Antiproliferation of Hepatoma Cell and Progression of Cell Cycle as Affected by Isoflavone Extracts from Soybean Cake

Abstract
:1. Introduction
2. Results and Discussion
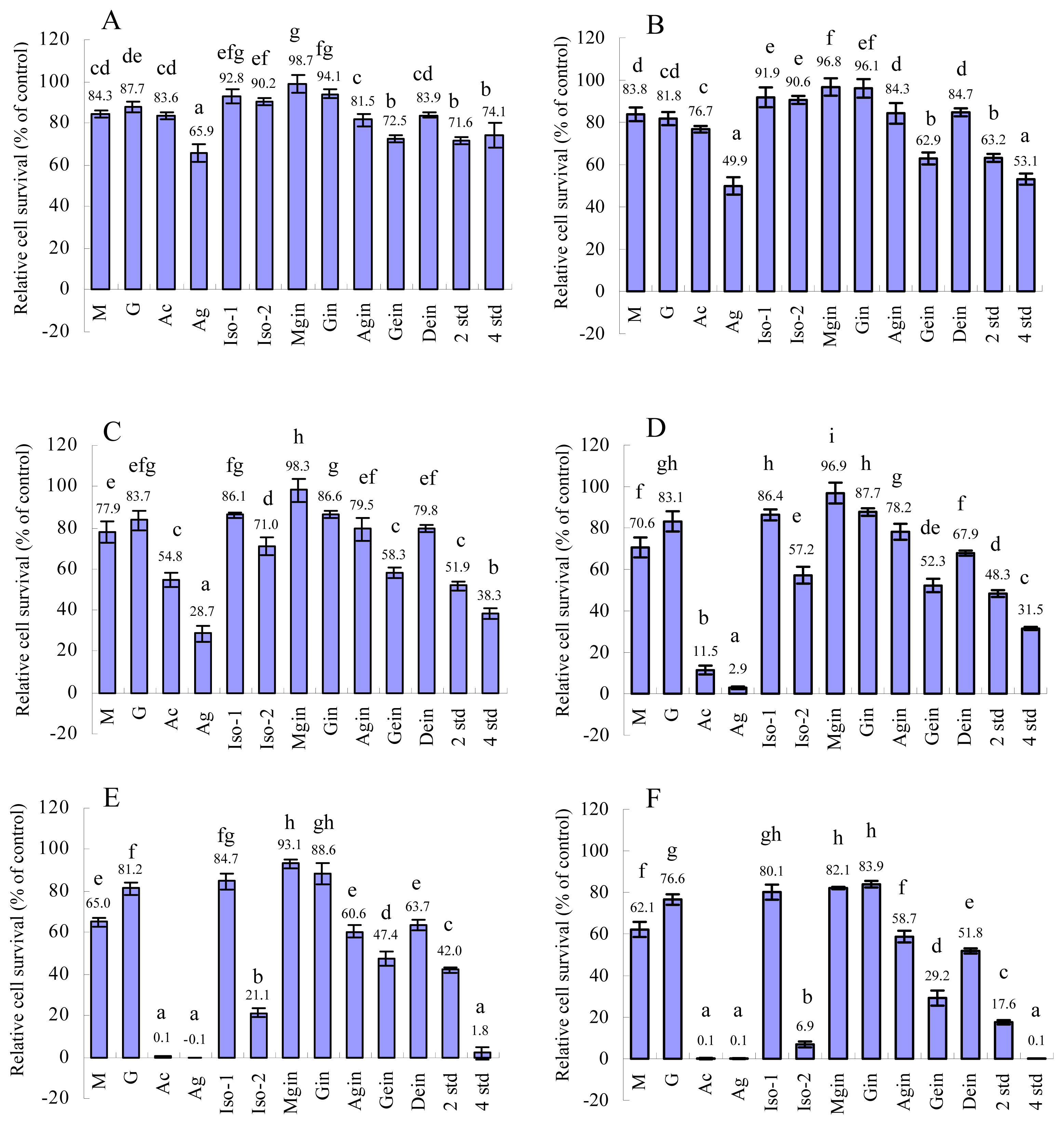
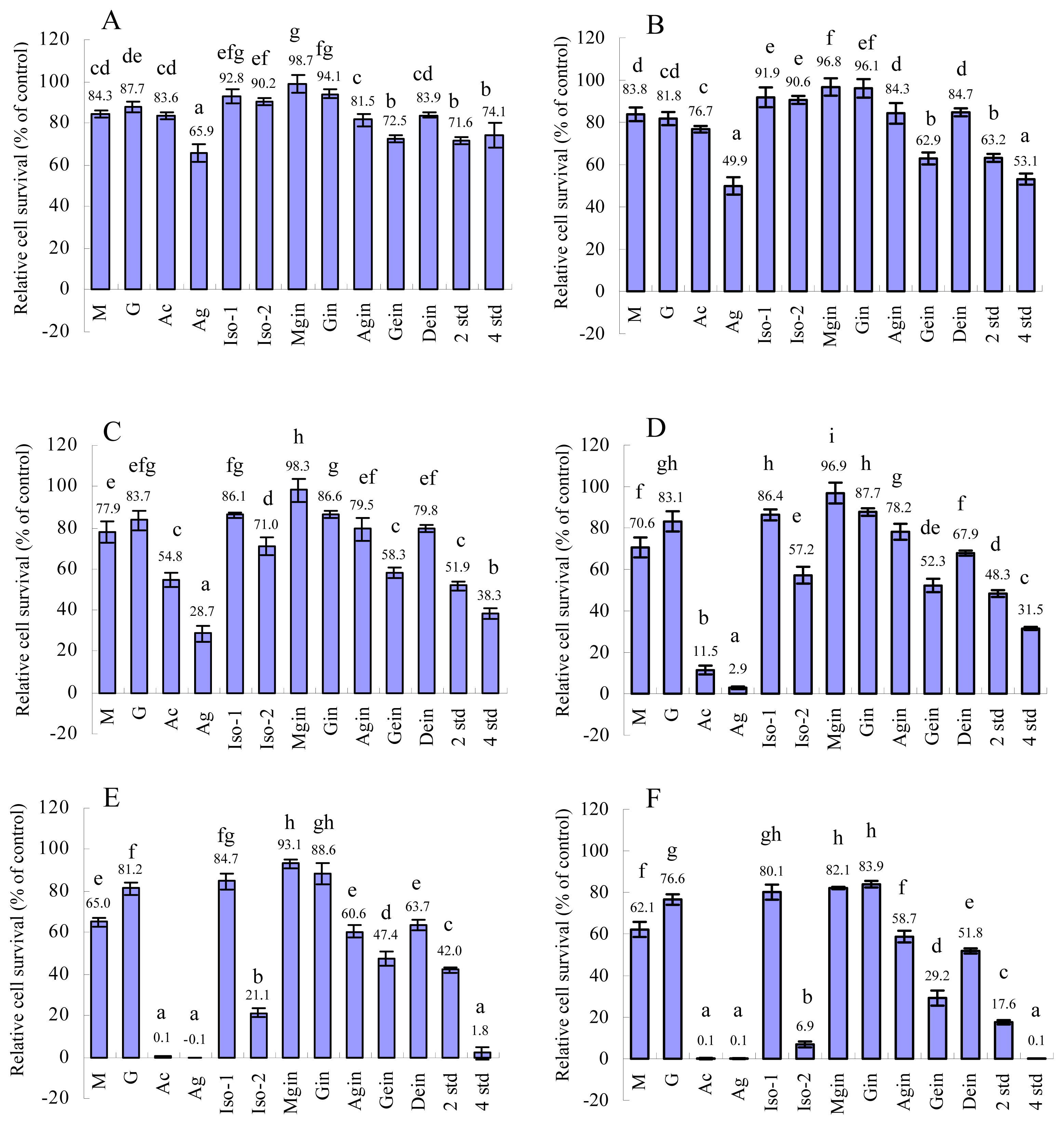
2.1 MTT test
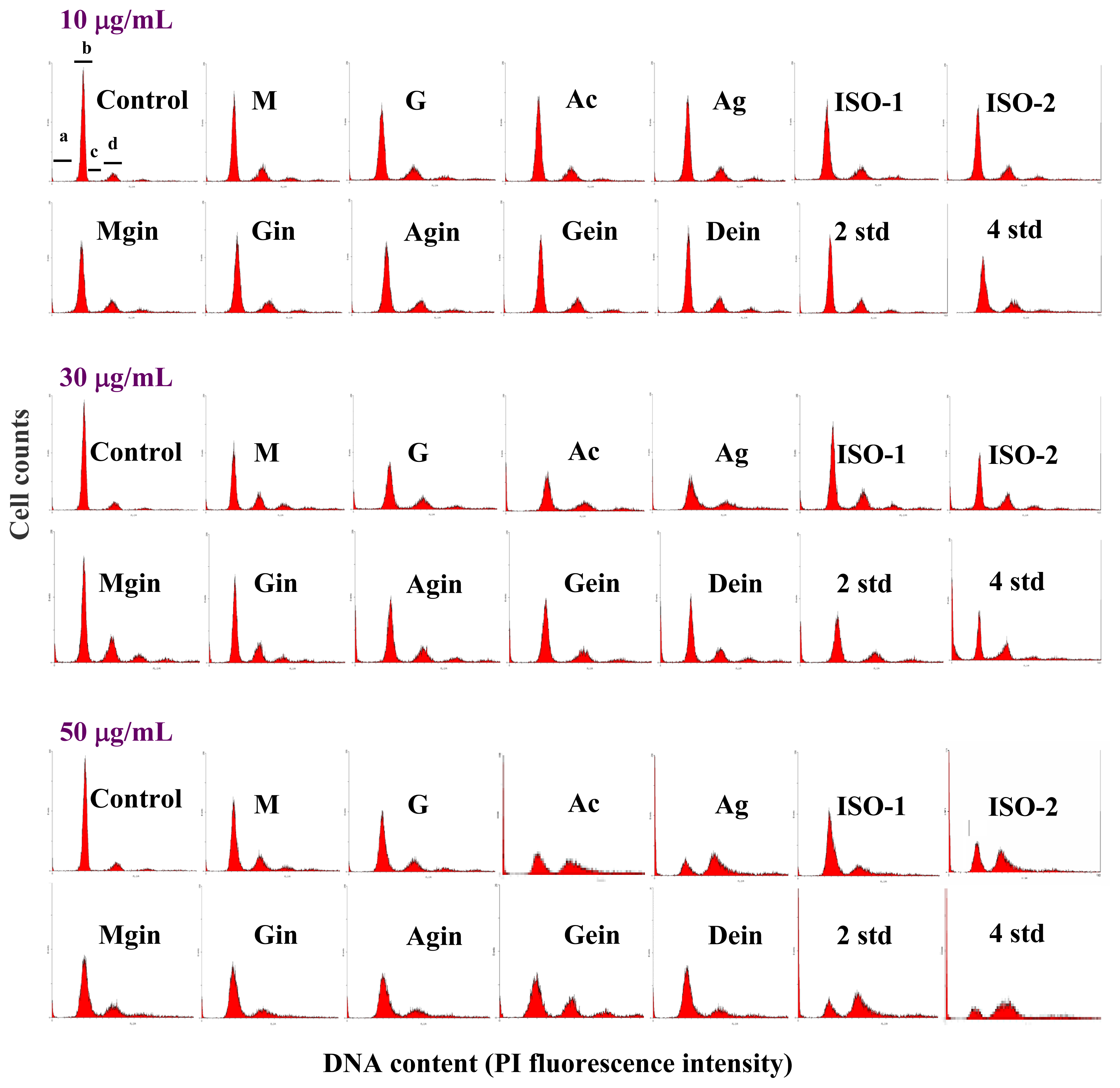
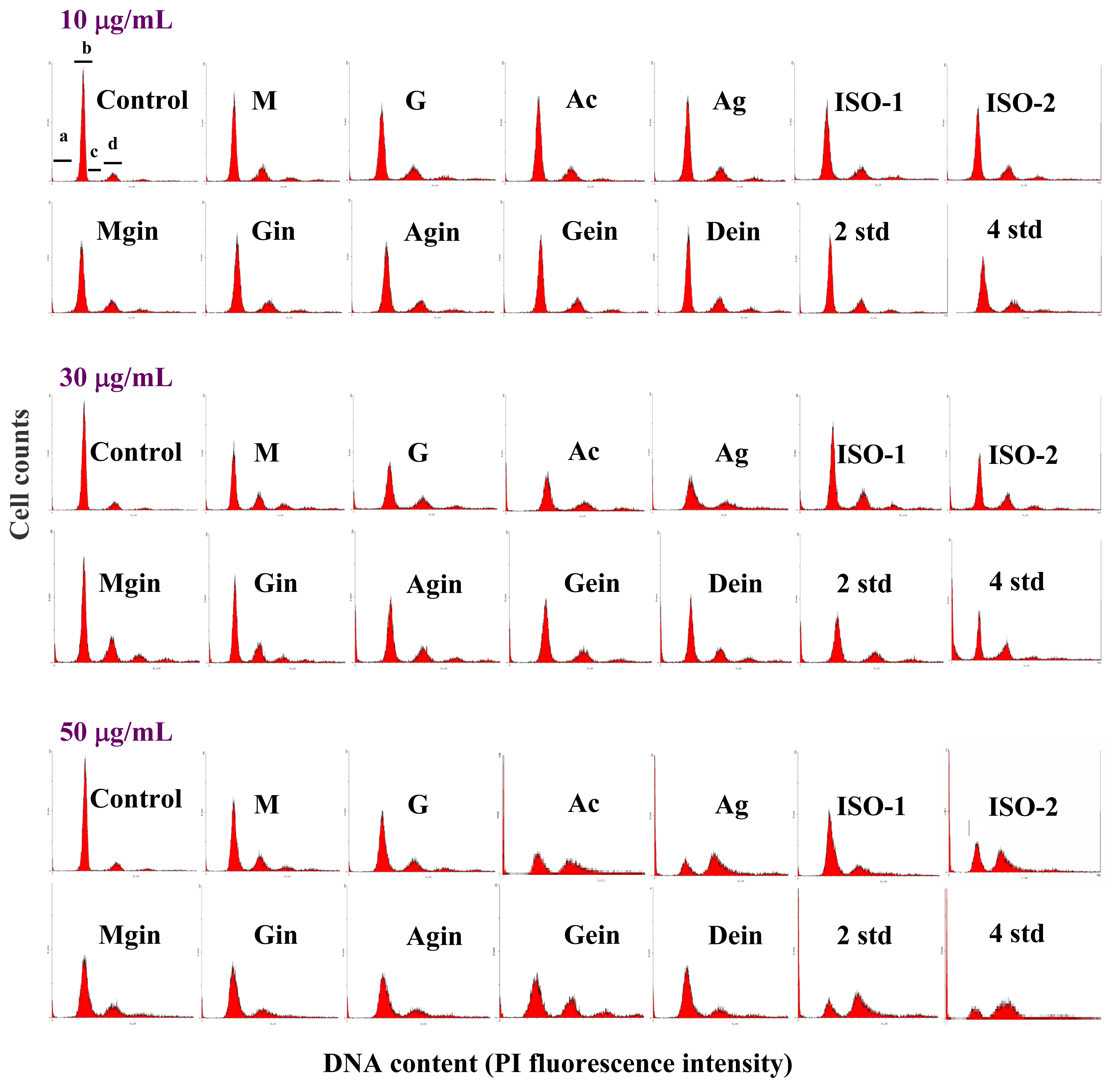
2.2 Cell cycle analysis
3. Experimental Section
3.1 Materials
3.2 Preparation of isoflavone extracts and fractions from soybean cake
3.3 Preparation of isoflavone extracts and fractions for cell antiproliferation study
3.4 Preparation of isoflavone standards for cell antiproliferation study
3.5 Preparation of final isoflavone concentration
3.6 Cell culture
3.7 MTT test
3.8 Cell cycle analysis
3.9 Statistical analysis

{kind=link}
{kind=link}
| Isoflavones | Isoflavone fractions | Isoflavone standard solutions | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| M | G | Ac | Ag | ISO-1 | ISO-2 | Mgin | Gin | Agin | Gein | Dein | 2 std | 4 std | |
| Malonyldaidzin | 48.8±0.3 | - | - | - | 22.9±0.0 | 12.0±0.0 | - | - | - | - | - | - | - |
| Malonylglycitin | 16.0±0.2 | - | - | - | 8.1±0.2 | 4.0±0.0 | - | - | - | - | - | - | - |
| Malonylgenistin | 35.1±1.2 | - | - | - | 17.0±0.1 | 8.8±0.1 | 100.0 | - | - | - | - | - | 100.0 |
| Daidzin | - | 25.3±0.1 | - | - | 9.1±0.1 | 6.3±0.1 | - | - | - | - | - | - | - |
| Glycitin | - | 29.2±0.2 | - | - | 9.0±0.1 | 7.3±0.1 | - | - | - | - | - | - | - |
| Genistin | - | 45.9±1.6 | - | - | 26.3±0.1 | 11.5±0.2 | - | 100.0 | - | - | - | - | 100.0 |
| Acetyldaidzin | - | - | 19.7±0.9 | - | 1.3±0.0 | 4.9±0.1 | - | - | - | - | - | - | - |
| Acetylglycitin | - | - | 12.8±0.7 | - | 1.0±0.0 | 3.2±0.2 | - | - | - | - | - | - | - |
| Acetylgenistin | - | - | 69.0±1.6 | - | 2.5±0.0 | 17.3±0.3 | - | - | 100.0 | - | - | 100.0 | |
