Photodynamic Effect of Hypericin on the Conformation and Catalytic Activity of Hemoglobin

1
School of Life Science and Shanghai Key Laboratory of Bio-Energy Crops, Shanghai University, Shanghai 200444, P. R. China
2
Department of Biochemistry and National Key Laboratory of Pharmaceutical Biotechnology, Nanjing University, Nanjing 210093, P. R. China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2008, 9(2), 145-153; https://doi.org/10.3390/ijms9020145
Submission received: 12 September 2007
/
Accepted: 30 January 2008
/
Published: 5 February 2008
(This article belongs to the Special Issue Interaction of Biological Molecules)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Hypericin, extracted from H. perforatum, can induce the generation of reactive oxygen species by visible light irradiation, which may consequently induce the conformational change of hemoglobin. We have not only employed UV-vis spectroscopy to observe the changes of UV-vis spectra of the protein, which reveals the conformational changes of the protein, but also employed electrochemical method to obtain its enhanced peroxidase activity. The photodynamic effect of hypericin on the conformation and catalytic activity of the protein has also been proven to be strongly dependent on the irradiation time, the hypericin concentration and the presence of oxygen. This work is beneficial not only to the fabrication of more sensitive hydrogen peroxide biosensor, but also to the guidance of the usage of this medicinal herb molecule, since the conformational change of the protein and the enhanced peroxidase can be easily obtained only by visible light irradiation on hypericin, the process of which is so common to happen.
1. Introduction
Study on photosensitizers has received more and more attention. However, the effect of the photosensitization on DNA, protein, cell, or even an organ should be given more attention, no matter whether the effect is harmful or helpful for the potential clinical application. Hypericin (HY, Scheme 1) is a phenanthroperylene quinine derivative. It can be extracted from H. perforatum, a popular medicine for the treatment of depression [1–8]. It has been used as a conventional medication for the treatment of depression and wound healing for a long time [9]. Recently, more and more interest has been given to some other important pharmaceutical potentials of this species, such as antivirus activity, anti-HIV, antibacterial activity and antitumor activity [10–12]. Most of the clinic functions are proposed to be related to the phototoxicity of HY. For instance, the cytotoxic activity can be greatly enhanced after light activation [9, 13–16]. The mechanism how it works is still to be explored, however, it is proposed that after being exposed in visible light at the wavelength of 540–600nm, HY will transfer light energy to oxygen to generate reactive oxygen species (ROS), which may further induce the apoptosis of cells [17]. Some other studies report that HY preferentially accumulates in the cell membranes, especially the mitochondria membrane, which might be the target in the photodynamic therapy [18]. Both the type I and type II photodynamic reactions can take place in the photoactivation of HY, resulting in the formation of radicals, such as singlet oxygen and superoxide radical [9].
In the paper, we report our studies of the photodynamic effect of HY on the structure and function of a protein by employing UV-vis spectroscopy and electrochemical methods. Hemoglobin (Hb), a kind of heme protein, is used as the target protein for this work. On one hand, the structure and function of Hb are relatively very clear, which is easier to get to know the information of the conformational and functional changes. On the other hand, it is relatively easier to design the experimental protocols, since its electrochemistry has been somewhat largely studied in our lab [19, 20]. Besides, we have found that the reactive molecules, such as hydroxyl radical, superoxide anions, may influence the peroxidase activity of the protein, which may be ascribed to the conformational transformation of the protein [21]. Based on the data obtained in this study, we found that visible light irradiation of the protein in the presence of HY would cause the change of its conformation. And, the peroxidase activity of the protein towards hydrogen peroxide (H2O2) can be obviously enhanced. Since the structural and formational changes can take place after the treatment of the protein with visible light, the process of which is so common to occur, this study should be very interesting to lots of scientists in different research areas.
2. Results and Discussion
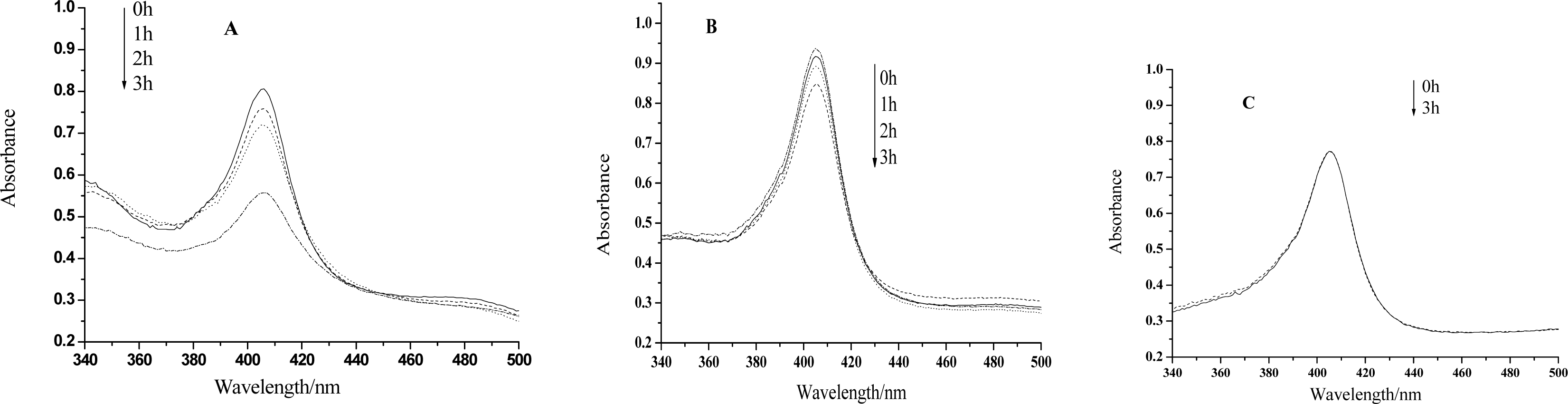
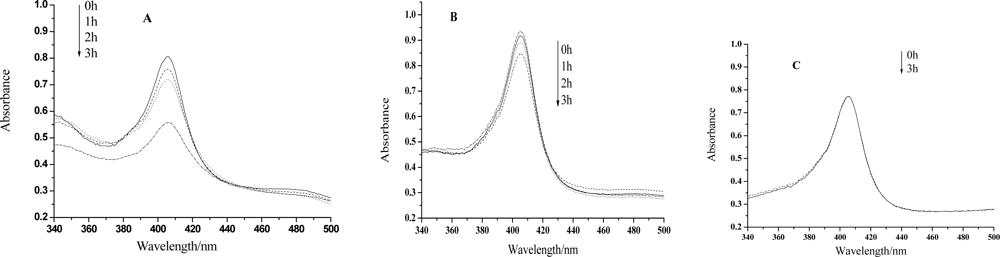
UV-vis spectroscopy is a very helpful technique to study the conformational changes of heme proteins, since the Soret band of the heme which is located at 407nm can provide very useful information on the secondary structure of heme proteins [23–26]. The position of the Soret band will shift or the absorption will decrease if the structure of a heme protein is transformed. Figure 1 shows that the absorption of the Soret band remains almost unchanged even after 3h irradiation by visible light in the absence of HY, however, the peak will decrease evidently only after 1 hour treatment if this medicinal species is in the presence. The longer the irradiation time is, the smaller the absorption peak is. Therefore, the heme moiety of the protein can be hardly influenced only by visible light. Nevertheless, irradiation treatment on Hb together with HY, the structure of the protein, especially the microenvironment of the heme ring, will be changed or even be badly damaged. We propose that the ROS, generated by the irradiation of HY, may react with the protein at the heme position, and change the microenvironment of the heme ring. It can be also observed from Fig.1 that the decrease of the absorption is much less significant for the case of lower HY concentration. So, the photodynamic effect of HY on Hb is not only a time dependent but also a concentration dependent process.
