The Distribution of Planktivorous Damselfishes (Pomacentridae) on the Great Barrier Reef and the Relative Influences of Habitat and Predation

Abstract
:1. Introduction
2. Materials and Methods
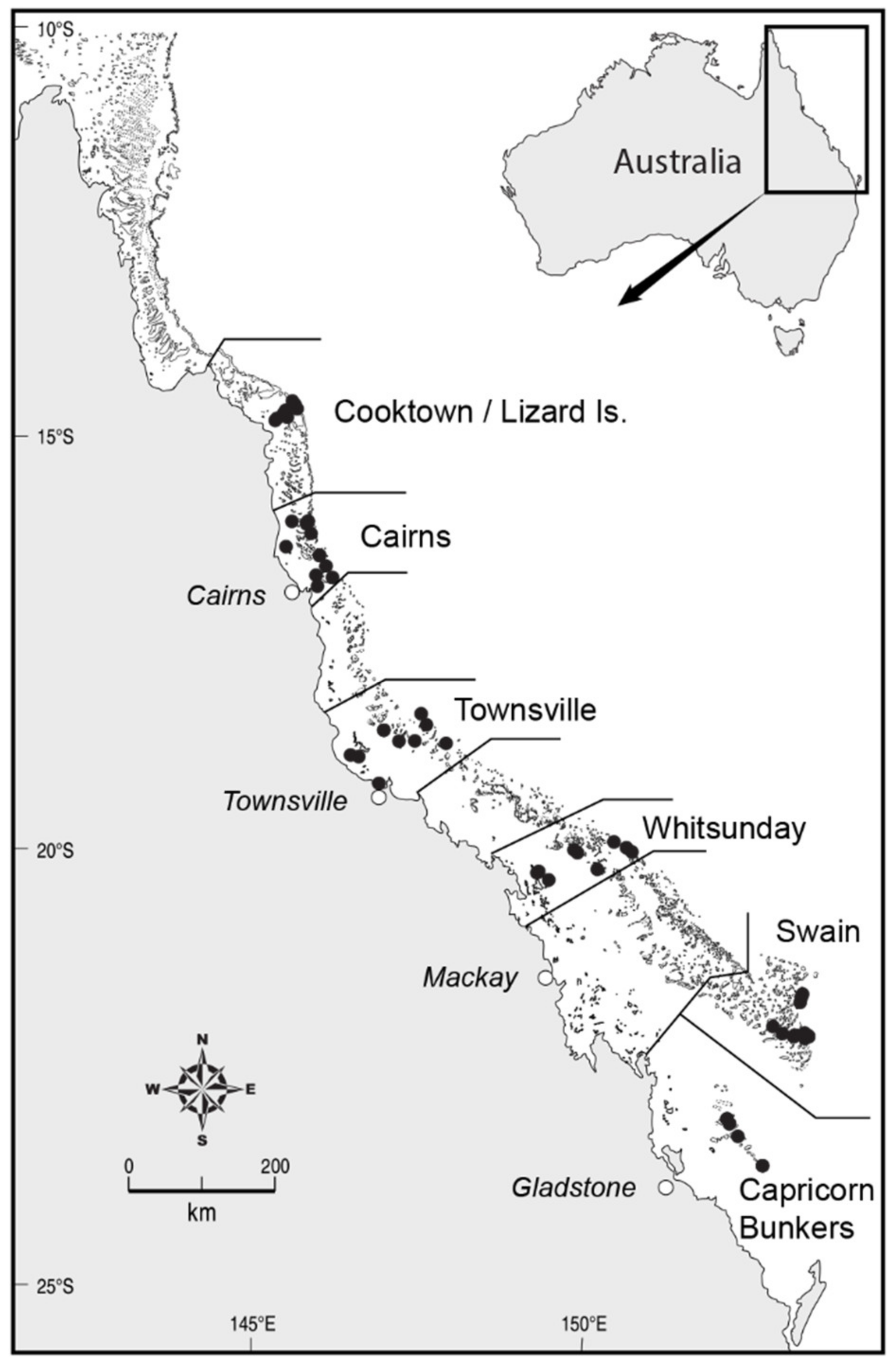
2.1. Survey Methods
2.2. Analyses of Latitudinal and Cross-Shelf Structure of Planktivorous Damselfish Assemblages
2.3. Top-Down versus Habitat Control of Planktivorous Damselfishes
3. Results
3.1. Distribution of Planktivorous Damselfishes
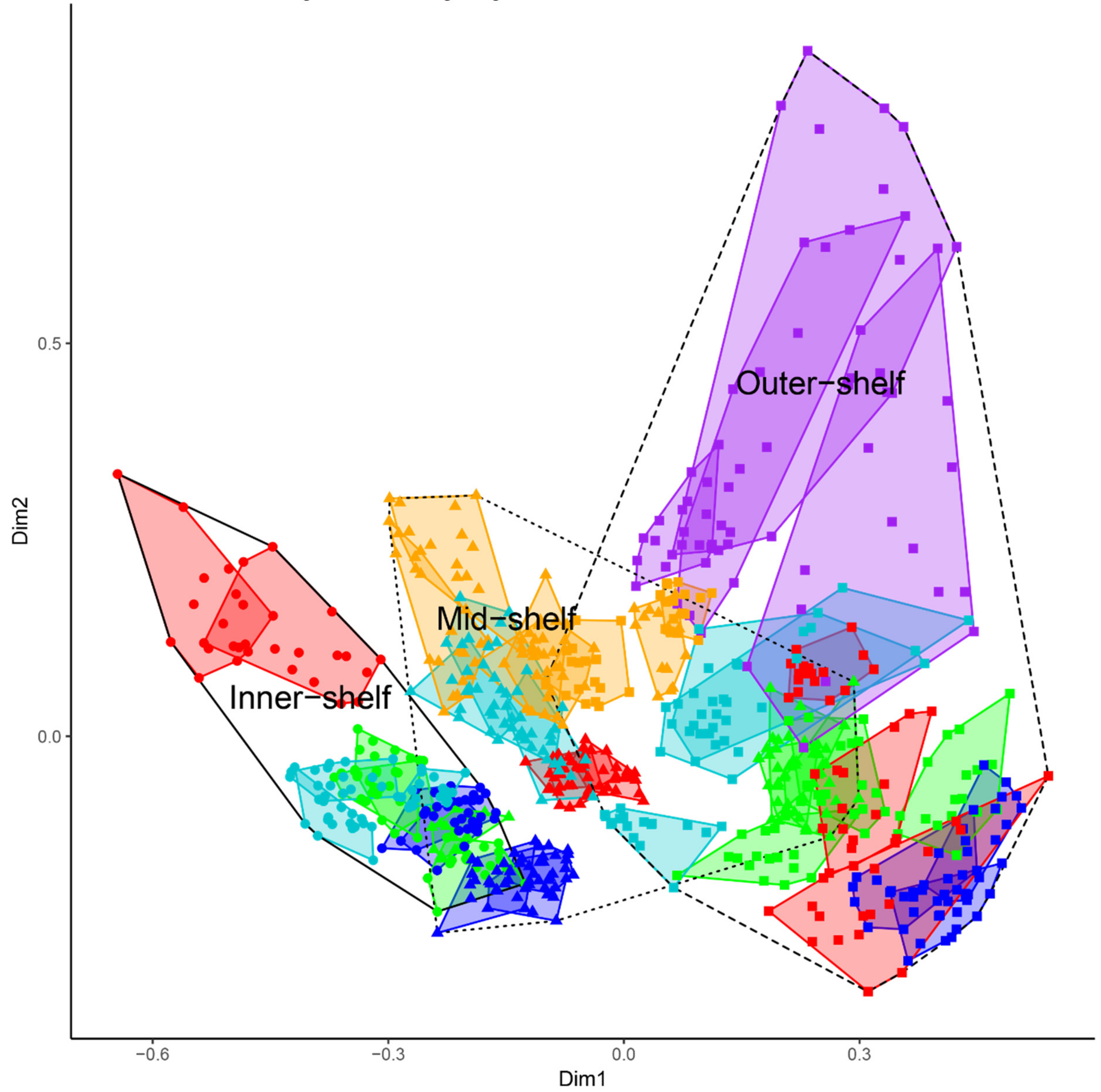
3.1.1. Cross-Shelf Patterns
3.1.2. Latitudinal Patterns
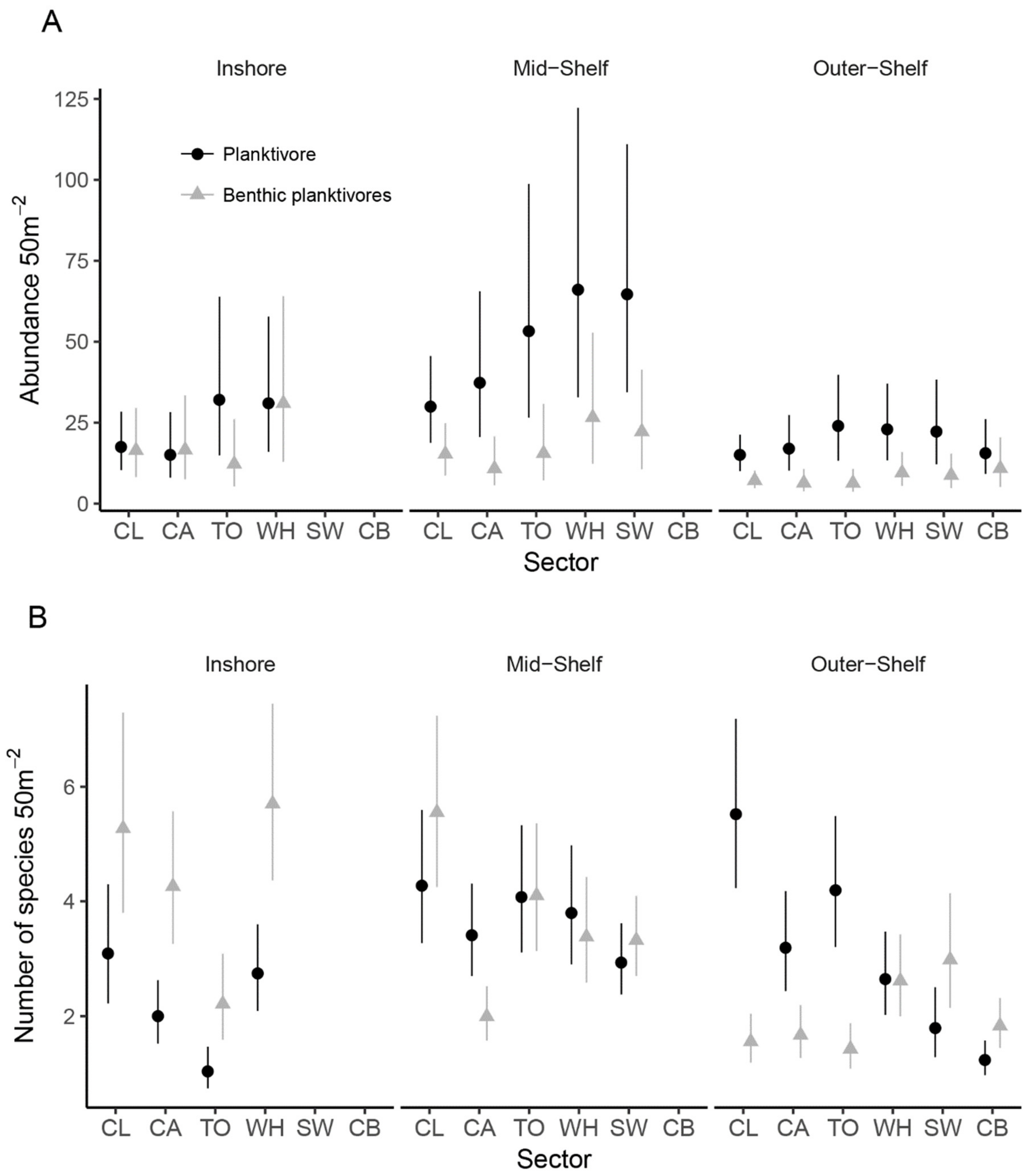
3.2. Abundance and Species Richness of Planktivorous Damselfishes
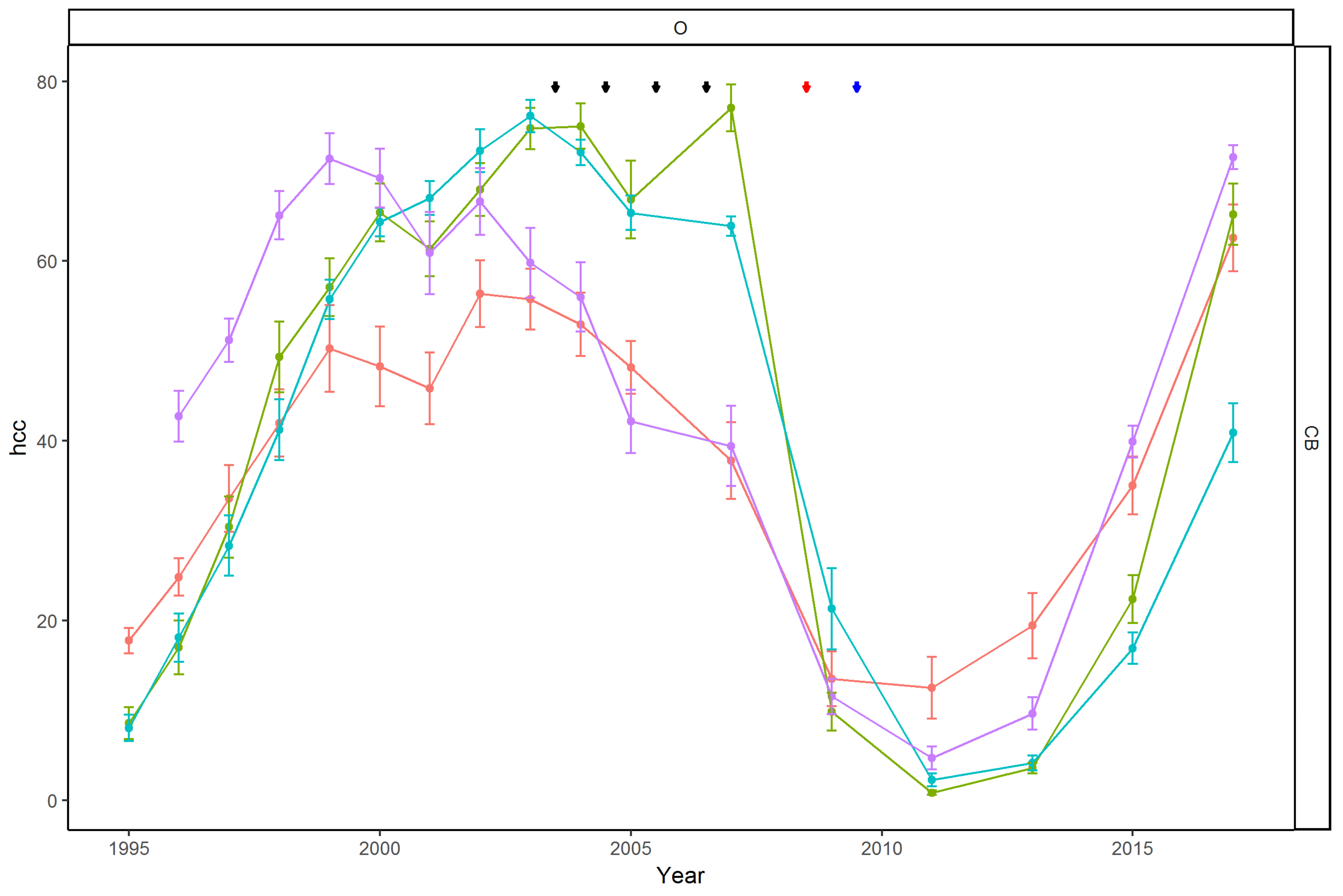
3.2.1. Sub-Regional Community Dynamics
3.2.2. Temporal Patterns
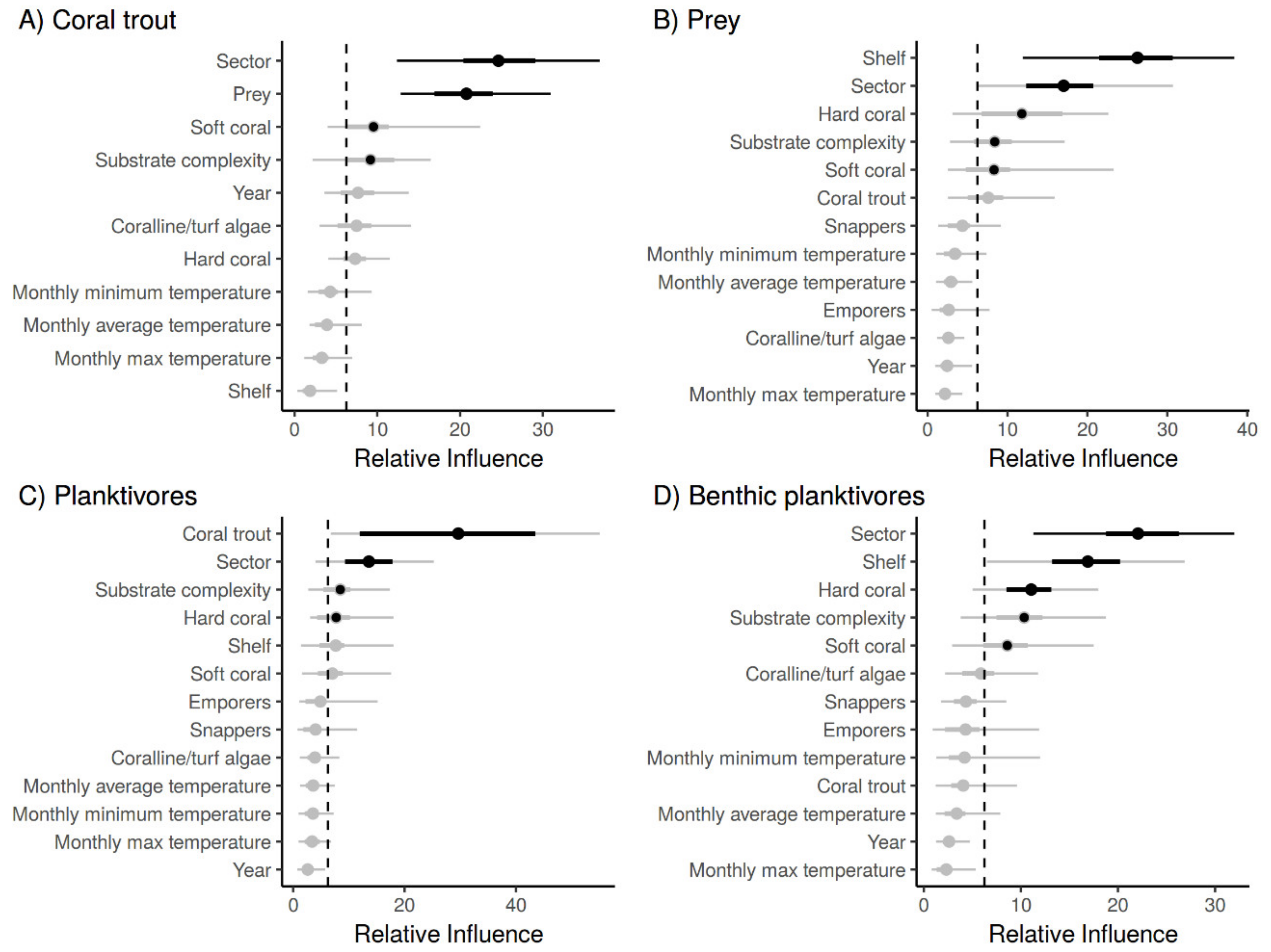
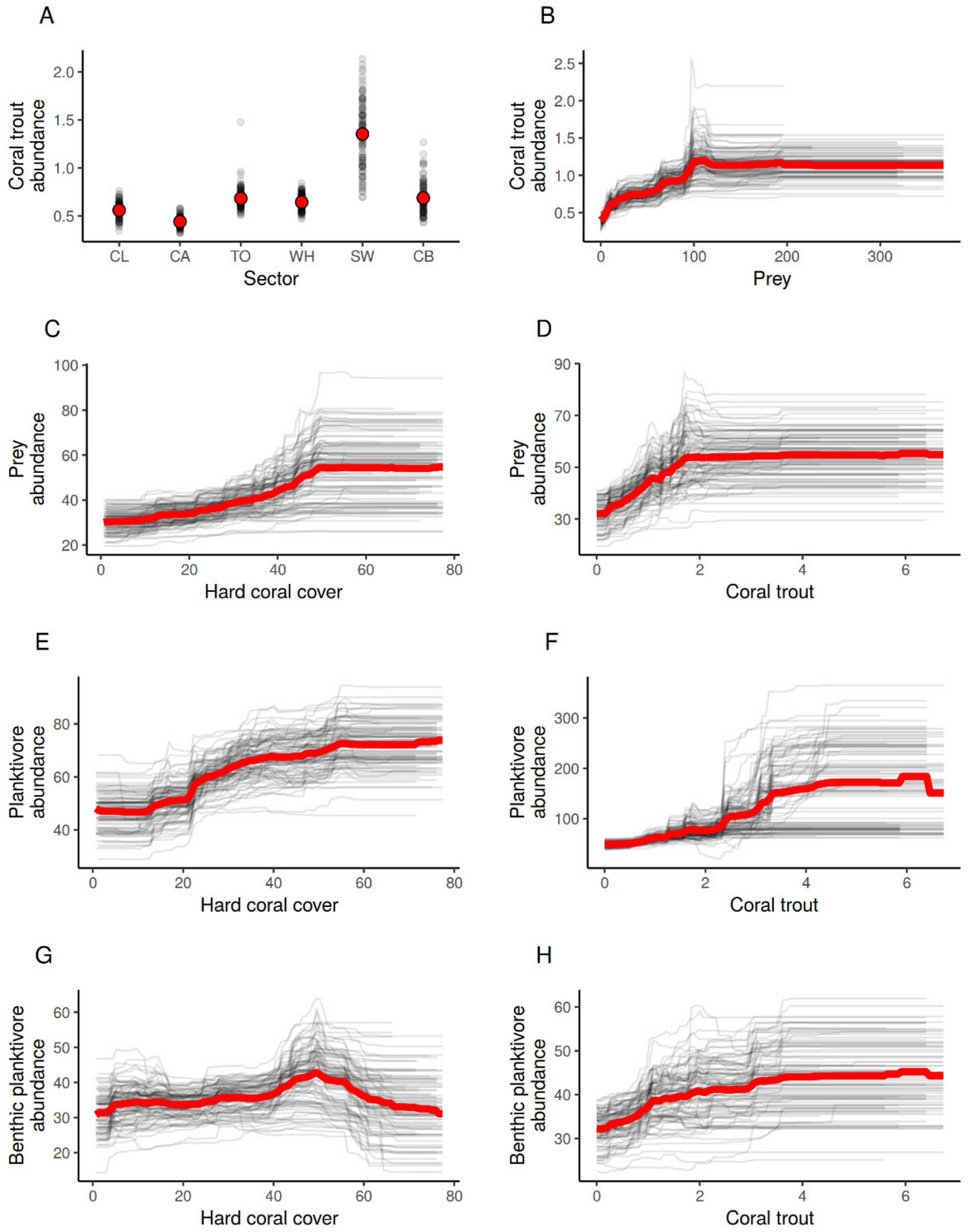
3.3. The Effects of Habitat and Predation on the Abundance of Planktivorous Damselfishes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A
Bootstrapped Gradient Boosted Regression Trees
Appendix B


References
- Hamner, W.M.; Jones, M.S.; Carleton, J.H.; Hauri, I.R.; Williams, D.M.B. Zooplankton, planktivorous fish, and water currents on a windward reef face: Great Barrier Reef, Australia. Bull. Mar. Sci. 1988, 42, 459–479. [Google Scholar]
- Pinnegar, J.K.; Polunin, N.V.C. Planktivorous damselfish support nitrogen and phosphorous fluxes to Mediterranean reefs. Mar. Biol. 2006, 148, 1089–1099. [Google Scholar] [CrossRef]
- Williams, D.M.B.; Hatcher, A.I. Structure of fish communities on outer slopes of inshore, mid-shelf and outer shelf reefs of the Great Barrier Reef. Mar. Ecol. Prog. Ser. 1983, 10, 239–250. [Google Scholar] [CrossRef]
- Cowan, Z.; Dworjanyn, S.A.; Caballes, C.F.; Pratchett, M.S. Predation on crown-of-thorns starfish larvae by damselfishes. Coral Reefs 2016, 35, 1253–1262. [Google Scholar] [CrossRef]
- Chase, T.J.; Pratchett, M.S.; Frank, G.E.; Hoogenboom, M.O. Coral-dwelling fish moderate bleaching susceptibility of coral hosts. PLoS ONE 2018, 13, e0208545. [Google Scholar] [CrossRef] [PubMed]
- Erez, J. On the importance of food sources in coral reef ecosystems. In Coral Reefs; Dubinsky, Z., Ed.; Elsevier: New York, NY, USA, 1990; pp. 411–418. [Google Scholar]
- St John, J. Feeding Ecology of the Coral Trout, Plectropomus leopardus (Serranidae), on the Great Barrier Reef, Australia. Ph.D. Thesis, James Cook University, Townsville, Australia, 1995. [Google Scholar]
- Frisch, A.J.; Ireland, M.; Baker, R. Trophic ecology of large predatory reef fishes: Energy pathways, trophic level and implications for fisheries in a changing climate. Mar. Biol. 2014, 161, 61–73. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Evans, R.D.; Russ, G.R. The effects of marine reserve protection on the trophic relationships of reef fishes on the Great Barrier Reef. Environ. Conserv. 2003, 30, 200–208. [Google Scholar] [CrossRef]
- Boaden, A.E.; Kingsford, M.J. Predators drive community structure in coral reef fish assemblages. Ecosphere 2015, 6, 1–33. [Google Scholar] [CrossRef]
- Gajdzik, L.; Parmentier, E.; Sturaro, N.; Frédérich, B. Trophic specializations of damselfishes are tightly associated with reef habitats and social behaviours. Mar. Biol. 2016, 163, 1–15. [Google Scholar] [CrossRef]
- Bellwood, D.R.; Hughes, T.P.; Folke, C.; Nyström, M. Confronting the coral reef crisis. Nature 2004, 429, 827–833. [Google Scholar] [CrossRef] [PubMed]
- Osborne, K.; Dolman, A.M.; Burgess, S.C.; Johns, K.A. Disturbance and the dynamics of coral cover on the Great Barrier Reef (1995–2009). PLoS ONE 2011, 6, e17516. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Alvarez-Noriega, M.; Alvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Berger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef] [PubMed]