| Daidzein | - | - | - | 47.3±0.5 | 1.4±0.1 | 11.9±0.1 | - | - | - | - | 100.0 | 100.0 | - |
| Glycitein | - | - | - | 16.3±0.2 | 0.6±0.0 | 4.1±0.0 | - | - | - | - | - | - | - |
| Genistein | - | - | - | 36.4±0.1 | 1.0±0.0 | 9.2±0.1 | - | - | - | 100.0 | - | 100.0 | 100.0 |
| Total | 99.9±1.7 | 100.4±0.8 | 101.5±2.1 | 100.0±0.6 | 100.2±0.0 | 100.5±0.2 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 200.0 | 400.0 |
| Isoflavone | M | G | Ac | Ag | Iso-1 | Iso-2 | Mgin | Gin | Agin | Gein | Dein | 2 std | 4 std |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| IC50(μg/mL) | 69.4 | 186.8 | 19.9 | 8.8 | 163.7 | 28.7 | 171.3 | 196.8 | 65.3 | 29.9 | 53.4 | 23.8 | 15.1 |
| Isoflavone | sub-G0/G1 (%) | G0/G1 (%) | S (%) | G2/M (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 10 μg/mL | 30 μg/mL | 50 μg/mL | 10 μg/mL | 30 μg/mL | 50 μg/mL | 10 μg/mL | 30 μg/mL | 50 μg/mL | 10 μg/mL | 30 μg/mL | 50 μg/mL | |
| Control | 2.2±0.1eA | 2.3±0.3hA | 2.2±0.3eA | 80.4±1.7aA | 80.7±2.0aA | 80.5±1.3aA | 6.6±0.6aA | 6.8±0.4aA | 6.8±0.9aA | 10.9±0.6cA | 10.2±1.3gA | 10.5±2.3fA |
| M | 3.1±0.1dB | 4.9±0.2gA | 5.7±0.5dA | 73.0±1.6bA | 67.1±3.4bAB | 63.8±2.6bC | 6.1±0.3aA | 5.6±0.6bA | 5.1±0.2bAB | 17.8±1.8abB | 22.4±2.6defAB | 25.4±2.5eA |
| G | 3.3±0.3cdB | 6.1±0.2deA | 6.0±0.1cdA | 74.5±0.8bA | 67.5±0.9bB | 65.0±1.6bB | 5.4±0.1bA | 5.3±0.7bAB | 4.9±0.1bB | 16.8±1.2abB | 21.1±1.4efA | 24.1±1.4eA |
| Ac | 3.2±0.4cdC | 7.4±0.8cB | 10.1±1.2aA | 73.1±1.6bA | 61.0±3.6cdB | 39.5±1.2dC | 6.7±0.7aA | 5.1±1.0bAB | 5.0±0.3bB | 17.0±1.5abC | 26.5±1.8bcdB | 45.5±3.1cA |
| Ag | 2.7±0.4deC | 7.2±0.3cdB | 9.8±1.2aA | 71.4±0.0bcA | 58.4±2.0dB | 27.9±4.0eC | 6.1±1.2abAB | 6.1±0.3aA | 5.0±0.4bB | 19.8±0.8abC | 28.3±2.0bcB | 57.3±4.0bA |
| ISO-1 | 3.4±0.2cdC | 5.0±0.1fgB | 6.3±0.7cdA | 72.7±2.9bcA | 69.0±2.8bA | 61.7±1.4bB | 6.0±0.7abA | 4.6±0.9bAB | 4.6±0.5bB | 18.0±2.6abB | 21.4±1.9efAB | 27.4±1.5eA |
