roles and, thus, support several different processes on coral reefs. These functional groups also have strikingly divergent ecologies and life histories, and exhibit marked differences in their susceptibility to hurricanes, sedimentation and other disturbances [15]. The extent to which reefs possess these functional groups is central to retain critical functions and provide useful goods and services [4]. Ultimately, coral reef processes and functions depend on and, at the same time, determine benthic structure, i.e., composition and relative abundance of functional groups.
The coral reefs of the Atlantic, the Caribbean and the Indo-Pacific do not differ fundamentally in their structural forms, their habitats and the interactions among their species [16], even though they exhibit striking and profoundly important regional differences in the species richness, functional composition, and dynamics [17,18]. For both fishes and corals, Caribbean reefs have only a fraction of the number of species found on the Great Barrier Reef alone, approximately 28% for fishes and 14% for corals due to the evolutionary history of isolation and loss of taxa in the Caribbean basin [19,20]. Main reef builders in the Indo-Pacific are a number of table Acropora corals along with massive Faviids and Porites spp. [21], while in the Caribbean the main reef builders, at least until 1983, were the elkhorn Acropora palmata and the staghorn Acropora cervicornis corals [22]. The opportunistic Pocillopora damicornis [23] is the ubiquitous coral species in the Red Sea [24]. Main algal grazers are herbivorous fishes in the Indo-Pacific, while, again until 1983, the sea urchin Diadema antillarum was the main algal grazer in the Caribbean, functionally replacing overfished grazers in the Caribbean [25,26,27]. Feather stars (Echinodermata, Chinoidea), which are a diverse and important filter-feeding component of the coral reef benthos on western Pacific reefs, are entirely absent from central and eastern Pacific reefs [17]. In the latter case, the transfer of nutrients and organics occurs without them, the role presumably being taken up entirely by other benthic filter feeders and planktivorous fishes [17]. Interestingly, while in the Indo-Pacific the majority of new coral recruits belong to the genus Acropora, main recruits in the Caribbean are Favia fragum, Agaricia agaricites, Porites asteroides and Siderastrea sp., which do not function as reef builders on Caribbean reefs [25]. Although corals in the Caribbean can achieve high adult numbers and cover that are comparable to the Great Barrier Reef (GBR), recruitment rates are orders of magnitude lower [28]. Thus, it has been predicted that reef resilience, i.e., recovery after a major environmental perturbation [29], might require longer periods of time in the Caribbean than on the GBR [17,18]. The diadematid disease of 1981 in the Pacific was apparently contained within the Hawaiian islands, while the diadematid disease of 1983 spread throughout the western Atlantic from Panama to Bermuda in about one year. Incidents of coral bleaching or disease of sponges have occurred over large areas of the Caribbean. Similarly, outbreaks of corallivore Acanthaster planci were generally restricted to individual provinces within the Indo-Pacific (e.g., the GBR, the Ryukyus, Samoa, Hawaii). Furthermore, except from the small seas on the Sunda Shelf, the Caribbean is far more terrestrially influenced than are most coral reef habitats in the IndoPacific [18]. Surrounded on three sides by land, influenced by major drainage basins from two continents, and containing a large number of islands, many of which are substantial in size and/or have a high human population density, the Caribbean as a whole may have made the operational transition from a partially “oceanic” to an effectively “inshore” basin [18].
Many coral reefs are affected by a number of acute natural disturbances, such as hurricanes, flooding, and outbreaks of coral predators, which lead to the creation of new “vacant” spaces due to mortality, maintenance of high diversity on coral reef communities, re-arrangement of benthic components, and successional regression towards a climax community [30]. Given time, coral reefs can recover to pre-disturbance structure [31,32,33] and have persisted on an evolutionary time scale [34,35]. Depending on location and strength of hydrodynamic activity a coral reef may act as a source of marine larvae for (a small or large area of) reefs downstream or as a sink of marine larvae from (a small or large area of) reefs upstream [36]. Nevertheless, replenishment of populations on coral reefs seem to base primarily on self-seeding since coral reefs are connected through hydrodynamic activity, in general, over moderate distance (tens to hundreds of kilometres) over tens to hundreds of years, while genetic mixing occurs over thousands of kilometres and over thousands of years [37,38]. Studies conducted at the largest spatial (hundreds of kilometres) and temporal (thousands of years) scales within biogeographic provinces show intermediate variance while studies conducted at intermediate spatial (tens of kilometres) and temporal (hundreds of years) scales show high degrees of order in coral composition [39]. Studies conducted on small spatial (<1 km) and temporal (<10 yrs) scales show the greatest degrees of variability in community composition for corals and fishes due to physical controls (light, water movement, sediment; [40]), biological interactions (herbivory, competition for space; [41]), disturbance (hurricanes, outbreaks of predators, disease) frequency [30], and recruit supply and post-recruitment mortality [42] as the interplay of multiple stochastic environmental and demographic factors [43]. On a whole single reef, however, there are distinct zones, i.e., belts of reef, usually a few meters to tens of meters wide that have characteristic combinations of substratum, benthic assemblage and fish assemblage; they occur down the sides of reefs and horizontally across their tops [44].
The balance among the critical groups of corals, algae, and herbivores underpin the formation of three-dimensional coral reefs, which are characterized by high biodiversity, heterogeneity, resistance to change at geological time scales, resilience to disturbance at ecological timescales, and by moderate and high connectivity at ecological and geological time scales, respectively.
2. The Coral Reef Crisis
A global summary of the magnitude of the chronic (in geologic time) degradation of coral reef ecosystems based on records of the status and trends of seven major guilds of carnivores, herbivores, and architectural species from 14 regions found that large animals declined before small animals and foundation species, and Atlantic reefs declined before reefs in the Red Sea and Australia [45]. The trajectories, however, of decline were strikingly similar worldwide pointing at the pervasive (in time and space) synergistic effects of fishing and land-derived pollution [45]. All reefs were substantially degraded long before we started gaining an understanding of their structure and function. The best managed coral reefs in the world, the Great Barrier Reef, were the closest to pristine, yet one-quarter to one-third away from ecological extinction [45]. The hardest hit was the Caribbean province while coral reefs of the Red Sea laid somewhere between reefs of the GBR and the Caribbean along the axis of ecological extinction. A recent report shows that about half of the coral reef ecosystem resources within the United States and Pacific Freely Associated States jurisdiction are considered by scientists to be in ‘poor’ or ‘fair’ condition and have declined over time due to several natural and anthropogenic threats [46]. Another consensus of opinion is that one-third of reef-building corals face elevated risk of extinction from climate change and local impacts and that the loss of reef ecosystems would lead to large-scale loss of global biodiversity [47]. A massive, Caribbean-wide decline of corals has been documented through meta-analysis with the average hard coral cover on reefs being reduced by 80%, from about 50% to 10% cover, during only the last three decades [48]. The rate and extent of coral loss in the Indo-Pacific (1% over the last twenty years and 2% between 1997 and 2003 or 3,168 km2 per year) was found greater than expected, too [49]. Coral cover was also surprisingly uniform among subregions (region-wide average was only 22.1% in 2003), suggesting that coral decline is a general global phenomenon, and declined decades earlier than previously assumed, even on the GBR [49].
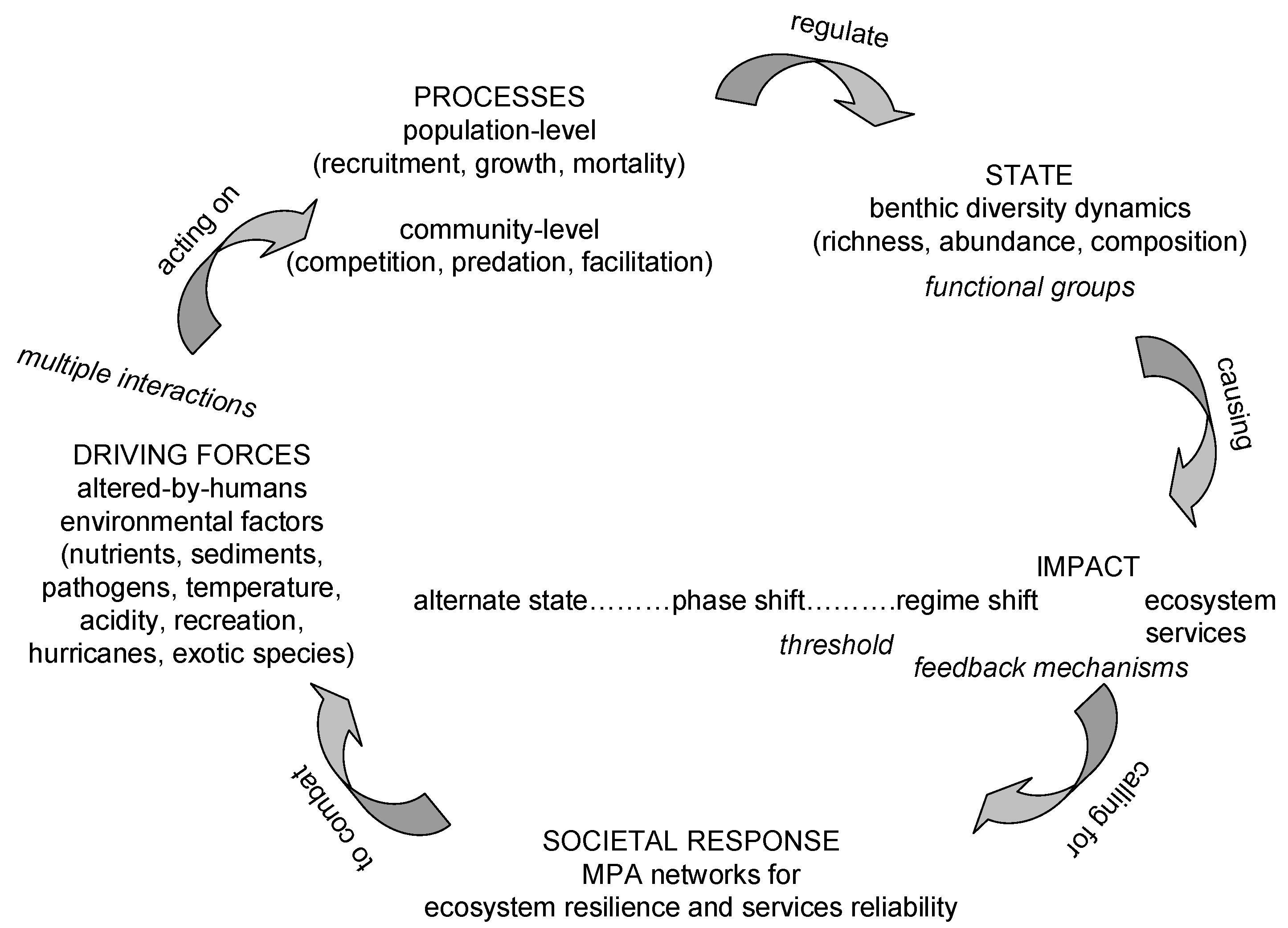