- Pratchett, M.S.; Hoey, A.S.; Wilson, S.; Hobbs, J.P.; Allen, G. Habitat-use and specialisation among coral reef damselfishes. In Biology of Damselfishes; Frédérich, B., Paramentier, E., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 84–121. [Google Scholar]
- Hughes, T.P. Catastrophes, phase shifts and large-scale degradation of a Caribbean coral Reef. Science 1994, 265, 1547–1551. [Google Scholar] [CrossRef] [PubMed]
- Graham, N.A.J.; Wilson, S.K.; Jennings, S.; Polunin, N.V.C.; Bijoux, J.P.; Robinson, J. Dynamic fragility of oceanic coral reef ecosystems. Proc. Natl. Acad. Sci. USA 2006, 103, 8425–8429. [Google Scholar] [CrossRef] [PubMed]
- De’ath, G.; Fabricius, K.E.; Sweatman, H.; Puotinen, M. The 27-year decline of coral cover on the Great Barrier Reef and its causes. Proc. Natl. Acad. Sci. USA 2012, 109, 17995–17999. [Google Scholar] [CrossRef] [PubMed]
- Tribble, G.W.; Nishikawa, H. An analysis of the diets of four spatially overlapping damselfishes of the genus Chromis. Jpn. J. Ichthyol. 1982, 29, 267–271. [Google Scholar]
- Sweatman, H.P.A. The influence of adults of some coral reef fishes on larval recruitment. Ecol. Monogr. 1985, 55, 469–485. [Google Scholar] [CrossRef]
- Holbrook, S.J.; Forrester, G.E.; Schmitt, R.J. Spatial patterns in abundance of damselfish reflect availability of suitable habitat. Oecologia 2000, 122, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Frederich, B.; Fabri, G.; Lepoint, G.; Vandewalle, P.; Parmentier, E. Trophic niches of thirteen damselfishes (Pomacentridae) at the Grand Recif of Toliara, Madagascar. Ichthyol. Res. 2009, 56, 10–17. [Google Scholar] [CrossRef]
- Gajdzik, L.; Parmentier, E.; Michel, L.N.; Sturaro, N.; Soong, K.; Lepoint, G.; Frédérich, B. Similar levels of trophic and functional diversity within damselfish assemblages across Indo-Pacific coral reefs. Func. Ecol. 2018, 32, 1358–1369. [Google Scholar] [CrossRef]
- Eurich, J.G.; Matley, J.K.; Baker, R.; McCormick, M.I.; Jones, G.P. Stable isotope analysis reveals trophic diversity and partitioning in territorial damselfishes on a low-latitude coral reef. Mar. Biol. 2019, 166, 17. [Google Scholar] [CrossRef]
- Wilson, S.K.; Graham, N.A.J.; Pratchett, M.S.; Jones, G.P.; Polunin, N.V.C. Multiple disturbances and the global degradation of coral reefs: Are reef fishes at risk or resilient? Glob. Chang. Biol. 2006, 12, 2220–2234. [Google Scholar] [CrossRef]
- Cheal, A.J.; Wilson, S.K.; Emslie, M.J.; Dolman, A.M.; Sweatman, H. Responses of reef fish communities to coral declines on the Great Barrier Reef. Mar. Ecol. Prog. Ser. 2008, 372, 211–223. [Google Scholar] [CrossRef]
- Emslie, M.J.; Logan, M.; Ceccarelli, D.M.; Cheal, A.J.; Hoey, A.S.; Miller, I.; Sweatman, H.P.A. Regional-scale variation in the distribution and abundance of farming damselfishes on Australia’s Great Barrier Reef. Mar. Biol. 2012, 159, 1293–1304. [Google Scholar] [CrossRef]
- Emslie, M.J.; Cheal, A.J.; Johns, K.A. Retention of habitat complexity minimizes disassembly of reef fish communities following disturbance: A large-scale natural experiment. PLoS ONE 2014, 9, e105384. [Google Scholar] [CrossRef] [PubMed]
- Ceccarelli, D.M.; Emslie, M.J.; Richards, Z.T. Post-disturbance stability of fish assemblages measured at coarse taxonomic resolution masks changes at finer scales. PLoS ONE 2014, 11, e0156232. [Google Scholar] [CrossRef] [PubMed]
- Doherty, P.; Fowler, T. An empirical test of recruitment limitation in a coral reef fish. Science 1994, 263, 935–939. [Google Scholar] [CrossRef] [PubMed]