| ISO-2 | 5.3±0.6aC | 9.1±0.2bB | 10.0±0.6aA | 70.9±1.2bcA | 61.1±2.3cdB | 33.5±3.0eC | 6.1±0.4abA | 4.8±0.6bB | 4.5±1.0bB | 17.6±1.7abB | 25.0±2.7cdeB | 52.0±3.3bA |
| Mgin | 4.3±0.4bC | 5.5±0.5efgB | 6.8±0.1bcdA | 73.2±1.7bA | 67.5±1.6bB | 62.5±1.6bC | 6.3±1.1abA | 6.3±0.5aA | 5.0±0.5abA | 16.2±1.7bC | 20.7±0.6efB | 25.7±0.6eA |
| Gin | 4.5±0.4abC | 5.3±0.2efgB | 6.6±0.4cdA | 71.9±1.8bcA | 69.2±3.6bB | 63.9±0.1bC | 6.5±1.7abA | 6.1±1.2aA | 5.1±0.2bA | 17.1±1.2abB | 19.5±2.2fAB | 24.5±2.2eA |
| Agin | 4.4±0.5bC | 6.3±0.5cdeB | 8.2±0.6bA | 73.4±0.3bA | 64.4±1.2bcB | 60.4±1.1bC | 5.7±0.7abA | 5.4±0.7bA | 4.4±0.8bA | 16.5±1.6abB | 24.0±1.4defA | 27.0±2.4eA |
| Gein | 4.5±0.3abC | 6.5±0.7cdeB | 10.6±0.9aA | 71.6±1.4bcA | 57.5±1.0dB | 47.9±1.4cC | 6.3±0.9abA | 6.7±0.8aA | 4.1±0.3bB | 17.6±0.8abC | 29.4±2.4bB | 37.4±3.2dA |
| Dein | 4.0±0.1bcC | 6.1±0.3defB | 7.4±0.8bcA | 73.1±2.7bcA | 69.5±0.2bB | 64.2±3.9bC | 6.2±0.9abA | 5.0±0.1bA | 4.9±0.1bB | 16.8±2.5abB | 19.5±0.2fAB | 23.5±2.2eA |
| 2 std | 4.7±0.3abC | 6.8±0.7cdB | 11.1±0.8aA | 71.9±0.2bA | 57.5±1.9dB | 32.3±4.2eC | 6.0±0.7abA | 5.2±0.6bA | 4.2±0.8bB | 17.4±0.3abC | 30.4±0.6bB | 52.4±2.8bA |
| 4 std | 4.6±0.5abC | 11.0±1.1aA | 11.1±0.2aA | 69.3±1.9cA | 48.8±2.1eB | 19.9±3.5fC | 6.1±0.3aA | 5.1±1.6bAB | 4.8±0.2bB | 20.0±1.1aC | 35.2±2.6aB | 64.2±3.6aA |
References and Notes
- World Cancer Research Fund, American Institute for Cancer Research, Food, nutrition and the prevention of cancer: a global perspective; American Institute for Cancer Research: Washington, 1997.
- Adlercreutz, H. Phyto-oestrogens and cancer. Lancet Oncol 2002, 3, 364–372. [Google Scholar]
- Block, G.; Patterson, B.; Subar, A. Fruits, vegetables, and cancer prevention: a review of the epidemiological evidence. Nutr. Cancer 1992, 18, 1–29. [Google Scholar]