Human population growth has led to unprecedented, both in extent and magnitude, chronic, accumulating and interacting alterations in physical, chemical and biological aspects of the ambient environment of coral reefs world-wide. This has been affected through increase in nutrient and sediment loads due to sewage, agriculture, land-clearing, and coastal development; removal of carnivorous and herbivorous fish due to overfishing; and climate change driven increase in bleaching, disease outbreak and acidification [50,51]. These alterations of the environment of coral reefs acting on population-level (recruitment, growth, mortality) and community-level (competition, energy flow through food webs, facilitation) processes modify benthic diversity dynamics (richness, abundance and composition) of functional groups, leading to transitions among alternate states in community structure of coral reefs named “phase shifts” (Scheme I). The initial and most frequent phase shift observed appear to be from a coral-dominated to an algal-dominated alternate state in community composition on coral reefs; it is mediated through change in top-down controls due to overfishing in most cases; and, again in most but not all cases, it is triggered by widespread coral mortality [52,53]. Depending on the persistence and magnitude of the disturbance, further sliding to a sea urchin barren state and finally to a rock state has been reported [54,55,56]. Phase shifts to corallimorpharia, soft corals or sponges have also been documented and they have been attributed to changes in bottom-up processes due to water quality degradation [57]. Positive or negative feedback mechanisms/loops reinforce the persistence or reversal, respectively, of the alternate state [58,59]. Once thresholds or “tipping points” are exceeded phase shifts turn into persistent “regime shifts” [60,61,62]. The latter are associated with changes in ecosystem goods and services provided by coral reefs, such as dramatic coral decline, local depletions of reef fisheries, and deficit of reef accretion compared to physical and biological erosion that eventually compromise options for social and economic development [63]. In the Caribbean province, increase in nutrients due to agriculture and sewage, loss of macro-fauna, reduced fish stocks, a shift from fish-dominated to echinoid-dominated herbivory as the ecological role of herbivorous fishes was increasingly replaced by a single species of echinoid, destructive overgrazing and bioerosion by food-limited sea urchins, and reduced coral recruitment constitute the trajectory of ecological symptoms that led to a regime shift from coral reefs to algal reefs throughout the province [64]. While the relative importance of top-down (nutrients) vs bottom-up (herbivory) changes on Caribbean reefs is still debated [12], a recent region-wide quantitative assessment of drivers of regime shifts in the Caribbean, indicated that herbivorous fish and Diadema were significant drivers of macroalgae abundance although their relative roles were smaller than that caused by cultivated land [65]. When re-assessing existing evidence on competitive interactions between corals and algae, the importance of the pre-condition of extensive coral mortality before macroalgae proliferation, i.e., space-availability, was stressed [66]. Furthermore, extensive coral mortality is a harbinger of coral recruitment failure. Regarding the stability of macroalgal communities on Caribbean coral reefs, modelling of a Caribbean forereef predicted that the stability of both coral-depauperate and coral dominated reef states on equilibrial time scales of decades to centuries and their emergence depend on the presence or absence of a bottleneck in coral recruitment, which, in turn, is determined by threshold levels of grazing intensity, among other process variables [59]. Even if local conditions improve, however, it should not be assumed that coral reef reorganization will resume exactly as before. It has been projected that the loss of brood stocks on degraded reefs is likely to select against self-seeding species and to shift the taxonomic composition of recruits in favour of those with longer planktonic durations, with potentially far-reaching consequences for community structure [64]. Although a number of studies have shown recent increases in coral cover [67,68] and recruitment [69], some have also reported a shift from communities dominated by framework builders such as Acropora spp. and Montastrea spp. toward those dominated by non-framework builders, such as Agaricia spp. and Porites spp., and sponges [70]. The long-term consequences of such species shifts are unknown, although some current opinion suggests that they may have detrimental consequences for the ability of coral reefs to keep pace with rising sea levels and temperatures, because many such opportunistic species are highly susceptible to temperature shifts and storm damage [71].
Scheme I.
The Driver Pressure State Impact Response model applied to explicate change on contemporary coral reefs.
Scheme I.
The Driver Pressure State Impact Response model applied to explicate change on contemporary coral reefs.
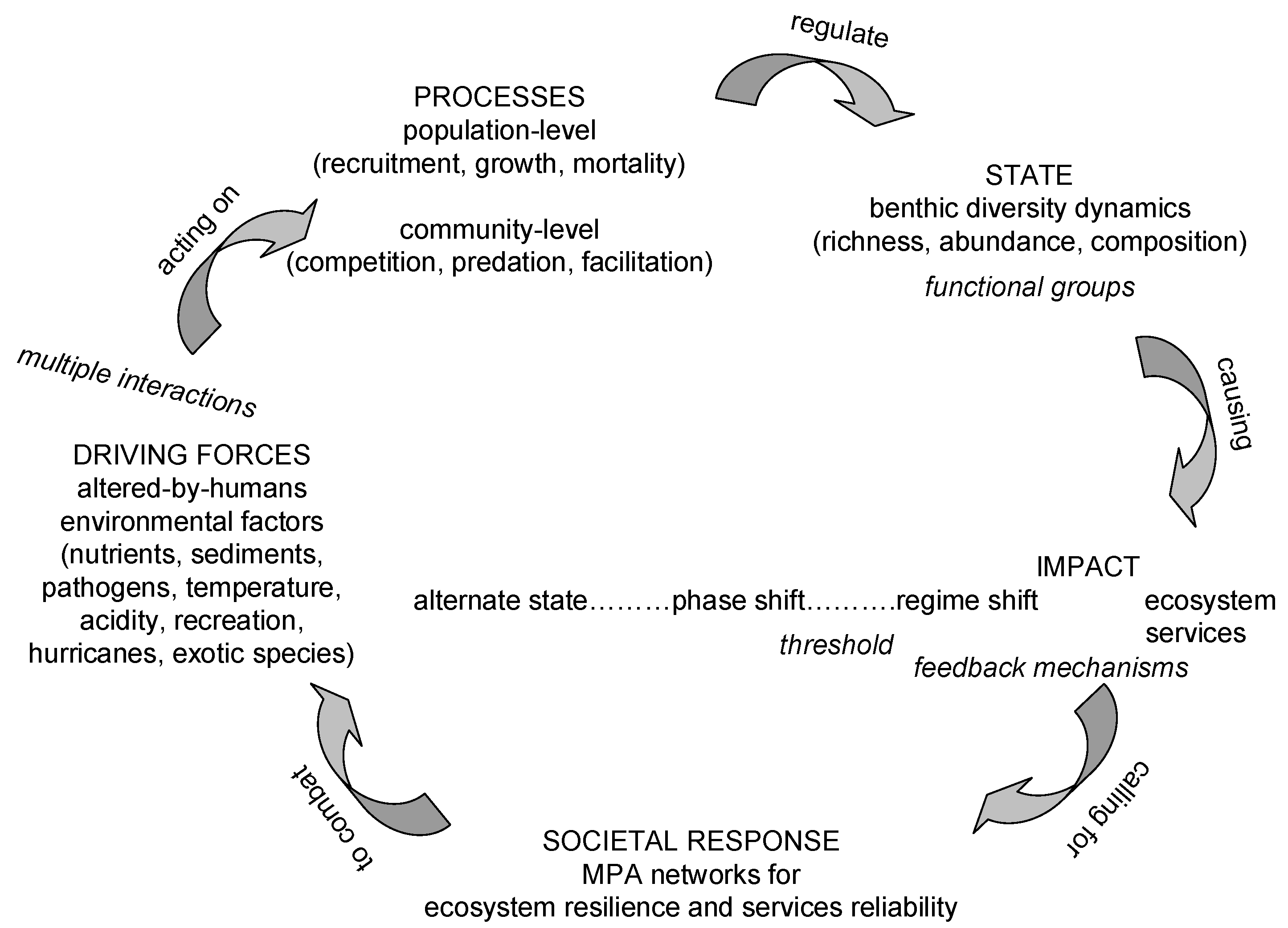
Reversing the alternate state may require restoring the state variables far beyond the threshold value, where the shift initially occurred; a pattern known as hysteresis [61]. The effect of hysteresis is particularly strong when phase shifts take place at large scales or involve large size, long living species, such as corals, or involve ecosystems with already compromised resistance and resilience to new perturbations due to chronic exposure to gradual alterations, as is the case with coral reefs [72,73]. There is some evidence that a similar hysteresis may apply to the succession of macroalgae, once a phase shift has been initiated—the macroalgae becoming larger and increasingly more resistant to perturbations and to infiltration by coral recruits, given a reasonable chance to proliferate [48]. It is alarming that although there is documentation of examples of a cascade of multiple phase shifts from coral reefs to algal reefs to barren reefs [74,75], there are only few examples of a reversal from macroalgal to scleractinian dominance [67,76,77]. Thus, avoiding changes in ecosystem processes and feedback mechanisms in the first place may seem more effective than restoring degraded ecosystem states once regime shifts have occurred [78].
Concurrently, we have little understanding of even the basic shape of the relationship between reef ecosystem resistance and resilience to phase shifts and key emergent and interrelated properties of reef ecosystems, such as trophic structure, biodiversity, heterogeneity, and connectivity [79]. The best-understood are probably aspects of coral resistance and resilience related to the effects of overfishing, degraded water quality, and increased macroalgal abundance on coral recruitment (resilience), and disease susceptibility (resistance). Increase in sediment load impacts primarily photosynthesis and calcification while increase in nutrient load impacts primarily fertilization success and larval development and survival of the reef-building corals [80]. Increase in nutrients also directly stimulates harmful fleshy algal blooms and inhibits the growth of ecologically beneficial reef-building corals while it indirectly influences competitive outcomes [12] and coral susceptibility to disease [81,82]. Herbivorous vertebrates and invertebrates regulate algal abundance and promote coral larval recruitment on hard substrates and, thus, facilitate coral over algal dominance on tropical and sub-tropical reefs provided algae are palatable to fish [83,84]. Chronic, indiscriminate overharvesting can alter a reef’s trophic structure and the redundancy and functional diversity within the critical group of fish on reefs [26,64]. Large amounts of macroalgae may also destabilize microbial communities [86], either by changing water chemistry near coral surfaces [82] or by serving as a reservoir for pathogens [87]. Steadily rising background sea temperature exposes corals to an increasingly hostile environment manifested through increased bleaching frequencies [88] in spite of their significant scope for physiological adaptation [89,90,91]. Robust experiments have shown dramatic declines in coral calcification rates and post-settlement survival and growth of coral larvae [92,93] with increase in acidity of their ambient environment. The cumulative impact of multiple stressors on the sedentary coral populations on reefs may involve narrowing acclimatization ranges, species pools, size of brooding stock, recruitment rates and eventually functional redundancy and response diversity within this critical functional group [15,50,51,94,95]. Coral loss inevitably leads to the decline of the plants and animals that live in the biogenic structures created by such foundation species, and contributes to the overall degradation of the coral reef ecosystem [5,79,96]. In the Caribbean, the demise of the two foundation corals species, the staghorn Acropora cervicornis and the tall, tabular coral Acropora palmata, which were abundant and widespread, commonly comprising more than 30–50% of the total coral cover down to a depth of 20 m, led to the demise of two major shallow-water reef habitats: the elkhorn ‘palmata zone’ and the staghorn ‘cervicornis zone’ [64].