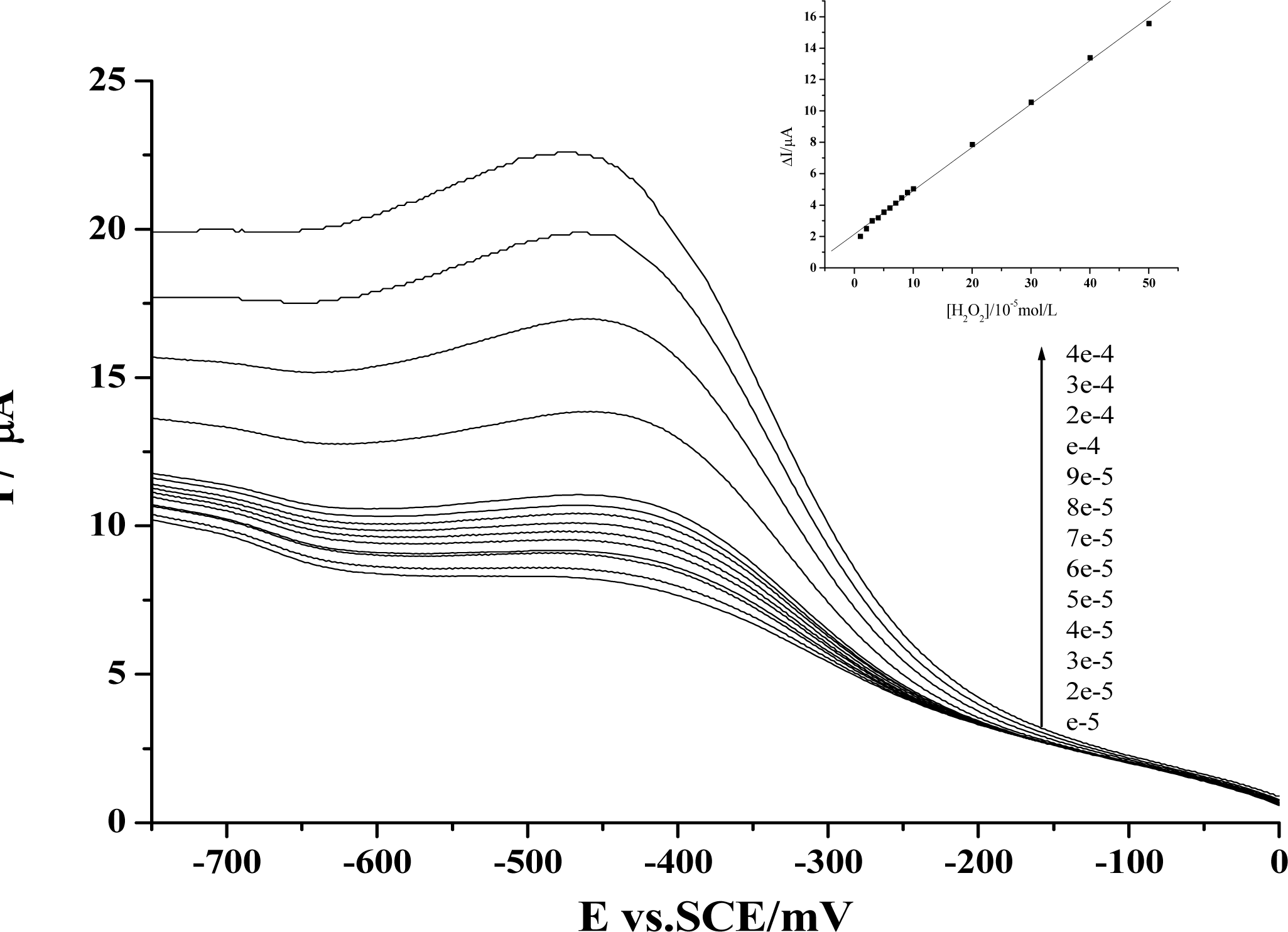
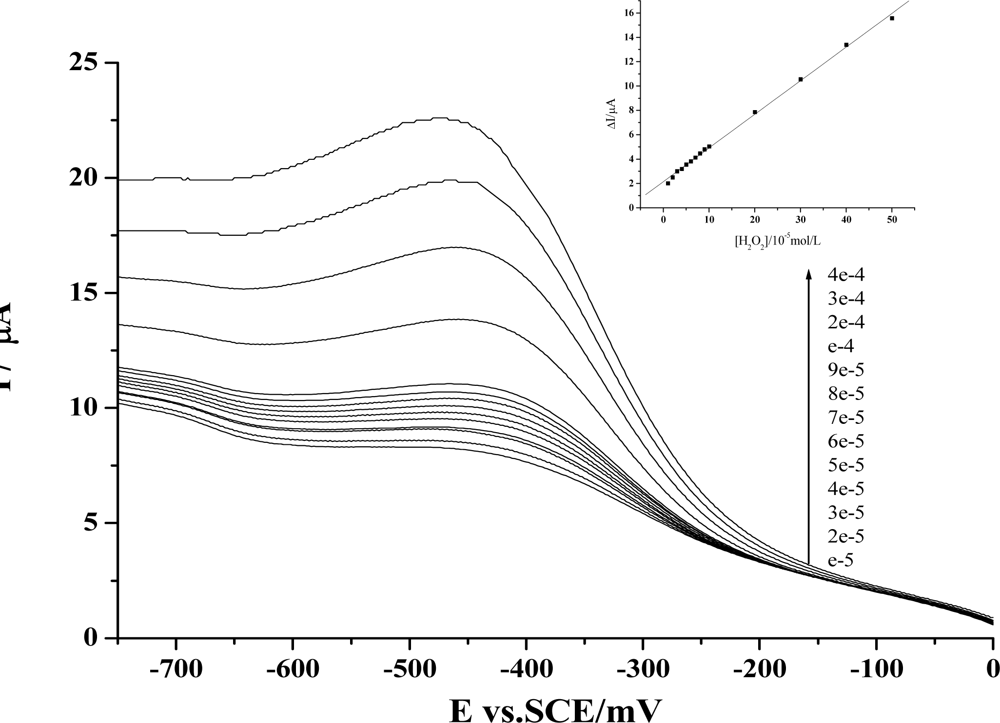
We have further employed electrochemical method to check whether the dynamic activity of HY may have any effect on the peroxidase activity of the protein. As is well known, the main physiological function of Hb is to carry oxygen. In the meantime, some other activities of this protein have also been revealed. Especially in recent years, the peroxidase activity of Hb has been largely studied, and these findings have been employed for the development of third-generation biosensors [27]. Based on our previous work of the preparation of protein-film modified electrodes and the fabrication of Hb-based H2O2 biosensors with this protein [19–21], we have first prepared an Hb-HY film modified electrode to study the catalytic activity of Hb towards H2O2. As is shown in Figure 2, the reduction peak of Hb will increase gradually with the addition of H2O2 in the test solution. Further studies reveal that a linear relationship between the increase of the reduction peak and the H2O2 concentration can be obtained from 1×10−5mol/L to 5×10−4mol/L, and the linear regression equation is: y = 2.15459 + 0.27599x, r = 0.999.