- Booth, D.J.; Wellington, G. Settlement preferences in coral-reef fishes: Effects on patterns of adult and juvenile distributions, individual fitness and population structure. Aust. J. Ecol. 1998, 23, 274–279. [Google Scholar] [CrossRef]
- Lynam, C.P.; Liope, M.; Möllman, C.; Helaouët, P.; Bayliss-Brown, G.A.; Stenseth, N.C. Interactions between top-down and bottom-up control in marine food webs. Proc. Natl. Acad. Sci. USA 2017, 114, 1952–1957. [Google Scholar] [CrossRef] [PubMed]
- Russ, G.R.; Questel, S.A.; Rizzari, J.R.; Alcala, A.C. The parrotfish-coral relationship: Refuting the ubiquity of a prevailing paradigm. Mar. Biol. 2015, 162, 2029–2045. [Google Scholar] [CrossRef]
- Carpenter, S.R.; Kitchem, J.E.; Hoixson, J.R. Cascading trophic interactions and lake productivity. Bioscience 1985, 35, 634–639. [Google Scholar] [CrossRef]
- Lapointe, B.E.; Littler, M.M.; Littler, D.S. Macroalgal overgrowth of fringing reefs ar Discovery Bay, Jamaica: Bottom-up versus top-down control. In Proceedings of the 8th International Coral Reef Symposium, Balboa, Panama, 24–29 June 1997; pp. 927–932. [Google Scholar]
- Hughes, T.P.; Smantz, A.M.; Steneck, R.; Carpenter, R.; Miller, S. Algal blooms on coral reefs: What are the causes? Limnol. Oceanogr. 1999, 44, 1583–1586. [Google Scholar] [CrossRef]
- Burkepile, D.E.; Hay, M.E. Nutrient versus herbivore control of macroalgal community development and coral growth on a Caribbean reef. Mar. Ecol. Prog. Ser. 2009, 389, 71–84. [Google Scholar] [CrossRef]
- Hixon, M.A.; Beets, J.P. Predation, prey refuges and the structure of coral-reef fish assemblages. Ecol. Monogr. 1993, 63, 77–101. [Google Scholar] [CrossRef]
- Carr, M.H.; Hixon, M.A. Predation effects on early postsettlement survivorship of coral-reef fishes. Mar. Ecol. Prog. Ser. 1995, 124, 31–42. [Google Scholar] [CrossRef]
- St., John, J.; Russ, G.R.; Brown, I.W.; Squire, L.C. The diet of the large coral reef serranid Plectropomus leopardus in two fishing zones on the Great Barrier Reef, Australia. Fish. Bull. 2001, 99, 180–192. [Google Scholar]
- Kingsford, M.J. Spatial and temporal variation in predation on reef fishes by coral trout (Plectropomus leopardus, Serranidae). Coral Reefs 1992, 11, 193–198. [Google Scholar] [CrossRef]
- Matley, J.K.; Tobin, A.J.; Simpfendorfer, C.A.; Fisk, A.T.; Heupel, M.R. Trophic niche and spatio-temporal changes in the feeding ecology of two sympatric species of coral trout (Plectropomus leopardus and P. laevis). Mar. Ecol. Prog. Ser. 2017, 563, 197–210. [Google Scholar] [CrossRef]
- Williams, D.M.B.; Dixon, P.; English, S. Cross-shelf distribution of copepods and fish larvae across the central Great Barrier Reef. Mar. Biol. 1988, 99, 577–589. [Google Scholar] [CrossRef]
- McKinnon, D.; Richardson, A.J.; Burford, M.A.; Furnas, M.J. Vulnerability of Great Barrier Reef plankton to climate change. In Climate Change and the Great Barrier Reef: A Vulnerability Assessment; Johnson, J.E., Marshall, P.A., Eds.; Great Barrier Reef Marine Park Authority and the Australian Greenhouse Office, Department of the Environment and Water Resources: Canberra, Australia, 2007; pp. 121–152. [Google Scholar]
- Sammarco, P.W.; Crenshaw, H. Plankton community dynamics of the central Great Barrier Reef lagoon: Analysis of data from Ikeda et al. Mar. Biol. 1984, 82, 167–180. [Google Scholar] [CrossRef]
- McKinnon, A.D.; Thorrold, S.R. Zooplankton community structure and copepod egg production in coastal waters of the central Great Barrier Reef lagoon. J. Plankton Res. 1993, 15, 1387–1411. [Google Scholar] [CrossRef]
- Emslie, M.J.; Cheal, A.J. Visual Census of Reef Fish: Standard Operational Procedure Number 3; Australian Institute of Marine Science: Townsville, Australia, 2018. [Google Scholar]