Areas of low species richness or ‘cool spots’, are expected to be less resistant to phase shifts [97]. Low-diversity reefs, such as in the Caribbean Basin, the Eastern Pacific, and many high-latitude or remote locations in the Indo-Pacific have low functional redundancy, where functional groups may be represented by a single species. In these systems, minor changes in biodiversity can have a major impact on ecosystem processes and consequently on the people whose livelihoods depend on the services that ecosystems generate [64]. Continental reefs may be less resilient than isolated reefs due to proximity to anthropogenic stressors. Isolated reefs, however, may be more vulnerable to global change. Studies have shown that climate change-driven loss of live coral, and ultimately structural complexity, in the Seychelles resulted in local extinctions, substantial reductions in species richness, reduced taxonomic distinctness, and a loss of species within key functional groups of reef fish [98,99]. The importance of deteriorating physical structure to these patterns demonstrates the longer-term impacts of bleaching on reefs and raises questions over the potential for recovery. In the Red Sea, modelling of the dynamics of corals based on field data from Yossi Loya’s laboratory showed poor recruitment and low coral abundances [100]. The model demonstrated high fragility in the face of disturbances and predicted relatively high species extinctions with mild habitat destruction [100].
Human-induced change in water quality, herbivory levels, and climate change rate exert individual, discrete stressors on species of the critical functional groups on coral reefs (calcifying organisms, herbivores, primary producers other than calcifying organisms). A single stress by acting on key processes removes certain genotypes, alters demographic structure, spatial structure, and life history traits of populations, and leads to faster mean turnover times, and, thus, reduces simultaneously “environmental envelopes” of remaining genotypes (species) and buffering capacity [101]. Multiple stressors interacting with each other, either through synergy or competition, will aggravate the problem. There is growing scientific consensus that the local, human-induced, chronic stresses of degraded seawater quality (due to sediment and nutrient loads from terrestrial runoff and untreated sewage discharge) and reduced herbivory (due to overfishing or disease) erode the capacity of coral reefs to resist change (resistance) or successfully re-establish themselves after disturbance (resilience) [50,51,102].
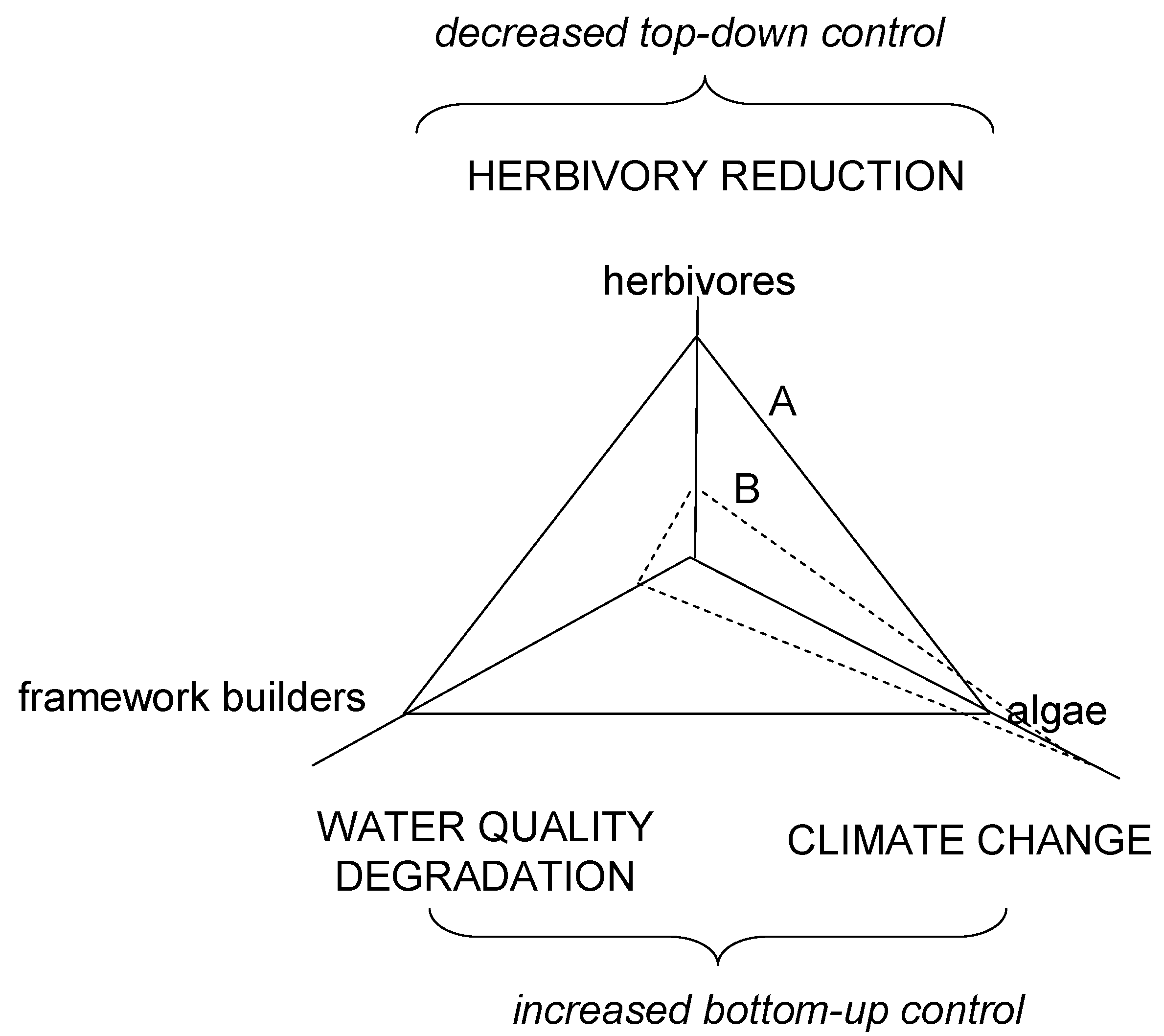
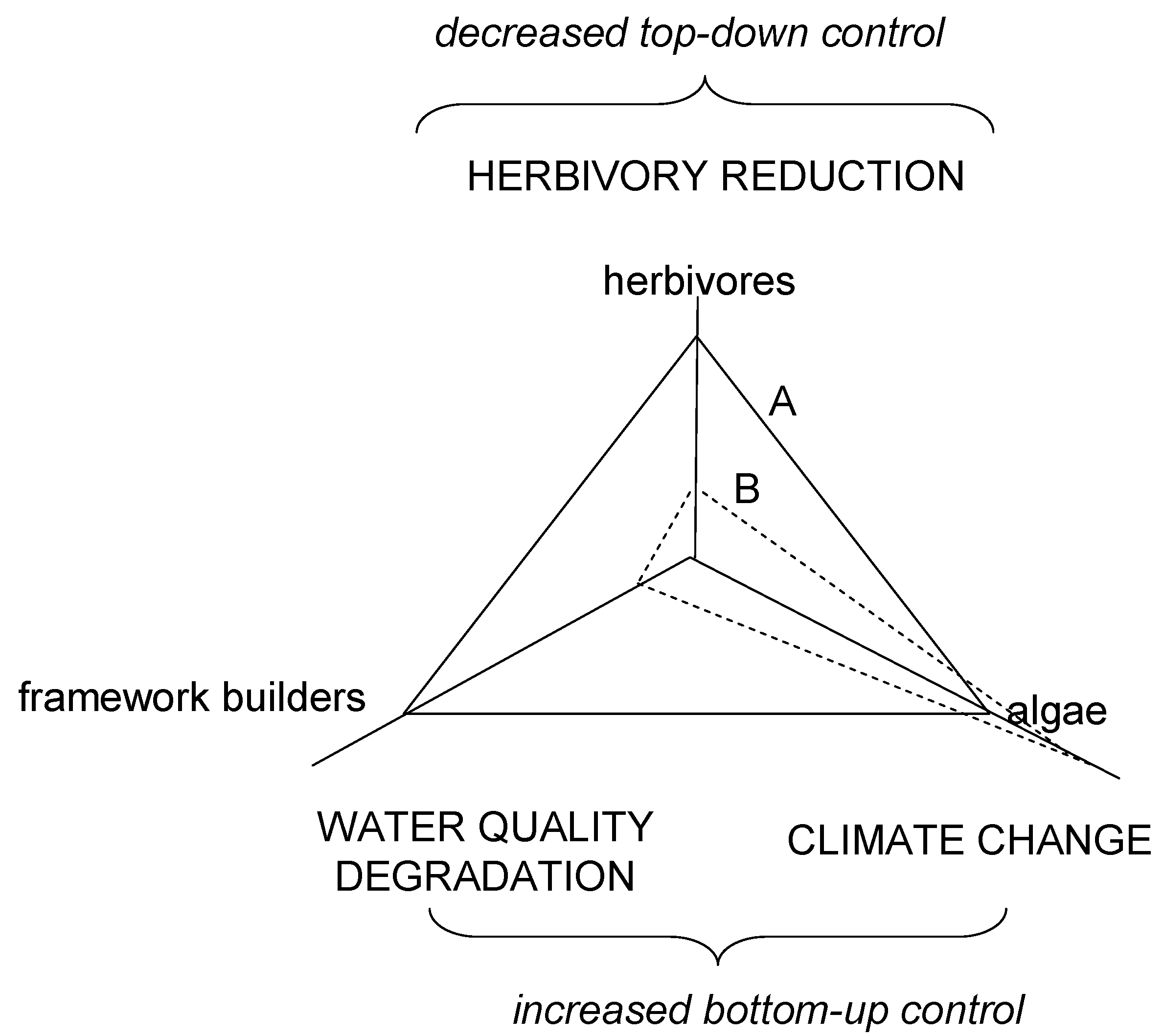
Climate forcing is postulated to exacerbate losses to a gene pool that is already reduced by effects of water quality degradation and herbivory reduction. The end result would be a reduced functional redundancy, response diversity, and ecosystem functioning or else a much simpler ecosystem with less distinguished functions. The result of homogenization would be algal beds on carbonate substrate resembling seagrasses on sandy bottoms. Furthermore, reduction in herbivory levels and water quality degradation may make fish and coral populations more strongly connected to climate variability at interannual to interdecadal scales. Increasing sensitivity implies an increasing correlation between fluctuations in population abundance (or another characteristic) and some climate signal [101]. The outcome of these interactions may be a change in the balance among the three functional groups and the persistence of algal dominance over large, formerly coral-dominated areas. Reefs that, before human interference, were controlled “top-down” by high levels of herbivory, and now widely held to be more strongly controlled from the “bottom-up” by degraded water quality and climate forcing (Scheme II).