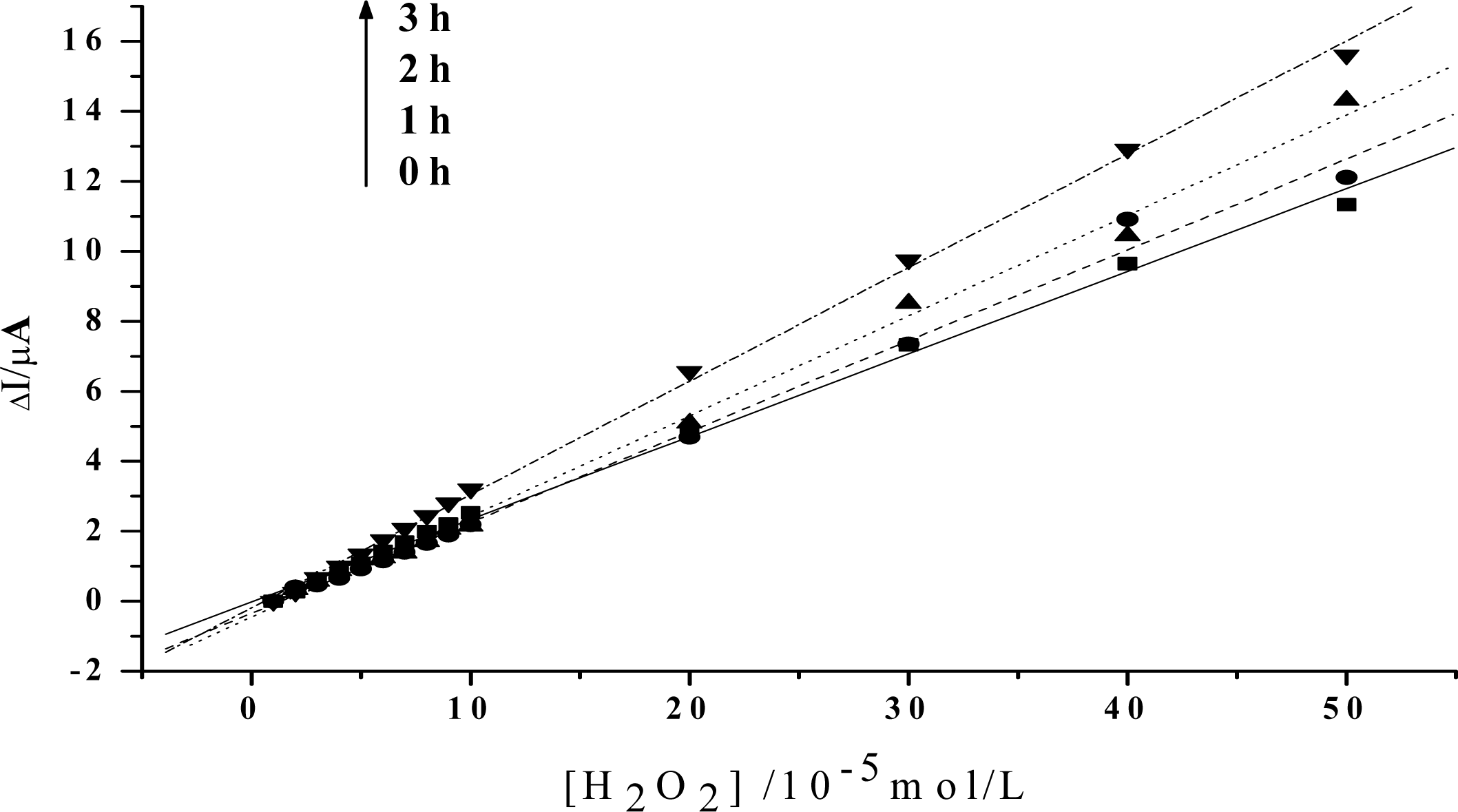
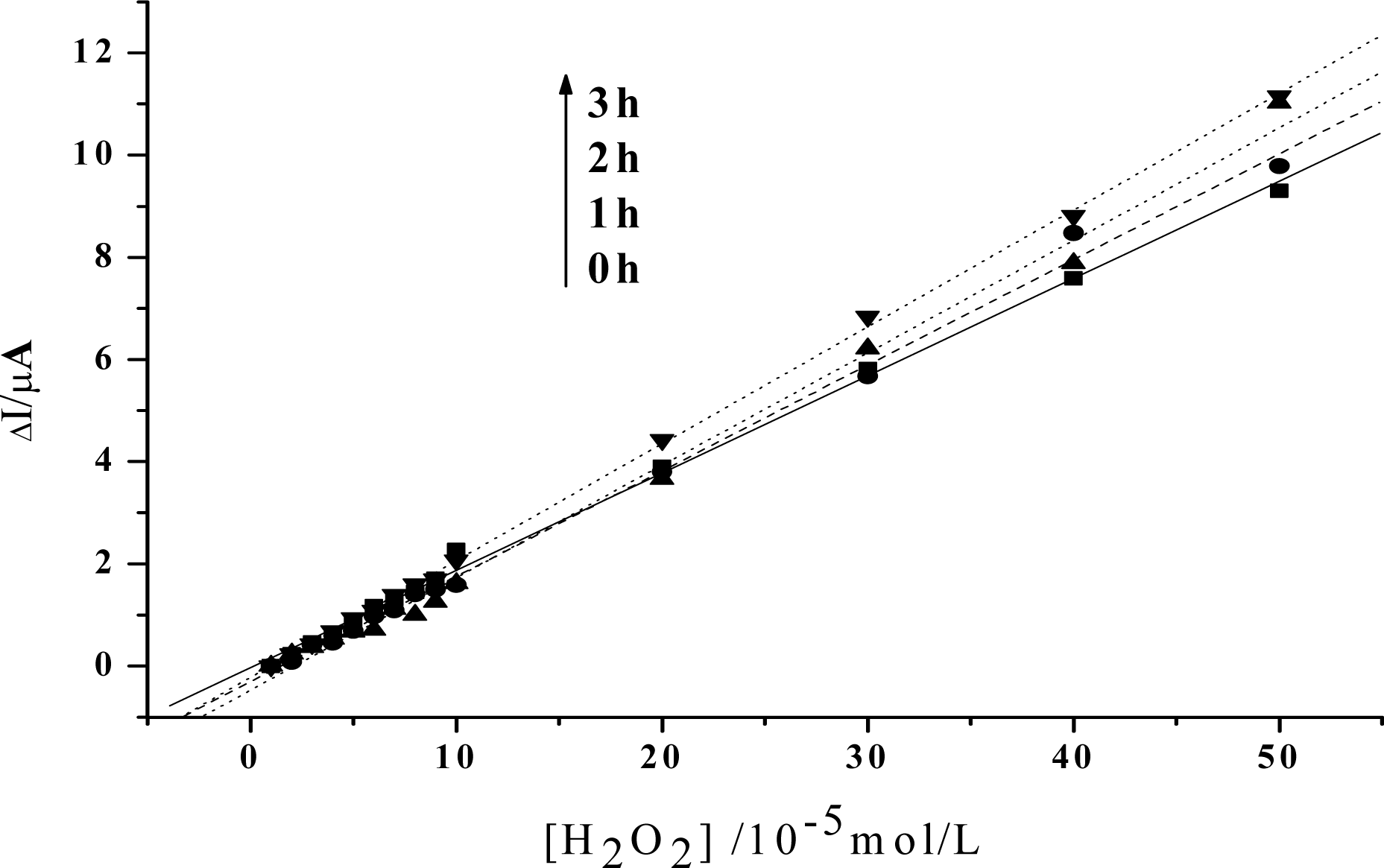
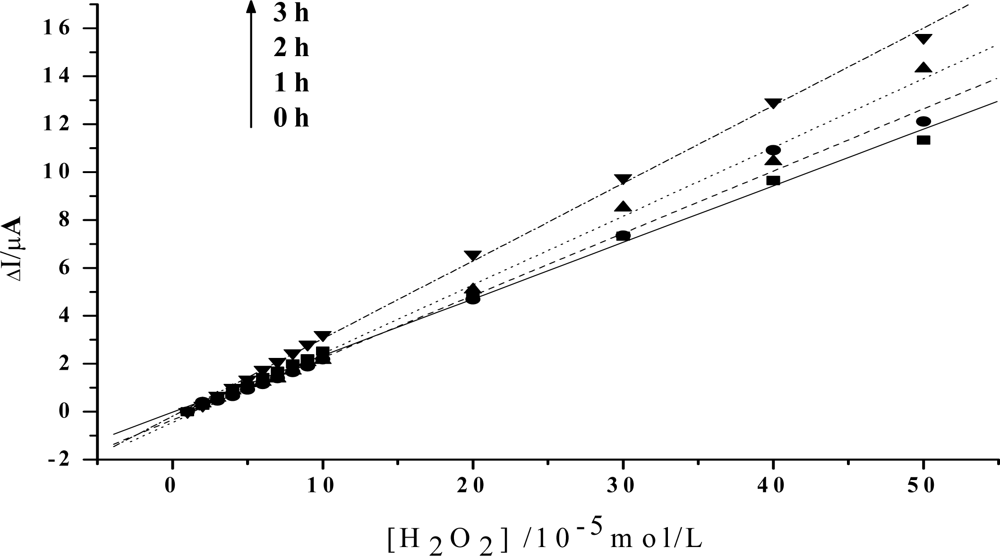
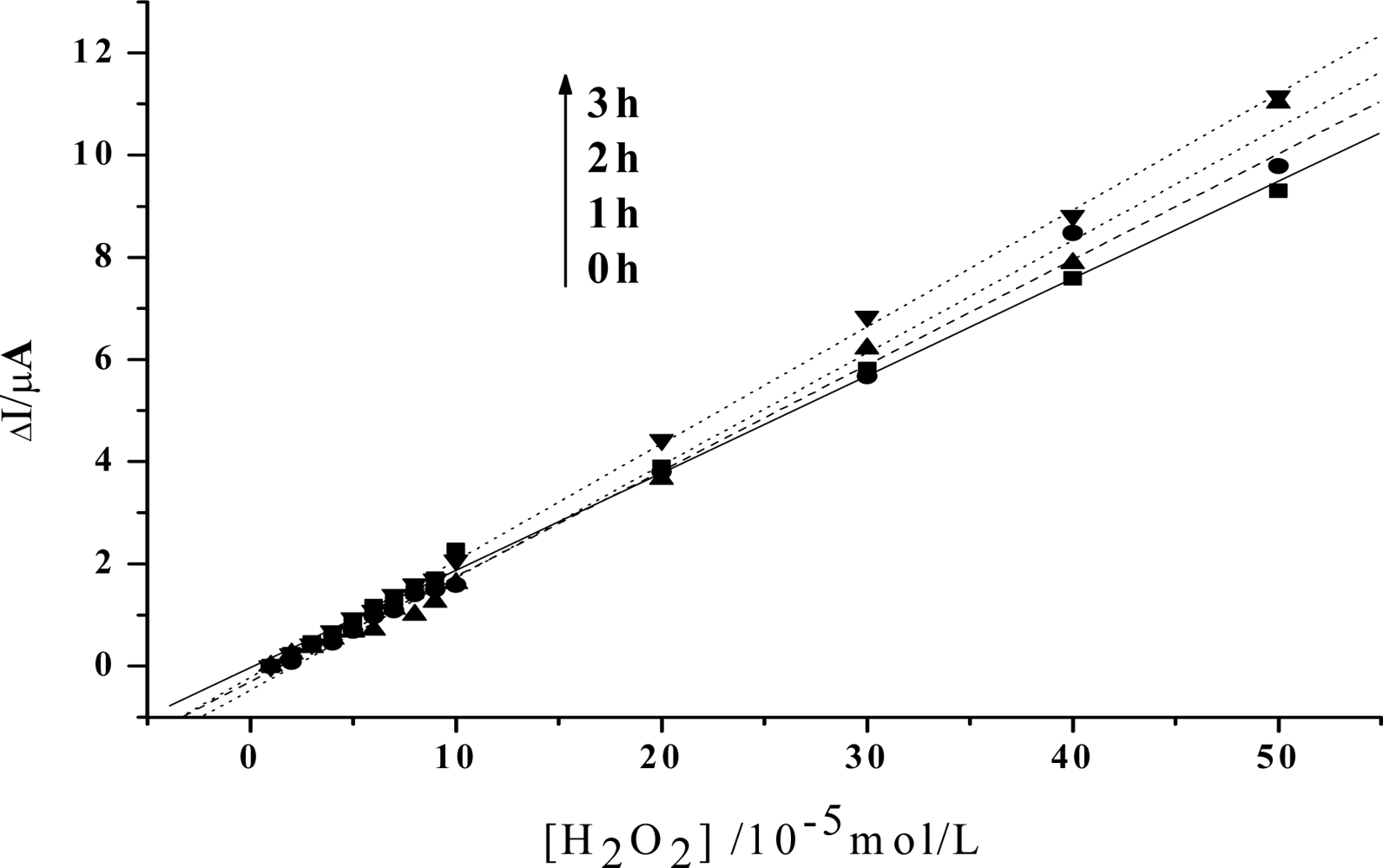
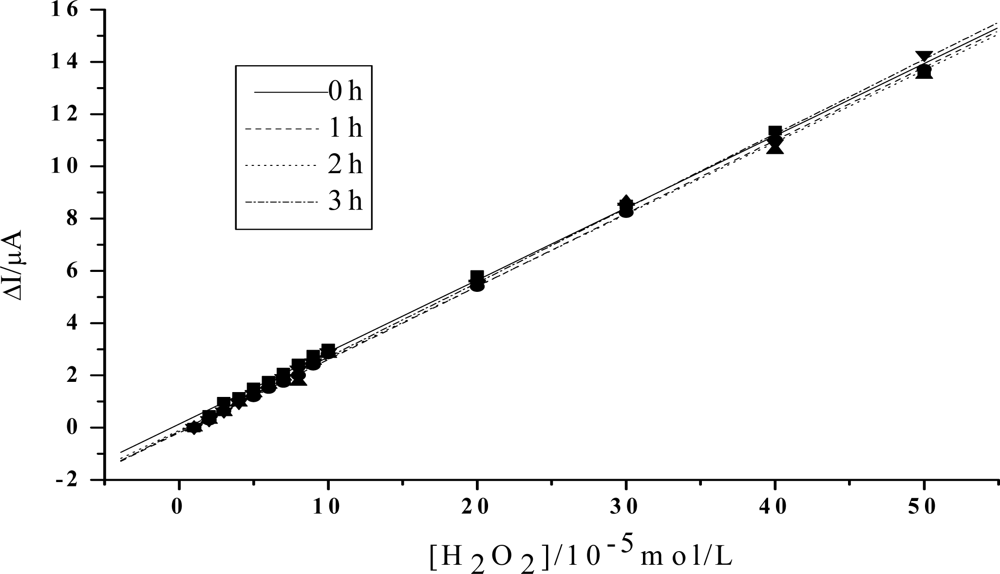
In order to obtain an obvious contrast, and to more clearly show the change of the peroxidase activity of the protein, the value of the reduction peak with the addition of 1×10−5mol/L H2O2 is chosen as the baseline, and is set to be 0. As is shown in Figure 3, the slope of the catalytic linearity is enhanced obviously after the irradiation of visible light on Hb together with HY. So, the catalytic activity of Hb towards H2O2 has been obviously improved. And, the slope increases with the irradiation time, which suggests that a longer time treatment on the samples will result in a higher catalytic ability of the protein. Meanwhile, concentration of HY is found to be another key factor. As is shown in Figure 4, although the slopes will also increase for the case of low HY concentration, the changes are indistinct in the first two hours. An obvious increase can be observed only after a 3 hours irradiation. And the increase is much smaller than that for the case with high HY concentration. These experimental results obtained with electrochemical method coincide very well with the results obtained by UV-vis spectroscopy, which further reveals that the photodynamic effect of HY is not only a time-dependent, but also a concentration-dependent process.