- Parmentier, E.; Gajdzik, L.; Olivier, D.; Frédérich, B. Trophic ecology of damselfishes. In Biology of Damselfishes; Frédérich, B., Paramentier, E., Eds.; CRC Press: Boca Raton, FL, USA, 2016; pp. 153–167. [Google Scholar]
- Jonker, M.; Johns, K.; Osborne, K. Surveys of Benthic Reef Communities Using Underwater Digital Photography and Counts of Juvenile Corals: Standard Operational Procedure Number 10; Australian Institute of Marine Science: Townsville, Australia, 2008. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Wickham, H. ggplot 2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2009. [Google Scholar]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Rue, H.; Martino, S.; Chopin, N. Approximate bayesian inference for latent Gaussian models by using integrated nested Laplace approximations. J. R. Stat. Soc. Ser. B. Stat. Method 2009, 71, 319–392. [Google Scholar] [CrossRef]
- De’ath, G. Boosted trees for ecological modelling and prediction. Ecology 2007, 88, 243–251. [Google Scholar] [CrossRef]
- Elith, J.; Leathwick, J.R.; Hastie, T. A working guide to boosted regression trees. J. Anim. Ecol. 2008, 77, 802–813. [Google Scholar] [CrossRef] [PubMed]
- Williamson, D.H.; Russ, G.R.; Ayling, A.M. No-take marine reserves increase abundance and biomass of reef fish on inshore fringing reefs of the Great Barrier Reef. Environ. Conserv. 2004, 31, 149–159. [Google Scholar] [CrossRef]
- Russ, G.R.; Cheal, A.J.; Dolman, A.M.; Emslie, M.J.; Evans, R.D.; Miller, I.; Sweatman, H.; Williamson, D.H. Rapid increase in fish numbers follows creation of world’s largest marine reserve network. Curr. Biol. 2008, 18, R514–R515. [Google Scholar] [CrossRef] [PubMed]
- Emslie, M.J.; Logan, M.; Williamson, D.H.; Ayling, A.M.; MacNeil, M.A.; Ceccarelli, D.; Cheal, A.J.; Evans, R.D.; Johns, K.A.; Jonker, M.J.; et al. Expectations and outcomes of reserve network performance following re-zoning of the Great Barrier Reef Marine Park. Curr. Biol. 2015, 25, 983–992. [Google Scholar] [CrossRef] [PubMed]
- Ridgeway, G. gbm: Generalized Boosted Regression Models, R package version 1:3; Foundation for Statistical Computing: Vienna, Austria, 2006. [Google Scholar]
- Drew, E.A. Halimeda biomass, growth rates and sediment generation on reefs in the central Great Barrier Reef province. Coral Reefs 1983, 2, 101–110. [Google Scholar] [CrossRef]
- Wismer, S.; Hoey, A.S.; Bellwood, D.R. Cross-shelf benthic community structure on the Great Barrier Reef: Relationships between macroalagal cover and herbivore biomass. Mar. Ecol. Prog. Ser. 2009, 376, 45–54. [Google Scholar] [CrossRef]
- Wilkinson, C.R.; Cheshire, A.C. Patterns in the distribution of sponge populations across the central Great Barrier Reef. Coral Reefs 1989, 8, 127–134. [Google Scholar] [CrossRef]
- Done, T.J. Patterns in the distribution of coral communities across the central Great Barrier Reef. Coral Reefs 1982, 1, 95–107. [Google Scholar] [CrossRef]
- Emslie, M.J.; Pratchett, M.S.; Cheal, A.J.; Osborne, K. Great Barrier Reef butterflyfish community structure: The role of shelf position and benthic community type. Coral Reefs 2010, 29, 705–715. [Google Scholar] [CrossRef]
- Williams, D.M.B. Patterns in the distribution of fish communities across the central Great Barrier Reef. Coral Reefs 1982, 1, 35–43. [Google Scholar] [CrossRef]
- Russ, G. Distribution and abundance of herbivorous grazing fishes in the central Great Barrier Reef. 1. Levels of variability across the entire continental shelf. Mar. Ecol. Prog. Ser. 1984, 20, 23–34. [Google Scholar] [CrossRef]
- Hoey, A.S.; Bellwood, D.R. Cross-shelf variation in the role of parrotfishes on the Great Barrier Reef. Coral Reefs 2008, 27, 37–47. [Google Scholar] [CrossRef]