Scheme II.
Effect of environmental drivers as selective forces on critical functional groups of coral reefs, namely framework builders (corals), algae and herbivores. Plane A, desired condition with balance attained among corals, algae and herbivores. Plane B, phase shift (to algal reefs) as the interaction of reduced top-down (over-fishing, disease) and increased bottom-up (degradation in water quality, climatic change) controls.
Scheme II.
Effect of environmental drivers as selective forces on critical functional groups of coral reefs, namely framework builders (corals), algae and herbivores. Plane A, desired condition with balance attained among corals, algae and herbivores. Plane B, phase shift (to algal reefs) as the interaction of reduced top-down (over-fishing, disease) and increased bottom-up (degradation in water quality, climatic change) controls.
Recent attention paid to global phenomena such as bleaching and coral disease, may be distracting attention from these long-term trends of reef degradation [45]. However, it is generally accepted that the coral–algal phase shift has been rapidly accelerating in recent decades, and this problem serves as an appropriate target for corrective coral reef management. Under these altered and increasingly unpredictable conditions, it may be necessary to re-evaluate the existing approaches and practices of how to analyze and anticipate the behaviour of coral reefs.
3. Coral Reef Management
The coral reef crisis attests to the ineffectiveness and inefficiency in the practice of contemporary management approaches, such as ecosystem-based management [103,104,105,106,107,108], integrated planning for environmental-economic-cultural disasters [109,110], and institutional change [111]. For example, a survey of the global network of Marine Protected Areas (MPAs) with coral reefs, which is considered a highly promising tool for marine ecosystem-based management [112] of, showed that whilst 18% of the world’s coral reefs are covered by MPAs, only ~2% is in areas of adequate conservation status, and less than 0.01% in areas of excellent status [113]. Stressors responsible for variations in macroalgae abundance (sewage and agriculture) and coral mortality (climate change) remained poorly, if at all, controlled inside MPAs throughout the Caribbean province [65].
Managing coral reef socio-ecological resilience has been proposed as an innovative way to manage the interaction between humanity and ecosystems [61,64,102,113,114,115,116] redirecting focus from monitoring the status of coral reefs to scrutinizing possibilities of how to better foresee ecosystem behavior in relation to socio-economic and environmental drivers that threaten coral reefs. It has been argued that combating the coral reef crisis should be based on a socio-ecological resilience paradigm as a basis for coping with uncertainty, future changes and ecological surprises in an increasingly human-dominated environment [64]. This outcome requires an improved understanding of the dynamics of key functional groups on coral reef ecosystems, of the processes that support or undermine resilience of key functional groups, of the socio-economic drivers and governance systems that regulate water quality and rates of extraction of reef resources; and a radical reassessment of the way reef resources are monitored and managed. Specific recommendations involve: (i) the rate of establishment and size of Non Take Areas (in regards to fishing), as a tool for resilience management, to be hugely increased to be sustainable within a seascape of degrading reefs and incorporate both functionally diverse and resilient “hot spots” and functionally depauperate and vulnerable “cool spots”, (ii) regional-scale active management of critical functional groups and habitat to support reef resilience, (iii) reef management be more inclusive, proactive and responsive, and (iv) markets for reef resources be reformed so as to reflect the ecological role of key functional groups on reefs [64].
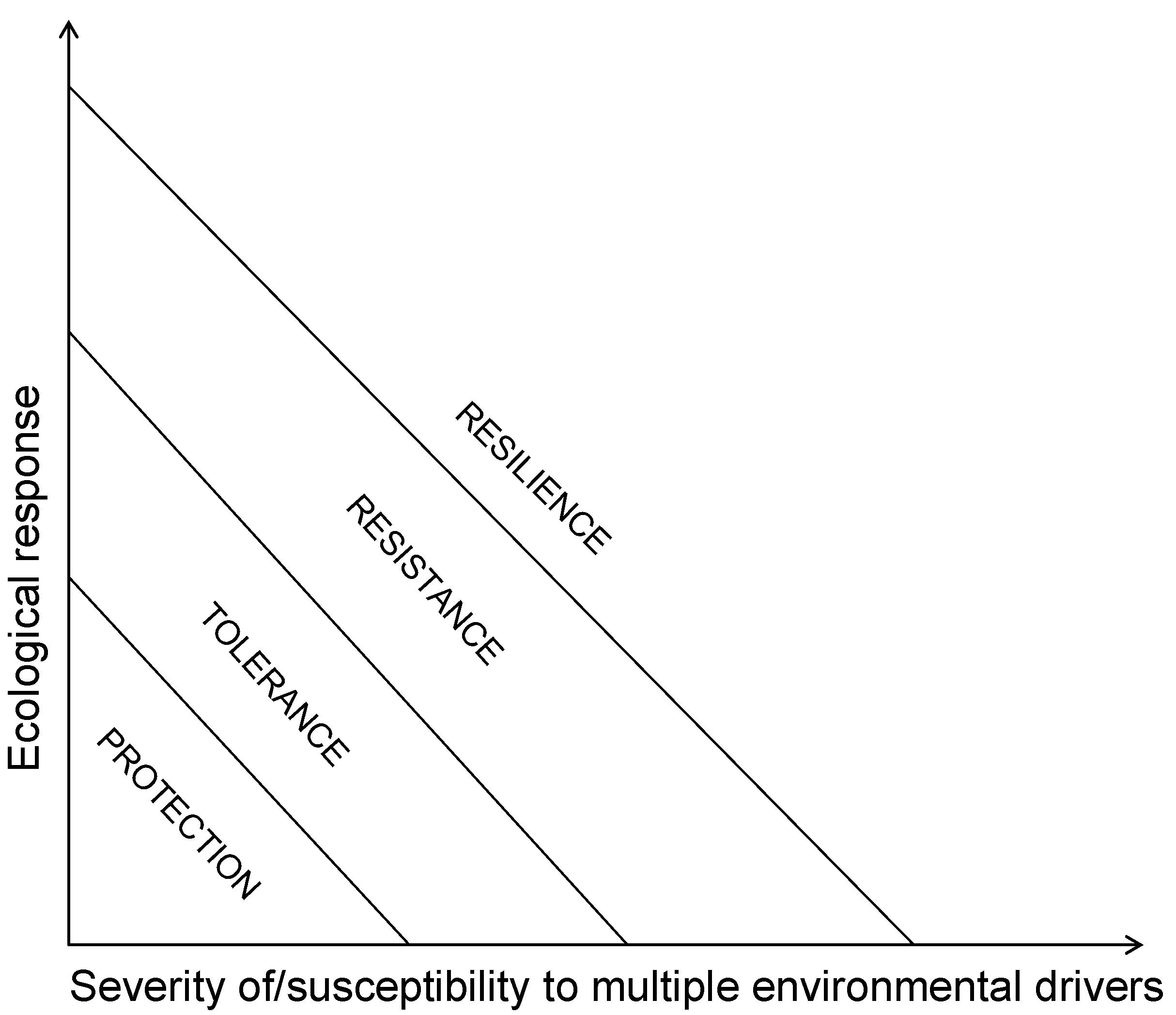
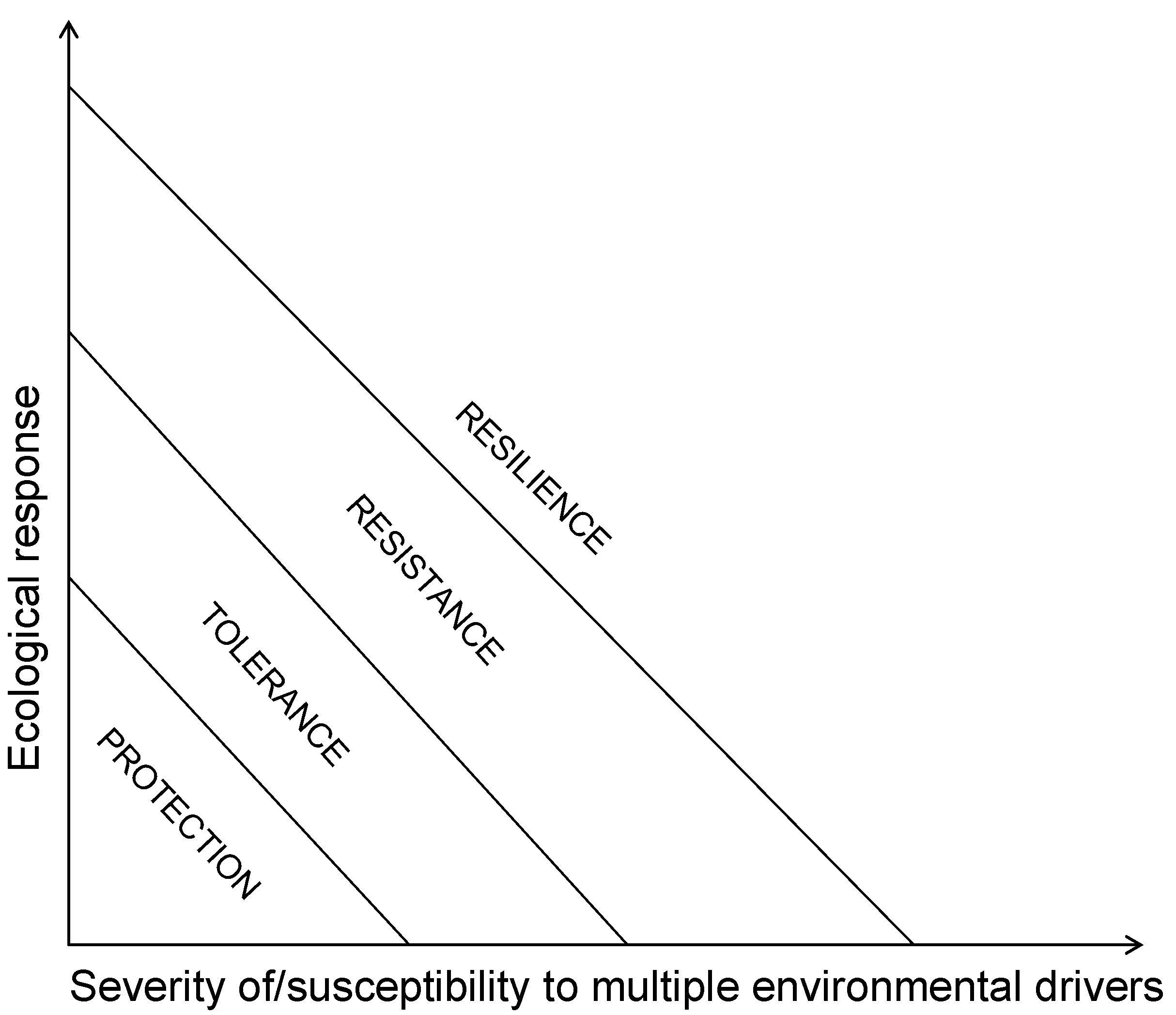
Marine resource managers are called to determine or/and direct the status of coral reef resources they manage across the gradient from protection, through tolerance, through resistance, to resilience towards a phase shift (Scheme III). Formal methods exist for the detection of a phase shift [117,118] while the relative abundance of the three key functional groups could help them position their reefs along the relative dominance paradigm [12]. Resilient areas may be identified through previous response to past climatic events; the ratio live/dead coral; the range of coral colony sizes; the fish age distribution; and the species diversity level [119]. Proposed, but yet untested, attributes of patterns and processes that lead to ecological resilience and ecosystem services reliability involve functional group approaches, ratios of “good” and “bad” colonizers of space, measurements of spatial heterogeneity, and estimates of potential space availability against grazing capacity, and increased variability [78]. Living coral cover is a key measure of reef habitat quality and quantity because corals feed and accomodate so many reef inhabitants [5,6]. Historic baseline coral cover, however, may have varied among subregions due to differences in disturbance frequency or the morphology of dominant species. For example, reefs dominated by plating acroporiid corals probably had higher baseline cover than reefs dominated by branching corals. Thus, similar current cover among subregions could actually reflect variability in the degree of coral loss. Additionally, the dependence of facilitation and other ecosystem functions on coral cover could vary among subregions (e.g., 20% cover might not be universally functionally equivalent) [49]. Thus, coral demographic approaches to monitoring “health” of reefs rather than live coral cover should be encouraged [121,122]. Also, modelling tools of the effects of fishing, water quality, and climate change on reef localities with a certain ratio of the three key functional groups may assist in obtaining trajectories of the coral reefs and adapt management as needed. Furthermore, ecosystem management should also be hierarchical and begin with the preservation of foundation species. Unfortunately, most marine conservation policies focus on the commercially harvested occupants of these habitats [49] utilising already failing optimization approaches based on Maximum Sustainable Yield [123]. Current aspirations towards sustaining fisheries need to fundamentally change their metrics from stock assessments to capturing the ecosystem performance and resilience that support long-term fisheries production [124]. MPA networks should be viewed not just as tools to halt further deterioration of corals reefs but as tools to reinforce resilience of coral reef ecosystems [113,125,126] (Scheme I). The long-term stability of coral reefs as a whole requires a holistic and regional approach to the control of human-related stressors in addition to the improvement and a three-fold increase in the establishment of new MPAs since failure to account for threats that affect corals and macroalgae may ultimately defeat the results of MPAs on fish populations [113] (Scheme I). In that respect, all countries whose socio-economic well-being depend on coral reefs have best interest to manage the totality of coral reefs under their jurisdiction as multiple zoned MPAs. The Large Marine Ecosystems, which are generally large regions of the order of 200,000 km2 or larger characterized by distinct bathymetry, hydrography, productivity and trophically dependent populations [126], may be the relevant scale upon which to design and implement networks of MPAs capable of protecting species, habitats, processes and functions of coral reefs [35,124,125,127].