From our further studies, this effect has also been known to be an oxygen-dependent process, although O2 is not required for the antivirus activity of HY [28]. As is shown in Fig. 5, if the samples are prepared and irradiated in an anaerobic situation, the catalytic slope will keep unchanged even if a 3h treatment has been made. This is reasonable, since O2 is required in a photodynamic process to generate ROS, which is crucial in photoactivating HY. So, no influence of this medicinal species on the peroxidase activity of Hb can be obtained without O2 being involved. These results have also confirmed that the phtosensitization of HY would be achieved only in the presence of oxygen. After absorbing the light energy, HY will be transformed to be an excited state, which may further transfer the energy to oxygen directly or indirectly, both of which induce the formation of the ROS, especially the singlet oxygen. ROS then react with Hb, which may induce the change of the microenvironment of the heme position. As a result, the catalytic ability of the protein towards H2O2 is enhanced.
In summary, with UV-vis spectroscopic and electrochemical techniques, we have found that irradiation on the mixture of Hb and HY by the very common visible light can activate HY to generate ROS, which will make the change of the structure of Hb and enhance the catalytic activity of the protein towards the H2O2 reduction. This process depends not only on the irradiation time but also on the concentration of HY. Meanwhile, O2, which is required for the generation of ROS, is also found to be a vital element in the photoactive effect. This work has not only confirmed the photosensitization of HY and revealed the enhanced peroxidase activity of Hb, but also be helpful to the development of more sensitive H2O2 biosensor and the usage of this medicinal herb molecule.
3. Experimental Section
3.1 Reagents
HY (~99%) was purchased from Shanghai Healthjoy Chemical Co. Ltd. Stock solutions were prepared by dissolving known amounts of HY in 1 mL of dimethyl sulphoxide (DMSO). Hb from bovin blood was purchased from Sigma and used without further purification. Stock solutions were prepared by dissolving known amounts of Hb in 1 mL of double-distilled water. DMSO and H2O2 (30% (w/v) solution) were provided from Shanghai Jinshan Tingxin Chemical Reagent Co. and Nanjing Chemical Reagent Co., respectively. Other chemicals were of analytical grade. Double-distilled water, which was purified with a Milli-Q purification system (Branstead, Boston, MA, U.S.A.) to a specific resistance of >18M ω cm, was used in all the experiments, and all the solutions were stored in the refrigerator at 4 °C.
3.2 Preparation of UV-vis spectroscopic samples and light treatment
- Sample I preparation: 1μL HY solution (0.1 mol/L), 5μL Hb (8mg/ml, pH 7.0) and 4μL Tris-HCl buffer (pH 7.0) were mixed in a microcentrifuge tube.
- Sample II preparation: 0.5μL HY solution (0.1 mol/L), 5μL Hb (8mg/ml, pH 7.0) and 4μL Tris-HCl buffer (pH 7.0) were mixed in a microcentrifuge tube.
- Sample III preparation:1μL HY solution (0.1 mol/L), 5μL Hb (8mg/ml, pH 7.0) and 4μL Tris-HCl buffer (pH 7.0) were mixed in a microcentrifuge tube. This sample was thoroughly deoxygenated by blowing high-purity nitrogen for at least 1 min, and then sealed by parafilm.
The samples were exposed in the visible light emitted by a 200W filament lamp, which was filtered by an orange filer to get a wide band illumination above 580nm, for 1h, 2h, 3h, respectively, at a distance of 20cm.
3.3 Spectrocopic measurements
UV-vis spectroscopy was performed using a UV-2550 spectrophotometer (Shimadzu, Japan). After the treatment with visible light irradiation, the samples were then twentyfold diluted and analyzed immediately with the spectrophotometer. The blank was a 0.1M tris-HCl solution (pH 7.0).
3.4 Preparation of the modified electrode
The substrate pyrolytic graphite (PG) electrode (A = 6.28 mm2) was prepared by inserting a PG rod in a glass tube and fixing it with epoxy resin. Electrical contact was made by attaching a copper wire to the rod with the help of Wood's alloy (a fusible bismuth-based alloy). Before the modification of the substrate electrode with HY and the protein, the substrate PG electrode was first polished using rough and fine sand papers. Its surface was then polished to mirror smoothness with alumina (particle size of approx. 0.05 μm)/water slurry on silk. Finally, the electrode was thoroughly washed by ultrasonicating in both double-distilled water and ethanol for approx. 5 min. 10μL of HY and Hb mixture with their concentration being 1×10−2mol/L and 6.2×10−5mol/L, respectively, was mixed with 10μL DMSO (20%) and then spread evenly on the surface of the substrate PG electrode. The HY and Hb mixture had been previously treated with visible light irradiation before its immobilization onto PG electrode surface. The modified electrode was dried overnight at room temperature in the dark. After that, this electrode was thoroughly rinsed with double-distilled water and could be ready for use.
3.5 Electrochemical Apparatus
Cyclic voltammetry (CV) was performed with a PARC 263A Potentiostat/Galvanostat (EG&G; Princeton, NJ, USA), using a three-electrode configuration at 25 ± 0.5 °C. The reference electrode was a saturated calomel electrode (SCE) and the counter electrode was a platinum electrode. Potentials are reported with respect to SCE unless specially stated. All the test solutions were thoroughly deoxygenated by bubbling high-purity nitrogen through the solution for at least 10 min. A stream of nitrogen was blown gently across the surface of the solution in order to maintain the solution anaerobic throughout the experiments.
Acknowledgments
This work is supported by the National Natural Science Foundation of China (Grant No. 90406005, 20575028), the Program for New Century Excellent Talents in University, the Chinese Ministry of Education (NCET-04-0452) and the Shanghai Municipal Education Commission (No.06AZ051).
References and Notes
- Pace, N; MacKinney, G. Hypericin. The Photodynamic Pigment from St. John's Wort. J Am Chem Soc 1941, 63, 2570–2574. [Google Scholar]