- Messina, M.; Deshcheemaker, K.; Erdman, J.W. The role of soy in preventing and treating chronic disease. Am. J. Clin. Nutr 1998, 68, 1329–1544. [Google Scholar]
- Adlercreutz, H.; Honjo, H.; Higash, A. Urinary excretion of lignans and isoflavonoid phytoestrogens in Japanese men and women consuming traditional Japanese diet. Am. J. Clin. Nutr 1991, 54, 1093–1100. [Google Scholar]
- Dixon, R.A.; Ferreira, D. Genistein. Phytochemistry 2002, 60, 205–211. [Google Scholar]
- Peterson, G.; Barnes, S. Genistein inhibition of the growth of human breast cancer cells: Independence from estrogen receptors and the multi-drug resistance gene. Biochem. Biophys. Res. Commun 1991, 179, 661–667. [Google Scholar]
- Scholar, E.M.; Toews, M.L. Inhibition of invasion of murine mammary carcinoma cells by the tyrosine kinase inhibitor genistein. Cancer Lett 1994, 87, 159–162. [Google Scholar]
- Frey, R.S.; Li, J.; Singletary, K.W. Effects of genistein on cell proliferation and cell cycle arrest in nonneoplastic human mammary epithelial cells involvement of Cdc2, p21waf/cip1, p27kip1, and Cdc25C expression. Biochem. Pharmacol 2001, 61, 979–989. [Google Scholar]
- Kumi-Diaka, J. Chemosensitivity of human prostate cancer cells PC3 and LNCaP to genistein isoflavone and β-lapachone. Bio. Cell 2002, 94, 37–44. [Google Scholar]
- Chang, K.L.; Kung, M.L.; Chow, N.H.; Su, S.J. Genistein arrests hepatoma cells at G2/M phase: involvement of ATM activation and upregulationof p21waf1/cip1 and Weel. Biochem. Pharmacol 2004, 67, 717–726. [Google Scholar]
- Di Virgilio, A.L.; Iwami, K.; Wätjjen, W.; Kahl, R.; Gegen, G.H. Genotoxicity of the isoflavones genistein, daidzein and equol in V79 cells. Toxicol. Lett 2004, 151, 151–162. [Google Scholar]