Scheme III.
Coral reef management options determined by the dependence of ecological resilience on severity of/susceptibility to environmental drivers.
Scheme III.
Coral reef management options determined by the dependence of ecological resilience on severity of/susceptibility to environmental drivers.
A recent review of recommendations for terrestrial and marine biodiversity management through protected areas during the last 22 years [128] offers important assistance to marine resource managers: (i) More recommendations advocate resilience (e.g., enhance diversity at various scales, a need to view a broader range of ecosystem states as desirable, re-evaluate operational definitions and guidelines regarding invasive species and species at risk of extinction) [64,128,129] than resistance strategies (e.g., restoration, increased monitoring of species distribution, increased investment in reserve protection). Resistance and resilience strategies are not mutually exclusive, however. Very special communities or organisms, such as the critical coral species Acropora palmata and the echinoid grazer Diadema antillarum on Caribbean reefs, may warrant highly invasive, intense and costly management regimes to maintain them [130]; (ii) Climate-informed conservation planning necessitates site-specific understanding of environmental susceptibility and societal capacity to cope and adapt; this process has been illustrated for five western Indian Ocean countries with respect to coral reef conservation [131]. Locations with high environmental susceptibility and low adaptive capacity will be most difficult to secure effectively in the future, while those with low environmental susceptibility and high adaptive capacity will be easiest. Locations with low environmental susceptibility and low adaptive capacity are good candidates for biodiversity investment, but to be effective these locations also require investments in human infrastructure, livelihood diversification and social capital [131]; (iii) Forecast-interventions bear significant risks if they are too deterministic, not robust to alternative futures or have negative unanticipated consequences [132]. They could, however, deliver great rewards and should be weighed with sensitivity analyses and scenarios, tested in pilot programs, and implemented initially at small scales [133]. Scenario building—done in ways that is amenable to local data limitations and useable by policymakers and managers—is particularly apt for exploring the range of magnitudes and direction of possible futures and trends without commitment to specific forecasts [134,135]; (iv) Widespread calls exist for immediate action to adapt conservation practice to ongoing climate change in order to ensure the persistence of many species and related ecosystem services. However, the majority of recommendations in the published journal literature lack elaboration necessary to operationalize them. Thus, greater effort to increase the availability and applicability of climate change adaptation options for conservation—through concrete strategies and case studies illustrating how and where to link research agendas, conservation programs and institutions—is needed. Well-documented case studies that focus not only on the outcome but also on the development process of adaptation plans are a promising avenue [136]. These efforts can best enhance and encourage more widespread climate change adaptation, particularly at smaller scales, by capturing what they learn and disseminating it widely; (v) Holistic strategies, engaging social sciences and driven by a vision of humans and other species co-mingling across reserves and developed lands are required because communities of local users are often in conflict with conservation objectives [137,138] and because the role of multi-use public and private lands in conservation practice becomes increasingly important [139]; (vi) While a common framework to provide practitioners with structure, principles, concepts, tools and roles awaits yet to evolve, large-scale adaptation efforts that incorporate many of the recommendations made are currently underway, including governmental efforts, such as by Parks Canada or Department of Environment Food and Rural Affairs in England, and by international non-governmental organizations, such as The Nature Conservancy and the Wildlife Conservation Society [127].
While management options will vary among regions, there must be a common goal of reversing common trajectories of degradation for coral reefs [45]. Attending to the most manageable human impacts (fishing and water quality) could be valuable but in the long term may not be sufficient to reverse the coral reef decline unless the global-scale causes (climate change) of coral mortality are reversed at the highest levels of government and society [66,109,110]. As in other cases of resource degradation, such as water quality, the acceptable level of (anthropogenic) change lays within societies [140]. Presumably, the social acceptance of an alternative coral reef state will depend on whether the new state provides desired functions to societies. A bleached reef, however, may be a lovely, all white image to a novice tourist snorkeler on reefs but a tragic image to a coral reef ecologist.
References
- Moberg, F.; Folke, C. Ecological goods and services of coral reef ecosystems. Ecol. Econ. 1999, 29, 215–233. [Google Scholar] [CrossRef]
- Costanza, R.; d’Arge, R.; de Groot, R.; Farber, S.; Grasso, M.; Hannon, B.; Limburg, K.; Naeem, S.; O’Neill, R.V.; Paruelo, J.; Raskin, R.G.; Sutton, P.; van den Belt, M. The value of ecosystem services: putting the issues in perspective. Ecol. Econ. 1998, 25, 67–72. [Google Scholar] [CrossRef]
- Dubinsky, Z. Ecosystems of the World: 25 Coral Reefs; Elsevier Science: Amsterdam, The Netherlands, 1990; p. 550. [Google Scholar]
- Done, T.J.; Ogden, J.C.; Wiebe, W.J. Biodiversity and Ecosystem Function on Coral Reefs. In Bodiversity and Ecosystem Function: a Global Perspective; Mooney, H.A., Cushman, J.H., Medina, E., Sala, O.E., Schulze, E.-D., Eds.; John Wiley and Sons Ltd: New York, NY, USA, 1996; Chapter 15; p. 36. [Google Scholar]
- Bruno, J.F.; Bertness, M.D. Habitat modification and facilitation in Benthic Marine Communities. In Marine Community Ecology; Bertness, M.D., Hay, M.E., Gaines, S.D., Eds.; Sinauer: Sunderland, MA, USA, 2001; p. 17. [Google Scholar]
- Idjadi, J.A.; Edmunds, P.J. Scleractinian corals act as facilitators for other invertebrates on a Caribbean reef. Mar. Ecol. Progr. Ser. 2006, 319, 117–127. [Google Scholar] [CrossRef]
- Fabricius, K.; De’Ath, G. Environmental factors associated with the spatial distribution of crustose coralline algae on the Great Barrier Reef. Coral Reefs 2001, 19, 303–309. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Choat, J.H. A functional analysis of grazing in parrotfishes (family Scaridae): the ecological implications. Environ. Biol. Fish. 1990, 28, 189–214. [Google Scholar] [CrossRef]
- Steneck , R.S. Herbivory on coral reefs: a synthesis. In Vol. 1: Plenary Addressess and Status review, Proceedings of the 6th International Coral Reef Symposium; Choat, J.H., Barnes, D., Borowitzka, M.A., Coll, J.C., Davies, P.J., Flood, P., Hatcher, B.G., Hopley, D., Hutchings, P.A., Kinsey, D., Orme, G.R., Pichon, M., Sale, P.F., Sammarco, P., Wallace, C.C., Wilkinson, C., Wolanski, E., Bellwood, O., Eds.; Brian Clouston Publishing: Canberra, Australia, 1988; pp. 37–49. [Google Scholar]
- McCook, L.J.; Jompa, J.; Diaz-Pulido, G. Competition between corals and algae on coral reefs: a review of evidence and mechanisms. Coral Reefs 2001, 19, 400–417. [Google Scholar] [CrossRef]
- Cvitanovic, C.; Fox, R.J.; Bellwood, D.R. Herbivory by Fishes on the Great Barrier Reef: A Review of Knowledge and Understanding. Unpublished Report to the Marine and Tropical Sciences Research Facility. Reef and Rainforest Research Centre Limited: Cairns, Australia; 33.