- Bilia, AR; Gallori, S; Vincieri, FF. St. John's Wort and depression: efficacy, safety and tolerability – an update. Life Sci 2002, 70, 3077–3096. [Google Scholar]
- Greeson, JM; Sanford, B; Monti, DA. St. John's Wort (Hypericum perforatum):a review of the current pharmacological, toxicological, and clinical literaturel. Psychopharmacology (Berl) 2001, 153, 402–414. [Google Scholar]
- Barnes, J; Anderson, LA; Phillipson, JD. St John's Wort (Hypericum perforatum L.): a review of its chemistry, pharmacology and clinical properties. J Pharm Pharmacol 2001, 53, 583–600. [Google Scholar]
- Miskovsky, P; Sureau, F; Chinsky, L; Turpin, PY. Subcellular distribution of hypericin in human cancer cells. Photochem Photobiol 1995, 62, 546–549. [Google Scholar]
- Kamuhabwa, AR; Augustijns, P; De Witte, PA. In vitro transport and uptake of protohypericin and hypericin in the Caco-2 model. Int J Pharm 1999, 188, 81–86. [Google Scholar]
- Chen, B; de Witte, PA. Photodynamic therapy efficacy and tissue distribution of hypericin in a mouse P388 lymphoma tumor model. Cancer Lett 2000, 150, 111–117. [Google Scholar]
- Andreoni, A; Colasanti, A; Colasanti, P; Mastrocinque, M; Riccio, P; Roberti, G. Laser photosensitization of cells by hypericin. Photochem Photobiol 1994, 59, 529–533. [Google Scholar]
- Guedes, RC; Eriksson, LA. Theoretical study of hypericin. J Photochem Photobiol A-Chem 2005, 172, 293–299. [Google Scholar]
- Miskovsky, P. A New Antiviral and Antitumor Photosensitizer: Mechanism of Action and Interaction with Biological Macromolecules. Curr Drug Targets 2002, 3, 55–84. [Google Scholar]
- Wang, S; Liang, J; Yang, L; Cui, Y; Shang, R; Luo, Y. Studies on the inhibitory effect of hypericin on foot-and-mouth virus in vitro. Chin J Vet med 2006, 42, 6–9. [Google Scholar]
- Ulicny, J; Laaksonen, A. Hypericin, an intriguing internally heterogenous molecule, forms a covalent intramolecular hydrogen bond. Chem Phys Lett 2000, 319, 396–402. [Google Scholar]
- Hadjur, C; Richard, MJ; Parat, MO; Jardon, P; Favier, A. Photodynamic effects of hypericin on lipid peroxidation and antioxidant status in melanoma cells. Photochem Photobiol 1996, 64, 375–381. [Google Scholar]
- Theodossiou, T; Spiro, MD; Jacobson, J; Hothersall, JS; Mac-Robert, AJ. Evidence for intracellular aggregation of hypericin and the impact on its photocytotoxicity in PAM 212 murine keratinocytes. Photochem Photobiol 2004, 80, 438–443. [Google Scholar]
- Thomas, C; MacGill, RS; Miller, GC; Pardini, RS. Photoactivation of hypericin generates singlet oxygen in mitochondria and inhibits succinoxidase. Photochem Photobiol 1992, 55, 47–53. [Google Scholar]
- Thomas, C; Pardini, RS. Oxygen dependence of hypericininduced phototoxicity to EMT6 mouse mammary carcinoma cells. Photochem Photobiol 1992, 55, 831–837. [Google Scholar]
- Lu, WD; Atkins, WM. A Novel Antioxidant Role for Ligandin Behavior of Glutathione S-Transferases: Attenuation of the Photodynamic Effects of Hypericin. Biochemistry 2004, 43, 12761–12769. [Google Scholar]
- Theodossiou, TA; Noronha-Dutra, A; Hothersall, JS. Mitochondria are a primary target of hypericin phototoxicity:Synergy of intracellular calcium mobilisation in cell killing. Int J Biochem Cell B 2006, 38, 1946–1956. [Google Scholar]
- Wang, H; Guan, R; Fan, C; Zhu, D; Li, G. A hydrogen peroxide biosensor based on the bioelectrocatalysis of hemoglobin incorporated in a kieselgubr film. Sensors and Actuators B 2002, 84, 214–218. [Google Scholar]
- Fan, C; Gao, Q; Zhu, D; Wagner, G; Li, G. An unmediated hydrogen peroxide biosensor based on hemoglobin incorporated in a montmorillonite membrane. Analyst 2001, 126, 1086–1089. [Google Scholar]
- Zhou, H; Gan, X; Wang, J; Zhu, X; Li, G. Hemoglobin-Based Hydrogen Peroxide Biosensor Tuned by the Photovoltaic Effect of Nano Titanium Dioxide. Anal Chem 2005, 77, 6102–6104. [Google Scholar]
- Geoge, P; Hanania, G. A spectrophotometric study of ionizations in methaemoglobin. Biochem J 1953, 55, 236–243. [Google Scholar]
- Sakai, H; Masada, Y; Onuma, H; Takeoka, S; Tsuchida, E. Reduction of Methemoglobin via Electron Transfer from Photoreduced Flavin: Restoration of O2-Binding of Concentrated Hemoglobin Solution Coencapsulated in Phospholipid Vesicles. Bioconjugate Chem 2004, 15, 1037–1045. [Google Scholar]
- Sakai, H; Onuma, H; Umeyama, M; Takeoka, S; Tsuchida, E. Photoreduction of Methemoglobin by Irradiation in the Near-Ultraviolet Region. Biochemistry 2000, 39, 14595–14602. [Google Scholar]
- Teramura, Y; Kanazawa, H; Sakai, H; Takeoka, S; Tsuchida, E. Prolonged Oxygen-Carrying Ability of Hemoglobin Vesicles by Coencapsulation of Catalase in Vivo. Bioconjugate Chem 2003, 14, 1171–1176. [Google Scholar]
- Brown, WD; Snyder, HE. Nonenzymatic reduction and oxidation of myoglobin and hemoglobin by nicotinamide adenine dinucleotides and flavins. J Boil Chem 1969, 244, 6702–6706. [Google Scholar]
- Li, G. Encyclopedia of Sensor; Grimes, CA, Dickey, EC, Pishko, MV, Eds.; American Scientific Publishers: Valencia, CA, 2006; pp. 301–313. [Google Scholar]
- Sanchez-Cortes, S; Miskovsky, P; Jancura, D; Bertoluzza, A. Specific Interactions of Antiretroviraly Active Drug Hypericin with DNA As Studied by Surface-Enhanced Resonance Raman Spectroscopy. J Phys Chem 1996, 100, 1938–1944. [Google Scholar]
Figure 1.
(A)UV-vis spectra of hemoglobin mixed with HY after visible light irradiation for 0h, 1h, 2h, 3h, respectively. HY concentration: 5×10−4mol/L. Hb concentration: 3.10×10−6mol/L. (B) is the case for HY concentration of 2.5×10−4mol/L. (C) is the case for the absence of HY.
Figure 1.
(A)UV-vis spectra of hemoglobin mixed with HY after visible light irradiation for 0h, 1h, 2h, 3h, respectively. HY concentration: 5×10−4mol/L. Hb concentration: 3.10×10−6mol/L. (B) is the case for HY concentration of 2.5×10−4mol/L. (C) is the case for the absence of HY.
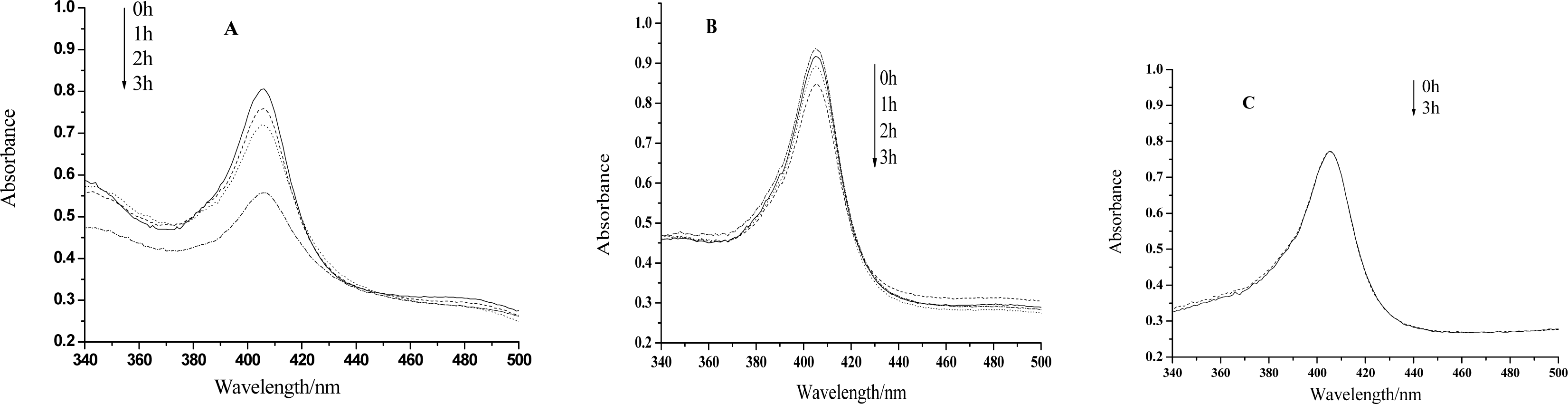
Figure 2.
Voltammograms of the reduction peak obtained at a Hb-HY modified electrode for pH 7.0 Tris-HCl buffer with the additions of different concentrations of H2O2. Inset is plots of the reduction peak current against H2O2 concentration. Scan rate: 200 mV s-1. For the Hb-HY modified electrode preparation, the HY and Hb concentrations are separately 1×10−2mol/L and 6.20×10−5mol/L
Figure 2.