- Oki, T.; Sowa, Y.; Hirose, T.; Takagaki, N.; Horinaka, M.; Nakanishi, R.; Yasuda, C.; Yoshida, T.; Kanazawa, M.; Satomi, Y.; Nishino, H.; Miki, T.; Sakai, T. Genistein induces Gadd45 gene and G2/M cell cycle arrest in the DU145 human prostate cancer cell line. FEBS Lett 2004, 577, 55–59. [Google Scholar]
- Kao, T.H.; Chen, B.H. Functional components in soybean cake and their effects on antioxidant activity. J. Agric. Food Chem 2006, 54, 7544–7555. [Google Scholar]
- Hewitt, A.L.; Singletary, K.W. Soy extract inhibits mammary adenocarcinoma growth in a syngeneic mouse model. Cancer Lett 2003, 192, 133–143. [Google Scholar]
- Constantinou, A.I.; Mehta, R.G.; Vaughan, A. Inhibition of N-methyl-N-nitrosourea-induced mammary tumors in rats by the soybean isoflavones. Anticancer Res 1996, 16, 3293–3298. [Google Scholar]
- Sakamoto, K. Synergistic effects of thearubigin and genistein on human prostate tumor cell (PC- 3) growth via cell cycle arrest. Cancer Lett 2000, 151, 103–109. [Google Scholar]
- Wei, H.; Wei, L.; Frenkel, K.; Bowen, R.; Barnes, S. Inhibition of tumor promoter-induced hydrogen peroxide formation in vitro and in vivo by genistein. Nutr. Cancer 1993, 20, 1–12. [Google Scholar]
- Wei, H.; Bower, R.; Zhang, X.; Lebwohl, M. Isoflavone genistein inhibits the initiation and promotion of two-stage skin carcinogenesis in mice. Carcinogenesis 1998, 19, 1509–1514. [Google Scholar]
- van Rossen, M.E.; Sluiter, W.; Bonthuis, F.; Jeekel, H.; Marquet, R.L.; van Eijck, C.H. Scavenging of reactive oxygen species leads to diminished peritoneal tumor recurrence. Cancer Res 2000, 60, 5625–5629. [Google Scholar]
- Bostwick, D.G.; Alexander, E.E.; Singh, R.; Shan, A.; Qian, J.; Santella, R.M.; Oberley, L.W.; Yan, T.; Zhong, W.; Jiang, X.; Oberley, T.D. Antioxidant enzyme expression and reactive oxygen species damage in prostatic intraepithelial neoplasia and cancer. Cancer 2000, 89, 123–134. [Google Scholar]