- Littler, M.M.; Littler, D.S.; Brooks, B.L. Harmful algae on tropical coral reefs: Bottom-up eutrophication and top-down herbivory. Harmful Algae 2006, 5, 565–585. [Google Scholar] [CrossRef]
- Mumby, P.J.; Edwards, A.J.; Arias-González, J.E.; Lindeman, K.C.; Blackwell, P.G.; Gall, A.; Gorczynska, M.I.; Harborne, A.R.; Pescod, C.L.; Henk, R.; Wabnitz, C.C.C.; Llewellyn, G. Mangroves enhance the biomass of coral reef fish communities in the Caribbean. Nature 2004, 427, 533–536. [Google Scholar] [CrossRef]
- Mumby, P.J.; Hastings, A. The impact of ecosystem connectivity on coral reef resilience. J. Applied Ecol. 2008, 45, 854–862. [Google Scholar] [CrossRef]
- Hughes, T.P.; Connell, J.H. Multiple stressors on coral reefs: A long-term perspective. Limnol. Oceanogr. 1999, 44, 932–940. [Google Scholar] [CrossRef]
- Goreau, T.F.; Goreau, N.I.; Goreau, T.J. Corals and Coral Reefs. Sci. Amer. 1979, 241, 124–136. [Google Scholar] [CrossRef]
- Comparison between Atlantic and Pacific Tropical Marine Coastal Ecosystems: Community Structure, Ecological Processes, and Productivity. Results and Scientific Papers of a Unesco/COMAR Workshop (Suva, Fiji, 24–29 March, 1986); Birkeland, C (Ed.) Unesco Reports in Marine Science 46; UNESCO: Suva, Fiji, 1987.
- Buddemeier, R.W.; Fautin, D.G. Large-scale dynamics: the state of the science, the state of the reef, and the research issues. Coral Reefs 2002, 21, 1–8. [Google Scholar] [CrossRef]
- Johnson, K.G.; Budd, A.F.; Steemann, T.A. Extinction selectivity and ecology of Neogene Caribbean reef corals. Paleobiology 1995, 21, 52–73. [Google Scholar]
- Bellwood, D.R.; Wainwright, P.C. The history and biogeography of fishes on coral reefs. In Coral Reef Fishes. In Coral Reef Fishes: Dynamics and Diversity in a Complex Ecosystem; Sale, P.F., Ed.; Academic Press: San Diego, CA, USA, 2002; p. 27. [Google Scholar]
- Done, T.J. Constancy and change in some Great Barrier Reef coral communities: 1980–1990. American Zoologist 1992, 32, 655–662. [Google Scholar]
- Goreau, T.F.; Goreau, T.F. The ecology of Jamaican coral reefs. 1. Species composition and zonation. Ecology 1959, 40, 67–89. [Google Scholar] [CrossRef]
- Loya, Y. The Red Sea coral Stylophora pistillata is an r strategist. Nature 1976, 259, 478–480. [Google Scholar] [CrossRef]
- Loya, Y. Community structure and species diversity of hermatypic corals at Eilat, Red Sea. Mar. Biol. 1972, 23, 100–123. [Google Scholar] [CrossRef]
- Sammarco, P.W. A Comparison of Some Ecological Processes on Coral Reefs of the Caribbean and the Great Barrier Reef; Unesco Reports in Marine Science 46; Unesco: Paris, France, 1987; pp. 148–187. [Google Scholar]
- Jackson, J.B.C.; Kirby, M.X.; Berger, W.H.; Bjorndal, K.A.; Botsford, L.W.; Bourque, B.J.; Bradbury, R.H.; Cooke, R.; Erlandson, J.; Estes, J.A.; Hughes, T.P.; Kidwell, S.; Lange, C.B.; Lenihan, H.S.; Pandolfi, J.M.; Peterson, C.H.; Steneck, R.S.; Tegner, M.J.; Warner, R.R. Historical overfishing and the recent collapse of coastal ecosystems. Science 2001, 293, 629–638. [Google Scholar] [CrossRef]
- Hughes, T.P. Catastrophes, phase-shifts, and largescale degradation of a Caribbean coral reef. Science 1994, 265, 1547–1551. [Google Scholar]
- Glassom, D.; Zakai, D; Chadwick-Furman, N.E. Coral recruitment: a spatio-temporal analysis along the coastline of Eilat, northern Red Sea. Mar. Biol. 2004, 144, 641–651. [Google Scholar] [CrossRef]
- Pimm, S.L. The complexity and stability of ecosystems. Nature 1984, 307, 321–327. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in Tropical Rain Forests and Coral Reefs. Science 1978, 199, 1302–1310. [Google Scholar]
- Pearson, R.G. Recovery and decolonisation of coral reefs. Mar. Ecol. Progr. Ser. 1981, 4, 105–122. [Google Scholar] [CrossRef]
- Bythell, J.C.; Hillis-Starr, Z.M.; Rogers, C.S. Local variability but landscape stability in coral reef communities following repeated hurricane impacts. Mar. Ecol. Progr. Ser. 2000, 204, 93–100. [Google Scholar] [CrossRef]
- Connell, J.H. Disturbance and recovery of coral assemblages. Coral Reefs 1997, 16S, 101–113. [Google Scholar] [CrossRef]
- Kaufmann, A.; Fagerstrom, J.A. The Phanero- zoic evolution of reef diversity. In Species Diversity in Ecological Communities: A Historical and Geographical Perspective; Ricklefs, R.E., Schulter, D., Eds.; University of Chicago Press: Chicago, IL, USA, 1993; pp. 315–329. [Google Scholar]
- Wood, R. The changing biology of reef-building. Palaios 1995, 10, 517–529. [Google Scholar] [CrossRef]
- Roberts, C.M. Connectivity and Management of Caribbean Coral Reefs. Science 1997, 278, 1454–1457. [Google Scholar] [CrossRef]
- Palumbi, S.R. Marine speciation on a small planet. Trends Ecol. Evol. 1992, 7, 114–118. [Google Scholar] [CrossRef]
- Benzie, J.A.H. Genetic structure of coral reef organisms: Ghosts of dispersal past. Am. Zool. 1999, 39, 131–145. [Google Scholar]
- Pandolfi, J.M. Coral reef ecology at multiple spatial and temporal scales. Coral Reefs 2002, 21, 13–23. [Google Scholar] [CrossRef]
- Chappell, J. Coral morphology, diversity and reef growth. Nature 1980, 286, 249–252. [Google Scholar] [CrossRef]
- Jackson, J.B.C. Community unity? Science 1994, 264, 1412–1413. [Google Scholar]
- Hughes, T.P.; Baird, A.H.; Dinsdale, E.A.; Moltschaniwskyj, N.A.; Pratchett, M.S.; Tanner, J.E.; Willis, B.L. Patterns of recruitment and abundance of corals along the Great Barrier Reef. Nature 1999, 397, 59–63. [Google Scholar] [CrossRef]
- Van Woesik, R. Processes regulating coral communities. Comment. Theor. Biol. 2002, 7, 201–214. [Google Scholar] [CrossRef]
- Done, T.J. Perspectives on Coral Reefs; Barnes, D.J., Ed.; Australian Institute of Marine Science: Cairns, Australia, 1983; pp. 95–147. [Google Scholar]
- Pandolfi, J.M.; Bradbury, R.H.; Sala, E.; Hughes, T.P.; Bjorndal, K.A.; Cooke, R.G.; McArdle, D.; McClenachan, L.; Newman, M.J.H.; Paredes, G.; Warner, R.R.; Jackson, J.B.C. Global trajectories of the long-term decline of coral reef ecosystems. Science 2003, 301, 955–958. [Google Scholar] [CrossRef]
- The State of Coral Reef Ecosystems of the United States and Pacific Freely Associated States; NOAA Technical Memorandum NOSNCCOS73; NOAA/NCCOS Center for Coastal Monitoring and Assessment’s Biogeography Team: Silver Spring, MD, USA, 2008.
- Carpenter, K.E.; Abrar, M.; Aeby, G.; Aronson, R.B.; Banks, S.; Bruckner, A.; Chiriboga, A.; Cortés, J.; Delbeek, J.C.; DeVantier, L.; Edgar, G.J.; Edwards, A.J.; Fenner, D.; Guzmán, H.M.; Hoeksema, B.W.; Hodgson, G.; Johan, O.; Licuanan, W.Y.; Livingstone, S.R.; Lovell, E.R.; Moore, J.A.; Obura, D.O.; Ochavillo, D.; Polidoro, B.A.; Precht, W.F.; Quibilan, M.C.; Reboton, C.; Richards, Z.T.; Rogers, A.D.; Sanciangco, J.; Sheppard, A.; Sheppard, C.; Smith, J.; Stuart, S.; Turak, E.; Veron, J.E.N.; Wallace, C.; Weil, E.; Wood, E. One-Third of Reef-Building Corals Face Elevated Extinction Risk from Climate Change and Local Impacts. Science 2008, 321, 560–563. [Google Scholar] [CrossRef]
- Gardner, T.A.; Côté, I.M.; Gill, J.A.; Grant, A.; Watkinson, A.R. Long-Term Region-Wide Declines in Caribbean Corals. Science 2003, 301, 958–960. [Google Scholar] [CrossRef]
- Bruno, J.F.; Selig, E.R. Regional Decline of Coral Cover in the Indo-Pacific: Timing, Extent, and Subregional Comparison. PLoS ONE 2007, 8, 1–8. [Google Scholar]
- Hughes, T.P.; Baird, A.H.; Bellwood, D.R.; Card, M.; Connolly, S.R.; Folke, C.; Grosberg, R.; Hoegh-Guldberg, O.; Jackson, J.B.C.; Kleypas, J.; Lough, J.M.; Marshall, P; Nyström, M.; Palumbi, S.R.; Pandolfi, J.M.; Rosen, B.; Roughgarden, J. Climate Change, Human Impacts, and the Resilience of Coral Reefs. Science 2003, 301, 929–933. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O.; Mumby, P.J.; Hooten, A.J.; Steneck, R.S.; Greenfield, P.; Gomez, E.; Harvell, C.D.; Sale, P.F.; Edwards, A.J.; Caldeira, K.; Knowlton, N.; Eakin, C.M.; Iglesias-Prieto, R.; Muthiga, N.; Bradbury, R.H.; Dubi, A.; Hatziolos, M.E. Coral reefs under rapid climate change and ocean acidification. Science 2007, 318, 1737–1742. [Google Scholar] [CrossRef]
- McManus, J.W.; Polsenberg, J.F. Coral-algal phase shifts on coral reefs: ecological and environmental aspects. Prog. Oceanogr. 2004, 60, 263–279. [Google Scholar] [CrossRef]
- Done, T.J. Phase shifts in coral reef communities and their ecological significance. Hydrobiologia 1992, 247, 121–132. [Google Scholar] [CrossRef]
- Ogden, J.C.; Brown, R.A.; Salesky, N. Grazing by the echinoid Diadema antillarum Philippi: Formation of halos around West Indian patch reefs. Science 1973, 182, 715–717. [Google Scholar]
- Hughes, T.P. Catastrophes, phase-shifts, and large-scale degradation of a Caribbean coral reef. Science 1994, 265, 1547–1551. [Google Scholar]