Voltammograms of the reduction peak obtained at a Hb-HY modified electrode for pH 7.0 Tris-HCl buffer with the additions of different concentrations of H2O2. Inset is plots of the reduction peak current against H2O2 concentration. Scan rate: 200 mV s-1. For the Hb-HY modified electrode preparation, the HY and Hb concentrations are separately 1×10−2mol/L and 6.20×10−5mol/L
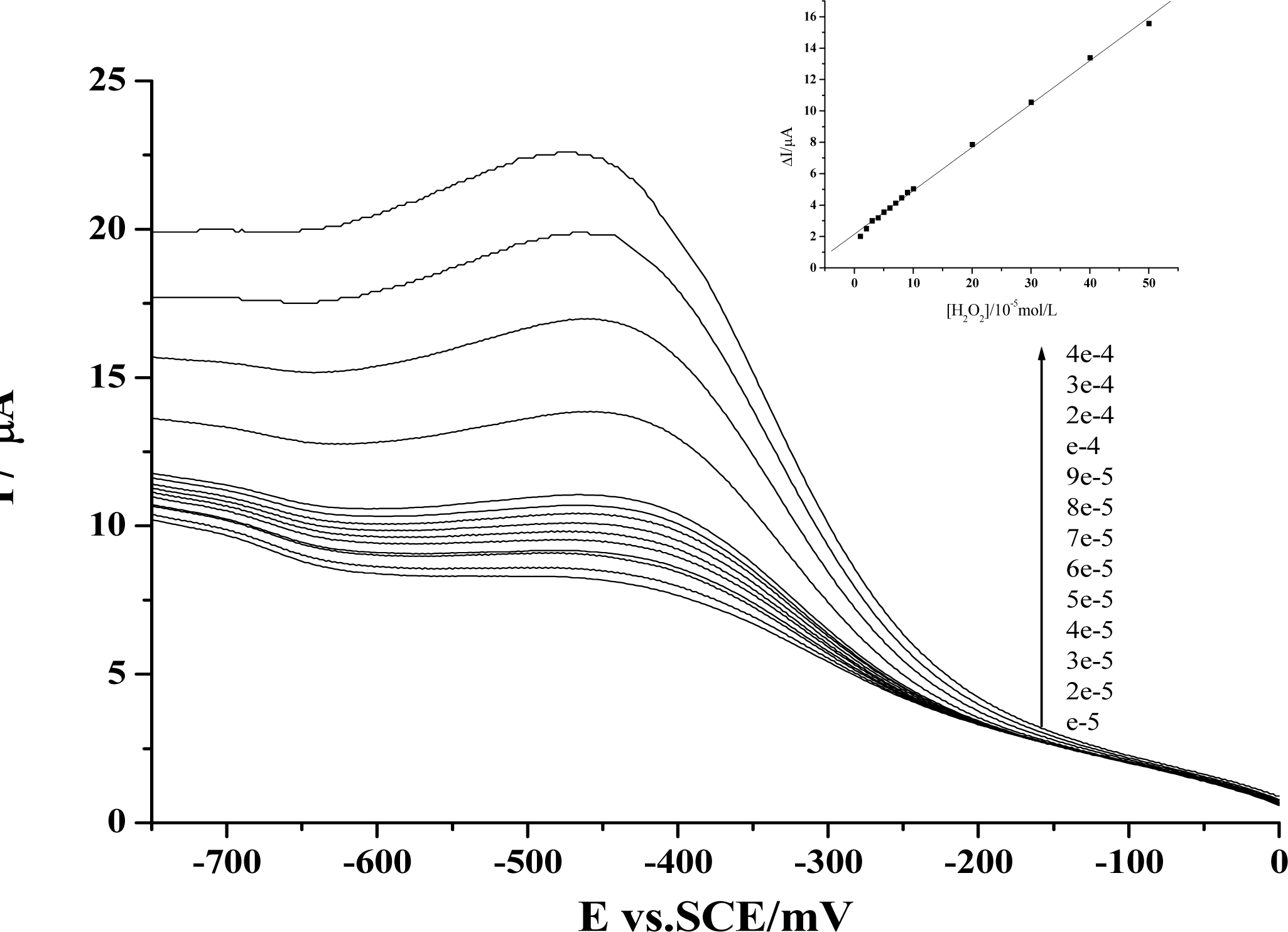
Figure 3.
Linear relationship between the increase of the reduction peak current and H2O2 concentration after visible light irradiation of 0h, 1h, 2h, 3h, respectively (The value of the reduction peak with 10−5 mol/L H2O2 is set as 0). Others same as in Figure 2.
Figure 3.
Linear relationship between the increase of the reduction peak current and H2O2 concentration after visible light irradiation of 0h, 1h, 2h, 3h, respectively (The value of the reduction peak with 10−5 mol/L H2O2 is set as 0). Others same as in Figure 2.
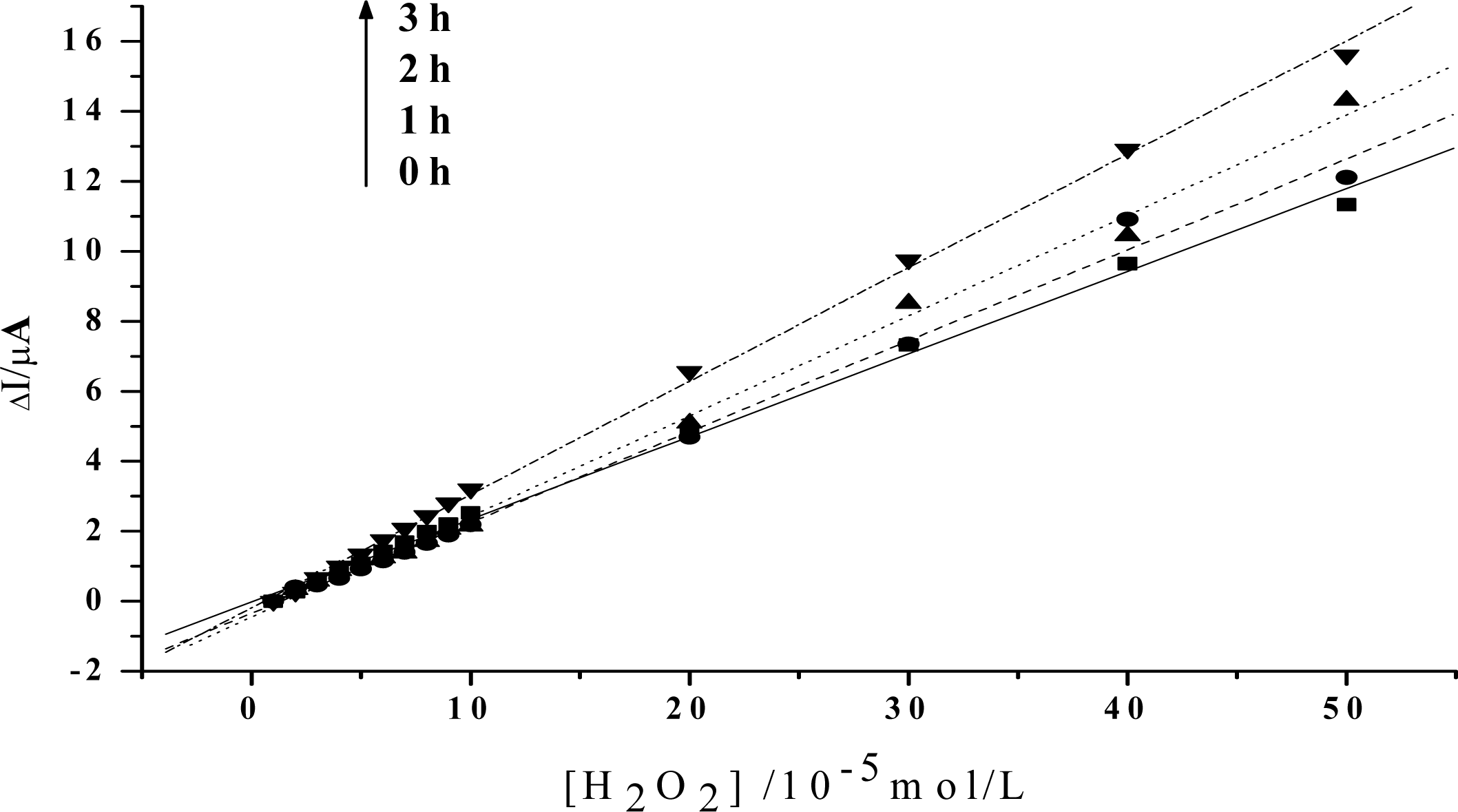
Figure 4.