- Emslie, M.J.; Cheal, A.J.; Logan, M. The distribution and abundance or reef-associated predatory fishes on the Great Barrier Reef. Coral Reefs 2017, 36, 829–846. [Google Scholar] [CrossRef]
- Cheal, A.; Emslie, M.; Miller, I.; Sweatman, H. The distribution of herbivorous fishes on the Great Barrier Reef. Mar. Biol. 2012, 159, 1143–1154. [Google Scholar] [CrossRef]
- Cheal, A.J.; Emslie, M.; MacNeil, M.A.; Miller, I.; Sweatman, H. Spatial variation in the functional characteristics of herbivorous fish communities and the resilience of coral reefs. Ecol. Appl. 2013, 23, 174–188. [Google Scholar] [CrossRef] [PubMed]
- Floeter, S.R.; Behrens, M.D.; Ferreira, C.E.L.; Paddack, N.J.; Horn, M.H. Geographical gradients of marine herbivorous fishes: Patterns and processes. Mar. Biol. 2005, 147, 1435–1447. [Google Scholar] [CrossRef]
- Epstein, H.E.; Kingsford, M.J. Are soft coral habitats unfavourable? A closer look at the association between reef fishes and their habitat. Environ. Biol. Fish. 2019. [Google Scholar] [CrossRef]
- Sano, M. Short-term effects of a mass coral bleaching event on a reef fish assemblage at Iriomote Island, Japan. Fish. Sci. 2004, 70, 41–46. [Google Scholar] [CrossRef]
- Bostrom-Einarsson, L.; Bonin, M.C.; Munday, P.L.; Jones, G.P. Loss of live coral compromises predator-avoidance behaviour in coral reef damselfish. Sci. Rep. 2018, 8, 7795. [Google Scholar] [CrossRef] [PubMed]
- Graham, N.A.J.; Nash, K.L. The importance of structural complexity in coral reef ecosystems. Coral Reefs 2013, 32, 315–326. [Google Scholar] [CrossRef]
- McClanahan, T.R.; Shafir, S.H. Causes and consequences of sea urchin abundance and diversity in Kenyan coral reef lagoons. Oecologia 1990, 83, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Grigg, R.W.; Polovina, J.J.; Atkinson, M.J. Model of a coral reef ecosystem III. Resource limitation, community regulation, fisheries yield and resource management. Coral Reefs 1984, 1, 23–27. [Google Scholar] [CrossRef]
- Polis, G.A.; Strong, D.R. Food web complexity and community dynamics. Am. Nat. 1996, 147, 813–846. [Google Scholar] [CrossRef]
- Shurin, J.B.; Borer, E.T.; Seabloom, E.W.; Anderson, K.; Blanchette, C.A.; Broitman, B.; Cooper, S.D.; Halpern, B.S. A cross-ecosystem comparison of the strength of trophic cascades. Ecol. Lett. 2002, 5, 785–791. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Coker, D.J.; Jones, G.P.; Munday, P.L. Specialization in habitat use by coral reef damselfishes and their susceptibility to habitat loss. Ecol. Evol. 2012, 2, 2168–2180. [Google Scholar] [CrossRef] [PubMed]
- Srinivasan, M. Depth distributions of coral reef fishes: The influence of microhabitat structure, settlement, and post-settlement processes. Oecologia 2003, 137, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Light, P.R.; Jones, G.P. Habitat preference in newly settled coral trout (Plectropomus leopardus, Serranidae). Coral Reefs 1997, 16, 117–126. [Google Scholar] [CrossRef]
- Kingsford, M.J. Contrasting patterns of reef utilization and recruitment of coral trout (Plectropomus leopardus) and snapper (Lutjanus carponotatus) at One Tree Island, southern Great Barrier Reef. Coral Reefs 2009, 28, 251–264. [Google Scholar] [CrossRef]
- Kerry, J.T.; Bellwood, D.R. The effect of coral morphology on shelter selection by coral reef fishes. Coral Reefs 2012, 31, 415–424. [Google Scholar] [CrossRef]
- Samoilys, M.A. Movement in a large predatory fish: Coral trout, Plectropomus leopardus (Pisces: Serranidae), on Heron Reef, Australia. Coral Reefs 1997, 16, 151–158. [Google Scholar] [CrossRef]