- McClanahan, T.R.; Shafir, S.H. Causes and consequences of sea urchin abundance and diversity in Kenyan coral reef lagoons. Oecologia 1990, 83, 362–370. [Google Scholar]
- Norström, A.V.; Nyström, M.; Lokrantz, J.; Folke, C. Alternative states on coral reefs: beyond coral-macroalgal phase shifts. Mar. Ecol. Progr. Ser. 2009, 376, 295–306. [Google Scholar] [CrossRef]
- Mumby, P.J.; Steneck, R.S. Coral reef management and conservation in light of rapidly evolving ecological paradigms. Trends Ecol. Evol. 2008, 23, 555–563. [Google Scholar] [CrossRef]
- Mumby, P. Phase shifts and the stability of macroalgal communities on Caribbean coral reefs. Coral Reefs 2009, 28, 761–773. [Google Scholar] [CrossRef]
- Knowlton, N. Thresholds and multiple stable states in coral reef community dynamics. Am. Zool. 1992, 32, 674–682. [Google Scholar]
- Scheffer, M.; Carpenter, S.; Foley, J.A.; Folke, C.; Walker, B. Catastrophic shifts in ecosystems. Nature 2001, 413, 591–596. [Google Scholar] [CrossRef]
- Steel, J.H. Regime shifts in the ocean: reconciling observations and theory. Progr. Oceanogr. 2004, 60, 135–141. [Google Scholar] [CrossRef]
- Cesar, H.; Burke, L.; Pet-Soede, L. The Economics of World-Wide Coral Reef Degradation; Cesar Environmental Economics Consulting (CEEC): Arnhem, The Netherlands, 2003. [Google Scholar]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef]
- Mora, C. A clear human footprint in the coral reefs of the Caribbean. P. Roy. Soc. B-Biol. Sci. 2008, 275, 767–773. [Google Scholar] [CrossRef]
- Aronson, R.B.; Precht, W.F. Conservation, precaution, and Caribbean reefs. Coral Reefs 2006, 25, 441–450. [Google Scholar] [CrossRef]
- Edmunds, P.J.; Carpenter, R.C. Recovery of Diadema antillarum reduces macroalgal cover andincreases abundance of juvenile corals on a Caribbean reef. Proc. Natl. Acad. Sci. USA 2001, 98, 5067–5073. [Google Scholar] [CrossRef] [Green Version]
- Done, T.J.; Turak, E.; Wakeford, M,; DeVantier, L.; McDonald, A.; Fisk, D. Decadal changes in turbid-water coral communities at Pandora Reef: loss of resilience or too soon to tell? Coral Reefs 2007, 26, 789–805. [Google Scholar] [CrossRef]
- Carpenter, R.C.; Edmunds, P.J. Local and regional scale recovery of Diadema promotes recruitment of scleractinian corals. Ecol. Lett. 2006, 9, 268–277. [Google Scholar]
- Aronson, R.B.; Macintyre, I.G.; Precht, W.F.; Murdoch, T.J.T.; Wapnicki, C.M. The expanding scale of species turnover events on coral reefs in Belize. Ecol. Monogr. 2002, 72, 233–249. [Google Scholar] [CrossRef]
- Knowlton, N. Sea urchin recovery from mass mortality: New hope for Caribbean coral reefs? Proc. Natl. Acad. Sci. USA 2001, 98, 4822–4824. [Google Scholar] [CrossRef]
- Van Nes, E.H.; Scheffer, M. Large species shifts triggered by small forces. Am. Nat. 2004, 164, 255–266. [Google Scholar] [CrossRef]
- Van Nes, E.H.; Scheffer, M. Slow recovery from perturbations as a generic indicator of a nearby catastrophic shift. Am. Nat. 2007, 170, 660–660. [Google Scholar] [CrossRef]
- McClanahan, T.R.; Mutere, J.C. Coral and sea-urchin assemblage structure and interrelationships in Kenyan reef lagoons. Hydrobiologia 1994, 286, 109–124. [Google Scholar] [CrossRef]
- Eakin, C.M. Where have all the carbonates gone? A model comparison of calcium carbonate budgets before and after the 1982–1983 El Niño at Uva Island in the eastern Pacific. Coral Reefs 1996, 15, 109–119. [Google Scholar]
- Hunter, C.L.; Evans, C.W. Coral reefs in Kaneohe Bay, Hawaii—two centuries of western influence and two decades of data. Bull. Mar. Sci. 1995, 57, 501–515. [Google Scholar]
- Idjadi, J.A.; Lee, S.C.; Bruno, J.F.; Precht, W.F.; Allen-Requa, L.; Edmunds, P.J. Rapid phase-shift reversal on a Jamaican coral reef. Coral Reefs 2006, 25, 209–211. [Google Scholar] [CrossRef]
- Nyström, M.; Graham, N.A.J.; Lokrantz, J.; Norström, A.V. Capturing the cornerstones of coral reef resilience: linking theory to practice. Coral Reefs 2008, 27, 795–809. [Google Scholar] [CrossRef]
- Knowlton, N.; Jackson, J.B.C. Shifting Baselines, Local Impacts, and Global Change on Coral Reefs. PLOS Biol. 2008, 6, 215–220. [Google Scholar] [CrossRef]
- Fabricius, K.E. Effects of terrestrial runoff on the ecology of corals and coral reefs: Review and synthesis. Mar. Pollut. Bull. 2005, 50, 125–146. [Google Scholar] [CrossRef]
- Bruno, J.F.; Petes, L.E.; Harvell, C.D.; Hettinger, A. Nutrient enrichment can increase the severity of coral diseases. Ecol. Lett. 2003, 6, 1056–1061. [Google Scholar] [CrossRef]
- Smith, J.E.; Shaw, M.; Edwards, R.A.; Obura, D.; Pantos, O.; Sala, E.; Sandin, S.A.; Smriga, S.; Hatay, M.; Rohwer, F.L. Indirect effects of algae on coral: algae-mediated, microbe-induced coral mortality. Ecol. Lett. 2006, 9, 835–845. [Google Scholar] [CrossRef]
- Szmant, A.M. Editorial Introduction to the special issue of Coral Reefs on “Coral Reef Algal Community Dynamics”. Why are coral reefs world-wide overgrown by algae? “Algae, algae everywhere, and nowhere a bit to eat!”. Coral Reefs 2001, 19, 299–302. [Google Scholar] [CrossRef]
- Burkepile, D.E.; Hay, M.E. Impact of Herbivore Identity on Algal Succession and Coral Growth on a Caribbean Reef. PLoS ONE 2010, 5, 1–9. [Google Scholar]
- Mumby, P.J.; Harborne, A.R.; Williams, J.; Kappel, C.V.; Brumbaugh, D.R.; Micheli, F.; Holmes, K.E.; Dahlgren, C.P.; Paris, C.B.; Blackwell, P.G. Trophic cascade facilitates coral recruitment in a marine reserve. Proc. Natl. Acad. Sci. USA 2007, 104, 8362–8367. [Google Scholar] [CrossRef]
- Kline, D.I.; Kuntz, N.M.; Breitbart, M.; Knowlton, N.; Rohwer, F. Role of elevated organic carbon levels and microbial activity in coral mortality. Mar. Ecol. Progr. Ser. 2006, 314, 119–125. [Google Scholar] [CrossRef]
- Nugues, M.M.; Smith, G.W.; van Hooidonk, R.J.; Seabra, M.I.; Bak, R.P.M. Algal contact as a trigger for coral disease. Ecol. Lett. 2004, 7, 919–923. [Google Scholar] [CrossRef]
- Donner, S.D.; Skirving, W.J.; Little, C.M.; Oppenheimer, M.; Hoegh-Guldberg, O. Global assessment of coral bleaching and required rates of adaptation under climate change. Global Change Biol. 2005, 11, 2251–2265. [Google Scholar] [CrossRef]
- Coles, S.L.; Brown, B.E. Coral Bleaching—Capacity for Acclimatization and Adaptation. Adv. Mar. Biol. 2003, 46, 183–223. [Google Scholar] [CrossRef]
- Baker, A.C.; Starger, C.J.; McClanahan, T.R.; Glynn, P.W. Corals’ adaptive response to climate change. Nature 2004, 430, 741. [Google Scholar] [CrossRef]
- Marshall, P.M.; Schuttenberg, H.Z. A Reef Manager’s Guide to Coral Bleaching; Great Barrier Reef Marine Park Authority: Townsville, Australia, 2006; p. 165. [Google Scholar]
- Fabricius, K.E. Theme section on “Ocean Acidification and Coral Reefs”. Coral Reefs 2008, 27, 455–457. [Google Scholar] [CrossRef]
- Randall, C.J.; Szmant, A.M. Elevated temperature reduces survivorship and settlement of the larvae of the Caribbean scleractinian coral, Favia fragum (Esper). Coral Reefs 2009, 28, 537–545. [Google Scholar] [CrossRef]
- Pörtner, H.O.; Langenbuch, M.; Michaelidis, B. Synergistic effects of temperature extremes, hypoxia, and increases in CO2 on marine animals: From Earth history to global change. J. Geophys. Res. 2005, 110, C09–S10. [Google Scholar]
- Bruno, J.F.; Selig, E.R.; Casey, K.S.; Page, C.A.; Willis, B.L.; Harvell, D.C.; Sweatman, H.; Melendy, A.M. Thermal Stress and Coral Cover as Drivers of Coral Disease Outbreaks. PLOS Biol. 2007, 5, 1–8. [Google Scholar] [CrossRef]
- Jones, G.P.; McCormick, M.I.; Srinivasan, M.; Eagle, J.V. Coral decline threatens fish biodiversity in marine reserves. Proc. Natl. Acad. Sci. USA 2004, 101, 8251–8253. [Google Scholar] [CrossRef]
- Kareiva, P.; Marvier, M. Conserving biodiversity coldspots. Am. Sci. 2003, 91, 344–351. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Wilson, S.K.; Jennings, S.; Polunin, N.V.C.; Robinson, J.; Bijoux, J.P.; Daw, T.M. Lag effects in the impacts of mass coral bleaching on coral reef fish, fisheries, and ecosystems. Conserv. Biol. 2007, 21, 1291–1300. [Google Scholar] [CrossRef] [Green Version]
- Hughes, T.P.; Rodrigues, M.J.; Bellwood, D.R.; Ceccarelli, D.; Hoegh-Guldberg, O.; McCook, L.; Moltschaniwskyj, N.; Pratchett, M.S.; Steneck, R.S.; Willis, B. Phase shifts, herbivory, and the resilience of coral reefs to climate change. Curr. Biol. 2007, 17, 360–365. [Google Scholar]
- Stone, L.; Eilam, E.; Abelson, A.; Llano, M. Modelling cora1 reef biodiversity and habitat destruction. Mar. Ecol. Progr. Ser. 1996, 134, 299–302. [Google Scholar] [CrossRef]
- Perry, R.I.; Philippe, C.; Brander, K.; Jennings, S.; Möllmann, C.; Planque, B. Sensitivity of marine systems to climate and fishing: Concepts, issues and management responses. J. Mar. Systems 2010, 79, 427–435. [Google Scholar] [CrossRef]