- Dudek, E.J.; Shang, F.; Taylor, A. H2O2-mediated oxidative stress activates NF-kappaB in lens epithelial cells. Free Radic. Biol. Med 2001, 31, 651–658. [Google Scholar]
- Popiolkiewicz, J.; Polkowski, K.; Skierski, S.; Mazurek, A.P. In vitro toxicity evaluation in the development of new anticancer drugs-genistein glycosides. Cancer Lett 2005, 229, 67–75. [Google Scholar]
- Papazisis, K.T.; Zambouli, D.; Kimoundri, O.T.; Papadakis, E.S.; Vala, V.; Deromichalos, D.; Voyatzi, S.; Markala, D.; Destouni, E.; Boutis, L.; Kortsaris, A.H. Protein tyrosine kinase inhibitor, genistein, enhances apoptosis and cell cycle arrest in K562 cells treated with γ-irradiation. Cancer Lett 2000, 160, 107–113. [Google Scholar]
- Casagrande, F.; Darbon, J.M. p21CIP1 is dispensable for the G2 arrest caused by genistein in human melanoma cells. Exp. Cell Res 2000, 258, 101–108. [Google Scholar]
- Ying, C.; Hsu, J.T.; Hung, H.C.; Lin, D.H.; Chen, L.F.O.; Wang, L.K. Growth and cell cycle regulation by isoflavones in human breast carcinoma cells. Reprod. Nutr. Dev 2002, 42, 55–64. [Google Scholar]
- Ismail, I.A.; Kang, K.S.; Kim, J.W.; Sohn, Y.K. Genistein induces G2/M cell cycle arrest and apoptosis in rat neuroblastoma B35 cells; involvement of p21waf1/cip1, Bax and Bcl-2. Korea J. Pathol 2006, 40, 39–347. [Google Scholar]
- Katdare, M.; Osborne, M.; Telang, N.T. Soy isoflavone genistein modulates cell cycle progression and induces apoptosis in HER-2/neu oncogene expressing human breast epithelial cells. Int. J. Oncol 2002, 21, 809–815. [Google Scholar]
- Sarkar, F.H.; Li, Y. Mechanisms of cancer chemoprevention by soy isoflavone genistein. Cancer Metastasis Rev 2002, 21, 265–280. [Google Scholar]