Linear relationship between the increase of the reduction peak current and H2O2 concentration after visible light irradiation of 0h, 1h, 2h, 3h, respectively. HY concentration: 5×10−3mol/L. Others same as in Figure 3.
Figure 4.
Linear relationship between the increase of the reduction peak current and H2O2 concentration after visible light irradiation of 0h, 1h, 2h, 3h, respectively. HY concentration: 5×10−3mol/L. Others same as in Figure 3.
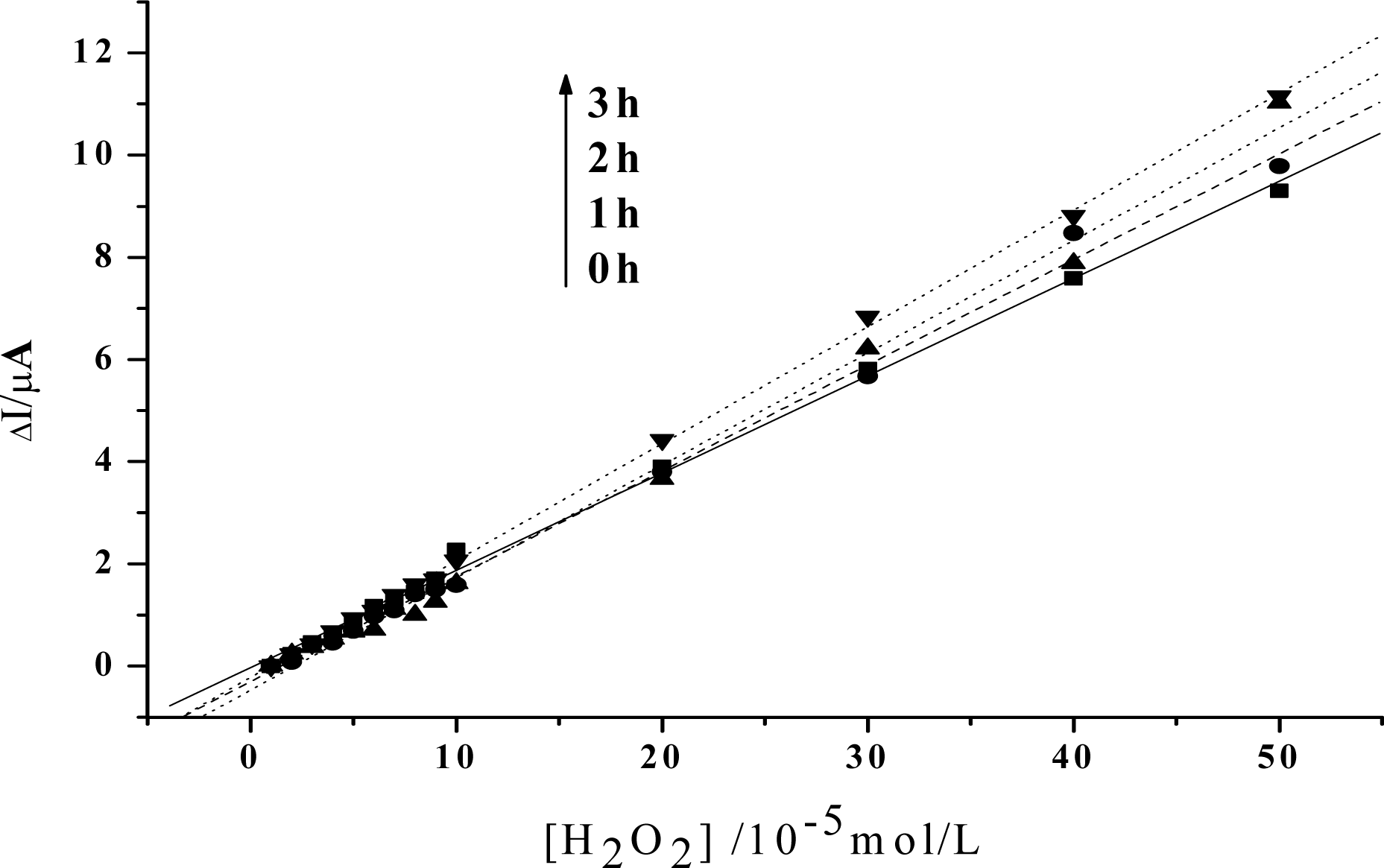
Figure 5.
Linear relationship between the increase of the reduction peak current and H2O2 concentration after visible light irradiation of 0h, 1h, 2h, 3h, respectively in an anaerobic condition. Others same as in Figure 3.
Figure 5.
Linear relationship between the increase of the reduction peak current and H2O2 concentration after visible light irradiation of 0h, 1h, 2h, 3h, respectively in an anaerobic condition. Others same as in Figure 3.

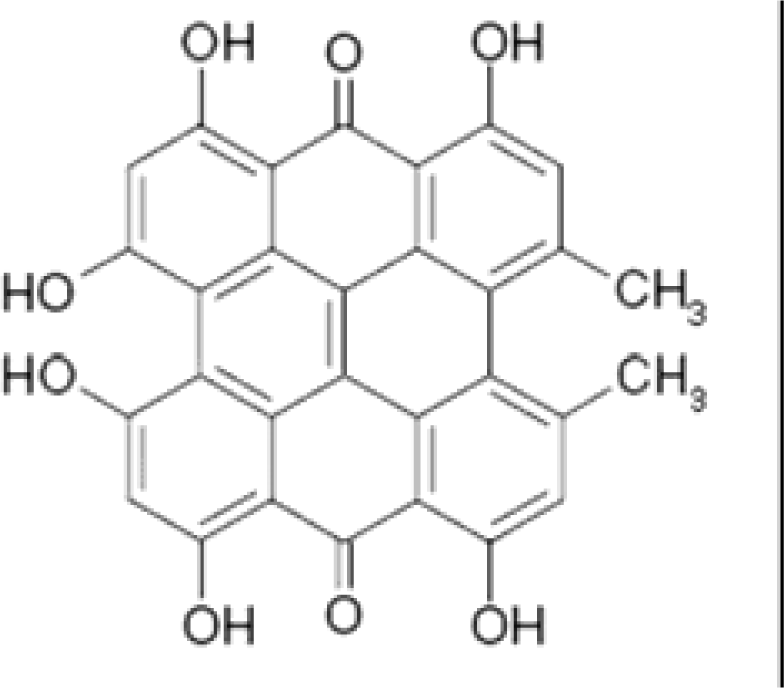
Scheme 1.
Chemical structure of hypericin.

Share and Cite
MDPI and ACS Style
Zhao, J.; Meng, W.; Miao, P.; Yu, Z.; Li, G. Photodynamic Effect of Hypericin on the Conformation and Catalytic Activity of Hemoglobin. Int. J. Mol. Sci. 2008, 9, 145-153. https://doi.org/10.3390/ijms9020145
AMA Style
Zhao J, Meng W, Miao P, Yu Z, Li G. Photodynamic Effect of Hypericin on the Conformation and Catalytic Activity of Hemoglobin. International Journal of Molecular Sciences. 2008; 9(2):145-153. https://doi.org/10.3390/ijms9020145
Chicago/Turabian StyleZhao, Jing, Wenying Meng, Peng Miao, Zhiguo Yu, and Genxi Li. 2008. "Photodynamic Effect of Hypericin on the Conformation and Catalytic Activity of Hemoglobin" International Journal of Molecular Sciences 9, no. 2: 145-153. https://doi.org/10.3390/ijms9020145