- Hughes, T.P.; Bellwood, D.R.; Folke, C.; Steneck, R.S.; Wilson, J. New paradigms for supporting the resilience of marine ecosystems. Trends Ecol. Evol. 2005, 20, 380–386. [Google Scholar] [CrossRef]
- Allison, G.W.; Gaines, S.D.; Lubchenco, J.; Possingham, H.P. Ensuring persistence of marine reserves: catastrophes require adopting an insurance factor. Ecol. Appl. 2003, 13, S8–S24. [Google Scholar] [CrossRef]
- Roberts, C.M.; Andelman, S.; Branch, G.; Bustamante, R.H.; Castilla, J.C.; Dugan, J.; Halpern, B.S.; Lafferty, K.S.; Leslie, H.; Lubchenco, J.; McArdle, D.; Possingham, H.P.; Ruckleshaus, M.; Warner, R.R. Ecological criteria for evaluating candidate sites for marine reserves. Ecol. Appl. 2003, 13, S199–S214. [Google Scholar] [CrossRef]
- Pikitch, E.K.; Santora, C.; Babcock, E.A.; Bakun, A.; Bonfil, R.; Conover, D.O.; Dayton, P.; Doukakis, P.; Fluharty, D.; Heneman, B.; Houde, E.D.; Link, J.; Livingston, P.A.; Mangel, M.; McAllister, M.K.; Pope, J.; Sainsbury, K.J. Ecosystem-based fishery management. Science 2004, 305, 346–347. [Google Scholar] [CrossRef]
- Guerry, A.D. Icarus and Daedalus: conceptual and tactical lessons for marine ecosystem-based management. Front. Ecol. Environ. 2005, 3, 202–211. [Google Scholar] [CrossRef]
- Fernandes, L.; Day, J.; Lewis, A.; Slegers, S.; Kerrigan, B.; Breen, D.; Cameron, D.; Jago, B.; Hall, J.; Lowe, D.; Innes, J.; Tanzer, J.; Chadwick, V.; Thompson, L.; Gorman, K.; Simmons, M.; Barnett, B.; Sampson, K.; De’Ath, G.; Mapstone, B.; Marsh, H.; Possingham, H.; Ball, I.; Ward, T.; Dobbs, K.; Aumend, J.; Slater, D.; Stapleton, K. Establishing representative no-take areas in the Great Barrier Reef: large-scale implementation of theory on marine protected areas. Conserv. Biol. 2005, 19, 1733–1744. [Google Scholar] [CrossRef]
- McClanahan, T.R.; Verheij, E.; Maina, J. Comparing the managementeffectiveness of a marine park and a multiple-use collaborative fisheries management area in East Africa. Aquat. Conserv. 2006, 16, 147–165. [Google Scholar] [CrossRef]
- McClanahan, T.R.; Polunin, N.V.C.; Done, T.J. Ecological States and the Resilience of Coral Reefs. In Resilience and the Behaviour of Large-Scale Systems; Gunderson, L.H., Pritchard, L., Jr., Eds.; Island Press: Washington, DC, USA, 2002; pp. 111–163. [Google Scholar]
- Ehrlich, P.R.; Kennedy, D. Millennium assessment of human behaviour. Science 2005, 309, 562–563. [Google Scholar] [CrossRef]
- Carney, D.; Farrington, J. Natural Resource Management and Institutional Change; Routledge: New York, NY, USA, 1998; p. 132. [Google Scholar]
- Scientific Consensus Statement on Marine Reserves and Marine Protected Areas; American Association for the Advancement of Science (AAAS): Santa Barbara, CA, USA, 2001.
- Mora, C.; Andréfouët, S.; Costello, M.J.; Kranenburg, C.; Rollo, A.; Veron, J.; Gaston, K.J.; Myers, R.A. Coral Reefs and the Global Network of Marine Protected Areas. Science 2006, 312, 1750–1751. [Google Scholar] [CrossRef]
- Nyström, M.; Folke, C.; Moberg, F. Coral reef disturbance and resilience in a human-dominated environment. Trends Ecol. Evol. 2000, 15, 413–417. [Google Scholar] [CrossRef]
- Nyström, M.; Folke, C. Spatial resilience of coral reefs. Ecosystems 2001, 4, 406–417. [Google Scholar] [CrossRef]
- Adger, N.; Hughes, T.P.; Folke, C.; Carpenter, S.R.; Rockström, J. Social-ecological resilience to coastal disasters. Science 2005, 309, 1036–1039. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Folke, C. Ecology for transformation. Trends Ecol. Evol. 2006, 21, 309–315. [Google Scholar] [CrossRef]
- Andersen, T.; Carstensen, J.; Hernández-García, E.; Duarte, C.M. Ecological thresholds and regime shifts: approaches to identification. Trends Ecol. Evol. 2008, 24, 49–57. [Google Scholar]
- deYoung, B.; Barange, M.; Beaugrand, G.; Harris, R.; Perry, R.I.; Scheffer, M.; Werner, F. Regime shifts in marine ecosystems: detection, prediction and management. Trends Ecol. Evol. 2008, 23, 402–409. [Google Scholar] [CrossRef]
- Schuttenberg, H.Z.; Corrigan, C.; McLeod, L.; Marshall, P.; Setiasih, N.; Obura, D.; Hoegh-Guldberg, O.; Causey, B.; Drew, M.; Hansen, L.; Grimsditch, G.; West, J.; Skeat, A.; Eakin, M.; McCook, L.; Crawford, M.; Kramer, P.; Campbell, S. Building resilience into coral reef management: Key findings & recommendations. In Global Problems Local Solutions, Proceedings of the 3rd International Tropical Marine Ecosystems Management Symposium, Cozumel, Mexico, 16–20 October 2006; ICRI and ICRAN: Cambridge, UK, 2006; p. 8. [Google Scholar]
- Smith, L.D.; Devlin, M.; Haynes, D.; Gilmour, J.P. A demographic approach to monitoring the health of coral reefs. Mar. Pollut. Bull. 2004, 51, 399–407. [Google Scholar]
- Done, T.J.; DeVantier, L.M.; Turak, E.; Fisk, D.A.; Wakeford, M.; van Woesik, R. Coral growth on three reefs: development of recovery benchmarks using a space for time approach. Coral Reefs 2010, (in press). [Google Scholar]
- Roberts, C.M. Ecological advice for the global fisheries crisis. Trends Ecol. Evol. 1997, 12, 35–38. [Google Scholar] [CrossRef]
- Pauly, D.; Christensen, V.; Guénette, S.; Pitcher, T.J.; Sumaila, U.R.; Walters, C.J.; Watson, R.; Zeller, D. Towards sustainability in world fisheries. Nature 2002, 418, 689–695. [Google Scholar] [CrossRef]
- Harris, J.A.; Hobbs, R.J.; Higgs, E.; Aronson, J. Ecological restoration and global climate change. Restor. Ecol. 2006, 14, 170–176. [Google Scholar] [CrossRef]
- Sherman, K.; Duda, A.M. Large marine ecosystems: an emerging paradigm for fishery sustainability. Fisheries 1999, 24, 15–26. [Google Scholar] [CrossRef]
- Establishing Marine Protected Area Networks—Making It Happen; IUCN-WCPA, National Oceanic and Atmospheric Administration and The Nature Conservancy: Washington, DC, USA, 2008; p. 118.
- Heller, N.E.; Zavaleta, E.S. Biodiversity management in the face of climate change: A review of 22 years of recommendations. Biol. Conserv. 2009, 142, 14–32. [Google Scholar] [CrossRef]
- Hulme, P.E. Adapting to climate change: is there scope for ecological management in the face of a global threat? J. Applied Ecol. 2005, 42, 784–794. [Google Scholar] [CrossRef]
- Threatened Elkhorn and Staghorn Corals (Acropora sp.); NOAA Fisheries: Silver Spring, MD, USA, November 25 2009.
- McClanahan, T.R.; Cinner, J.E.; Maina, J.; Graham, N.A.; Daw, T.M.; Stead, S.M.; Wamukota, A.; Brown, K.; Ateweberhan, M.; Venus, V.; Polunin, N.V.C. Conservation action in a changing climate. Conserv. Lett. 2008, 1, 53–59. [Google Scholar] [Green Version]
- Suffling, R.; Scott, D. Assessment of climate change effects on Canada’s national park system. Environ. Monit. Assess. 2002, 74, 117–139. [Google Scholar] [CrossRef]
- McLachlan, J.; Hellmann, J.; Schwartz, M. A framework for debate of assisted migration in an era of climate change. Conserv. Biol. 2007, 21, 297–302. [Google Scholar] [CrossRef]
- Brown, I. Modelling future landscape change on coastalfloodplains using a rule-based GIS. Environ. Modell. Softw. 2006, 21, 1479–1490. [Google Scholar] [CrossRef]
- Millar, C.I.; Stephenson, N.L.; Stephens, S.L. Climate changeand forests of the future: managing in the face of uncertainty. Ecol. Appl. 2007, 17, 2145–2151. [Google Scholar] [CrossRef]
- McLeod, L. Integrating Resilience Into MPA Design. In Global Problems Local Solutions, Proceedings of the 3rd International Tropical Marine Ecosystems Management Symposium, Cozumel, Mexico, 16–20 October 2006; ICRI and ICRAN: Cambridge, UK, 2006; p. 8. [Google Scholar]
- Camargo, C.; Maldonado, J.H.; Alvarado, E.; Moreno-Sánchez, R.; Mendoza, S.; Manrique, N.; Mogollón, A,; Osorio, J.D.; Grajales, A.; Sánchez, J.A. Community involvement in management for maintaining coral reef resilience and biodiversity in southern Caribbean marine protected areas. Biodivers. Conserv. 2009, 18, 935–956. [Google Scholar] [CrossRef]
- Stern, M.J. The power of trust: toward a theory of local opposition to neighboring protected areas. Society Nat. Resources 2008, 21, 859–875. [Google Scholar] [CrossRef]
- Christie, P.; White, A.T. Best practices for improved governance of coral reef marine protected areas. Coral Reefs 2007, 26, 1047–1056. [Google Scholar] [CrossRef]
- Clark, R.B. Marine Pollution; Oxford University Press: Oxford, UK, 2001; p. 248. [Google Scholar]
© 2010 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
MDPI and ACS Style
Dikou, A. Ecological Processes and Contemporary Coral Reef Management. Diversity 2010, 2, 717-737. https://doi.org/10.3390/d2050717
AMA Style
Dikou A. Ecological Processes and Contemporary Coral Reef Management. Diversity. 2010; 2(5):717-737. https://doi.org/10.3390/d2050717
Chicago/Turabian StyleDikou, Angela. 2010. "Ecological Processes and Contemporary Coral Reef Management" Diversity 2, no. 5: 717-737. https://doi.org/10.3390/d2050717