- Matley, J.K.; Tobin, A.J.; Lédée, E.J.I.; Heupel, M.R.; Simpfendorfer, C.A. Contrasting patterns of vertical and horizontal space use of two sympatric coral reef fish. Mar. Biol. 2016, 163, 253. [Google Scholar] [CrossRef]
- Hobson, E.S.; Chess, J.R. Trophic relationships among fishes and plankton in the lagoon at Enewetak Atoll, Marshall Islands. Fish. Bull. 1978, 76, 133–153. [Google Scholar]
- Sano, M.; Shimizu, M.; Nose, Y. Food Habits of Teleostean Reef Fishes in Okinawa Island, Southern Japan; University of Tokyo Bulletin, No. 25; University of Tokyo Press: Tokyo, Japan, 1984. [Google Scholar]
- Wilson, S.K.; Graham, N.A.J.; Polunin, N.V.C. Appraisal of visual assessments of habitat complexity and benthic composition on coral reefs. Mar. Biol. 2007, 151, 1069–1076. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CL | CA | TO | WH | SW | CB | Inshore | Mid-Shelf | Outer-Shelf | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AB | % Reefs (n = 47) | % | % | % | % | % | % | % | % | % | ||||||||||
| Planktivores | 711,500 | 100 | 14.01 | 49.05 | 13.51 | 38.24 | 21.33 | 76.05 | 23.77 | 74.88 | 24.29 | 97.70 | 3.10 | 21.95 | 15.95 | 45.76 | 56.75 | 89.20 | 27.30 | 42.54 |
| Pomacentrus lepidogenys | 229,555 | 100 | 5.57 | 6.29 | 16.44 | 15.01 | 20.67 | 23.78 | 21.53 | 21.89 | 31.49 | 40.86 | 4.31 | 9.84 | 1.86 | 1.72 | 65.19 | 33.06 | 32.95 | 16.56 |
| Neopomacentrus azysron | 150,068 | 98 | 14.27 | 10.54 | 14.00 | 8.36 | 22.06 | 16.60 | 42.89 | 28.51 | 5.59 | 4.75 | 1.18 | 1.76 | 17.26 | 10.45 | 70.66 | 23.42 | 12.08 | 3.97 |
| Chromis nitida | 106,722 | 41 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 18.08 | 8.55 | 78.05 | 47.09 | 3.86 | 4.10 | 13.50 | 5.81 | 81.21 | 19.14 | 5.30 | 1.24 |
| Neopomacentrus bankieri | 51,853 | 24 | 0.67 | 0.17 | 4.84 | 1.00 | 71.95 | 18.70 | 22.54 | 5.18 | 0.00 | 0.00 | 0.00 | 0.00 | 100 | 20.92 | 0.00 | 0.00 | 0.00 | 0.00 |
| Chromis margaritifer | 32,827 | 67 | 54.33 | 8.78 | 22.32 | 2.91 | 20.39 | 3.35 | 1.25 | 0.18 | 0.20 | 0.04 | 1.51 | 0.49 | 0.01 | <0.01 | 8.78 | 0.64 | 91.21 | 6.56 |
| Pomacentrus philippinus | 22,418 | 83 | 25.69 | 2.83 | 21.72 | 1.94 | 32.30 | 3.63 | 13.16 | 1.31 | 6.95 | 0.88 | 0.18 | 0.04 | 0.08 | 0.01 | 37.46 | 1.85 | 62.46 | 3.07 |
| Chrysiptera rollandi | 22,158 | 74 | 25.84 | 2.82 | 19.69 | 1.74 | 8.29 | 0.92 | 35.85 | 3.52 | 10.26 | 1.29 | 0.06 | 0.01 | 47.09 | 4.21 | 50.34 | 2.46 | 2.57 | 0.12 |
| Chromis ternatensis | 20,969 | 93 | 62.75 | 6.48 | 23.63 | 1.97 | 4.83 | 0.51 | 5.65 | 0.52 | 2.85 | 0.34 | 0.30 | 0.06 | 2.73 | 0.23 | 31.99 | 1.48 | 65.27 | 3.00 |
| Chromis weberi | 10,430 | 72 | 48.05 | 2.47 | 24.41 | 1.01 | 17.96 | 0.94 | 8.21 | 0.38 | 0.01 | <0.01 | 1.36 | 0.14 | 1.48 | 0.06 | 36.92 | 0.85 | 61.60 | 1.41 |
| Chromis atripectoralis | 4663 | 93 | 12.81 | 2.29 | 18.43 | 2.67 | 22.62 | 4.13 | 21.98 | 3.54 | 9.98 | 2.05 | 14.18 | 5.15 | 9.51 | 1.40 | 51.76 | 4.16 | 38.73 | 3.09 |
| Dascyllus reticulatus | 4600 | 78 | 44.65 | 1.01 | 34.67 | 0.63 | 10.35 | 0.24 | 5.80 | 0.12 | 0.26 | 0.01 | 4.26 | 0.20 | 11.51 | 0.22 | 55.98 | 0.57 | 32.17 | 0.32 |
| Chrysiptera talboti | 4362 | 83 | 46.03 | 0.99 | 5.23 | 0.09 | 22.95 | 0.50 | 23.77 | 0.46 | 1.97 | 0.05 | 0.05 | 0.00 | 7.04 | 0.12 | 82.99 | 0.80 | 9.97 | 0.10 |
| Chromis lepidolepis | 3344 | 67 | 29.93 | 0.49 | 31.55 | 0.42 | 30.59 | 0.51 | 7.57 | 0.11 | 0.06 | 0.00 | 0.30 | 0.01 | 1.11 | 0.01 | 31.85 | 0.24 | 67.05 | 0.49 |
| Chromis xanthura | 3101 | 54 | 83.10 | 1.27 | 9.77 | 0.12 | 5.51 | 0.09 | 1.19 | 0.02 | 0.32 | 0.01 | 0.10 | 0.00 | 0.03 | <0.01 | 8.13 | 0.06 | 91.84 | 0.62 |
| Chromis chrysura | 2454 | 30 | 5.13 | 0.06 | 1.51 | 0.01 | 87.45 | 1.08 | 4.32 | 0.05 | 0.04 | 0.00 | 1.55 | 0.04 | 0.00 | 0.00 | 0.73 | <0.01 | 99.27 | 0.53 |