- Li, Y.; Bhuiyan, M.; Alhasan, S.; Senderowicz, A.M.; Sarkar, F.H. Induction of apoptosis and inhibition of c-erbB-2 in breast cancer cells by flavopiridol. Clin. Cancer Res 2000, 6, 223–229. [Google Scholar]
- Su, S.J.; Yeh, T.M.; Chuang, W.J.; Ho, C.L.; Chang, K.L.; Cheng, H.L.; Liu, H.S.; Cheng, H.L.; Hsu, P.Y.; Chow, N.H. The novel targets for anti-angiogenesis of genistein on human cancer cells. Biochem. Pharmacol 2005, 69, 307–318. [Google Scholar]
- Conklin, C.M.J.; Bechberger, J.F.; MacFabe, D.; Guthrie, N.; Kurowska, E.M.; Naus, C.C. Genistein and quercetin increase connexin 43 and suppress growth of breast cancer cells. Carcinogenesis 2007, 28, 93–100. [Google Scholar]
- Kim, H.Y.; Yu, R.; Kim, J.S.; Kim, Y.K.; Sung, M.K. Antiproliferative crude soy saponin extract modulates the expression of IκBα, protein kinase C, and cyclooxygenase-2 in human colon cancer cells. Cancer Lett 2004, 210, 1–6. [Google Scholar]
- Ellington, A.A.; Berhow, M.; Singletary, K.W. Induction of macroautophagy in human colon cancer cells by soybean B-group triterpenoid saponins. Carcinogenesis 2005, 26, 159–167. [Google Scholar]
- Ellington, A.A.; Berhow, M.; Singletary, K.W. Inhibitionof Akt signaling and enhanced ERK1/2 activity are involved in induction of marcoautophagy by triterpenoid B-group soyasaponins in colon cancer cells. Carcinogenesis 2006, 27, 298–306. [Google Scholar]
- Dalais, F.S.D.; Meliala, A.; Wattanapenpaiboon, N.; Frydenberg, M.; Siter, D.A.I.S; Thomson, W.K.; Wahlqvist, M.L. Effects of a diet rich in phytoestrogens on prostate-specific antigen and sex hormones in men diagnosed with prostate cancer. Urology 2004, 64, 510–515. [Google Scholar]
- Stephens, F.O. Phytoestrogens and prostate cancer: possible preventive role. Med. J. Aust 1997, 167, 138–140. [Google Scholar]