| Chromis vanderbilti | 2147 | 35 | 60.74 | 0.64 | 7.59 | 0.06 | 28.60 | 0.31 | 0.09 | 0.00 | 0.05 | 0.00 | 2.93 | 0.06 | 0.00 | 0.00 | 0.28 | <0.01 | 99.72 | 0.47 |
| Pomachromis richardsoni | 1031 | 67 | 28.23 | 0.14 | 7.95 | 0.03 | 15.32 | 0.08 | 14.26 | 0.07 | 33.85 | 0.20 | 0.39 | 0.00 | 25.22 | 0.10 | 61.20 | 0.14 | 13.58 | 0.03 |
| Chromis viridis | 800 | 43 | 59.00 | 0.23 | 22.00 | 0.07 | 7.38 | 0.03 | 11.38 | 0.04 | 0.25 | <0.01 | 0.00 | 0.00 | 58.88 | 0.19 | 30.63 | 0.05 | 10.50 | 0.02 |
| Chromis iomelas | 627 | 28 | 27.91 | 0.09 | 3.67 | 0.01 | 63.16 | 0.20 | 5.26 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.64 | <0.01 | 99.36 | 0.14 |
| Neopomacentrus cyanomos | 622 | 30 | 19.61 | 0.06 | 4.02 | 0.01 | 1.29 | 0.00 | 75.08 | 0.21 | 0.00 | 0.00 | 0.00 | 0.00 | 89.07 | 0.22 | 10.77 | 0.01 | 0.16 | <0.01 |
| Chrysiptera flavipinnis | 598 | 37 | 37.12 | 0.11 | 0.00 | 0.00 | 0.17 | <0.01 | 26.76 | 0.07 | 30.77 | 0.10 | 5.18 | 0.03 | 3.51 | 0.01 | 65.05 | 0.09 | 31.44 | 0.04 |
| Chromis lineata | 305 | 28 | 62.38 | 0.09 | 29.51 | 0.04 | 7.21 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 14.75 | 0.01 | 85.25 | 0.06 |
| Dascyllus aruanus | 280 | 24 | 89.64 | 0.12 | 6.79 | 0.01 | 2.50 | <0.01 | 0.71 | <0.01 | 0.36 | <0.01 | 0.00 | 0.00 | 11.43 | 0.01 | 87.86 | 0.05 | 0.71 | <0.01 |
| Dascyllus trimaculatus | 205 | 61 | 14.63 | 0.01 | 24.39 | 0.02 | 29.27 | 0.03 | 10.24 | 0.01 | 11.71 | 0.01 | 9.76 | 0.02 | 2.93 | <0.01 | 67.32 | 0.03 | 29.76 | 0.01 |
| Chromis retrofasciata | 128 | 24 | 15.63 | 0.01 | 60.16 | 0.03 | 0.00 | 0.00 | 17.97 | 0.01 | 6.25 | <0.01 | 0.00 | 0.00 | 76.56 | 0.04 | 22.66 | 0.01 | 0.78 | <0.01 |
| Neoglyphidodon polyacanthus | 55 | 20 | 1.82 | <0.01 | 0.00 | 0.00 | 5.45 | <0.01 | 18.18 | <0.01 | 74.55 | 0.02 | 0.00 | 0.00 | 0.00 | 0.00 | 70.91 | 0.01 | 29.09 | <0.01 |
| Chromis amboinensis | 41 | 28 | 53.66 | 0.01 | 0.00 | 0.00 | 4.88 | <0.01 | 26.83 | <0.01 | 14.63 | <0.01 | 0.00 | 0.00 | 24.39 | <0.01 | 26.83 | <0.01 | 48.78 | <0.01 |
| Amblyglyphidodon aureus | 17 | 17 | 76.47 | 0.01 | 5.88 | 0.01 | 11.76 | 0.01 | 5.88 | 0.01 | 0.00 | 0.01 | 0.00 | 0.01 | 5.88 | <0.01 | 82.35 | <0.01 | 11.76 | <0.01 |
| Chromis agilis | 12 | 7 | 63.16 | 0.01 | 0.00 | 0.00 | 36.84 | <0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 100.0 | <0.01 |
| Benthic planktivores | 390,036 | 100 | 26.91 | 51.67 | 11.29 | 17.51 | 9.05 | 17.68 | 22.92 | 39.59 | 15.44 | 34.04 | 14.39 | 55.84 | 36.44 | 57.32 | 42.48 | 36.60 | 21.08 | 18.01 |
| Pomacentrus moluccensis | 173,246 | 89 | 26.91 | 22.94 | 13.47 | 9.28 | 10.58 | 9.18 | 25.59 | 19.64 | 22.27 | 21.81 | 1.18 | 2.03 | 43.81 | 30.62 | 52.47 | 20.08 | 3.72 | 1.41 |
| Pomacentrus coelestis | 62,661 | 87 | 2.24 | 0.69 | 1.16 | 0.29 | 2.75 | 0.86 | 3.29 | 0.91 | 7.04 | 2.49 | 83.53 | 52.08 | 1.53 | 0.39 | 11.24 | 1.56 | 87.23 | 11.97 |
| Pomacentrus brachialis | 54,702 | 74 | 46.54 | 12.53 | 9.59 | 2.09 | 1.86 | 0.51 | 38.23 | 9.26 | 3.78 | 1.17 | 0.01 | <0.01 | 50.60 | 11.16 | 48.78 | 5.89 | 0.62 | 0.07 |
| Acanthochromis polyacanthus | 35,186 | 100 | 38.05 | 6.59 | 13.73 | 6.59 | 21.55 | 6.59 | 22.19 | 6.59 | 3.48 | 6.59 | 1.00 | 6.59 | 45.33 | 6.43 | 29.27 | 2.28 | 25.40 | 1.96 |
| Amblyglyphidodon curacao | 31,096 | 96 | 22.43 | 3.43 | 12.31 | 1.52 | 12.36 | 1.93 | 18.07 | 2.49 | 34.50 | 6.06 | 0.32 | 0.10 | 27.80 | 3.49 | 57.33 | 3.94 | 14.87 | 1.01 |
| Pomacentrus amboinensis | 12,911 | 72 | 49.99 | 3.18 | 11.72 | 0.60 | 5.62 | 0.36 | 20.57 | 1.18 | 12.08 | 0.88 | 0.02 | <0.01 | 42.58 | 2.22 | 54.51 | 1.55 | 2.90 | 0.08 |
| Pomacentrus nagaskiensis | 6206 | 61 | 22.86 | 0.70 | 18.92 | 0.47 | 0.26 | 0.01 | 56.74 | 1.56 | 1.11 | 0.04 | 0.11 | 0.01 | 88.58 | 2.22 | 11.04 | 0.15 | 0.39 | 0.01 |
| Chrysiptera rex | 4976 | 67 | 17.26 | 0.42 | 36.09 | 0.71 | 15.03 | 0.37 | 17.20 | 0.38 | 2.93 | 0.08 | 11.48 | 0.57 | 0.16 | <0.01 | 13.79 | 0.15 | 86.05 | 0.94 |
| Amblyglyphidodon leucogaster | 4395 | 74 | 48.03 | 1.04 | 23.46 | 0.41 | 11.24 | 0.25 | 16.04 | 0.31 | 1.21 | 0.03 | 0.02 | 0.00 | 33.38 | 0.59 | 60.73 | 0.59 | 5.89 | 0.06 |
| Neoglyphidodon melas | 2327 | 100 | 4.17 | 0.05 | 14.48 | 0.13 | 8.94 | 0.10 | 30.30 | 0.31 | 39.79 | 0.52 | 2.32 | 0.05 | 19.25 | 0.18 | 29.18 | 0.15 | 51.57 | 0.26 |
| Amphiprion akindynos | 909 | 85 | 13.42 | 0.06 | 10.67 | 0.04 | 32.89 | 0.15 | 9.24 | 0.04 | 30.91 | 0.16 | 2.86 | 0.03 | 1.76 | 0.01 | 61.17 | 0.12 | 37.07 | 0.07 |
| Pomacentrus australis | 710 | 20 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 12.11 | 0.04 | 2.25 | 0.01 | 85.63 | 0.60 | 0.28 | <0.01 | 14.08 | 0.02 | 85.63 | 0.13 |
| Amphiprion melanopus | 447 | 48 | 0.45 | <0.01 | 12.98 | 0.02 | 52.35 | 0.12 | 2.68 | 0.01 | 31.54 | 0.08 | 0.00 | 0.00 | 0.45 | <0.01 | 82.10 | 0.08 | 17.45 | 0.02 |
| Amphiprion perideraion | 176 | 37 | 27.84 | 0.02 | 10.80 | 0.01 | 34.66 | 0.03 | 18.75 | 0.01 | 6.25 | 0.01 | 1.70 | 0.00 | 2.84 | <0.01 | 69.32 | 0.03 | 27.84 | 0.01 |
| Premnas biaculeatus | 61 | 17 | 62.30 | 0.02 | 37.70 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 45.90 | 0.01 | 54.10 | 0.01 | 0.00 | 0.00 |
| Factor | Degrees of Freedom | Sums of Squares | Mean Square | F Model | R2 | Pr (>F) |
|---|---|---|---|---|---|---|
| Sector | 5 | 114.60 | 22.920 | 139.13 | 0.161 | 0.001 |
| Shelf | 2 | 130.86 | 65.428 | 397.18 | 0.183 | 0.001 |
| Year | 1 | 2.86 | 2.859 | 17.36 | 0.004 | 0.001 |
| Sector*Shelf | 7 | 79.86 | 11.409 | 69.26 | 0.112 | 0.001 |
| Sector*Year | 5 | 4.28 | 0.855 | 5.19 | 0.006 | 0.001 |
| Shelf*Year | 2 | 1.52 | 0.759 | 4.61 | 0.002 | 0.001 |
| Sector*Shelf*Year | 7 | 3.59 | 0.513 | 3.11 | 0.005 | 0.001 |
| Residuals | 2286 | 376.57 | 0.165 | 0.527 | ||
| Total | 2315 | 714.13 | 1.000 |
| Chi-Square Test Statistics | Chi-Square Residuals | ||||||
|---|---|---|---|---|---|---|---|
| Comparison | χ2 | d.f. | p | NN | NP | PN | PP |
| HC versus prey | 23.817 | 3 | <0.001 | −1.356 | −3.064 | 1.021 | 3.398 |
| HC versus planktivores | 22.029 | 3 | <0.001 | −1.504 | −2.544 | 0.427 | 3.621 |
| HC versus benthic planktivores | 19.182 | 3 | <0.001 | −1.059 | 1.764 | −3.064 | 2.358 |
| HC versus Trout | 63.166 | 3 | <0.001 | −3.138 | −0.613 | −2.915 | 6.667 |
| Trout versus prey | 75.513 | 3 | <0.001 | 0.427 | −4.401 | −2.916 | 6.889 |
| Trout versus planktivores | 77.974 | 3 | <0.001 | −0.167 | −3.733 | −3.361 | 7.261 |
| Trout versus benthic planktivores | 58.961 | 3 | <0.001 | 0.427 | −2.173 | −4.252 | 5.998 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emslie, M.J.; Logan, M.; Cheal, A.J. The Distribution of Planktivorous Damselfishes (Pomacentridae) on the Great Barrier Reef and the Relative Influences of Habitat and Predation. Diversity 2019, 11, 33. https://doi.org/10.3390/d11030033
Emslie MJ, Logan M, Cheal AJ. The Distribution of Planktivorous Damselfishes (Pomacentridae) on the Great Barrier Reef and the Relative Influences of Habitat and Predation. Diversity. 2019; 11(3):33. https://doi.org/10.3390/d11030033
Chicago/Turabian StyleEmslie, Michael J., Murray Logan, and Alistair J. Cheal. 2019. "The Distribution of Planktivorous Damselfishes (Pomacentridae) on the Great Barrier Reef and the Relative Influences of Habitat and Predation" Diversity 11, no. 3: 33. https://doi.org/10.3390/d11030033
APA StyleEmslie, M. J., Logan, M., & Cheal, A. J. (2019). The Distribution of Planktivorous Damselfishes (Pomacentridae) on the Great Barrier Reef and the Relative Influences of Habitat and Predation. Diversity, 11(3), 33. https://doi.org/10.3390/d11030033