- Galati, G.; O’Brien, P.J. Potential toxicity of flavonoids and other dietary phenolics: significance for their chemopreventive and anticancer properties. Free Radic. Biol. Med 2004, 37, 287–303. [Google Scholar]
- McClain, R.M.; Wolz, E.; Davidovich, A.; Bausch, J. Genetic toxicity studies with genistein. Food Chem. Toxicol 2006, 44, 42–55. [Google Scholar]
- McClain, R.M.; Wolz, E.; Davidovich, A.; Edwards, J.; Bausch, J. Reproductive safety studies with genistein in rats. Food Chem. Toxicol 2007, 45, 1319–1332. [Google Scholar]
- Kao, T.H.; Lu, Y.F.; Chen, B.H. Preparative chromatography of four groups of isoflavones from soybean cake. Eur. Food Res. Technol 2005, 221, 459–465. [Google Scholar]
- Hsieh, H.C.; Kao, T.H.; Chen, B.H. A fast HPLC method for analysis of isoflavones in soybean. J. Liquid Chrom. Rel. Technol 2004, 27, 315–324. [Google Scholar]
- SAS, SAS Procedures and SAS/Graph User’s Guide, version 6, Software Release 8.2 (TS2MO) of the SAS System for Microsoft Windows; SAS Institute, Inc: Cary, NC, 2004.
© 2007 by MDPI Reproduction is permitted for noncommercial purposes.
Share and Cite
Kao, T.-H.; Huang, R.-F.S.; Chen, B.-H. Antiproliferation of Hepatoma Cell and Progression of Cell Cycle as Affected by Isoflavone Extracts from Soybean Cake. Int. J. Mol. Sci. 2007, 8, 1095-1110. https://doi.org/10.3390/i8111092
Kao T-H, Huang R-FS, Chen B-H. Antiproliferation of Hepatoma Cell and Progression of Cell Cycle as Affected by Isoflavone Extracts from Soybean Cake. International Journal of Molecular Sciences. 2007; 8(11):1095-1110. https://doi.org/10.3390/i8111092
Chicago/Turabian StyleKao, Tsai-Hua, Rwei-Fen S. Huang, and Bing-Huei Chen. 2007. "Antiproliferation of Hepatoma Cell and Progression of Cell Cycle as Affected by Isoflavone Extracts from Soybean Cake" International Journal of Molecular Sciences 8, no. 11: 1095-1110. https://doi.org/10.3390/i8111092